
Cannabinoids in Medicine: A Multifaceted Exploration of Types, Therapeutic Applications, and Emerging Opportunities in Neurodegenerative Diseases and Cancer Therapy


 , ,
, ,
Abstract
:1. Introduction
1.1. Background on Cannabinoids
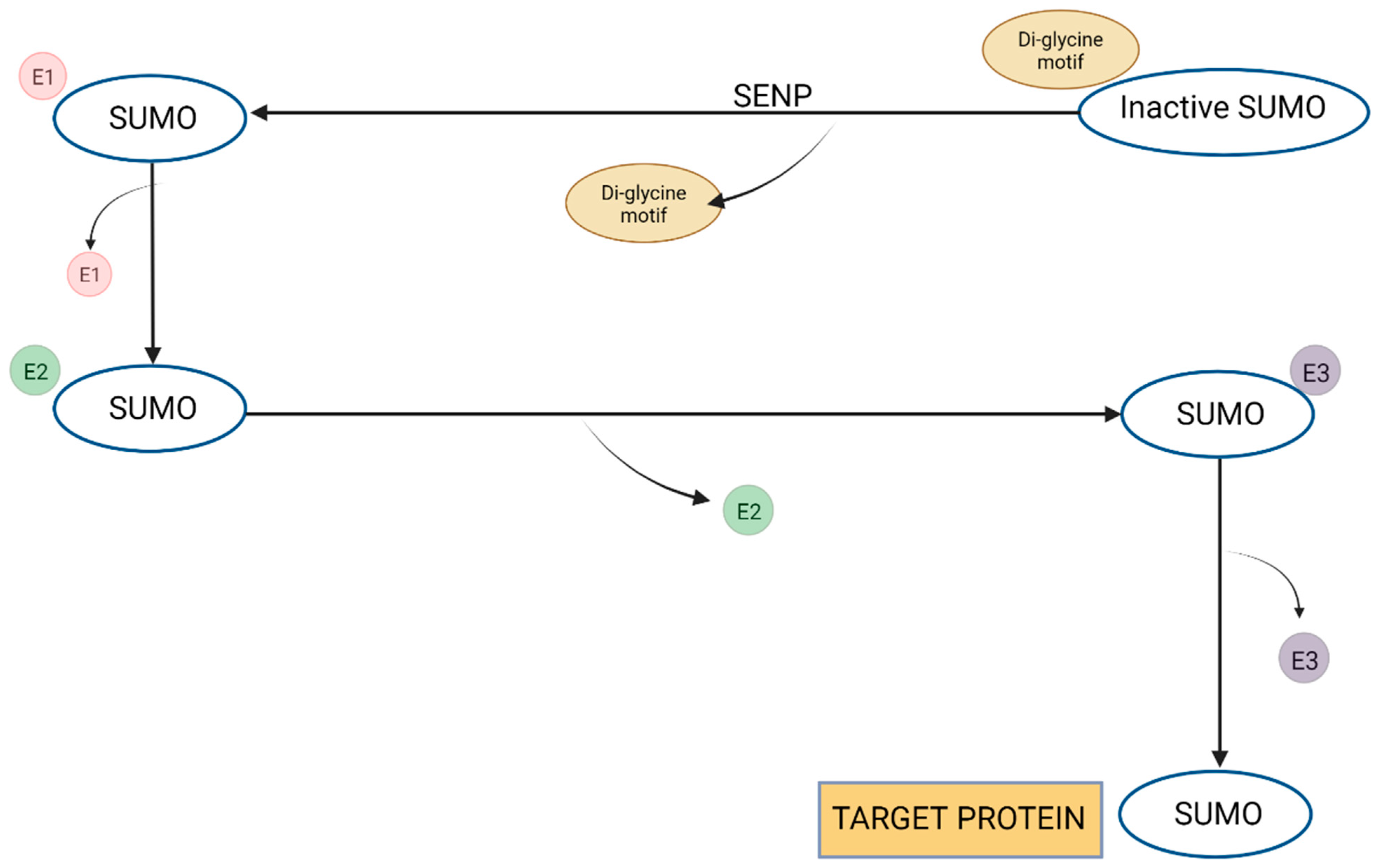
1.2. Importance of Endogenous Cannabinoids
1.3. Overview of Neurodegenerative Diseases and Cancer
1.4. Significance of Studying Cannabinoid Effects
2. Cannabinoids: Types and Mechanisms of Action
2.1. Phytocannabinoids
2.2. Endocannabinoids
2.3. Synthetic Cannabinoids
2.4. Cannabinoid Receptors and Signaling Pathways
3. The Pharmacology of Cannabinoids
3.1. Absorption, Distribution, Metabolism, and Excretion (ADME)
3.2. Pharmacokinetics and Pharmacodynamics
3.3. Factors Influencing Cannabinoid Effects
4. Cannabinoids and Neurodegenerative Diseases
4.1. Alzheimer’s Disease
4.2. Parkinson’s Disease
4.3. Huntington’s Disease
4.4. Multiple Sclerosis
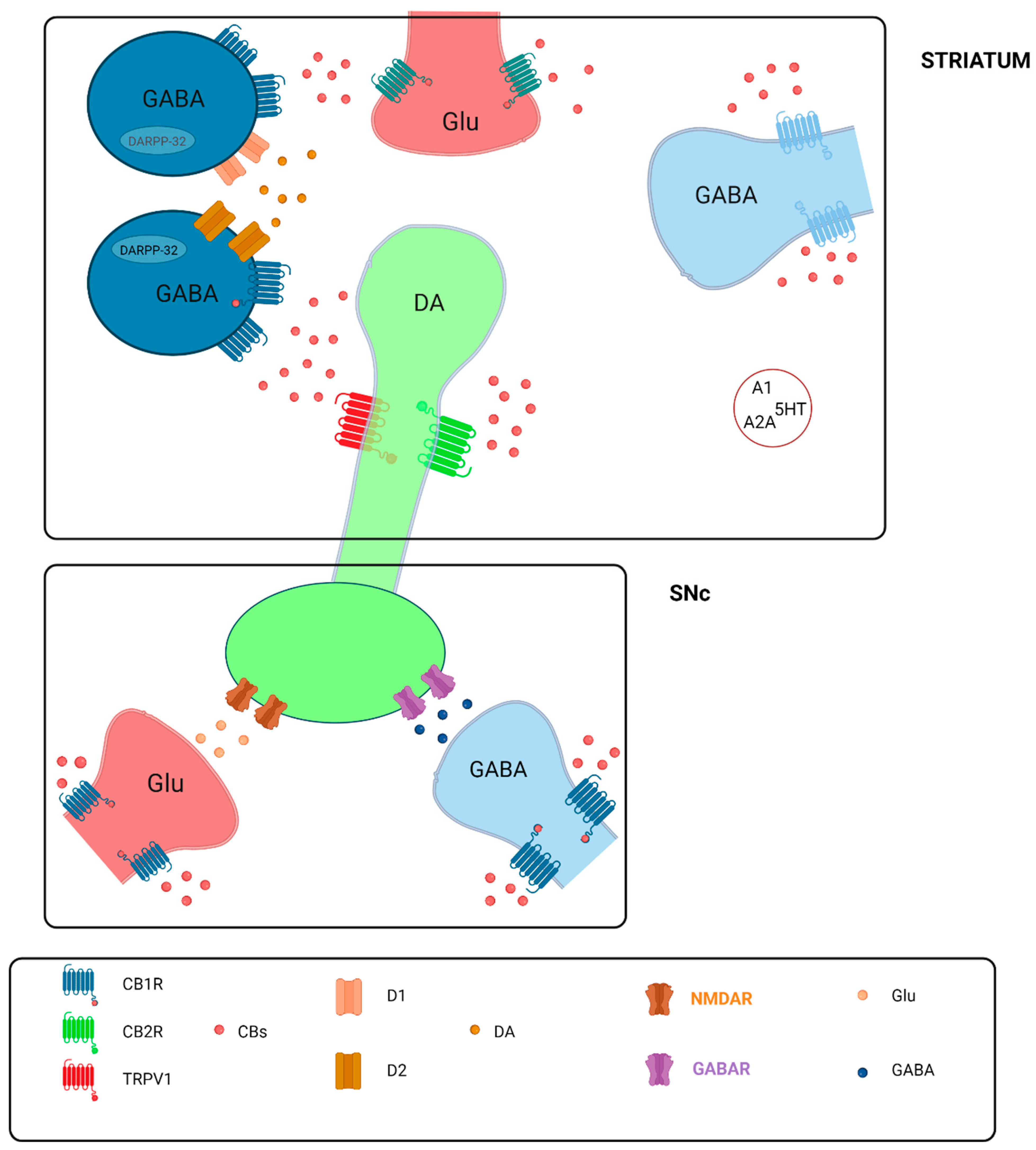
4.5. Mechanisms of Cannabinoid Action in Neurodegeneration
5. Cannabinoids and Cancer
5.1. Antitumor Effects of Cannabinoids
5.2. Cannabinoids in Cancer Therapy
5.3. Potential Mechanisms of Cannabinoid-Mediated Anticancer Effects
6. Clinical Applications and Challenges
6.1. Cannabinoids as Therapeutic Agents
6.2. Clinical Trials and Evidence-Based Medicine
6.3. Safety Considerations and Adverse Effects
7. Future Perspectives and Conclusions
7.1. Promising Avenues for Future Research
7.2. Implications for Cannabinoid-Based Therapies
7.3. Concluding Remarks on the Potential of Cannabinoids
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CHS | Cannabinoid Hyperemesis Syndrome |
| THC | Δ9-tetrahydrocannabinol THC |
| ECS | The Endocannabinoid System |
| CNS | Central nervous system |
| eCBs | Endocannabinoids |
| CBD | Cannabidiol |
| THCV | D9-tetrahydrocannabivarin |
| ROS | Reactive Oxygen Species |
| CBRs | Cannabinoid Receptors |
| DRO | Dronabinol |
| CBG | Cannbigerol |
| CBC | Cannabichromene |
| CBDV | Cannabidivarin |
| GPCR | G Protein-Coupled Receptor |
| CBDA | Cannabidiolic Acid |
| CBN | Cannabinol |
| CBL | Cannabicyclol |
| CBE | Cannabielsoin |
| CBF | Cannabifuran |
| TRP | Transient Receptor Potential |
| PPAR | Peroxisome Proliferator-Activated Receptors |
| TRPV1 | Transient Receptor Potential Vanilloid Type 1 |
| DSI | Depolarization-Induced Suppression of Inhibition |
| DSE | Depolarization-Induced Suppression of Excitation |
| 5-HT | 5-hydroxytryptamine |
| CYP450 | Cytochrome P450 |
| D11-OH-THC | D11-hydroxy-THC |
| D11-COOH-THC | D11-carboxy-THC |
| CBND | Cannabinodiol |
| EEG | Electroencephalography |
| ERP | Event Related Potentials |
| AD | Alzheimer’s Disease |
| Ab | Amyloid Beta |
| PSEN1 | Presenilin-1 |
| PSEN2 | Presenilin-2 |
| BDNF | Brain-Derived Neurotrophic Factor |
| LOAD | Late-Onset Alzheimer’s Disease |
| PD | Parkinson’s disease |
| LIDs | Levodopa-Induced Dyskinesias |
| GPl | Globus Pallidus |
| HD | Huntington’s Disease |
| MS | Multiple Sclerosis |
| CREA | Chronic Relapsing Experimental Allergic Encephalomyelitis |
| AEA | Anandamide |
| FAAH | Fatty Acid Amide Hydrolase |
| MCAO | Middle Cerebral Artery Occlusion |
| NMDA | N-methyl-D-aspartate |
| PEA | Palmitoylethanolamide |
| OEA | Oleoylethanolamide |
| AraS | N-arachidonoyl-L-serine |
| MAPK | Mitogen-Activated Protein Kinase |
| CRC | Colorectal Cancer |
| MAGL | Monoacylglycerol Lipase |
| LC3 | Light Chain 3 |
| NFAT | Nuclear Factor of Activated T Cells |
| LNCaP | Lymph Node Carcinoma of the Prostate |
| GB | Glioblastoma |
| TMZ | Temozolomide |
| CSCs | Cancer Stem Cells |
| PWH | People Living with HIV |
| ART | Antiretroviral Therapy |
| DDIs | Drug–Drug Interactions |
| SSRIs | Selective Serotonin Reuptake Inhibitors |
| OTC | Over the Counter |
| IBD | Inflammatory Bowel Disease |
| UC | Ulcerative Colitis |
| CD | Crohn’s Disease |
| BMT | Bone Marrow Transplant |
| EAE | Experimental Autoimmune Encephalomyelitis |
| MM | Medical Marijuana |
References
- Vogel, Z.; Barg, J.; Levy, R.; Saya, D.; Heldman, E.; Mechoulam, R. Anandamide, a Brain Endogenous Compound, Interacts Specifically with Cannabinoid Receptors and Inhibits Adenylate Cyclase. J. Neurochem. 1993, 61, 352–355. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, C.Y.; Gilchrist, B.L. A Case of Cannabinoid Hyperemesis Syndrome Caused by Synthetic Cannabinoids. J. Emerg. Med. 2013, 45, 544–546. [Google Scholar] [CrossRef] [PubMed]
- Sawzdargo, M.; Nguyen, T.; Lee, D.K.; Lynch, K.R.; Cheng, R.; Heng, H.H.Q.; George, S.R.; O’Dowd, B.F. Identification and cloning of three novel human G protein-coupled receptor genes GPR52, ΨGPR53 and GPR55: GPR55 is extensively expressed in human brain. Mol. Brain Res. 1999, 64, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Overton, H.A.; Babbs, A.J.; Doel, S.M.; Fyfe, M.C.T.; Gardner, L.S.; Griffin, G.; Jackson, H.C.; Procter, M.J.; Rasamison, C.M.; Tang-Christensen, M.; et al. Deorphanization of a G protein-coupled receptor for oleoylethanolamide and its use in the discovery of small-molecule hypophagic agents. Cell Metab. 2006, 3, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Hansen, K.B.; Rosenkilde, M.M.; Knop, F.K.; Wellner, N.; Diep, T.A.; Rehfeld, J.F.; Andersen, U.B.; Holst, J.J.; Hansen, H.S. 2-Oleoyl Glycerol Is a GPR119 Agonist and Signals GLP-1 Release in Humans. J. Clin. Endocrinol. Metab. 2011, 96, E1409–E1417. [Google Scholar] [CrossRef]
- Luk, T.; Jin, W.; Zvonok, A.; Lu, D.; Lin, X.-Z.; Chavkin, C.; Makriyannis, A.; Mackie, K. Identification of a potent and highly efficacious, yet slowly desensitizing CB1 cannabinoid receptor agonist: An efficacious, slowly desensitizing CB1 agonist. Br. J. Pharmacol. 2004, 142, 495–500. [Google Scholar] [CrossRef]
- Gonsiorek, W.; Lunn, C.; Fan, X.; Narula, S.; Lundell, D.; Hipkin, R.W. Endocannabinoid 2-arachidonyl glycerol is a full agonist through human type 2 cannabinoid receptor: Antagonism by anandamide. Mol. Pharmacol. 2000, 57, 1045–1050. [Google Scholar]
- Mackie, K.; Devane, W.A.; Hille, B. Anandamide, an endogenous cannabinoid, inhibits calcium currents as a partial agonist in N18 neuroblastoma cells. Mol. Pharmacol. 1993, 44, 498–503. [Google Scholar]
- Hanuš, L.; Abu-Lafi, S.; Fride, E.; Breuer, A.; Vogel, Z.; Shalev, D.E.; Kustanovich, I.; Mechoulam, R. 2-Arachidonyl glyceryl ether, an endogenous agonist of the cannabinoid CB1 receptor. Proc. Natl. Acad. Sci. USA 2001, 98, 3662–3665. [Google Scholar] [CrossRef]
- Porter, A.C.; Sauer, J.-M.; Knierman, M.D.; Becker, G.W.; Berna, M.J.; Bao, J.; Nomikos, G.G.; Carter, P.; Bymaster, F.P.; Leese, A.B.; et al. Characterization of a Novel Endocannabinoid, Virodhamine, with Antagonist Activity at the CB1 Receptor. J. Pharmacol. Exp. Ther. 2002, 301, 1020–1024. [Google Scholar] [CrossRef]
- Huang, S.M.; Bisogno, T.; Trevisani, M.; Al-Hayani, A.; De Petrocellis, L.; Fezza, F.; Tognetto, M.; Petros, T.J.; Krey, J.F.; Chu, C.J.; et al. An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc. Natl. Acad. Sci. USA 2002, 99, 8400–8405. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.M.; Bisogno, T.; Petros, T.J.; Chang, S.Y.; Zavitsanos, P.A.; Zipkin, R.E.; Sivakumar, R.; Coop, A.; Maeda, D.Y.; De Petrocellis, L.; et al. Identification of a New Class of Molecules, the Arachidonyl Amino Acids, and Characterization of One Member That Inhibits Pain. J. Biol. Chem. 2001, 276, 42639–42644. [Google Scholar] [CrossRef] [PubMed]
- Michael Rossi, A.; Kuehnle, J.C.; Mendelson, J.H. Marihuana and mood in human volunteers. Pharmacol. Biochem. Behav. 1978, 8, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Abel, E.L. Cannabis: Effects on hunger and thirst. Behav. Biol. 1975, 15, 255–281. [Google Scholar] [CrossRef]
- Schmid, P.C.; Paria, B.C.; Krebsbach, R.J.; Schmid, H.H.O.; Dey, S.K. Changes in anandamide levels in mouse uterus are associated with uterine receptivity for embryo implantation. Proc. Natl. Acad. Sci. USA 1997, 94, 4188–4192. [Google Scholar] [CrossRef]
- Paria, B.C.; Huet-Hudson, Y.M.; Dey, S.K. Blastocyst’s state of activity determines the “window” of implantation in the receptive mouse uterus. Proc. Natl. Acad. Sci. USA 1993, 90, 10159–10162. [Google Scholar] [CrossRef]
- Paria, B.C.; Deutsch, D.D.; Dey, S.K. The uterus is a potential site for anandamide synthesis and hydrolysis: Differential profiles of anandamide synthase and hydrolase activities in the mouse uterus during the periimplantation period. Mol. Reprod. Dev. 1996, 45, 183–192. [Google Scholar] [CrossRef]
- Kunos, G.; Osei-Hyiaman, D.; Bátkai, S.; Sharkey, K.A.; Makriyannis, A. Should peripheral CB1 cannabinoid receptors be selectively targeted for therapeutic gain? Trends Pharmacol. Sci. 2009, 30, 1–7. [Google Scholar] [CrossRef]
- Scutt, A.; Williamson, E.M. Cannabinoids Stimulate Fibroblastic Colony Formation by Bone Marrow Cells Indirectly via CB2 Receptors. Calcif. Tissue Int. 2007, 80, 50–59. [Google Scholar] [CrossRef]
- Mori, M.A.; Meyer, E.; Soares, L.M.; Milani, H.; Guimarães, F.S.; De Oliveira, R.M.W. Cannabidiol reduces neuroinflammation and promotes neuroplasticity and functional recovery after brain ischemia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 75, 94–105. [Google Scholar] [CrossRef]
- Pazos, M.R.; Cinquina, V.; Gómez, A.; Layunta, R.; Santos, M.; Fernández-Ruiz, J.; Martínez-Orgado, J. Cannabidiol administration after hypoxia–ischemia to newborn rats reduces long-term brain injury and restores neurobehavioral function. Neuropharmacology 2012, 63, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Hartsel, J.A.; Eades, J.; Hickory, B.; Makriyannis, A. Cannabis sativa and Hemp. In Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2016; pp. 735–754. ISBN 978-0-12-802147-7. [Google Scholar] [CrossRef]
