
Age-Related Changes in Serum N-Glycome in Men and Women—Clusters Associated with Comorbidity
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Methods
2.1. Study Design and Setting
2.2. Ethical Issues
2.3. Assessment of Smoking
2.4. Assessment of Alcohol Drinking
2.5. Usual Physical Activity History
2.6. Definition of Metabolic Disorders
2.7. Definition of Comorbidities and Their Quantification
2.8. Routine Analytical Determinations
2.9. Determination of Inflammation Markers
2.10. Determination of Glycation Markers (Markers of Glycaemic Control)
2.11. Serum N-Glycan Analyses
2.12. Statistical Analyses
3. Results
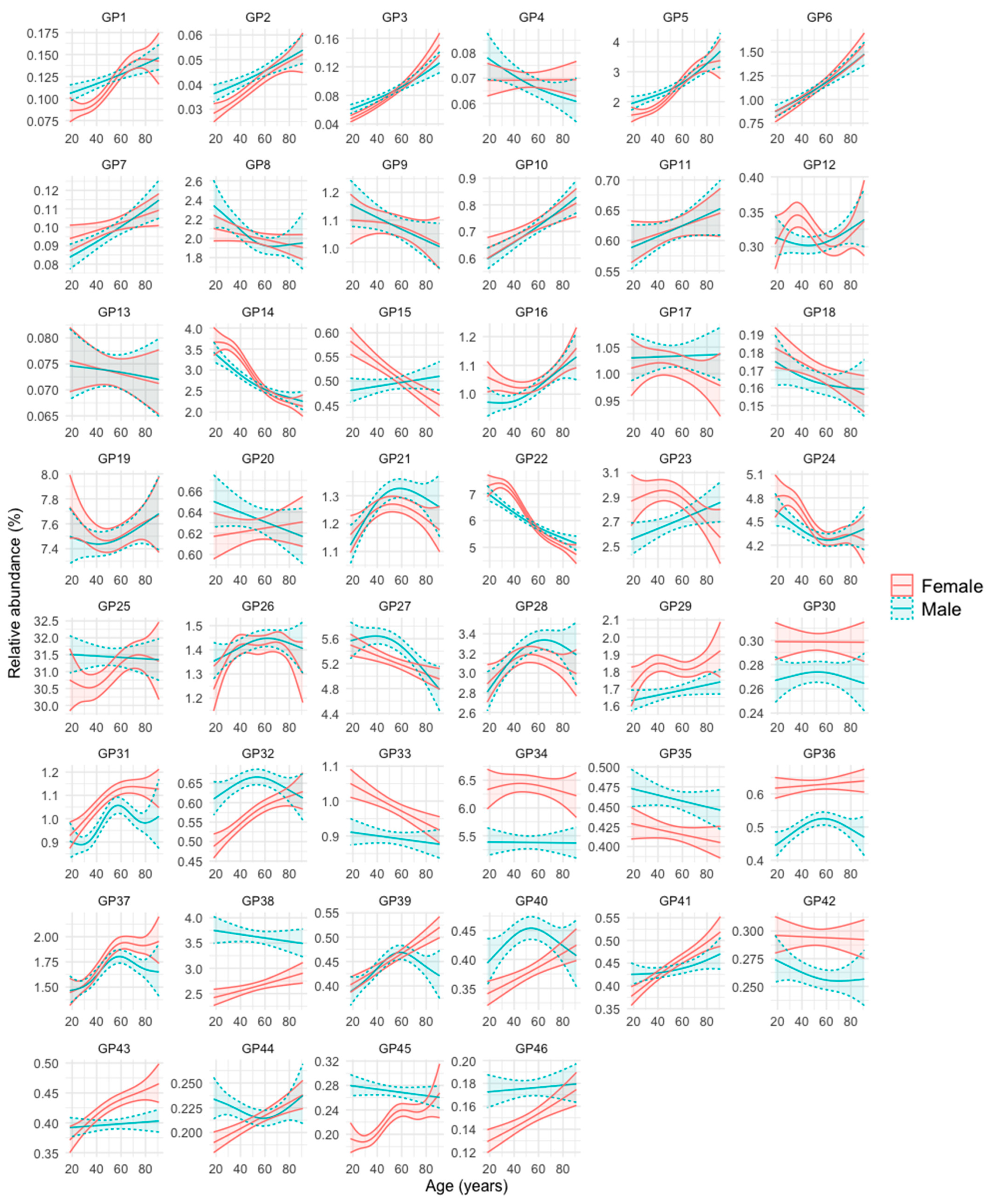
3.1. N-Glycan Peaks (GPs) in Relation to Age and Sex
- Group 1: GPs that are less abundant in women at younger ages, but increase in abundance over the years at a faster rate in this sex, so that at advanced ages their abundance becomes either equal or even higher than that of men. These are GP1, GP2, GP3, GP5, GP32, GP40, GP41, GP44, GP45, and GP46.
- Group 2: GPs that are more abundant in women at younger ages, but their abundance becomes equal or even higher in men in advances ages because they increase faster in men over the years (GP7, GP10, GP15, GP16, and GP23), or they decrease faster in women over the years (GP14, GP22, and GP24).
- Group 3: GPs that are similarly abundant in both sexes at young ages, but their abundance increases more prominently in women than in men over the years (GP39 and GP43).
- Group 4: GPs that are more abundant in one sex, regardless of age. They are GP29, GP30, GP31, GP33, GP34, GP36, and GP42 (more abundant in women), and GP35 and GP38 (more abundant in men).
