
Pharmacology of Adenosine Receptors: Recent Advancements
 , , ,
, , ,  ,
,
Abstract
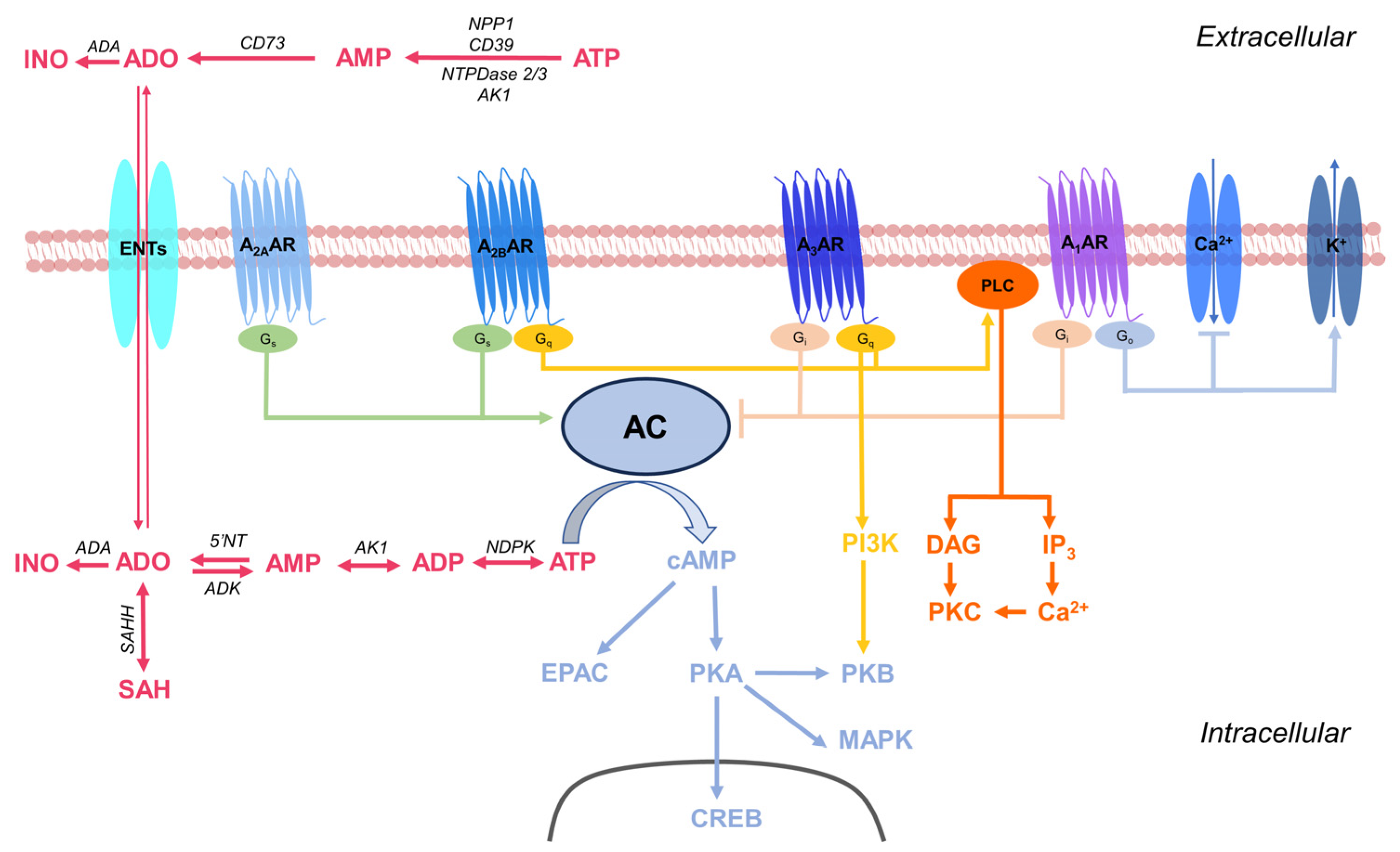
:1. Introduction
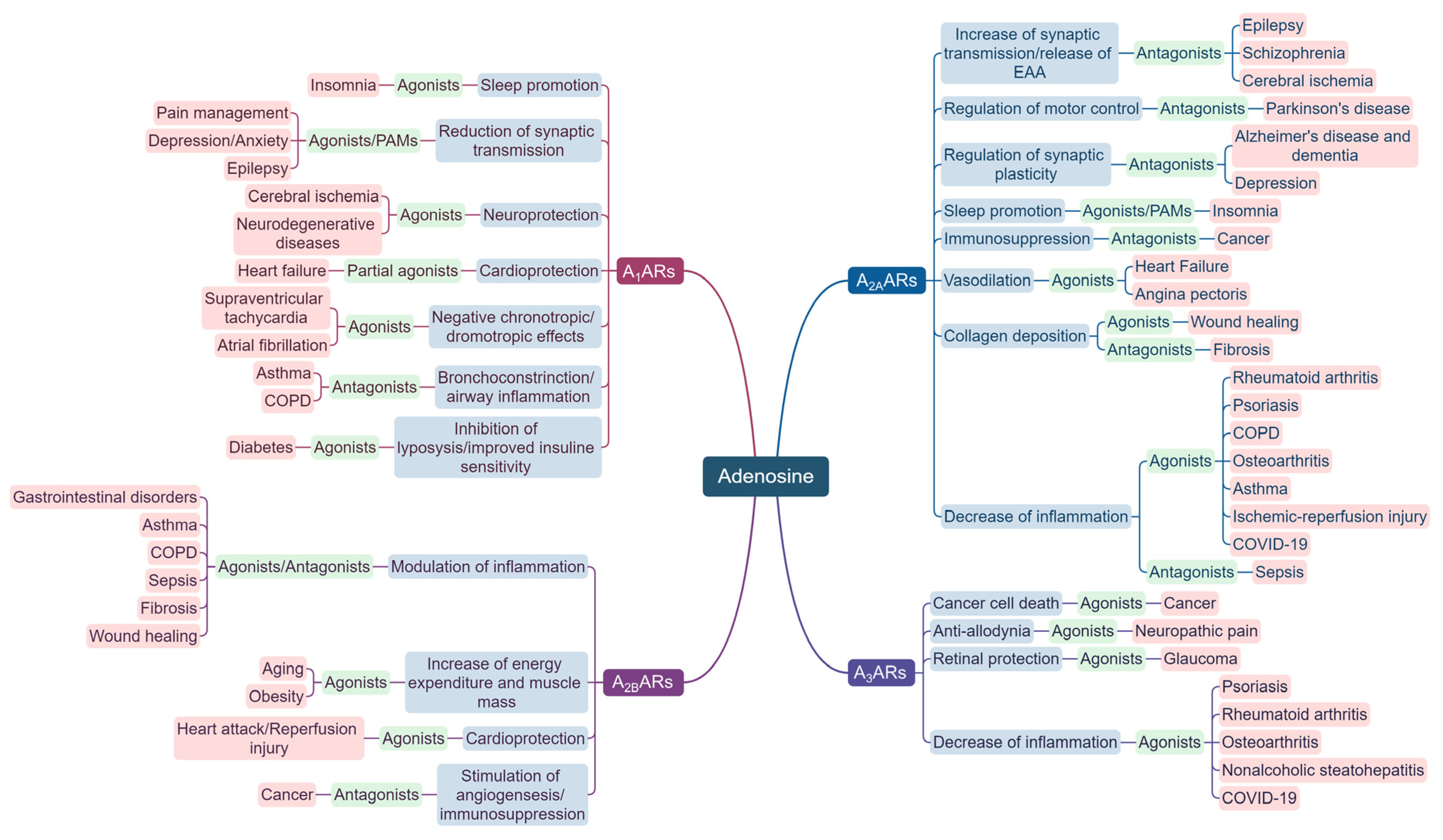
2. ARs in CNS Diseases
2.1. Recent Advancements in AR Modulation for Pain Management
2.2. Recent Advancements in AR Modulation for Neurodegenerative Diseases
2.2.1. Alzheimer’s Disease
2.2.2. Parkinson’s Disease
2.2.3. Multiple Sclerosis
2.3. Recent Advancements in AR Modulation for Brain Injury
2.3.1. Traumatic Brain Injury
2.3.2. Cerebral Ischemia
2.3.3. Chemotherapy-Induced Neurotoxicity
2.4. Recent Advancements in AR Modulation for Epilepsy
2.5. Recent Advancements in AR Modulation for Neuropsychiatric Disorders
2.5.1. Depression
2.5.2. Anxiety
2.5.3. Schizophrenia
2.6. Recent Advancements in AR Modulation for Sleep Disorders
2.7. Recent Advancements in AR Modulation for Eye Diseases
3. ARs in Cardiovascular and Metabolic Diseases
3.1. Recent Advancements in AR Modulation for Ischemic Heart Disease
3.2. Recent Advancements in AR Modulation for Hypertension
3.3. Recent Advancements in AR Modulation for the Regulation of Angiogenesis
3.4. Recent Advancements in AR Modulation for Metabolic Diseases
4. ARs in Inflammation and Autoimmunity
4.1. Recent Advancements in AR Modulation for Autoimmunity Diseases
4.2. Recent Advancements in AR Modulation for Osteoarthritis
4.3. Recent Advancements in AR Modulation for Respiratory Diseases
4.4. Recent Advancements in AR Modulation for Sepsis
5. ARs in Cancer