- Aziz, A.; Nguyen, L.C.; Oumeslakht, L.; Bensussan, A.; Ben Mkaddem, S. Cannabinoids as Immune System Modulators: Cannabidiol Potential Therapeutic Approaches and Limitations. Cannabis Cannabinoid Res. 2023, 8, 254–269. [Google Scholar] [CrossRef] [PubMed]
- Coghe, G.; Pau, M.; Corona, F.; Frau, J.; Lorefice, L.; Fenu, G.; Spinicci, G.; Mamusa, E.; Musu, L.; Massole, S.; et al. Walking improvements with nabiximols in patients with multiple sclerosis. J. Neurol. 2015, 262, 2472–2477. [Google Scholar] [CrossRef] [PubMed]
- Flachenecker, P.; Henze, T.; Zettl, U.K. Nabiximols (THC/CBD Oromucosal Spray, Sativex®) in Clinical Practice—Results of a Multicenter, Non-Interventional Study (MOVE 2) in Patients with Multiple Sclerosis Spasticity. Eur. Neurol. 2014, 71, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Rosales, F.; Walsh, D. Intractable nausea and vomiting due to gastrointestinal mucosal metastases relieved by tetrahydrocannabinol (Dronabinol). J. Pain Symptom Manag. 1997, 14, 311–314. [Google Scholar] [CrossRef]
- Hernandez, S.L.; Sheyner, I.; Stover, K.T.; Stewart, J.T. Dronabinol Treatment of Refractory Nausea and Vomiting Related to Peritoneal Carcinomatosis. Am. J. Hosp. Palliat. Care 2015, 32, 5–7. [Google Scholar] [CrossRef]
- Beal, J.E.; Olson, R.; Laubenstein, L.; Morales, J.O.; Bellman, P.; Yangco, B.; Lefkowitz, L.; Plasse, T.F.; Shepard, K.V. Dronabinol as a treatment for anorexia associated with weight loss in patients with AIDS. J. Pain Symptom Manag. 1995, 10, 89–97. [Google Scholar] [CrossRef]
- DeJesus, E.; Rodwick, B.M.; Bowers, D.; Cohen, C.J.; Pearce, D. Use of Dronabinol Improves Appetite and Reverses Weight Loss in HIV/AIDS-Infected Patients. J. Int. Assoc. Physicians AIDS Care 2007, 6, 95–100. [Google Scholar] [CrossRef]
- McPartland, J.M.; Glass, M.; Pertwee, R.G. Meta-analysis of cannabinoid ligand binding affinity and receptor distribution: Interspecies differences: Ligand affinity and receptor distribution of CB1. Br. J. Pharmacol. 2007, 152, 583–593. [Google Scholar] [CrossRef]
- Stockings, E.; Campbell, G.; Hall, W.D.; Nielsen, S.; Zagic, D.; Rahman, R.; Murnion, B.; Farrell, M.; Weier, M.; Degenhardt, L. Cannabis and cannabinoids for the treatment of people with chronic noncancer pain conditions: A systematic review and meta-analysis of controlled and observational studies. Pain 2018, 159, 1932–1954. [Google Scholar] [CrossRef]
- Castellanos, D.; Singh, S.; Thornton, G.; Avila, M.; Moreno, A. Synthetic Cannabinoid Use: A Case Series of Adolescents. J. Adolesc. Health 2011, 49, 347–349. [Google Scholar] [CrossRef]
- Esfandyari, T.; Camilleri, M.; Ferber, I.; Burton, D.; Baxter, K.; Zinsmeister, A.R. Effect of a cannabinoid agonist on gastrointestinal transit and postprandial satiation in healthy human subjects: A randomized, placebo-controlled study. Neurogastroenterol. Motil. 2006, 18, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Toyota, M.; Shimamura, T.; Ishii, H.; Renner, M.; Braggins, J.; Asakawa, Y. New Bibenzyl Cannabinoid from the New Zealand Liverwort Radula marginata. Chem. Pharm. Bull. 2002, 50, 1390–1392. [Google Scholar] [CrossRef] [PubMed]
- Iwata, N.; Kitanaka, S. New Cannabinoid-Like Chromane and Chromene Derivatives from Rhododendron anthopogonoides. Chem. Pharm. Bull. 2011, 59, 1409–1412. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular Targets of the Phytocannabinoids: A Complex Picture. In Phytocannabinoids; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J., Eds.; Progress in the Chemistry of Organic Natural Products; Springer International Publishing: Cham, Switzerland, 2017; Volume 103, pp. 103–131. ISBN 978-3-319-45539-6. [Google Scholar] [CrossRef]
- Howlett, A.C. International Union of Pharmacology. XXVII. Classification of Cannabinoid Receptors. Pharmacol. Rev. 2002, 54, 161–202. [Google Scholar] [CrossRef]
- Kogan, N.M.; Rabinowitz, R.; Levi, P.; Gibson, D.; Sandor, P.; Schlesinger, M.; Mechoulam, R. Synthesis and Antitumor Activity of Quinonoid Derivatives of Cannabinoids. J. Med. Chem. 2004, 47, 3800–3806. [Google Scholar] [CrossRef]
- Granja, A.G.; Carrillo-Salinas, F.; Pagani, A.; Gómez-Cañas, M.; Negri, R.; Navarrete, C.; Mecha, M.; Mestre, L.; Fiebich, B.L.; Cantarero, I.; et al. A Cannabigerol Quinone Alleviates Neuroinflammation in a Chronic Model of Multiple Sclerosis. J. Neuroimmune Pharmacol. 2012, 7, 1002–1016. [Google Scholar] [CrossRef]
- Carrillo-Salinas, F.J.; Navarrete, C.; Mecha, M.; Feliú, A.; Collado, J.A.; Cantarero, I.; Bellido, M.L.; Muñoz, E.; Guaza, C. A Cannabigerol Derivative Suppresses Immune Responses and Protects Mice from Experimental Autoimmune Encephalomyelitis. PLoS ONE 2014, 9, e94733. [Google Scholar] [CrossRef]
- Dos Santos, R.G.; Hallak, J.E.C.; Crippa, J.A.S. Neuropharmacological Effects of the Main Phytocannabinoids: A Narrative Review. In Cannabinoids and Neuropsychiatric Disorders; Murillo-Rodriguez, E., Pandi-Perumal, S.R., Monti, J.M., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2021; Volume 1264, pp. 29–45. ISBN 978-3-030-57368-3. [Google Scholar] [CrossRef]
- Shrader, S.H.; Tong, Y.-G.; Duff, M.B.; Freedman, J.H.; Song, Z.-H. Involvement of dopamine receptor in the actions of non-psychoactive phytocannabinoids. Biochem. Biophys. Res. Commun. 2020, 533, 1366–1370. [Google Scholar] [CrossRef]
- Smaga, I.; Bystrowska, B.; Gawlinski, D.; Przegalinski, E.; Filip, M. The Endocannabinoid/Endovanilloid System and Depression. Curr. Neuropharmacol. 2014, 12, 462–474. [Google Scholar] [CrossRef]
- Shen, C.-J.; Zheng, D.; Li, K.-X.; Yang, J.-M.; Pan, H.-Q.; Yu, X.-D.; Fu, J.-Y.; Zhu, Y.; Sun, Q.-X.; Tang, M.-Y.; et al. Cannabinoid CB1 receptors in the amygdalar cholecystokinin glutamatergic afferents to nucleus accumbens modulate depressive-like behavior. Nat. Med. 2019, 25, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Boulebd, H. Insights on the antiradical capacity and mechanism of phytocannabinoids: H-abstraction and electron transfer processes in physiological media and the influence of the acid-base equilibrium. Phytochemistry 2023, 208, 113608. [Google Scholar] [CrossRef] [PubMed]
- Hua, T.; Vemuri, K.; Pu, M.; Qu, L.; Han, G.W.; Wu, Y.; Zhao, S.; Shui, W.; Li, S.; Korde, A.; et al. Crystal Structure of the Human Cannabinoid Receptor CB1. Cell 2016, 167, 750–762.e14. [Google Scholar] [CrossRef] [PubMed]
- Navarro, G.; Varani, K.; Lillo, A.; Vincenzi, F.; Rivas-Santisteban, R.; Raïch, I.; Reyes-Resina, I.; Ferreiro-Vera, C.; Borea, P.A.; Sánchez De Medina, V.; et al. Pharmacological data of cannabidiol- and cannabigerol-type phytocannabinoids acting on cannabinoid CB1, CB2 and CB1/CB2 heteromer receptors. Pharmacol. Res. 2020, 159, 104940. [Google Scholar] [CrossRef]
- Devane, W.A.; Hanuš, L.; Breuer, A.; Pertwee, R.G.; Stevenson, L.A.; Griffin, G.; Gibson, D.; Mandelbaum, A.; Etinger, A.; Mechoulam, R. Isolation and Structure of a Brain Constituent That Binds to the Cannabinoid Receptor. Science 1992, 258, 1946–1949. [Google Scholar] [CrossRef]
- Wang, H.; Matsumoto, H.; Guo, Y.; Paria, B.C.; Roberts, R.L.; Dey, S.K. Differential G protein-coupled cannabinoid receptor signaling by anandamide directs blastocyst activation for implantation. Proc. Natl. Acad. Sci. USA 2003, 100, 14914–14919. [Google Scholar] [CrossRef]
- Maccarrone, M.; Rossi, S.; Bari, M.; De Chiara, V.; Fezza, F.; Musella, A.; Gasperi, V.; Prosperetti, C.; Bernardi, G.; Finazzi-Agrò, A.; et al. Anandamide inhibits metabolism and physiological actions of 2-arachidonoylglycerol in the striatum. Nat. Neurosci. 2008, 11, 152–159. [Google Scholar] [CrossRef]
- Piomelli, D.; Beltramo, M.; Giuffrida, A.; Stella, N. Endogenous Cannabinoid Signaling. Neurobiol. Dis. 1998, 5, 462–473. [Google Scholar] [CrossRef]
- Liu, J.; Wang, L.; Harvey-White, J.; Osei-Hyiaman, D.; Razdan, R.; Gong, Q.; Chan, A.C.; Zhou, Z.; Huang, B.X.; Kim, H.-Y.; et al. A biosynthetic pathway for anandamide. Proc. Natl. Acad. Sci. USA 2006, 103, 13345–13350. [Google Scholar] [CrossRef]
- González, S.; Mena, M.A.; Lastres-Becker, I.; Serrano, A.; De Yébenes, J.G.; Ramos, J.A.; Fernández-Ruiz, J. Cannabinoid CB1 receptors in the basal ganglia and motor response to activation or blockade of these receptors in parkin-null mice. Brain Res. 2005, 1046, 195–206. [Google Scholar] [CrossRef]
- Bidaut-Russell, M.; Hewlett, A.C. Cannabinoid Receptor-Regulated Cyclic AMP Accumulation in the Rat Striatum. J. Neurochem. 1991, 57, 1769–1773. [Google Scholar] [CrossRef]
- Fan, S.-F.; Yazulla, S. Inhibitory interaction of cannabinoid CB1 receptor and dopamine D2 receptor agonists on voltage-gated currents of goldfish cones. Vis. Neurosci. 2004, 21, 69–77. [Google Scholar] [CrossRef]
- Navarro, M.; Hernández, E.; Muñoz, R.M.; Del Arco, I.; Villanúa, M.A.; Carrera, M.R.A.; Rodríguez De Fonseca, F. Acute administration of the CB1 cannabinoid receptor antagonist SR 141716A induces anxiety-like responses in the rat. NeuroReport 1997, 8, 491–496. [Google Scholar] [CrossRef]
- Muller, C.; Lynch, D.L.; Hurst, D.P.; Reggio, P.H. A Closer Look at Anandamide Interaction With TRPV1. Front. Mol. Biosci. 2020, 7, 144. [Google Scholar] [CrossRef]
- Russo, R.; LoVerme, J.; La Rana, G.; D’Agostino, G.; Sasso, O.; Calignano, A.; Piomelli, D. Synergistic antinociception by the cannabinoid receptor agonist anandamide and the PPAR-α receptor agonist GW7647. Eur. J. Pharmacol. 2007, 566, 117–119. [Google Scholar] [CrossRef]
- Bouaboula, M.; Hilairet, S.; Marchand, J.; Fajas, L.; Fur, G.L.; Casellas, P. Anandamide induced PPARγ transcriptional activation and 3T3-L1 preadipocyte differentiation. Eur. J. Pharmacol. 2005, 517, 174–181. [Google Scholar] [CrossRef]