- Group 5: GPs whose abundance increases or decreases over the years of age, but without clear differences between men and women. They are GP6, GP11, GP26, GP28, and GP37 (they increase over the years) and GP8, GP9, GP13, and GP18 (they decrease over the years).
- Group 6: GPs whose profile does not fit into any of the previous groups (GP4, GP12, GP17, GP19, GP20, GP21, GP25, and GP27).
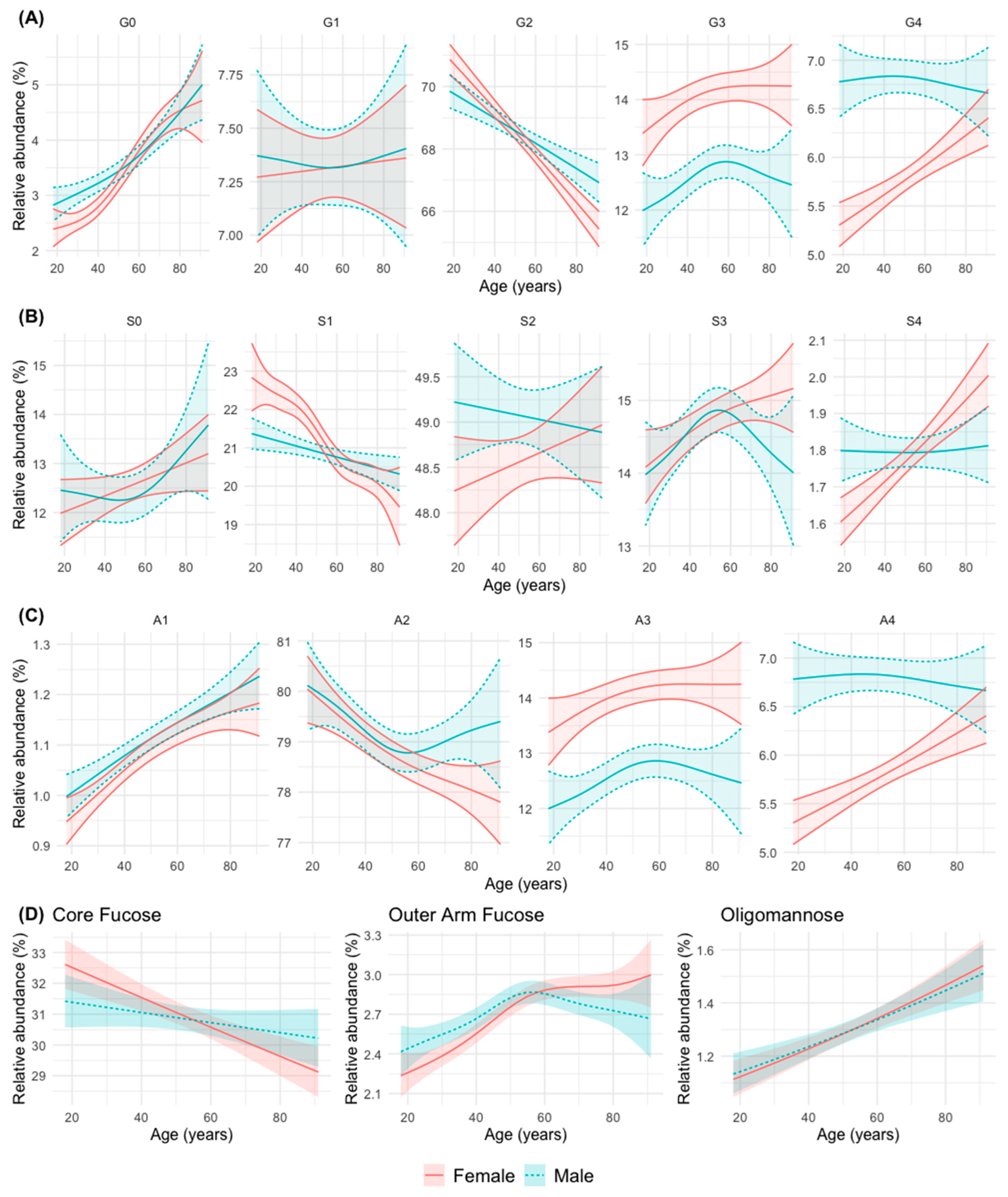
3.2. N-Glycan Groups (as Defined by Common Features) in Relation to Age and Sex
- Group 1: An increase in abundance with age in men and women is observed for simple N-glycans such as agalactosylated (G0), monoantennary (A1), non-sialylated (S0, although not statistically significant in this case), and oligomannose (OM) N-glycans. The abundance of N-glycans with peripheral (outer-arm, OF) fucosylation also increase significantly with age, although it stabilizes after 50 years of age in both sexes.
- Group 2: A decrease in abundance with age in men and women is observed for digalactosylated (G2), biantennary (A2, although it stabilizes in men from middle age onwards), monosialylated (S1, although more significantly in women) and core-fucosylated (CF) N-glycans.
- Group 3: A trend to increase in abundance with age is observed for with tetrasialylated (S4), tetragalactosylated (G4), and tetraantennary (A4) N-glycans in women, while in men the abundance remains stable at a higher level, which tends to equal that of women at advanced ages.
- Group 4: An increase in abundance with age in both sexes is observed for trigalactosylated (G3) and triantennary (A3, with a non-significant trend in this case) N-glycans, although always at a higher level of abundance in women.