5.1. Recent Advancements in A2AAR Modulation for Cancer
5.2. Recent Advancements in A2BAR Modulation for Cancer
5.3. Recent Advancements in A1 and A3 AR Modulation for Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Linden, J.; Koch-Nolte, F.; Dahl, G. Purine Release, Metabolism, and Signaling in the Inflammatory Response. Annu. Rev. Immunol. 2019, 37, 325–347. [Google Scholar] [CrossRef]
- Burnstock, G. Purine and Purinergic Receptors. Brain Neurosci. Adv. 2018, 2, 2398212818817494. [Google Scholar] [CrossRef]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pharmacology of Adenosine Receptors: The State of the Art. Physiol. Rev. 2018, 98, 1591–1625. [Google Scholar] [CrossRef]
- Vincenzi, F.; Pasquini, S.; Borea, P.A.; Varani, K. Targeting Adenosine Receptors: A Potential Pharmacological Avenue for Acute and Chronic Pain. Int. J. Mol. Sci. 2020, 21, 8710. [Google Scholar] [CrossRef]
- Pasquini, S.; Contri, C.; Borea, P.A.; Vincenzi, F.; Varani, K. Adenosine and Inflammation: Here, There and Everywhere. Int. J. Mol. Sci. 2021, 22, 7685. [Google Scholar] [CrossRef]
- Chang, C.-P.; Wu, K.-C.; Lin, C.-Y.; Chern, Y. Emerging Roles of Dysregulated Adenosine Homeostasis in Brain Disorders with a Specific Focus on Neurodegenerative Diseases. J. Biomed. Sci. 2021, 28, 70. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, S.; Contri, C.; Cappello, M.; Borea, P.A.; Varani, K.; Vincenzi, F. Update on the Recent Development of Allosteric Modulators for Adenosine Receptors and Their Therapeutic Applications. Front. Pharmacol. 2022, 13, 1030895. [Google Scholar] [CrossRef]