- Falenski, K.W.; Thorpe, A.J.; Schlosburg, J.E.; Cravatt, B.F.; Abdullah, R.A.; Smith, T.H.; Selley, D.E.; Lichtman, A.H.; Sim-Selley, L.J. FAAH−/− Mice Display Differential Tolerance, Dependence, and Cannabinoid Receptor Adaptation after Δ9-Tetrahydrocannabinol and Anandamide Administration. Neuropsychopharmacology 2010, 35, 1775–1787. [Google Scholar] [CrossRef]
- Xiang, W.; Shi, R.; Kang, X.; Zhang, X.; Chen, P.; Zhang, L.; Hou, A.; Wang, R.; Zhao, Y.; Zhao, K.; et al. Monoacylglycerol lipase regulates cannabinoid receptor 2-dependent macrophage activation and cancer progression. Nat. Commun. 2018, 9, 2574. [Google Scholar] [CrossRef]
- Ueda, N.; Tsuboi, K.; Uyama, T.; Ohnishi, T. Biosynthesis and degradation of the endocannabinoid 2-arachidonoylglycerol. BioFactors 2011, 37, 1–7. [Google Scholar] [CrossRef]
- Palese, F.; Pontis, S.; Realini, N.; Piomelli, D. NAPE-specific phospholipase D regulates LRRK2 association with neuronal membranes. In Advances in Pharmacology; Elsevier: Amsterdam, The Netherlands, 2021; Volume 90, pp. 217–238. ISBN 978-0-12-822516-5. [Google Scholar]
- Hillard, C.J.; Edgemond, W.S.; Jarrahian, A.; Campbell, W.B. Accumulation of N-Arachidonoylethanolamine (Anandamide) into Cerebellar Granule Cells Occurs via Facilitated Diffusion. J. Neurochem. 2002, 69, 631–638. [Google Scholar] [CrossRef]
- Pertwee, R.G. GPR55: A new member of the cannabinoid receptor clan?: Commentary. Br. J. Pharmacol. 2007, 152, 984–986. [Google Scholar] [CrossRef]
- Sharir, H.; Abood, M.E. Pharmacological characterization of GPR55, a putative cannabinoid receptor. Pharmacol. Ther. 2010, 126, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.G.; Teixeira, F.M.; Garção, P.; Agostinho, P.; Ledent, C.; Cortes, L.; Mackie, K.; Köfalvi, A. Presynaptic CB1 cannabinoid receptors control frontocortical serotonin and glutamate release—Species differences. Neurochem. Int. 2012, 61, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Heifets, B.D.; Chevaleyre, V.; Castillo, P.E. Interneuron activity controls endocannabinoid-mediated presynaptic plasticity through calcineurin. Proc. Natl. Acad. Sci. USA 2008, 105, 10250–10255. [Google Scholar] [CrossRef]
- Chung, Y.C.; Bok, E.; Huh, S.H.; Park, J.-Y.; Yoon, S.-H.; Kim, S.R.; Kim, Y.-S.; Maeng, S.; Hyun Park, S.; Jin, B.K. Cannabinoid Receptor Type 1 Protects Nigrostriatal Dopaminergic Neurons against MPTP Neurotoxicity by Inhibiting Microglial Activation. J. Immunol. 2011, 187, 6508–6517. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J. The endocannabinoid system as a target for the treatment of motor dysfunction: Endocannabinoids and motor disorders. Br. J. Pharmacol. 2009, 156, 1029–1040. [Google Scholar] [CrossRef]
- Kannampalli, P.; Sengupta, J.N. Role of principal ionotropic and metabotropic receptors in visceral pain. J. Neurogastroenterol. Motil. 2015, 21, 147–158. [Google Scholar] [CrossRef] [PubMed]
- García, M.C.; Cinquina, V.; Palomo-Garo, C.; Rábano, A.; Fernández-Ruiz, J. Identification of CB2 receptors in human nigral neurons that degenerate in Parkinson’s disease. Neurosci. Lett. 2015, 587, 1–4. [Google Scholar] [CrossRef]
- Spaderna, M.; Addy, P.H.; D’Souza, D.C. Spicing things up: Synthetic cannabinoids. Psychopharmacology 2013, 228, 525–540. [Google Scholar] [CrossRef]
- Ogata, J.; Uchiyama, N.; Kikura-Hanajiri, R.; Goda, Y. DNA sequence analyses of blended herbal products including synthetic cannabinoids as designer drugs. Forensic Sci. Int. 2013, 227, 33–41. [Google Scholar] [CrossRef]
- Navarrete, F.; García-Gutiérrez, M.S.; Aracil-Fernández, A.; Lanciego, J.L.; Manzanares, J. Cannabinoid CB1 and CB2 Receptors, and Monoacylglycerol Lipase Gene Expression Alterations in the Basal Ganglia of Patients with Parkinson’s Disease. Neurotherapeutics 2018, 15, 459–469. [Google Scholar] [CrossRef]
- Sierra, S.; Luquin, N.; Rico, A.J.; Gómez-Bautista, V.; Roda, E.; Dopeso-Reyes, I.G.; Vázquez, A.; Martínez-Pinilla, E.; Labandeira-García, J.L.; Franco, R.; et al. Detection of cannabinoid receptors CB1 and CB2 within basal ganglia output neurons in macaques: Changes following experimental parkinsonism. Brain Struct. Funct. 2015, 220, 2721–2738. [Google Scholar] [CrossRef]
- Duncan, M.; Mouihate, A.; Mackie, K.; Keenan, C.M.; Buckley, N.E.; Davison, J.S.; Patel, K.D.; Pittman, Q.J.; Sharkey, K.A. Cannabinoid CB2 receptors in the enteric nervous system modulate gastrointestinal contractility in lipopolysaccharide-treated rats. Am. J. Physiol.-Gastrointest. Liver Physiol. 2008, 295, G78–G87. [Google Scholar] [CrossRef] [PubMed]
- Morgese, M.G.; Cassano, T.; Cuomo, V.; Giuffrida, A. Anti-dyskinetic effects of cannabinoids in a rat model of Parkinson’s disease: Role of CB1 and TRPV1 receptors. Exp. Neurol. 2007, 208, 110–119. [Google Scholar] [CrossRef]
- Valjent, E.; Pagès, C.; Rogard, M.; Besson, M.-J.; Maldonado, R.; Caboche, J. Δ9-tetrahydrocannabinol-induced MAPK/ERK and Elk-1 activation in vivo depends on dopaminergic transmission: THC and ERK activation in vivo. Eur. J. Neurosci. 2001, 14, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Mackie, K.; Hille, B. Cannabinoids inhibit N-type calcium channels in neuroblastoma-glioma cells. Proc. Natl. Acad. Sci. USA 1992, 89, 3825–3829. [Google Scholar] [CrossRef]
- Leterrier, C.; Bonnard, D.; Carrel, D.; Rossier, J.; Lenkei, Z. Constitutive Endocytic Cycle of the CB1 Cannabinoid Receptor. J. Biol. Chem. 2004, 279, 36013–36021. [Google Scholar] [CrossRef]
- Griffin, G.; Tao, Q.; Abood, M.E. Cloning and pharmacological characterization of the rat CB(2) cannabinoid receptor. J. Pharmacol. Exp. Ther. 2000, 292, 886–894. [Google Scholar]
- Sugiura, T.; Waku, K. Cannabinoid Receptors and Their Endogenous Ligands. J. Biochem. 2002, 132, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Herkenham, M.; Lynn, A.B.; Little, M.D.; Johnson, M.R.; Melvin, L.S.; De Costa, B.R.; Rice, K.C. Cannabinoid receptor localization in brain. Proc. Natl. Acad. Sci. USA 1990, 87, 1932–1936. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, S.; Manogar, P.; Prabhu, S.; Pugazhenthi, M.; Praseetha, P.K. A pharmacoinformatic approach on Cannabinoid receptor 2 (CB2) and different small molecules: Homology modelling, molecular docking, MD simulations, drug designing and ADME analysis. Comput. Biol. Chem. 2019, 78, 95–107. [Google Scholar] [CrossRef]
- Altyar, A.E.; Youssef, F.S.; Kurdi, M.M.; Bifari, R.J.; Ashour, M.L. The Role of Cannabis sativa L. as a Source of Cannabinoids against Coronavirus 2 (SARS-CoV-2): An In Silico Study to Evaluate Their Activities and ADMET Properties. Molecules 2022, 27, 2797. [Google Scholar] [CrossRef]
- Sekiguchi, K.; Kanazu, T.; Takeuchi, M.; Hasegawa, H.; Yamaguchi, Y. Non-clinical evaluation of the metabolism, pharmacokinetics and excretion of S-777469, a new cannabinoid receptor 2 selective agonist. Xenobiotica 2014, 44, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Cvan Trobec, K.; Kerec Kos, M.; Trontelj, J.; Grabnar, I.; Tschirner, A.; Palus, S.; Anker, S.D.; Springer, J.; Lainscak, M. Influence of cancer cachexia on drug liver metabolism and renal elimination in rats: Drug liver metabolism and renal elimination in cancer cachexia. J. Cachexia Sarcopenia Muscle 2015, 6, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Caillé, F.; Cacheux, F.; Peyronneau, M.-A.; Jego, B.; Jaumain, E.; Pottier, G.; Ullmer, C.; Grether, U.; Winkeler, A.; Dollé, F.; et al. From Structure–Activity Relationships on Thiazole Derivatives to the In Vivo Evaluation of a New Radiotracer for Cannabinoid Subtype 2 PET Imaging. Mol. Pharm. 2017, 14, 4064–4078. [Google Scholar] [CrossRef] [PubMed]
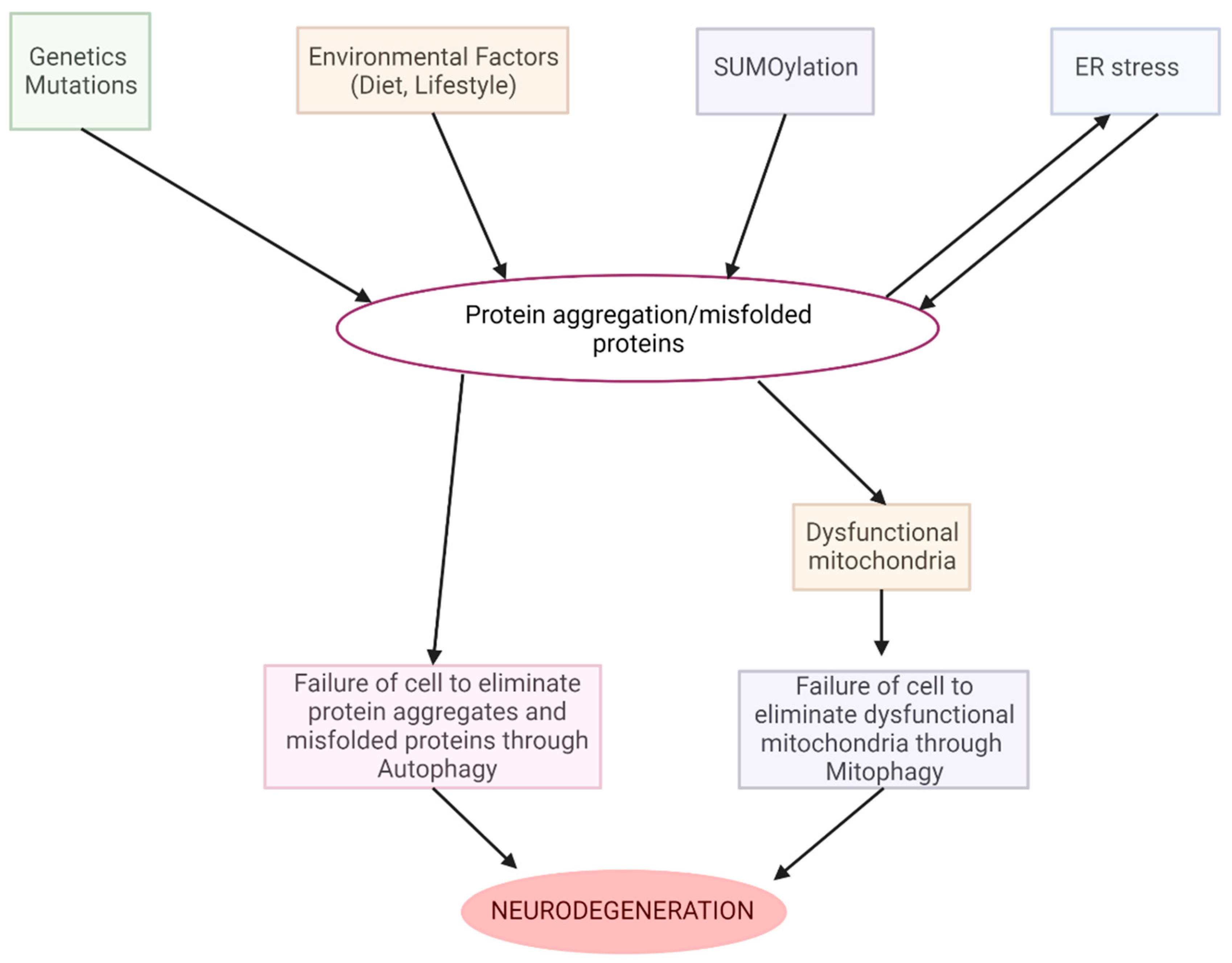
- Basha, F.H.; Waseem, M.; Srinivasan, H. Promising Action of Cannabinoids on ER Stress-Mediated Neurodegeneration: An In Silico Investigation. J. Env. Pathol. Toxicol. Oncol. 2022, 41, 39–54. [Google Scholar] [CrossRef]
- De La Torre, R.; Pichini, S. Usefulness of Sweat Testing for the Detection of Cannabis Smoke. Clin. Chem. 2004, 50, 1961–1962. [Google Scholar] [CrossRef] [PubMed]
- Solowij, N.; Broyd, S.J.; Van Hell, H.H.; Hazekamp, A. A protocol for the delivery of cannabidiol (CBD) and combined CBD and ∆9-tetrahydrocannabinol (THC) by vaporisation. BMC Pharmacol. Toxicol. 2014, 15, 58. [Google Scholar] [CrossRef]
- Martin, J.H.; Schneider, J.; Lucas, C.J.; Galettis, P. Exogenous Cannabinoid Efficacy: Merely a Pharmacokinetic Interaction? Clin. Pharmacokinet. 2018, 57, 539–545. [Google Scholar] [CrossRef]
- Ohlsson, A.; Lindgren, J.-E.; Andersson, S.; Agurell, S.; Gillespie, H.; Hollister, L.E. Single-dose kinetics of deuterium-labelled cannabidiol in man after smoking and intravenous administration. Biol. Mass. Spectrom. 1986, 13, 77–83. [Google Scholar] [CrossRef]
- Grotenhermen, F. Pharmacokinetics and Pharmacodynamics of Cannabinoids: Clin. Pharmacokinet. 2003, 42, 327–360. [Google Scholar] [CrossRef]
- Krotulski, A.J.; Garibay, N.; Walther, D.; Walton, S.E.; Mohr, A.L.A.; Logan, B.K.; Baumann, M.H. Pharmacokinetics and pharmacodynamics of the synthetic cannabinoid, 5F-MDMB-PICA, in male rats. Neuropharmacology 2021, 199, 108800. [Google Scholar] [CrossRef] [PubMed]
- Zamarripa, C.A.; Spindle, T.R.; Surujunarain, R.; Weerts, E.M.; Bansal, S.; Unadkat, J.D.; Paine, M.F.; Vandrey, R. Assessment of Orally Administered Δ9-Tetrahydrocannabinol When Coadministered with Cannabidiol on Δ9-Tetrahydrocannabinol Pharmacokinetics and Pharmacodynamics in Healthy Adults: A Randomized Clinical Trial. JAMA Netw. Open 2023, 6, e2254752. [Google Scholar] [CrossRef]