- Group 5: A divergent change in abundance with age between men and women starting at middle age (possibly coinciding with menopause in women) is observed for trisialylated (S3, which decrease in men with a continuous increase in women), and biantennary (A2, that stabilize at that age in men, with a continuous decline in women) N-glycans.
- Group 6: A difference in the abundance between men and women up to middle age is observed for agalactosylated (G0) N-glycans (more abundant in men), and monosialylated (S1) N-glycans (more abundant in women) (Figure 2).
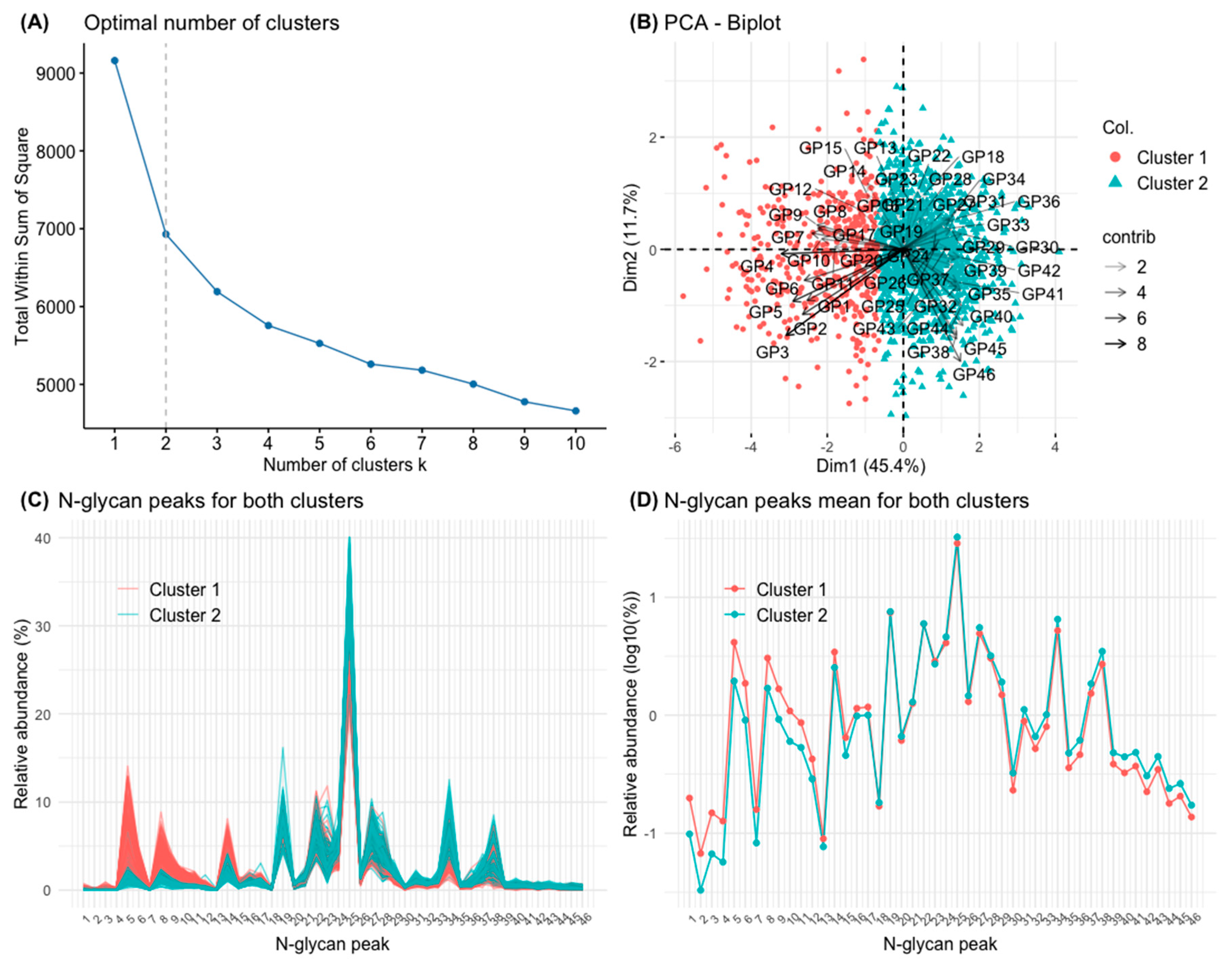
3.3. Clusters Defined by Principal Component Analysis (PCA) and k-Means Clustering
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scheper, A.F.; Schofield, J.; Bohara, R.; Ritter, T.; Pandit, A. Understanding glycosylation: Regulation through the metabolic flux of precursor pathways. Biotechnol. Adv. 2023, 67, 108184. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Lauc, G.; Pezer, M.; Rudan, I.; Campbell, H. Mechanisms of disease: The human N-glycome. Biochim. Biophys. Acta 2016, 1860, 1574–1582. [Google Scholar] [CrossRef] [PubMed]
- Cummings, R.D.; Pierce, J.M. The challenge and promise of glycomics. Chem. Biol. 2014, 21, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Axford, J.; Alavi, A.; Cummings, R.; Lauc, G.; Opdenakker, G.; Reis, C.; Rudd, P. Translational glycobiology: From bench to bedside. J. R. Soc. Med. 2019, 112, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Stöckmann, H.; O’Flaherty, R.; Adamczyk, B.; Saldova, R.; Rudd, P.M. Automated, high-throughput serum glycoprofiling platform. Integr. Biol. 2015, 7, 1026–1032. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, R.; Muniyappa, M.; Walsh, I.; Stöckmann, H.; Hilliard, M.; Hutson, R.; Saldova, R.; Rudd, P.M. A robust and versatile automated glycoanalytical technology for serum antibodies and acute phase proteins: Ovarian Cancer Case Study. Mol. Cell Proteom. 2019, 18, 2191–2206. [Google Scholar] [CrossRef]
- Kyselova, Z.; Mechref, Y.; Kang, P.; Goetz, J.A.; Dobrolecki, L.E.; Sledge, G.W.; Schnaper, L.; Hickey, R.J.; Malkas, L.H.; Novotny, M.V. Breast cancer diagnosis and prognosis through quantitative measurements of serum glycan profiles. Clin. Chem. 2008, 54, 1166–1175. [Google Scholar] [CrossRef]
- Saldova, R.; Fan, Y.; Fitzpatrick, J.M.; Watson, R.W.G.; Rudd, P.M. Core fucosylation and α2-3 sialylation in serum N-glycome is significantly increased in prostate cancer comparing to benign prostate hyperplasia. Glycobiology 2011, 21, 195–205. [Google Scholar] [CrossRef]
- Saldova, R.; Asadi Shehni, A.; Haakensen, V.D.; Steinfeld, I.; Hilliard, M.; Kifer, I.; Helland, A.; Yakhini, Z.; Børresen-Dale, A.L.; Rudd, P.M. Association of N-glycosylation with breast carcinoma and systemic features using high-resolution quantitative UPLC. J. Proteome Res. 2014, 13, 2314–2327. [Google Scholar] [CrossRef]
- Saldova, R.; Haakensen, V.D.; Rødland, E.; Walsh, I.; Stöckmann, H.; Engebraaten, O.; Børresen-Dale, A.L.; Rudd, P.M. Serum N-glycome alterations in breast cancer during multimodal treatment and follow-up. Mol. Oncol. 2017, 11, 1361–1379. [Google Scholar] [CrossRef] [PubMed]
- Huffman, J.E.; Pučić-Baković, M.; Klarić, L.; Hennig, R.; Selman, M.H.; Vučković, F.; Novokmet, M.; Krištić, J.; Borowiak, M.; Muth, T.; et al. Comparative performance of four methods for high-throughput glycosylation analysis of immunoglobulin G in genetic and epidemiological research. Mol. Cell Proteom. 2014, 13, 1598–1610. [Google Scholar] [CrossRef] [PubMed]
- Cindrić, A.; Krištić, J.; Martinić Kavur, M.; Pezer, M. Glycosylation and aging. Adv. Exp. Med. Biol. 2021, 1325, 341–373. [Google Scholar] [CrossRef] [PubMed]