- Borea, P.A.; Gessi, S.; Merighi, S.; Vincenzi, F.; Varani, K. Pathological Overproduction: The Bad Side of Adenosine. Br. J. Pharmacol. 2017, 174, 1945–1960. [Google Scholar] [CrossRef]
- Shaw, S.; Uniyal, A.; Gadepalli, A.; Tiwari, V.; Belinskaia, D.A.; Shestakova, N.N.; Venugopala, K.N.; Deb, P.K.; Tiwari, V. Adenosine Receptor Signalling: Probing the Potential Pathways for the Ministration of Neuropathic Pain. Eur. J. Pharmacol. 2020, 889, 173619. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Tosh, D.K.; Jain, S.; Gao, Z.-G. Historical and Current Adenosine Receptor Agonists in Preclinical and Clinical Development. Front. Cell. Neurosci. 2019, 13, 124. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Giancotti, L.A.; Lauro, F.; Mufti, F.; Salvemini, D. Treatment of Chronic Neuropathic Pain: Purine Receptor Modulation. Pain 2020, 161, 1425–1441. [Google Scholar] [CrossRef]
- Vincenzi, F.; Pasquini, S.; Battistello, E.; Merighi, S.; Gessi, S.; Borea, P.A.; Varani, K. A1 Adenosine Receptor Partial Agonists and Allosteric Modulators: Advancing Toward the Clinic? Front. Pharmacol. 2020, 11, 625134. [Google Scholar] [CrossRef]
- Vincenzi, F.; Targa, M.; Romagnoli, R.; Merighi, S.; Gessi, S.; Baraldi, P.G.; Borea, P.A.; Varani, K. TRR469, a Potent A(1) Adenosine Receptor Allosteric Modulator, Exhibits Anti-Nociceptive Properties in Acute and Neuropathic Pain Models in Mice. Neuropharmacology 2014, 81, 6–14. [Google Scholar] [CrossRef]