- Lucas, C.J.; Galettis, P.; Schneider, J. The pharmacokinetics and the pharmacodynamics of cannabinoids. Br. J. Clin. Pharmacol. 2018, 84, 2477–2482. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.; Scheidweiler, K.B.; Wohlfarth, A.; Zhu, M.; Pang, S.; Huestis, M.A. Strategies to distinguish new synthetic cannabinoid FUBIMINA (BIM-2201) intake from its isomer THJ-2201: Metabolism of FUBIMINA in human hepatocytes. Forensic Toxicol. 2016, 34, 256–267. [Google Scholar] [CrossRef]
- Morral, A.R.; McCaffrey, D.F.; Paddock, S.M. Reassessing the marijuana gateway effect: Marijuana gateway effect. Addiction 2002, 97, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Stansfield, C.R.; Somerville, R.F.; Hassan, V.R.; Kolbe, E.; Partington, H.K.; Walsh, K.A.J.; Johnson, C.S. Effects of external influences on synthetic cannabinoid trends in New Zealand, 2014 to 2020. Forensic Sci. Int. 2020, 316, 110485. [Google Scholar] [CrossRef]
- Day, N.L.; Goldschmidt, L.; Day, R.; Larkby, C.; Richardson, G.A. Prenatal marijuana exposure, age of marijuana initiation, and the development of psychotic symptoms in young adults. Psychol. Med. 2015, 45, 1779–1787. [Google Scholar] [CrossRef]
- Tapert, S.; Schweinsburg, A.; Brown, S. The Influence of Marijuana Use on Neurocognitive Functioning in Adolescents. Curr. Drug Abuse Rev. 2008, 1, 99–111. [Google Scholar] [CrossRef]
- Sevigny, E.L.; Pacula, R.L.; Heaton, P. The effects of medical marijuana laws on potency. Int. J. Drug Policy 2014, 25, 308–319. [Google Scholar] [CrossRef]
- Agarwal, C.; Máthé, K.; Hofmann, T.; Csóka, L. Ultrasound-Assisted Extraction of Cannabinoids from Cannabis sativa L. Optimized by Response Surface Methodology. J. Food Sci. 2018, 83, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Ilan, A.B.; Gevins, A.; Coleman, M.; ElSohly, M.A.; De Wit, H. Neurophysiological and subjective profile of marijuana with varying concentrations of cannabinoids: Behav. Pharmacol. 2005, 16, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Volicer, L.; Stelly, M.; Morris, J.; McLaughlin, J.; Volicer, B.J. Effects of dronabinol on anorexia and disturbed behavior in patients with Alzheimer’s disease. Int. J. Geriatr. Psychiatry 1997, 12, 913–919. [Google Scholar] [CrossRef]
- Walther, S.; Mahlberg, R.; Eichmann, U.; Kunz, D. Delta-9-tetrahydrocannabinol for nighttime agitation in severe dementia. Psychopharmacology 2006, 185, 524–528. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Alvarez, A.; Pérez, C.A.; Moreno, R.D.; Vicente, M.; Linker, C.; Casanueva, O.I.; Soto, C.; Garrido, J. Acetylcholinesterase Accelerates Assembly of Amyloid-β-Peptides into Alzheimer’s Fibrils: Possible Role of the Peripheral Site of the Enzyme. Neuron 1996, 16, 881–891. [Google Scholar] [CrossRef]
- Eubanks, L.M.; Rogers, C.J.; Beuscher; Koob, G.F.; Olson, A.J.; Dickerson, T.J.; Janda, K.D. A Molecular Link between the Active Component of Marijuana and Alzheimer’s Disease Pathology. Mol. Pharm. 2006, 3, 773–777. [Google Scholar] [CrossRef]
- Ehrhart, J.; Obregon, D.; Mori, T.; Hou, H.; Sun, N.; Bai, Y.; Klein, T.; Fernandez, F.; Tan, J.; Shytle, R.D. Stimulation of cannabinoid receptor 2 (CB2) suppresses microglial activation. J. Neuroinflamm. 2005, 2, 29. [Google Scholar] [CrossRef]
- Facchinetti, F.; Del Giudice, E.; Furegato, S.; Passarotto, M.; Leon, A. Cannabinoids ablate release of TNF? in rat microglial cells stimulated with lypopolysaccharide. Glia 2003, 41, 161–168. [Google Scholar] [CrossRef]
- Stella, N.; Schweitzer, P.; Piomelli, D. A second endogenous cannabinoid that modulates long-term potentiation. Nature 1997, 388, 773–778. [Google Scholar] [CrossRef]
- Marsicano, G.; Goodenough, S.; Monory, K.; Hermann, H.; Eder, M.; Cannich, A.; Azad, S.C.; Cascio, M.G.; Gutiérrez, S.O.; Van Der Stelt, M.; et al. CB1 Cannabinoid Receptors and On-Demand Defense Against Excitotoxicity. Science 2003, 302, 84–88. [Google Scholar] [CrossRef]
- Van Der Stelt, M.; Mazzola, C.; Esposito, G.; Matias, I.; Petrosino, S.; Filippis, D.D.; Micale, V.; Steardo, L.; Drago, F.; Iuvone, T.; et al. Endocannabinoids and β-amyloid-induced neurotoxicity in vivo: Effect of pharmacological elevation of endocannabinoid levels. Cell. Mol. Life Sci. 2006, 63, 1410–1424. [Google Scholar] [CrossRef] [PubMed]
- Koppel, J.; Vingtdeux, V.; Marambaud, P.; d’Abramo, C.; Jimenez, H.; Stauber, M.; Friedman, R.; Davies, P. CB2 Receptor Deficiency Increases Amyloid Pathology and Alters Tau Processing in a Transgenic Mouse Model of Alzheimer’s Disease. Mol. Med. 2014, 20, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Aso, E.; Sánchez-Pla, A.; Vegas-Lozano, E.; Maldonado, R.; Ferrer, I. Cannabis-Based Medicine Reduces Multiple Pathological Processes in AβPP/PS1 Mice. J. Alzheimer’s Dis. 2014, 43, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Aso, E.; Andrés-Benito, P.; Ferrer, I. Delineating the Efficacy of a Cannabis-Based Medicine at Advanced Stages of Dementia in a Murine Model. J. Alzheimer’s Dis. 2016, 54, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Marsicano, G.; Lafenêtre, P. Roles of the Endocannabinoid System in Learning and Memory. In Behavioral Neurobiology of the Endocannabinoid System; Kendall, D., Alexander, S., Eds.; Current Topics in Behavioral Neurosciences; Springer: Berlin/Heidelberg, Germany, 2009; Volume 1, pp. 201–230. ISBN 978-3-540-88954-0. [Google Scholar] [CrossRef]
- Butovsky, E.; Juknat, A.; Goncharov, I.; Elbaz, J.; Eilam, R.; Zangen, A.; Vogel, Z. In vivo up-regulation of brain-derived neurotrophic factor in specific brain areas by chronic exposure to Delta9-tetrahydrocannabinol. J. Neurochem. 2005, 93, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Schubert, D.; Kepchia, D.; Liang, Z.; Dargusch, R.; Goldberg, J.; Maher, P. Efficacy of Cannabinoids in a Pre-Clinical Drug-Screening Platform for Alzheimer’s Disease. Mol. Neurobiol. 2019, 56, 7719–7730. [Google Scholar] [CrossRef]
- Ahmed, A.; Van Der Marck, M.; Van Den Elsen, G.; Olde Rikkert, M. Cannabinoids in late-onset Alzheimer’s disease. Clin. Pharmacol. Ther. 2015, 97, 597–606. [Google Scholar] [CrossRef]
- Carroll, C.B.; Bain, P.G.; Teare, L.; Liu, X.; Joint, C.; Wroath, C.; Parkin, S.G.; Fox, P.; Wright, D.; Hobart, J.; et al. Cannabis for dyskinesia in Parkinson disease: A randomized double-blind crossover study. Neurology 2004, 63, 1245–1250. [Google Scholar] [CrossRef]
- Kim, S.H.; Won, S.J.; Mao, X.O.; Jin, K.; Greenberg, D.A. Molecular Mechanisms of Cannabinoid Protection from Neuronal Excitotoxicity. Mol. Pharmacol. 2006, 69, 691–696. [Google Scholar] [CrossRef]
- Pérez-Olives, C.; Rivas-Santisteban, R.; Lillo, J.; Navarro, G.; Franco, R. Recent Advances in the Potential of Cannabinoids for Neuroprotection in Alzheimer’s, Parkinson’s, and Huntington’s Diseases. In Cannabinoids and Neuropsychiatric Disorders; Murillo-Rodriguez, E., Pandi-Perumal, S.R., Monti, J.M., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2021; Volume 1264, pp. 81–92. ISBN 978-3-030-57368-3. [Google Scholar] [CrossRef]
- Garcia-Arencibia, M.; Garcia, C.; Fernandez-Ruiz, J. Cannabinoids and Parkinsons Disease. CNS Neurol. Disord.-Drug Targets 2009, 8, 432–439. [Google Scholar] [CrossRef]
- Brotchie, J.M. Nondopaminergic mechanisms in levodopa-induced dyskinesia. Mov. Disord. 2005, 20, 919–931. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, B.; Asbrock, N.; Kathuria, S.; Piomelli, D.; Giuffrida, A. Effects of levodopa on endocannabinoid levels in rat basal ganglia: Implications for the treatment of levodopa-induced dyskinesias. Eur. J. Neurosci. 2003, 18, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, V.; Houeto, J.L.; Bonnet, A.M.; Clavier, I.; Arnulf, I.; Cattelin, F.; Le Fur, G.; Damier, P.; Welter, M.L.; Agid, Y. Neurokinin B, Neurotensin, and Cannabinoid Receptor Antagonists and Parkinson Disease: Clin. Neuropharmacol. 2004, 27, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Chagas, M.H.N.; Zuardi, A.W.; Tumas, V.; Pena-Pereira, M.A.; Sobreira, E.T.; Bergamaschi, M.M.; Dos Santos, A.C.; Teixeira, A.L.; Hallak, J.E.; Crippa, J.A.S. Effects of cannabidiol in the treatment of patients with Parkinson’s disease: An exploratory double-blind trial. J. Psychopharmacol. 2014, 28, 1088–1098. [Google Scholar] [CrossRef] [PubMed]
- Sieradzan, K.A.; Fox, S.H.; Hill, M.; Dick, J.P.R.; Crossman, A.R.; Brotchie, J.M. Cannabinoids reduce levodopa-induced dyskinesia in Parkinson’s disease: A pilot study. Neurology 2001, 57, 2108–2111. [Google Scholar] [CrossRef]
- Lastres-Becker, I.; Molina-Holgado, F.; Ramos, J.A.; Mechoulam, R.; Fernández-Ruiz, J. Cannabinoids provide neuroprotection against 6-hydroxydopamine toxicity in vivo and in vitro: Relevance to Parkinson’s disease. Neurobiol. Dis. 2005, 19, 96–107. [Google Scholar] [CrossRef]
- Klegeris, A.; Bissonnette, C.J.; McGeer, P.L. Reduction of human monocytic cell neurotoxicity and cytokine secretion by ligands of the cannabinoid-type CB2 receptor: Antineurotoxic action of cannabinoids. Br. J. Pharmacol. 2003, 139, 775–786. [Google Scholar] [CrossRef]
- Martín-Moreno, A.M.; Reigada, D.; Ramírez, B.G.; Mechoulam, R.; Innamorato, N.; Cuadrado, A.; De Ceballos, M.L. Cannabidiol and Other Cannabinoids Reduce Microglial Activation In Vitro and In Vivo: Relevance to Alzheimer’s Disease. Mol. Pharmacol. 2011, 79, 964–973. [Google Scholar] [CrossRef]
- López-Sendón Moreno, J.L.; García Caldentey, J.; Trigo Cubillo, P.; Ruiz Romero, C.; García Ribas, G.; Alonso Arias, M.A.A.; García De Yébenes, M.J.; Tolón, R.M.; Galve-Roperh, I.; Sagredo, O.; et al. A double-blind, randomized, cross-over, placebo-controlled, pilot trial with Sativex in Huntington’s disease. J. Neurol. 2016, 263, 1390–1400. [Google Scholar] [CrossRef]
- Russo, E.; Guy, G.W. A tale of two cannabinoids: The therapeutic rationale for combining tetrahydrocannabinol and cannabidiol. Med. Hypotheses 2006, 66, 234–246. [Google Scholar] [CrossRef]