- Vanhooren, V.; Desmyter, L.; Liu, X.E.; Cardelli, M.; Franceschi, C.; Federico, A.; Libert, C.; Laroy, W.; Dewaele, S.; Contreras, R.; et al. N-glycomic changes in serum proteins during human aging. Rejuvenation Res. 2007, 10, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Vanhooren, V.; Liu, X.E.; Franceschi, C.; Gao, C.F.; Libert, C.; Contreras, R.; Chen, C. N-glycan profiles as tools in diagnosis of hepatocellular carcinoma and prediction of healthy human ageing. Mech. Ageing Dev. 2009, 130, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Vanhooren, V.; Dewaele, S.; Libert, C.; Engelborghs, S.; De Deyn, P.P.; Toussaint, O.; Debacq-Chainiaux, F.; Poulain, M.; Glupczynski, Y.; Franceschi, C.; et al. Serum N-glycan profile shift during human ageing. Exp. Gerontol. 2010, 45, 738–743. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Nie, H.; Sun, X.; Sun, W.; Qu, Y.; Liu, X.; Yao, Y.; Liang, X.; Chen, C.C.; Li, Y. Human serum N-glycan profiles are age and sex dependent. Age Ageing 2011, 40, 568–575. [Google Scholar] [CrossRef]
- Borelli, V.; Vanhooren, V.; Lonardi, E.; Reiding, K.R.; Capri, M.; Libert, C.; Garagnani, P.; Salvioli, S.; Franceschi, C.; Wuhrer, M. Plasma N-glycome signature of Down syndrome. J. Proteome Res. 2015, 14, 4232–4245. [Google Scholar] [CrossRef]
- Catera, M.; Borelli, V.; Malagolini, N.; Chiricolo, M.; Venturi, G.; Reis, C.A.; Osorio, H.; Abruzzo, P.M.; Capri, M.; Monti, D.; et al. Identification of novel plasma glycosylation-associated markers of aging. Oncotarget 2016, 7, 7455–7468. [Google Scholar] [CrossRef]
- Knezević, A.; Polasek, O.; Gornik, O.; Rudan, I.; Campbell, H.; Hayward, C.; Wright, A.; Kolcic, I.; O’Donoghue, N.; Bones, J.; et al. Variability, heritability and environmental determinants of human plasma N-glycome. J. Proteome Res. 2009, 8, 694–701. [Google Scholar] [CrossRef]
- Knežević, A.; Gornik, O.; Polašek, O.; Pučić, M.; Redžić, I.; Novokmet, M.; Rudd, P.M.; Wright, A.F.; Campbell, H.; Rudan, I.; et al. Effects of aging, body mass index, plasma lipid profiles, and smoking on human plasma N-glycans. Glycobiology 2010, 20, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Pucic, M.; Muzinic, A.; Novokmet, M.; Skledar, M.; Pivac, N.; Lauc, G.; Gornik, O. Changes in plasma and IgG N-glycome during childhood and adolescence. Glycobiology 2012, 22, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Ruhaak, L.R.; Uh, H.W.; Beekman, M.; Hokke, C.H.; Westendorp, R.G.; Houwing-Duistermaat, J.; Wuhrer, M.; Deelder, A.M.; Slagboom, P.E. Plasma protein N-glycan profiles are associated with calendar age, familial longevity and health. J. Proteome Res. 2011, 10, 1667–1674. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Hashii, N.; Tsumoto, H.; Takakura, D.; Ohta, Y.; Abe, Y.; Arai, Y.; Kawasaki, N.; Hirose, N.; Endo, T. SONIC (Septuagenarians, Octogenarians, Nonagenarians Investigation with Centenarians). Change in N-glycosylation of plasma proteins in Japanese semisupercentenarians. PLoS ONE 2015, 10, e0142645. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, M.M.; Singh, P.; Rizvi, S.I. Erythrocyte sialic acid content during aging in humans: Correlation with markers of oxidative stress. Dis. Markers 2012, 32, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Merleev, A.A.; Park, D.; Xie, Y.; Kailemia, M.J.; Xu, G.; Ruhaak, L.R.; Kim, K.; Hong, Q.; Li, Q.; Patel, F.; et al. A site-specific map of the human plasma glycome and its age and gender-associated alterations. Sci. Rep. 2020, 10, 17505. [Google Scholar] [CrossRef]