- Palazuelos, J.; Aguado, T.; Pazos, M.R.; Julien, B.; Carrasco, C.; Resel, E.; Sagredo, O.; Benito, C.; Romero, J.; Azcoitia, I.; et al. Microglial CB2 cannabinoid receptors are neuroprotective in Huntington’s disease excitotoxicity. Brain 2009, 132, 3152–3164. [Google Scholar] [CrossRef] [PubMed]
- Saft, C.; Von Hein, S.M.; Lücke, T.; Thiels, C.; Peball, M.; Djamshidian, A.; Heim, B.; Seppi, K. Cannabinoids for Treatment of Dystonia in Huntington’s Disease. J. Huntington’s Dis. 2018, 7, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Dowie, M.J.; Howard, M.L.; Nicholson, L.F.B.; Faull, R.L.M.; Hannan, A.J.; Glass, M. Behavioural and molecular consequences of chronic cannabinoid treatment in Huntington’s disease transgenic mice. Neuroscience 2010, 170, 324–336. [Google Scholar] [CrossRef]
- Farkas, S.; Nagy, K.; Palkovits, M.; Kovács, G.G.; Jia, Z.; Donohue, S.; Pike, V.; Halldin, C.; Máthé, D.; Harkany, T.; et al. [125I]SD-7015 reveals fine modalities of CB1 cannabinoid receptor density in the prefrontal cortex during progression of Alzheimer’s disease. Neurochem. Int. 2012, 60, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, B.G.; Blázquez, C.; Del Pulgar, T.G.; Guzmán, M.; De Ceballos, M.L. Prevention of Alzheimer’s Disease Pathology by Cannabinoids: Neuroprotection Mediated by Blockade of Microglial Activation. J. Neurosci. 2005, 25, 1904–1913. [Google Scholar] [CrossRef] [PubMed]
- Solas, M.; Francis, P.T.; Franco, R.; Ramirez, M.J. CB2 receptor and amyloid pathology in frontal cortex of Alzheimer’s disease patients. Neurobiol. Aging 2013, 34, 805–808. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.-M.; Sepers, M.; Henstridge, C.M.; Lassalle, O.; Neuhofer, D.; Martin, H.; Ginger, M.; Frick, A.; DiPatrizio, N.V.; Mackie, K.; et al. Uncoupling of the endocannabinoid signalling complex in a mouse model of fragile X syndrome. Nat. Commun. 2012, 3, 1080. [Google Scholar] [CrossRef]
- Mulder, J.; Zilberter, M.; Pasquaré, S.J.; Alpár, A.; Schulte, G.; Ferreira, S.G.; Köfalvi, A.; Martín-Moreno, A.M.; Keimpema, E.; Tanila, H.; et al. Molecular reorganization of endocannabinoid signalling in Alzheimer’s disease. Brain 2011, 134, 1041–1060. [Google Scholar] [CrossRef]
- Bisogno, T. Endogenous Cannabinoids: Structure and Metabolism. J. Neuroendocr. 2008, 20, 1–9. [Google Scholar] [CrossRef]
- Van Laere, K.; Casteels, C.; Lunskens, S.; Goffin, K.; Grachev, I.D.; Bormans, G.; Vandenberghe, W. Regional changes in type 1 cannabinoid receptor availability in Parkinson’s disease in vivo. Neurobiol. Aging 2012, 33, 620.e1–620.e8. [Google Scholar] [CrossRef]
- Pisani, V.; Madeo, G.; Tassone, A.; Sciamanna, G.; Maccarrone, M.; Stanzione, P.; Pisani, A. Homeostatic changes of the endocannabinoid system in Parkinson’s disease: eCB Changes in PD. Mov. Disord. 2011, 26, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Glass, M.; Dragunow, M.; Faull, R.L.M. The pattern of neurodegeneration in Huntington’s disease: A comparative study of cannabinoid, dopamine, adenosine and GABAA receptor alterations in the human basal ganglia in Huntington’s disease. Neuroscience 2000, 97, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R. The Pharmacohistory of Cannabis Sativa. In Cannabinoids as Therapeutic Agents, 1st ed.; Mechoulam, R., Ed.; Chapman and Hall/CRC: Boca Raton, FL, USA, 2019; pp. 1–20. ISBN 978-0-429-26066-7. [Google Scholar]
- Zajicek, J.P.; Apostu, V.I. Role of Cannabinoids in Multiple Sclerosis. CNS Drugs 2011, 25, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G. Pharmacology of cannabinoid CB1 and CB2 receptors. Pharmacol. Ther. 1997, 74, 129–180. [Google Scholar] [CrossRef] [PubMed]
- Zajicek, J.; Ball, S.; Wright, D.; Vickery, J.; Nunn, A.; Miller, D.; Cano, M.G.; McManus, D.; Mallik, S.; Hobart, J. Effect of dronabinol on progression in progressive multiple sclerosis (CUPID): A randomised, placebo-controlled trial. Lancet Neurol. 2013, 12, 857–865. [Google Scholar] [CrossRef]
- Arévalo-Martín, Á.; Vela, J.M.; Molina-Holgado, E.; Borrell, J.; Guaza, C. Therapeutic Action of Cannabinoids in a Murine Model of Multiple Sclerosis. J. Neurosci. 2003, 23, 2511–2516. [Google Scholar] [CrossRef]
- Panikashvili, D.; Simeonidou, C.; Ben-Shabat, S.; Hanuš, L.; Breuer, A.; Mechoulam, R.; Shohami, E. An endogenous cannabinoid (2-AG) is neuroprotective after brain injury. Nature 2001, 413, 527–531. [Google Scholar] [CrossRef]
- Franklin, A.; Parmentier-Batteur, S.; Walter, L.; Greenberg, D.A.; Stella, N. Palmitoylethanolamide Increases after Focal Cerebral Ischemia and Potentiates Microglial Cell Motility. J. Neurosci. 2003, 23, 7767–7775. [Google Scholar] [CrossRef]
- Muthian, S.; Rademacher, D.J.; Roelke, C.T.; Gross, G.J.; Hillard, C.J. Anandamide content is increased and CB1 cannabinoid receptor blockade is protective during transient, focal cerebral ischemia. Neuroscience 2004, 129, 743–750. [Google Scholar] [CrossRef]
- Hetz, C.; Saxena, S. ER stress and the unfolded protein response in neurodegeneration. Nat. Rev. Neurol. 2017, 13, 477–491. [Google Scholar] [CrossRef]
- Fernández-Ruiz, J.; Moro, M.A.; Martínez-Orgado, J. Cannabinoids in Neurodegenerative Disorders and Stroke/Brain Trauma: From Preclinical Models to Clinical Applications. Neurotherapeutics 2015, 12, 793–806. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.A.; Wang, Z.H.; Zhang, R.; Piao, M.J.; Kim, K.C.; Kang, S.S.; Kim, Y.W.; Lee, J.; Park, D.; Hyun, J.W. Myricetin Protects Cells against Oxidative Stress-Induced Apoptosis via Regulation of PI3K/Akt and MAPK Signaling Pathways. Int. J. Mol. Sci. 2010, 11, 4348–4360. [Google Scholar] [CrossRef] [PubMed]
- Jazvinšćak Jembrek, M.; Radovanović, V.; Vlainić, J.; Vuković, L.; Hanžić, N. Neuroprotective effect of zolpidem against glutamate-induced toxicity is mediated via the PI3K/Akt pathway and inhibited by PK11195. Toxicology 2018, 406–407, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Jazvinšćak Jembrek, M.; Vlainić, J.; Čadež, V.; Šegota, S. Atomic force microscopy reveals new biophysical markers for monitoring subcellular changes in oxidative injury: Neuroprotective effects of quercetin at the nanoscale. PLoS ONE 2018, 13, e0200119. [Google Scholar] [CrossRef]
- Zakharova, I.O.; Sokolova, T.V.; Bayunova, L.V.; Zorina, I.I.; Rychkova, M.P.; Shpakov, A.O.; Avrova, N.F. The Protective Effect of Insulin on Rat Cortical Neurons in Oxidative Stress and Its Dependence on the Modulation of Akt, GSK-3beta, ERK1/2, and AMPK Activities. Int. J. Mol. Sci. 2019, 20, 3702. [Google Scholar] [CrossRef]
- Sánchez-Blázquez, P.; Rodríguez-Muñoz, M.; Vicente-Sánchez, A.; Garzón, J. Cannabinoid Receptors Couple to NMDA Receptors to Reduce the Production of NO and the Mobilization of Zinc Induced by Glutamate. Antioxid. Redox Signal. 2013, 19, 1766–1782. [Google Scholar] [CrossRef]
- Rocha, F.C.M.; Dos Santos Júnior, J.G.; Stefano, S.C.; Da Silveira, D.X. Systematic review of the literature on clinical and experimental trials on the antitumor effects of cannabinoids in gliomas. J. Neurooncol. 2014, 116, 11–24. [Google Scholar] [CrossRef]
- Massi, P.; Vaccani, A.; Ceruti, S.; Colombo, A.; Abbracchio, M.P.; Parolaro, D. Antitumor Effects of Cannabidiol, a Nonpsychoactive Cannabinoid, on Human Glioma Cell Lines. J. Pharmacol. Exp. Ther. 2004, 308, 838–845. [Google Scholar] [CrossRef]
- Becker, W.; Alrafas, H.R.; Wilson, K.; Miranda, K.; Culpepper, C.; Chatzistamou, I.; Cai, G.; Nagarkatti, M.; Nagarkatti, P.S. Activation of Cannabinoid Receptor 2 Prevents Colitis-Associated Colon Cancer through Myeloid Cell De-activation Upstream of IL-22 Production. iScience 2020, 23, 101504. [Google Scholar] [CrossRef]
- Silva-Reis, R.; Silva, A.M.S.; Oliveira, P.A.; Cardoso, S.M. Antitumor Effects of Cannabis sativa Bioactive Compounds on Colorectal Carcinogenesis. Biomolecules 2023, 13, 764. [Google Scholar] [CrossRef]
- Sánchez-Sánchez, L.; García, J.; Fernández, R.; Noskova, E.; Egiguren-Ortiz, J.; Gulak, M.; Ochoa, E.; Laso, A.; Oiarbide, M.; Santos, J.I.; et al. Characterization of the Antitumor Potential of Extracts of Cannabis sativa Strains with High CBD Content in Human Neuroblastoma. Int. J. Mol. Sci. 2023, 24, 3837. [Google Scholar] [CrossRef]
- Ligresti, A.; Moriello, A.S.; Starowicz, K.; Matias, I.; Pisanti, S.; De Petrocellis, L.; Laezza, C.; Portella, G.; Bifulco, M.; Di Marzo, V. Antitumor Activity of Plant Cannabinoids with Emphasis on the Effect of Cannabidiol on Human Breast Carcinoma. J. Pharmacol. Exp. Ther. 2006, 318, 1375–1387. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.S.; Marie, M.A.; Sheweita, S.A. Novel mechanism of cannabidiol-induced apoptosis in breast cancer cell lines. Breast 2018, 41, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Leyva-Illades, D.; DeMorrow, S. Orphan G protein receptor GPR55 as an emerging target in cancer therapy and management. Cancer Manag. Res. 2013, 5, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Niphakis, M.J.; Cognetta, A.B.; Chang, J.W.; Buczynski, M.W.; Parsons, L.H.; Byrne, F.; Burston, J.J.; Chapman, V.; Cravatt, B.F. Evaluation of NHS Carbamates as a Potent and Selective Class of Endocannabinoid Hydrolase Inhibitors. ACS Chem. Neurosci. 2013, 4, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Curry, Z.A.; Wilkerson, J.L.; Bagdas, D.; Kyte, S.L.; Patel, N.; Donvito, G.; Mustafa, M.A.; Poklis, J.L.; Niphakis, M.J.; Hsu, K.-L.; et al. Monoacylglycerol Lipase Inhibitors Reverse Paclitaxel-Induced Nociceptive Behavior and Proinflammatory Markers in a Mouse Model of Chemotherapy-Induced Neuropathy. J. Pharmacol. Exp. Ther. 2018, 366, 169–183. [Google Scholar] [CrossRef]
- Seltzer, E.S.; Watters, A.K.; MacKenzie, D.; Granat, L.M.; Zhang, D. Cannabidiol (CBD) as a Promising Anti-Cancer Drug. Cancers 2020, 12, 3203. [Google Scholar] [CrossRef]
- Pagano, E.; Borrelli, F.; Orlando, P.; Romano, B.; Monti, M.; Morbidelli, L.; Aviello, G.; Imperatore, R.; Capasso, R.; Piscitelli, F.; et al. Pharmacological inhibition of MAGL attenuates experimental colon carcinogenesis. Pharmacol. Res. 2017, 119, 227–236. [Google Scholar] [CrossRef]
- Mücke, M.; Weier, M.; Carter, C.; Copeland, J.; Degenhardt, L.; Cuhls, H.; Radbruch, L.; Häuser, W.; Conrad, R. Systematic review and meta-analysis of cannabinoids in palliative medicine: Cannabinoids in palliative medicine. J. Cachexia Sarcopenia Muscle 2018, 9, 220–234. [Google Scholar] [CrossRef]
- Noskova, E.; Fernández, R.; García, J.; Ochoa, E.; Domínguez-Fernández, C.; Adell, A.; Laso, A.; Andrés, M.F.; González-Coloma, A.; Astigarraga, E.; et al. Screening System of Cannabis sativa Extracts Based on Their Mitochondrial Safety Profile Using Cytochrome c Oxidase Activity as a Biomarker. Int. J. Mol. Sci. 2023, 24, 1315. [Google Scholar] [CrossRef]
- Harkany, T.; Horvath, T.L. (S)Pot on Mitochondria: Cannabinoids Disrupt Cellular Respiration to Limit Neuronal Activity. Cell Metab. 2017, 25, 8–10. [Google Scholar] [CrossRef] [PubMed]
- Fišar, Z.; Singh, N.; Hroudová, J. Cannabinoid-induced changes in respiration of brain mitochondria. Toxicol. Lett. 2014, 231, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Lim, S.; Ryu, J.; Lee, C.-W.; Kim, Y.; Kang, J.-H.; Kang, S.-S.; Ahn, Y.K.; Park, C.-S.; Kim, J.J. CB1 and CB2 cannabinoid receptors differentially regulate the production of reactive oxygen species by macrophages. Cardiovasc. Res. 2009, 84, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Liang, Z.; Mi, Q.; Guo, Y. Limonene terpenoid obstructs human bladder cancer cell (T24 cell line) growth by inducing cellular apoptosis, caspase activation, G2/M phase cell cycle arrest and stops cancer metastasis. J. BUON Off. J. Balk. Union Oncol. 2020, 25, 280–285. [Google Scholar]
- Hafidh, R.R.; Hussein, S.Z.; MalAllah, M.Q.; Abdulamir, A.S.; Abu Bakar, F. A High-throughput Quantitative Expression Analysis of Cancer-related Genes in Human HepG2 Cells in Response to Limonene, a Potential Anticancer Agent. Curr. Cancer Drug Targets 2018, 18, 807–815. [Google Scholar] [CrossRef]
- Zhang, X.-Z.; Wang, L.; Liu, D.-W.; Tang, G.-Y.; Zhang, H.-Y. Synergistic Inhibitory Effect of Berberine and d-Limonene on Human Gastric Carcinoma Cell Line MGC803. J. Med. Food 2014, 17, 955–962. [Google Scholar] [CrossRef]
- Russo, R.; Cassiano, M.G.V.; Ciociaro, A.; Adornetto, A.; Varano, G.P.; Chiappini, C.; Berliocchi, L.; Tassorelli, C.; Bagetta, G.; Corasaniti, M.T. Role of D-Limonene in Autophagy Induced by Bergamot Essential Oil in SH-SY5Y Neuroblastoma Cells. PLoS ONE 2014, 9, e113682. [Google Scholar] [CrossRef]
- Yu, X.; Lin, H.; Wang, Y.; Lv, W.; Zhang, S.; Qian, Y.; Deng, X.; Feng, N.; Yu, H.; Qian, B. D-limonene exhibits antitumor activity by inducing autophagy and apoptosis in lung cancer. OncoTargets Ther. 2018, 11, 1833–1847. [Google Scholar] [CrossRef]
- Del Toro-Arreola, S.; Flores-Torales, E.; Torres-Lozano, C.; Del Toro-Arreola, A.; Tostado-Pelayo, K.; Guadalupe Ramirez-Dueñas, M.; Daneri-Navarro, A. Effect of d-limonene on immune response in BALB/c mice with lymphoma. Int. Immunopharmacol. 2005, 5, 829–838. [Google Scholar] [CrossRef]
- Manassero, C.A.; Girotti, J.R.; Mijailovsky, S.; García De Bravo, M.; Polo, M. In vitro comparative analysis of antiproliferative activity of essential oil from mandarin peel and its principal component limonene. Nat. Prod. Res. 2013, 27, 1475–1478. [Google Scholar] [CrossRef]
- Colvin, E.K.; Hudson, A.L.; Anderson, L.L.; Kumar, R.P.; McGregor, I.S.; Howell, V.M.; Arnold, J.C. An Examination of the Anti-Cancer Properties of Plant Cannabinoids in Preclinical Models of Mesothelioma. Cancers 2022, 14, 3813. [Google Scholar] [CrossRef]
- Lauckner, J.E.; Jensen, J.B.; Chen, H.-Y.; Lu, H.-C.; Hille, B.; Mackie, K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc. Natl. Acad. Sci. USA 2008, 105, 2699–2704. [Google Scholar] [CrossRef] [PubMed]
- Kurosawa, M.; Arakaki, R.; Yamada, A.; Tsunematsu, T.; Kudo, Y.; Sprent, J.; Ishimaru, N. NF-κB2 Controls the Migratory Activity of Memory T Cells by Regulating Expression of CXCR4 in a Mouse Model of Sjögren’s Syndrome. Arthritis Rheumatol. 2017, 69, 2193–2202. [Google Scholar] [CrossRef] [PubMed]
- Notaro, A.; Sabella, S.; Pellerito, O.; Di Fiore, R.; De Blasio, A.; Vento, R.; Calvaruso, G.; Giuliano, M. Involvement of PAR-4 in Cannabinoid-Dependent Sensitization of Osteosarcoma Cells to TRAIL-Induced Apoptosis. Int. J. Biol. Sci. 2014, 10, 466–478. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Tiedra, S.; Fabriàs, G.; Dávila, D.; Salanueva, Í.J.; Casas, J.; Montes, L.R.; Antón, Z.; García-Taboada, E.; Salazar-Roa, M.; Lorente, M.; et al. Dihydroceramide accumulation mediates cytotoxic autophagy of cancer cells via autolysosome destabilization. Autophagy 2016, 12, 2213–2229. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.; Carracedo, A.; Salanueva, Í.J.; Hernández-Tiedra, S.; Lorente, M.; Egia, A.; Vázquez, P.; Blázquez, C.; Torres, S.; García, S.; et al. Cannabinoid action induces autophagy-mediated cell death through stimulation of ER stress in human glioma cells. J. Clin. Investig. 2009, 119, 1359–1372. [Google Scholar] [CrossRef]
- Fisher, T.; Golan, H.; Schiby, G.; PriChen, S.; Smoum, R.; Moshe, I.; Peshes-Yaloz, N.; Castiel, A.; Waldman, D.; Gallily, R.; et al. In Vitro and In Vivo Efficacy of Non-Psychoactive Cannabidiol in Neuroblastoma. Curr. Oncol. 2016, 23, 15–22. [Google Scholar] [CrossRef]
- Borrell, B. How accurate are cancer cell lines? Nature 2010, 463, 858. [Google Scholar] [CrossRef]
- Sredni, S.T.; Huang, C.-C.; Suzuki, M.; Pundy, T.; Chou, P.; Tomita, T. Spontaneous involution of pediatric low-grade gliomas: High expression of cannabinoid receptor 1 (CNR1) at the time of diagnosis may indicate involvement of the endocannabinoid system. Childs Nerv. Syst. 2016, 32, 2061–2067. [Google Scholar] [CrossRef]
- Foroughi, M.; Hendson, G.; Sargent, M.A.; Steinbok, P. Spontaneous regression of septum pellucidum/forniceal pilocytic astrocytomas—Possible role of Cannabis inhalation. Childs Nerv. Syst. 2011, 27, 671–679. [Google Scholar] [CrossRef]
- Blasco-Benito, S.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; García-Taboada, E.; Wade, J.; Smith, S.; Guzmán, M.; Pérez-Gómez, E.; et al. Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in preclinical models of breast cancer. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef]
- LaVigne, J.; Hecksel, R.; Streicher, J.M. In Defense of the “Entourage Effect”: Terpenes Found in Cannabis sativa Activate the Cannabinoid Receptor 1 In Vivo. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- Nithipatikom, K.; Gomez-Granados, A.D.; Tang, A.T.; Pfeiffer, A.W.; Williams, C.L.; Campbell, W.B. Cannabinoid Receptor Type 1 (CB1) Activation Inhibits Small GTPase RhoA Activity and Regulates Motility of Prostate Carcinoma Cells. Endocrinology 2012, 153, 29–41. [Google Scholar] [CrossRef]
- Roberto, D.; Klotz, L.H.; Venkateswaran, V. Cannabinoid WIN 55,212-2 induces cell cycle arrest and apoptosis, and inhibits proliferation, migration, invasion, and tumor growth in prostate cancer in a cannabinoid-receptor 2 dependent manner. Prostate 2019, 79, 151–159. [Google Scholar] [CrossRef]
- Allister, S.D.M.; Chan, C.; Taft, R.J.; Luu, T.; Abood, M.E.; Moore, D.H.; Aldape, K.; Yount, G. Cannabinoids selectively inhibit proliferation and induce death of cultured human glioblastoma multiforme cells. J. Neurooncol. 2005, 74, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Doherty, G.J.; De Paula, B.H.R. Cannabinoids in glioblastoma multiforme—Hype or hope? Br. J. Cancer 2021, 124, 1341–1343. [Google Scholar] [CrossRef] [PubMed]
- Lah, T.T.; Novak, M.; Pena Almidon, M.A.; Marinelli, O.; Žvar Baškovič, B.; Majc, B.; Mlinar, M.; Bošnjak, R.; Breznik, B.; Zomer, R.; et al. Cannabigerol Is a Potential Therapeutic Agent in a Novel Combined Therapy for Glioblastoma. Cells 2021, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Stachtari, C.C.; Thomareis, O.N.; Tsaousi, G.G.; Karakoulas, K.A.; Chatzimanoli, F.I.; Chatzopoulos, S.A.; Vasilakos, D.G. Interaction of a Cannabinoid-2 Agonist with Tramadol on Nociceptive Thresholds and Immune Responses in a Rat Model of Incisional Pain. Am. J. Ther. 2016, 23, e1484–e1492. [Google Scholar] [CrossRef] [PubMed]
- Gardin, A.; Kucher, K.; Kiese, B.; Appel-Dingemanse, S. Cannabinoid Receptor Agonist 13, a Novel Cannabinoid Agonist: First in Human Pharmacokinetics and Safety. Drug Metab. Dispos. 2009, 37, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, J.; Annas, P.; Huizar, K.; Clarke, C.; Zettergren, A.; Karlsten, R.; Segerdahl, M. Evaluation of the analgesic efficacy and psychoactive effects of AZD1940, a novel peripherally acting cannabinoid agonist, in human capsaicin-induced pain and hyperalgesia. Clin. Exp. Pharmacol. Physiol. 2013, 40, 212–218. [Google Scholar] [CrossRef]
- Banister, S.D.; Krishna Kumar, K.; Kumar, V.; Kobilka, B.K.; Malhotra, S.V. Selective modulation of the cannabinoid type 1 (CB1) receptor as an emerging platform for the treatment of neuropathic pain. Med. Chem. Commun. 2019, 10, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, J.; Segerdahl, M.; Webster, L.; Reimfelt, A.; Huizar, K.; Annas, P.; Karlsten, R.; Quiding, H. Evaluation of the analgesic efficacy of AZD1940, a novel cannabinoid agonist, on post-operative pain after lower third molar surgical removal. Scand. J. Pain 2013, 4, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Bradford, H.F. Glutamate, GABA and epilepsy. Prog. Neurobiol. 1995, 47, 477–511. [Google Scholar] [CrossRef] [PubMed]