- Ząbczyńska, M.; Link-Lenczowski, P.; Pocheć, E. Glycosylation in autoimmune diseases. Adv. Exp. Med. Biol. 2021, 1325, 205–218. [Google Scholar] [CrossRef]
- Ruhaak, L.R.; Uh, H.W.; Beekman, M.; Koeleman, C.A.; Hokke, C.H.; Westendorp, R.G.; Wuhrer, M.; Houwing-Duistermaat, J.J.; Slagboom, P.E.; Deelder, A.M. Decreased levels of bisecting GlcNAc glycoforms of IgG are associated with human longevity. PLoS ONE 2010, 5, e12566. [Google Scholar] [CrossRef]
- Krištić, J.; Vučković, F.; Menni, C.; Klarić, L.; Keser, T.; Beceheli, I.; Pučić-Baković, M.; Novokmet, M.; Mangino, M.; Thaqi, K.; et al. Glycans are a novel biomarker of chronological and biological ages. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69, 779–789. [Google Scholar] [CrossRef]
- Krištić, J.; Lauc, G.; Pezer, M. Immunoglobulin G glycans—Biomarkers and molecular effectors of aging. Clin. Chim. Acta. 2022, 535, 30–45. [Google Scholar] [CrossRef]
- Shkunnikova, S.; Mijakovac, A.; Sironic, L.; Hanic, M.; Lauc, G.; Kavur, M.M. IgG glycans in health and disease: Prediction, intervention, prognosis, and therapy. Biotechnol. Adv. 2023, 67, 108169. [Google Scholar] [CrossRef] [PubMed]
- Mijakovac, A.; Frkatović, A.; Hanić, M.; Ivok, J.; Martinić Kavur, M.; Pučić-Baković, M.; Spector, T.; Zoldoš, V.; Mangino, M.; Lauc, G. Heritability of the glycan clock of biological age. Front. Cell Dev. Biol. 2022, 10, 982609. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflammaging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C. Inflammaging as a major characteristic of old people: Can it be prevented or cured? Nutr. Rev. 2007, 65, S173–S176. [Google Scholar] [CrossRef] [PubMed]
- Dall’Olio, F.; Vanhooren, V.; Chen, C.C.; Slagboom, P.E.; Wuhrer, M.; Franceschi, C. N-glycomic biomarkers of biological aging and longevity: A link with inflammaging. Ageing Res. Rev. 2013, 12, 685–698. [Google Scholar] [CrossRef]
- Dall’Olio, F. Glycobiology of aging. Subcell. Biochem. 2018, 90, 505–526. [Google Scholar] [CrossRef]
- Alende-Castro, V.; Alonso-Sampedro, M.; Vazquez-Temprano, N.; Tuñez, C.; Rey, D.; García-Iglesias, C.; Sopeña, B.; Gude, F.; Gonzalez-Quintela, A. Factors influencing erythrocyte sedimentation rate in adults: New evidence for an old test. Medicine 2019, 98, e16816. [Google Scholar] [CrossRef]
- O’Flaherty, R.; Simon, Á.; Alonso-Sampedro, M.; Sánchez-Batán, S.; Fernández-Merino, C.; Gude, F.; Saldova, R.; González-Quintela, A. Changes in serum N-glycome for risk drinkers: A comparison with standard markers for alcohol abuse in men and women. Biomolecules 2022, 12, 241. [Google Scholar] [CrossRef]
- Gual, A.; Martos, A.R.; Lligoña, A.; Llopis, J.J. Does the concept of a standard drink apply to viticultural societies? Alcohol Alcohol. 1999, 34, 153–160. [Google Scholar] [CrossRef]
- Craig, C.L.; Marshall, A.L.; Sjöström, M.; Bauman, A.E.; Booth, M.L.; Ainsworth, B.E.; Pratt, M.; Ekelund, U.; Yngve, A.; Sallis, J.F.; et al. International physical activity questionnaire: 12-country reliability and validity. Med. Sci. Sports Exerc. 2003, 35, 1381–1395. [Google Scholar] [CrossRef]
- Román Viñas, B.; Ribas Barba, L.; Ngo, J.; Serra Majem, L. Validity of the international physical activity questionnaire in the Catalan population (Spain). Gac. Sanit. 2013, 27, 254–257. [Google Scholar] [CrossRef] [PubMed]
- Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults. Executive Summary of The Third Report of The National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, And Treatment of High Blood Cholesterol In Adults (Adult Treatment Panel III). JAMA 2001, 285, 2486–2497. [Google Scholar] [CrossRef] [PubMed]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2019. Diabetes Care 2019, 42 (Suppl. S1), S13–S28. [Google Scholar] [CrossRef] [PubMed]