- Karler, R.; Turkanis, S.A. The Cannabinoids as Potential Antiepileptics. J. Clin. Pharmacol. 1981, 21, 437S–448S. [Google Scholar] [CrossRef]
- Iannotti, F.A.; Hill, C.L.; Leo, A.; Alhusaini, A.; Soubrane, C.; Mazzarella, E.; Russo, E.; Whalley, B.J.; Di Marzo, V.; Stephens, G.J. Nonpsychotropic Plant Cannabinoids, Cannabidivarin (CBDV) and Cannabidiol (CBD), Activate and Desensitize Transient Receptor Potential Vanilloid 1 (TRPV1) Channels in Vitro: Potential for the Treatment of Neuronal Hyperexcitability. ACS Chem. Neurosci. 2014, 5, 1131–1141. [Google Scholar] [CrossRef]
- Kollipara, R.; Langille, E.; Tobin, C.; French, C.R. CBD can be combined with additional cannabinoids for optimal seizure reduction and requires GPR55 for its anticonvulsant effects. bioRxiv 2023. [Google Scholar] [CrossRef]
- Goerl, B.; Watkins, S.; Metcalf, C.; Smith, M.; Beenhakker, M. Cannabidiolic acid exhibits entourage-like improvements of anticonvulsant activity in an acute rat model of seizures. Epilepsy Res. 2021, 169, 106525. [Google Scholar] [CrossRef]
- Ryan, D.; Drysdale, A.J.; Lafourcade, C.; Pertwee, R.G.; Platt, B. Cannabidiol Targets Mitochondria to Regulate Intracellular Ca2+ Levels. J. Neurosci. 2009, 29, 2053–2063. [Google Scholar] [CrossRef]
- Chaves, Y.C.; Genaro, K.; Crippa, J.A.; Da Cunha, J.M.; Zanoveli, J.M. Cannabidiol induces antidepressant and anxiolytic-like effects in experimental type-1 diabetic animals by multiple sites of action. Metab. Brain Dis. 2021, 36, 639–652. [Google Scholar] [CrossRef]
- Lucas, P.; Walsh, Z.; Crosby, K.; Callaway, R.; Belle-Isle, L.; Kay, R.; Capler, R.; Holtzman, S. Substituting cannabis for prescription drugs, alcohol and other substances among medical cannabis patients: The impact of contextual factors: Cannabis substitution. Drug Alcohol. Rev. 2016, 35, 326–333. [Google Scholar] [CrossRef]
- Kostanda, E.; Khatib, S. Biotic stress caused by Tetranychus urticae mites elevates the quantity of secondary metabolites, cannabinoids and terpenes, in Cannabis sativa L. Ind. Crops Prod. 2022, 176, 114331. [Google Scholar] [CrossRef]
- Loss, C.M.; Teodoro, L.; Rodrigues, G.D.; Moreira, L.R.; Peres, F.F.; Zuardi, A.W.; Crippa, J.A.; Hallak, J.E.C.; Abílio, V.C. Is Cannabidiol During Neurodevelopment a Promising Therapy for Schizophrenia and Autism Spectrum Disorders? Front. Pharmacol. 2021, 11, 635763. [Google Scholar] [CrossRef] [PubMed]
- Karoly, H.C.; Milburn, M.A.; Brooks-Russell, A.; Brown, M.; Streufert, J.; Bryan, A.D.; Lovrich, N.P.; DeJong, W.; Cinnamon Bidwell, L. Effects of High-Potency Cannabis on Psychomotor Performance in Frequent Cannabis Users. Cannabis Cannabinoid Res. 2022, 7, 107–115. [Google Scholar] [CrossRef]
- Wallace, A.L.; Wade, N.E.; Lisdahl, K.M. Impact of 2 Weeks of Monitored Abstinence on Cognition in Adolescent and Young Adult Cannabis Users. J. Int. Neuropsychol. Soc. 2020, 26, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Schuster, R.M.; Mermelstein, R.J.; Vassileva, J.; Martin, E.M.; Diviak, K.R. Performance of young adult cannabis users on neurocognitive measures of impulsive behavior and their relationship to symptoms of cannabis use disorders. J. Clin. Exp. Neuropsychol. 2012, 34, 962–976. [Google Scholar] [CrossRef]
- Collin, C.; Ehler, E.; Waberzinek, G.; Alsindi, Z.; Davies, P.; Powell, K.; Notcutt, W.; O’Leary, C.; Ratcliffe, S.; Nováková, I.; et al. A double-blind, randomized, placebo-controlled, parallel-group study of Sativex, in subjects with symptoms of spasticity due to multiple sclerosis. Neurol. Res. 2010, 32, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Markovà, J.; Essner, U.; Akmaz, B.; Marinelli, M.; Trompke, C.; Lentschat, A.; Vila, C. Sativex ® as add-on therapy vs. further optimized first-line ANTispastics (SAVANT) in resistant multiple sclerosis spasticity: A double-blind, placebo-controlled randomised clinical trial. Int. J. Neurosci. 2019, 129, 119–128. [Google Scholar] [CrossRef]
- Meuth, S.G.; Henze, T.; Essner, U.; Trompke, C.; Vila Silván, C. Tetrahydrocannabinol and cannabidiol oromucosal spray in resistant multiple sclerosis spasticity: Consistency of response across subgroups from the SAVANT randomized clinical trial. Int. J. Neurosci. 2020, 130, 1199–1205. [Google Scholar] [CrossRef]
- Wade, D.T.; Collin, C.; Stott, C.; Duncombe, P. Meta-analysis of the efficacy and safety of Sativex (nabiximols), on spasticity in people with multiple sclerosis. Mult. Scler. 2010, 16, 707–714. [Google Scholar] [CrossRef]
- Serpell, M.G.; Notcutt, W.; Collin, C. Sativex long-term use: An open-label trial in patients with spasticity due to multiple sclerosis. J. Neurol. 2013, 260, 285–295. [Google Scholar] [CrossRef]
- Ambrose, T.; Simmons, A. Cannabis, Cannabinoids, and the Endocannabinoid System—Is there Therapeutic Potential for Inflammatory Bowel Disease? J. Crohn’s Colitis 2019, 13, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Kienzl, M.; Storr, M.; Schicho, R. Cannabinoids and Opioids in the Treatment of Inflammatory Bowel Diseases. Clin. Transl. Gastroenterol. 2020, 11, e00120. [Google Scholar] [CrossRef] [PubMed]
- Mboumba Bouassa, R.-S.; Comeau, E.; Alexandrova, Y.; Pagliuzza, A.; Yero, A.; Samarani, S.; Needham, J.; Singer, J.; Lee, T.; Bobeuf, F.; et al. Effects of Oral Cannabinoids on Systemic Inflammation and Viral Reservoir Markers in People with HIV on Antiretroviral Therapy: Results of the CTN PT028 Pilot Clinical Trial. Cells 2023, 12, 1811. [Google Scholar] [CrossRef] [PubMed]
- Couch, D.G.; Cook, H.; Ortori, C.; Barrett, D.; Lund, J.N.; O’Sullivan, S.E. Palmitoylethanolamide and Cannabidiol Prevent Inflammation-induced Hyperpermeability of the Human Gut In Vitro and In Vivo—A Randomized, Placebo-controlled, Double-blind Controlled Trial. Inflamm. Bowel Dis. 2019, 25, 1006–1018. [Google Scholar] [CrossRef]
- Mboumba Bouassa, R.-S.; Needham, J.; Nohynek, D.; Singer, J.; Lee, T.; Bobeuf, F.; Samarani, S.; Del Balso, L.; Paisible, N.; Vertzagias, C.; et al. Safety and Tolerability of Oral Cannabinoids in People Living with HIV on Long-Term ART: A Randomized, Open-Label, Interventional Pilot Clinical Trial (CTNPT 028). Biomedicines 2022, 10, 3168. [Google Scholar] [CrossRef]
- Whiting, P.F.; Wolff, R.F.; Deshpande, S.; Di Nisio, M.; Duffy, S.; Hernandez, A.V.; Keurentjes, J.C.; Lang, S.; Misso, K.; Ryder, S.; et al. Cannabinoids for Medical Use: A Systematic Review and Meta-analysis. JAMA 2015, 313, 2456. [Google Scholar] [CrossRef]
- Stuyt, E. The Problem with the Current High Potency THC Marijuana from the Perspective of an Addiction Psychiatrist. Mo. Med. 2018, 115, 482–486. [Google Scholar]
- Tramer, M.R. Cannabinoids for control of chemotherapy induced nausea and vomiting: Quantitative systematic. BMJ 2001, 323, 16. [Google Scholar] [CrossRef]
- Gobbi, G.; Atkin, T.; Zytynski, T.; Wang, S.; Askari, S.; Boruff, J.; Ware, M.; Marmorstein, N.; Cipriani, A.; Dendukuri, N.; et al. Association of Cannabis Use in Adolescence and Risk of Depression, Anxiety, and Suicidality in Young Adulthood: A Systematic Review and Meta-analysis. JAMA Psychiatry 2019, 76, 426. [Google Scholar] [CrossRef]
- Sarne, Y. Beneficial and deleterious effects of cannabinoids in the brain: The case of ultra-low dose THC. Am. J. Drug Alcohol. Abus. 2019, 45, 551–562. [Google Scholar] [CrossRef]
- Emrich, H.M.; Leweke, F.M.; Schneider, U. Towards a Cannabinoid Hypothesis of Schizophrenia: Cognitive Impairments Due to Dysregulation of the Endogenous Cannabinoid System. Pharmacol. Biochem. Behav. 1997, 56, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Iacopetti, C.L.; Packer, C.D. Cannabinoid Hyperemesis Syndrome: A Case Report and Review of Pathophysiology. Clin. Med. Res. 2014, 12, 65–67. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, S.; Strawn, J.; Poweleit, E.; Sarangdhar, M.; Ramsey, L. The Impact of Marijuana on Antidepressant Treatment in Adolescents: Clinical and Pharmacologic Considerations. J. Pers. Med. 2021, 11, 615. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Statist. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Deutsch, D.G. A Personal Retrospective: Elevating Anandamide (AEA) by Targeting Fatty Acid Amide Hydrolase (FAAH) and the Fatty Acid Binding Proteins (FABPs). Front. Pharmacol. 2016, 7, 370. [Google Scholar] [CrossRef]
- Elmes, M.W.; Kaczocha, M.; Berger, W.T.; Leung, K.; Ralph, B.P.; Wang, L.; Sweeney, J.M.; Miyauchi, J.T.; Tsirka, S.E.; Ojima, I.; et al. Fatty Acid-binding Proteins (FABPs) Are Intracellular Carriers for Δ9-Tetrahydrocannabinol (THC) and Cannabidiol (CBD). J. Biol. Chem. 2015, 290, 8711–8721. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Misawa, K.; Yamamoto, I.; Watanabe, K. Cannabidiolic Acid as a Selective Cyclooxygenase-2 Inhibitory Component in Cannabis. Drug Metab. Dispos. 2008, 36, 1917–1921. [Google Scholar] [CrossRef]
- Devinsky, O.; Patel, A.D.; Thiele, E.A.; Wong, M.H.; Appleton, R.; Harden, C.L.; Greenwood, S.; Morrison, G.; Sommerville, K.; On behalf of the GWPCARE1 Part A Study Group. Randomized, dose-ranging safety trial of cannabidiol in Dravet syndrome. Neurology 2018, 90, e1204–e1211. [Google Scholar] [CrossRef]
- Massi, P.; Valenti, M.; Vaccani, A.; Gasperi, V.; Perletti, G.; Marras, E.; Fezza, F.; Maccarrone, M.; Parolaro, D. 5-Lipoxygenase and anandamide hydrolase (FAAH) mediate the antitumor activity of cannabidiol, a non-psychoactive cannabinoid. J. Neurochem. 2008, 104, 1091–1100. [Google Scholar] [CrossRef]
- Ghosh, A.; Chen, F.; Thakur, A.; Hong, H. Cysteinyl Leukotrienes and Their Receptors: Emerging Therapeutic Targets in Central Nervous System Disorders. CNS Neurosci. Ther. 2016, 22, 943–951. [Google Scholar] [CrossRef]