- Charlson, M.E.; Pompei, P.; Ales, K.L.; MacKenzie, C.R. A new method of classifying prognostic comorbidity in longitudinal studies: Development and validation. J. Chronic Dis. 1987, 40, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Alende-Castro, V.; Alonso-Sampedro, M.; Fernández-Merino, C.; Sánchez-Castro, J.; Sopeña, B.; Gude, F.; Gonzalez-Quintela, A. C-Reactive protein versus erythrocyte sedimentation rate: Implications among patients with no known inflammatory conditions. J. Am. Board Fam. Med. 2021, 34, 974–983. [Google Scholar] [CrossRef] [PubMed]
- Alende-Castro, V.; Alonso-Sampedro, M.; Gude, F.; Gonzalez-Quintela, A. Serum concentrations of interleukin 6 in the general adult population: Possible implications for anti-IL-6 therapy in SARS-Cov-2 infection and IL-6-related diseases. J. Investig. Allergol. Clin. Immunol. 2021, 31, 75–78. [Google Scholar] [CrossRef]
- Hoelzel, W.; Weykamp, C.; Jeppsson, J.O.; Miedema, K.; Barr, J.R.; Goodall, I.; Hoshino, T.; John, W.G.; Kobold, U.; Little, R.; et al. IFCC reference system for measurement of hemoglobin A1c in human blood and the national standardization schemes in the United States, Japan, and Sweden: A method-comparison study. Clin. Chem. 2004, 50, 166–174. [Google Scholar] [CrossRef]
- Wu, W.C.; Ma, W.Y.; Wei, J.N.; Yu, T.Y.; Lin, M.S.; Shih, S.R.; Hua, C.H.; Liao, Y.J.; Chuang, L.M.; Li, H.Y. Serum glycated albumin to guide the diagnosis of diabetes mellitus. PLoS ONE. 2016, 11, e0146780. [Google Scholar] [CrossRef]
- Royle, L.; Campbell, M.P.; Radcliffe, C.M.; White, D.M.; Harvey, D.J.; Abrahams, J.L.; Kim, Y.G.; Henry, G.W.; Shadick, N.A.; Weinblatt, M.E.; et al. HPLC-based analysis of serum N-glycans on a 96-well plate platform with dedicated database software. Anal. Biochem. 2008, 376, 1–12. [Google Scholar] [CrossRef]
- Cheng, K.; Zhou, Y.; Neelamegham, S. DrawGlycan-SNFG: A robust tool to render glycans and glycopeptides with fragmentation information. Glycobiology 2016, 27, 200–205. [Google Scholar] [CrossRef]
- Neelamegham, S.; Aoki-Kinoshita, K.; Bolton, E.; Frank, M.; Lisacek, F.; Lütteke, T.; O’Boyle, N.; Packer, N.; Stanley, P.; Toukach, P.; et al. Updates to the Symbol Nomenclature for Glycans guidelines. Glycobiology 2019, 29, 620–624. [Google Scholar] [CrossRef]
- Zhao, S.; Walsh, I.; Abrahams, J.; Royle, L.; Nguyen-Khuong, T.; Spencer, D.; Fernandes, D.L.; Packer, N.; Rudd, P.M.; Campbell, M.P. GlycoStore: A database of retention properties for glycan analysis. Bioinform. 2018, 34, 3231–3232. [Google Scholar] [CrossRef]
- Kynclova, P.; Hron, K.; Filzmoser, P. Correlation between compositional parts based on symmetric balances. Math. Geosci. 2017, 49, 777–796. [Google Scholar] [CrossRef]
- Cao, Y.; Lin, W.; Li, H. Two-sample tests of high-dimensional means for compositional data. Biometrika 2018, 105, 115–132. [Google Scholar] [CrossRef]
- Filzmoser, P.; Hron, K.; Reimann, C. Principal component analysis for compositional data with outliers. Environmetrics 2009, 20, 621–632. [Google Scholar] [CrossRef]
- Filzmoser, P.; Hron, K.; Reimann, C. Interpretation of multivariate outliers for compositional data. Comput. Geosci. 2012, 39, 77–85. [Google Scholar] [CrossRef]
- Charrad, M.; Ghazzali, N.; Boiteau, V.; Niknafs, A. NbClust: An R package for determining the relevant number of clusters in a data set. J. Stat. Soft. 2014, 61, 1–36. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Tsagris, M.; Athineou, G.; Alenazi, A.; Adam, C. Compositional: Compositional Data Analysis. R Package Version 5.4. 2022. Available online: https://CRAN.R-project.org/package=Compositional (accessed on 15 December 2023).
- Van den Boogaart, K.G.; Tolosana-Delgado, R.; Bren, M. Compositions: Compositional Data Analysis. R Package Version 2.0-4. 2022. Available online: https://CRAN.R-project.org/package=compositions (accessed on 15 December 2023).
- Filzmoser, P.; Hron, K.; Templ, M. Applied Compositional Data Analysis; Springer: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Filzmoser, P.; Gschwandtner, M. Mvoutlier: Multivariate Outlier Detection Based on Robust Methods. R Package Version 2.1.1. 2021. Available online: https://cran.r-project.org/web/packages/mvoutlier/mvoutlier.pdf (accessed on 15 December 2023).
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2017. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016. [Google Scholar]
- Ercan, A.; Kohrt, W.M.; Cui, J.; Deane, K.D.; Pezer, M.; Yu, E.W.; Hausmann, J.S.; Campbell, H.; Kaiser, U.B.; Rudd, P.M.; et al. Estrogens regulate glycosylation of IgG in women and men. JCI Insight 2017, 2, e89703. [Google Scholar] [CrossRef]
- Baechle, J.J.; Chen, N.; Makhijani, P.; Winer, S.; Furman, D.; Winer, D.A. Chronic inflammation and the hallmarks of aging. Mol. Metab. 2023, 74, 101755. [Google Scholar] [CrossRef]
- Kuribara, T.; Totani, K. Oligomannose-type glycan processing in the endoplasmic reticulum and its importance in misfolding diseases. Biology 2022, 11, 199. [Google Scholar] [CrossRef]
- Miyahara, K.; Nouso, K.; Saito, S.; Hiraoka, S.; Harada, K.; Takahashi, S.; Morimoto, Y.; Kobayashi, S.; Ikeda, F.; Miyake, Y.; et al. Serum glycan markers for evaluation of disease activity and prediction of clinical course in patients with ulcerative colitis. PLoS ONE 2013, 8, e74861. [Google Scholar] [CrossRef]
- Becker, D.J.; Lowe, J.B. Fucose: Biosynthesis and biological function in mammals. Glycobiology 2003, 13, 41R–53R. [Google Scholar] [CrossRef]
- Sillanaukee, P.; Pönniö, M.; Jääskeläinen, I.P. Occurrence of sialic acids in healthy humans and different disorders. Eur. J. Clin. Investig. 1999, 29, 413–425. [Google Scholar] [CrossRef]
- Clerc, F.; Reiding, K.R.; Jansen, B.C.; Lageveen-Kammeijer, G.; Bondt, A.; Wuhrer, M. Human plasma protein N-glycosylation. Glycoconj. J. 2016, 33, 309–343. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| N-Glycan Trait (%) | Total Sample (n = 1516) | Cluster 1 (n = 431) | Cluster 2 (n = 1085) | p-Value |
|---|---|---|---|---|
| G0 | 3.19 (2.40, 4.53) | 5.75 (4.47, 7.92) | 2.76 (2.20, 3.41) | <0.001 |
| G1 | 6.95 (6.05, 8.26) | 9.59 (8.44, 11.60) | 6.41 (5.72, 7.14) | <0.001 |
| G2 | 67.98 (65.43, 69.98) | 64.98 (61.46, 68.21) | 68.57 (66.82, 70.45) | <0.001 |
| G3 | 13.48 (11.54, 15.41) | 11.28 (9.83, 12.95) | 14.30 (12.52, 15.92) | <0.001 |
| G4 | 6.22 (5.09, 7.58) | 5.22 (4.29, 6.23) | 6.66 (5.62, 7.89) | <0.001 |
| A1 | 1.08 (0.95, 1.22) | 1.30 (1.16, 1.49) | 1.01 (0.91, 1.12) | <0.001 |
| A2 | 77.76 (75.70, 79.82) | 80.05 (78.34, 81.48) | 76.92 (75.07, 78.63) | <0.001 |
| A3 | 13.48 (11.54, 15.41) | 11.28 (9.83, 12.95) | 14.30 (12.52, 15.92) | <0.001 |
| A4 | 6.22 (5.09, 7.58) | 5.22 (4.29, 6.23) | 6.66 (5.62, 7.89) | <0.001 |
| S0 | 11.83 (9.82, 14.69) | 17.92 (15.23, 23.93) | 10.66 (9.26, 12.08) | <0.001 |
| S1 | 21.15 (19.48, 22.85) | 21.37 (19.59, 23.27) | 21.08 (19.45, 22.63) | 0.085 |
| S2 | 49.50 (47.16, 51.29) | 45.02 (42.05, 47.60) | 50.39 (49.00, 52.03) | <0.001 |
| S3 | 14.87 (13.06, 16.59) | 12.57 (11.17, 13.96) | 15.68 (14.35, 17.15) | <0.001 |
| S4 | 1.82 (1.50, 2.13) | 1.48 (1.26, 1.76) | 1.93 (1.68, 2.23) | <0.001 |
| CF | 30.44 (27.23, 33.93) | 35.68 (32.65, 39.53) | 28.76 (26.09, 31.45) | <0.001 |