- Rossi, S.; De Chiara, V.; Musella, A.; Sacchetti, L.; Cantarella, C.; Castelli, M.; Cavasinni, F.; Motta, C.; Studer, V.; Bernardi, G.; et al. Preservation of Striatal Cannabinoid CB1 Receptor Function Correlates with the Antianxiety Effects of Fatty Acid Amide Hydrolase Inhibition. Mol. Pharmacol. 2010, 78, 260–268. [Google Scholar] [CrossRef]
- De Chiara, V.; Angelucci, F.; Rossi, S.; Musella, A.; Cavasinni, F.; Cantarella, C.; Mataluni, G.; Sacchetti, L.; Napolitano, F.; Castelli, M.; et al. Brain-Derived Neurotrophic Factor Controls Cannabinoid CB1 Receptor Function in the Striatum. J. Neurosci. 2010, 30, 8127–8137. [Google Scholar] [CrossRef]
- Rubino, T.; Guidali, C.; Vigano, D.; Realini, N.; Valenti, M.; Massi, P.; Parolaro, D. CB1 receptor stimulation in specific brain areas differently modulate anxiety-related behaviour. Neuropharmacology 2008, 54, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.A.; Purrio, M.; Viveros, M.-P.; Lutz, B. Biphasic Effects of Cannabinoids in Anxiety Responses: CB1 and GABAB Receptors in the Balance of GABAergic and Glutamatergic Neurotransmission. Neuropsychopharmacology 2012, 37, 2624–2634. [Google Scholar] [CrossRef] [PubMed]
- Haller, J.; Bakos, N.; Szirmay, M.; Ledent, C.; Freund, T.F. The effects of genetic and pharmacological blockade of the CB1 cannabinoid receptor on anxiety: Anxiety and cannabinoids. Eur. J. Neurosci. 2002, 16, 1395–1398. [Google Scholar] [CrossRef]
- Su, F.; Yi, H.; Xu, L.; Zhang, Z. Fluoxetine and S-citalopram inhibit M1 activation and promote M2 activation of microglia in vitro. Neuroscience 2015, 294, 60–68. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.-P.; Patel, S.; Meozzi, P.A.; Myers, L.; Perchuk, A.; Mora, Z.; Tagliaferro, P.A.; Gardner, E.; et al. Functional Expression of Brain Neuronal CB2 Cannabinoid Receptors Are Involved in the Effects of Drugs of Abuse and in Depression. Ann. N. Y. Acad. Sci. 2008, 1139, 434–449. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; Manzanares, J. Overexpression of CB2 cannabinoid receptors decreased vulnerability to anxiety and impaired anxiolytic action of alprazolam in mice. J. Psychopharmacol. 2011, 25, 111–120. [Google Scholar] [CrossRef]
- García-Gutiérrez, M.S.; García-Bueno, B.; Zoppi, S.; Leza, J.C.; Manzanares, J. Chronic blockade of cannabinoid CB2 receptors induces anxiolytic-like actions associated with alterations in GABAA receptors: Chronic blockade of CB2 receptors is anxiolytic. Br. J. Pharmacol. 2012, 165, 951–964. [Google Scholar] [CrossRef]
- Liu, Q.-R.; Canseco-Alba, A.; Zhang, H.-Y.; Tagliaferro, P.; Chung, M.; Dennis, E.; Sanabria, B.; Schanz, N.; Escosteguy-Neto, J.C.; Ishiguro, H.; et al. Cannabinoid type 2 receptors in dopamine neurons inhibits psychomotor behaviors, alters anxiety, depression and alcohol preference. Sci. Rep. 2017, 7, 17410. [Google Scholar] [CrossRef] [PubMed]
- Bradley, S.G.; Munson, A.E.; Dewey, W.L.; Harris, L.S. Enhanced susceptibility of mice to combinations of delta 9-tetrahydrocannabinol and live or killed gram-negative bacteria. Infect. Immun. 1977, 17, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Morahan, P.S.; Klykken, P.C.; Smith, S.H.; Harris, L.S.; Munson, A.E. Effects of cannabinoids on host resistance to Listeria monocytogenes and herpes simplex virus. Infect. Immun. 1979, 23, 670–674. [Google Scholar] [CrossRef] [PubMed]
- Bradley, S.G. Drugs of Abuse and Infections. In Drugs of Abuse, Immunity, and Immunodeficiency; Friedman, H., Specter, S., Klein, T.W., Eds.; Advances in Experimental Medicine and Biology; Springer: Boston, MA, USA, 1991; Volume 288, pp. 119–125. ISBN 978-1-4684-5927-2. [Google Scholar] [CrossRef]
- Kraft, B.; Wintersberger, W.; Kress, H.G. Cannabinoid receptor-independent suppression of the superoxide generation of human neutrophils (PMN) by CP55 940, but not by anandamide. Life Sci. 2004, 75, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Akinola, O.; Ogbeche, E.O.; Olumoh-Abdul, H.A.; Alli-Oluwafuyi, A.O.; Oyewole, A.L.; Amin, A.; AbdulMajeed, W.I.; Olajide, O.J.; Nafiu, A.B.; Njan, A.A.; et al. Oral Ingestion of Cannabis sativa: Risks, Benefits, and Effects on Malaria-Infected Hosts. Cannabis Cannabinoid Res. 2018, 3, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Oxenius, A. Interleukin 2: From immunostimulation to immunoregulation and back again. EMBO Rep. 2007, 8, 1142–1148. [Google Scholar] [CrossRef]
- Ito, T.; Connett, J.M.; Kunkel, S.L.; Matsukawa, A. The linkage of innate and adaptive immune response during granulomatous development. Front. Immun. 2013, 4, 10. [Google Scholar] [CrossRef]
- Dotsey, E.; Ushach, I.; Pone, E.; Nakajima, R.; Jasinskas, A.; Argueta, D.A.; Dillon, A.; DiPatrizio, N.; Davies, H.; Zlotnik, A.; et al. Transient Cannabinoid Receptor 2 Blockade during Immunization Heightens Intensity and Breadth of Antigen-specific Antibody Responses in Young and Aged mice. Sci. Rep. 2017, 7, 42584. [Google Scholar] [CrossRef]
- Sohal, A.; Chaudhry, H.; Dhaliwal, A.; Sharma, R.; Singla, P.; Sodhi, S.; Gupta, G.; Dukovic, D.; Kowdley, K. S1364 Effect of Cannabis on Mortality and Resource Utilisation in Patients with Acute on Chronic Liver Failure: Nationwide Analysis. Am. J. Gastroenterol. 2022, 117, e979–e980. [Google Scholar] [CrossRef]
- Yekhtin, Z.; Khuja, I.; Meiri, D.; Or, R.; Almogi-Hazan, O. Differential Effects of D9 Tetrahydrocannabinol (THC)- and Cannabidiol (CBD)-Based Cannabinoid Treatments on Macrophage Immune Function In Vitro and on Gastrointestinal Inflammation in a Murine Model. Biomedicines 2022, 10, 1793. [Google Scholar] [CrossRef]
- Aswad, M.; Hamza, H.; Pechkovsky, A.; Zikrach, A.; Popov, T.; Zohar, Y.; Shahar, E.; Louria-Hayon, I. High-CBD Extract (CBD-X) Downregulates Cytokine Storm Systemically and Locally in Inflamed Lungs. Front. Immunol. 2022, 13, 875546. [Google Scholar] [CrossRef]
- Khuja, I.; Yekhtin, Z.; Or, R.; Almogi-Hazan, O. Cannabinoids Reduce Inflammation but Inhibit Lymphocyte Recovery in Murine Models of Bone Marrow Transplantation. Int. J. Mol. Sci. 2019, 20, 668. [Google Scholar] [CrossRef] [PubMed]
- Manoharan, R.; Kemper, J.; Young, J. Exploring the medical cannabis prescribing behaviours of New Zealand physicians. Drug Alcohol. Rev. 2022, 41, 1355–1366. [Google Scholar] [CrossRef] [PubMed]
- Knaub, K.; Sartorius, T.; Dharsono, T.; Wacker, R.; Wilhelm, M.; Schön, C. A Novel Self-Emulsifying Drug Delivery System (SEDDS) Based on VESIsorb® Formulation Technology Improving the Oral Bioavailability of Cannabidiol in Healthy Subjects. Molecules 2019, 24, 2967. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Brain Region | Drug | R6/1 | WT | |
|---|---|---|---|---|
| Aggregate number | Striatum | HU210 | Increased | N/A |
| CB1 ligand binding | Striatum | URB597 | Increased | None |
| Hippocampus | THC | None | Decreased | |
| CB1 mRNA | Striatum | HU210 URB597 | None | Decreased |
| GABAA ligand binding | Globus pallidus | URB597 | None | Increased |
| 5HT2A ligand binding | Striatum | HU210 URB597 | None | Decreased |
| Hippocampus | URB597 | None | Decreased | |
| Motor cortex | URB597 | None | Decreased |
| CB1 Receptor | CB2 Receptor | Endocannabinoid Levels | Endocannabinoid Synthesis | Endocannabinoid Degradation | |
|---|---|---|---|---|---|
| Alzheimer’s disease | CB1 receptor expression initially increased, followed by a decline during disease progression [140]. CB1 receptor was functionally impaired [141]. | CB2 receptor increased in the entorhinal cortex and parahippocamus [142]. | Decreased AEA levels in the midfrontal and temporal cortex [143]. | DGLalfa and DGLbeta levels were increased in AD patients (Braak stage IV) [144]. | Increased FAAH levels [145]. Increased MGL levels in AD patients (Braak stage IV) [144]. |
| Parkinson’s disease | CB1 receptor expression decreased in the substantia nigra. CB1 receptor expression increased in dopaminergic projecting areas [146] | AEA levels increased in cerebrospinal fluid. A sevenfold increase in 2AG levels in the globus pallidus [147]. | - | Decreased levels of anandamide membrane transporter and FAAH [145]. | |
| Huntington’s disease | CB1 receptor expression decreased in the caudate nucleus, putamen, and globus pallidus [148]. | CB2 receptor expression increased in striatal microglia [137]. | AEA and 2AG levels decreased in the striatum. AEA levels increased and 2AG levels decreased in the cortex [145]. | NAPE-PLD and DGL levels decreased in the striatum [145]. | FAAH levels increased and MGL levels decreased in the cortex [145]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voicu, V.; Brehar, F.-M.; Toader, C.; Covache-Busuioc, R.-A.; Corlatescu, A.D.; Bordeianu, A.; Costin, H.P.; Bratu, B.-G.; Glavan, L.-A.; Ciurea, A.V. Cannabinoids in Medicine: A Multifaceted Exploration of Types, Therapeutic Applications, and Emerging Opportunities in Neurodegenerative Diseases and Cancer Therapy. Biomolecules 2023, 13, 1388. https://doi.org/10.3390/biom13091388
Voicu V, Brehar F-M, Toader C, Covache-Busuioc R-A, Corlatescu AD, Bordeianu A, Costin HP, Bratu B-G, Glavan L-A, Ciurea AV. Cannabinoids in Medicine: A Multifaceted Exploration of Types, Therapeutic Applications, and Emerging Opportunities in Neurodegenerative Diseases and Cancer Therapy. Biomolecules. 2023; 13(9):1388. https://doi.org/10.3390/biom13091388
Chicago/Turabian StyleVoicu, Victor, Felix-Mircea Brehar, Corneliu Toader, Razvan-Adrian Covache-Busuioc, Antonio Daniel Corlatescu, Andrei Bordeianu, Horia Petre Costin, Bogdan-Gabriel Bratu, Luca-Andrei Glavan, and Alexandru Vlad Ciurea. 2023. "Cannabinoids in Medicine: A Multifaceted Exploration of Types, Therapeutic Applications, and Emerging Opportunities in Neurodegenerative Diseases and Cancer Therapy" Biomolecules 13, no. 9: 1388. https://doi.org/10.3390/biom13091388