| OF | 2.67 (2.28, 3.12) | 2.30 (1.96, 2.61) | 2.83 (2.45, 3.25) | <0.001 |
| OM | 1.10 (0.91, 1.40) | 1.76 (1.40, 2.65) | 0.99 (0.86, 1.15) | <0.001 |
| Cluster 1 (n = 431) | Cluster 2 (n = 1085) | p-Value | |
|---|---|---|---|
| Age, years | 56 (40, 70) | 51 (38.00, 65) | 0.002 |
| Women, n (%) | 232 (53.8) | 606 (55.8) | 0.608 |
| Men, n (%) | 199 (46.2) | 479 (44.2) | |
| Smoking status | |||
| Never smokers, n (%) | 237 (55.0) | 588 (54.2) | 0.031 |
| Ex-smokers, n (%) | 128 (29.7) | 267 (24.6) | |
| Smokers, n (%) | 66 (15.3) | 230 (21.2) | |
| Physical activity | |||
| Low, n (%) | 165 (38.3) | 431 (39.7) | 0.891 |
| Medium, n (%) | 161 (37.3) | 391 (36.0) | |
| High, n (%) | 105 (24.4) | 263 (24.2) | |
| Alcohol consumption (g/day) | |||
| 0–9, n (%) | 159 (36.9) | 387 (35.7) | 0.468 |
| 10–139, n (%) | 162 (37.6) | 436 (40.2) | |
| 140–279, n (%) | 78 (18.1) | 163 (15.0) | |
| ≥280, n (%) | 32 (7.4) | 99 (9.1) | |
| Body mass index, kg/m2 | 27.7 (24.5, 31.5) | 27.7 (24.6, 31.3) | 0.726 |
| Diabetes mellitus, n (%) | 61 (14.2) | 126 (11.6) | 0.358 |
| Metabolic syndrome, n (%) | 95 (22.0) | 219 (20.2) | 0.608 |
| Serum glucose, mg/dL | 91 (83, 102) | 88 (81, 98) | 0.006 |
| Blood glycated hemoglobin (HbA1c), % | 5.4 (5.2, 5.8) | 5.4 (5.2, 5.7) | 0.752 |
| Serum glycated albumin, % | 14.1 (13.0, 15.5) | 13.6 (12.4, 14.8) | <0.001 |
| Serum fructosamine, μmol/L | 262 (237, 292) | 251 (220, 279) | <0.001 |
| Serum HDL-cholesterol, mg/dL | 59 (49, 71) | 57 (46, 68) | 0.006 |
| Serum LDL-cholesterol, mg/dL | 114 (93, 137) | 113 (94, 134) | 0.905 |
| Erythrocyte sedimentation rate (ESR), mm/h | 9 (5, 17) | 9 (5, 16) | 0.326 |
| Serum C-reactive protein, mg/dL | 0.13 (0.04, 0.33) | 0.14 (0.05, 0.41) | 0.358 |
| Serum TNF-alpha, pg/mL | 7.6 (6.4, 9.6) | 7.4 (6.0, 8.8) | 0.001 |
| Serum interleukin-8 (IL-8), pg/mL | 7.0 (5.0, 10.0) | 7.0 (5.0, 11.0) | 0.608 |
| Serum interleukin-6 (IL-6), pg/mL | 2.1 (2.0, 3.2) | 2.2 (2.0, 3.6) | 0.358 |
| Serum soluble interleukin-2 receptor, U/mL | 414 (321, 527) | 411 (313, 530) | 0.608 |
| Serum aspartate aminotransferase (AST), IU/L | 23 (20, 28) | 22 (19, 27) | 0.002 |
| Gamma-glutamyl transferase (GGT), IU/L | 21 (14, 34) | 19 (13, 33) | 0.069 |
| Serum triiodothyronine (T3), pg/mL | 3.30 (3.05, 3.56) | 3.39 (3.15, 3.67) | 0.001 |
| Glomerular filtration rate, mL/min/1.7 m2 | 96.8 (84.5, 111.8) | 101.9 (88.3, 117.2) | 0.001 |
| Comorbidity index | |||
| 0 points | 296 (68.7) | 789 (72.7) | 0.015 |
| 1 point | 84 (19.5) | 222 (20.5) | |
| ≥2 points | 51 (11.8) | 74 (6.8) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lado-Baleato, Ó.; Torre, J.; O’Flaherty, R.; Alonso-Sampedro, M.; Carballo, I.; Fernández-Merino, C.; Vidal, C.; Gude, F.; Saldova, R.; González-Quintela, A. Age-Related Changes in Serum N-Glycome in Men and Women—Clusters Associated with Comorbidity. Biomolecules 2024, 14, 17. https://doi.org/10.3390/biom14010017
Lado-Baleato Ó, Torre J, O’Flaherty R, Alonso-Sampedro M, Carballo I, Fernández-Merino C, Vidal C, Gude F, Saldova R, González-Quintela A. Age-Related Changes in Serum N-Glycome in Men and Women—Clusters Associated with Comorbidity. Biomolecules. 2024; 14(1):17. https://doi.org/10.3390/biom14010017
Chicago/Turabian StyleLado-Baleato, Óscar, Jorge Torre, Róisín O’Flaherty, Manuela Alonso-Sampedro, Iago Carballo, Carmen Fernández-Merino, Carmen Vidal, Francisco Gude, Radka Saldova, and Arturo González-Quintela. 2024. "Age-Related Changes in Serum N-Glycome in Men and Women—Clusters Associated with Comorbidity" Biomolecules 14, no. 1: 17. https://doi.org/10.3390/biom14010017