- Draper-Joyce, C.J.; Bhola, R.; Wang, J.; Bhattarai, A.; Nguyen, A.T.N.; Cowie-Kent, I.; O’Sullivan, K.; Chia, L.Y.; Venugopal, H.; Valant, C.; et al. Positive Allosteric Mechanisms of Adenosine A1 Receptor-Mediated Analgesia. Nature 2021, 597, 571–576. [Google Scholar] [CrossRef]
- Kan, H.-W.; Chang, C.-H.; Lin, C.-L.; Lee, Y.-C.; Hsieh, S.-T.; Hsieh, Y.-L. Downregulation of Adenosine and Adenosine A1 Receptor Contributes to Neuropathic Pain in Resiniferatoxin Neuropathy. Pain 2018, 159, 1580–1591. [Google Scholar] [CrossRef]
- Wall, M.J.; Hill, E.; Huckstepp, R.; Barkan, K.; Deganutti, G.; Leuenberger, M.; Preti, B.; Winfield, I.; Carvalho, S.; Suchankova, A.; et al. Selective Activation of Gαob by an Adenosine A1 Receptor Agonist Elicits Analgesia without Cardiorespiratory Depression. Nat. Commun. 2022, 13, 4150. [Google Scholar] [CrossRef]
- Luongo, L.; Salvemini, D. Targeting Metabotropic Adenosine Receptors for Neuropathic Pain: Focus on A2A. Brain. Behav. Immun. 2018, 69, 60–61. [Google Scholar] [CrossRef]
- Kwilasz, A.J.; Green Fulgham, S.M.; Ellis, A.; Patel, H.P.; Duran-Malle, J.C.; Favret, J.; Harvey, L.O.; Rieger, J.; Maier, S.F.; Watkins, L.R. A Single Peri-Sciatic Nerve Administration of the Adenosine 2A Receptor Agonist ATL313 Produces Long-Lasting Anti-Allodynia and Anti-Inflammatory Effects in Male Rats. Brain. Behav. Immun. 2019, 76, 116–125. [Google Scholar] [CrossRef]
- Betti, M.; Catarzi, D.; Varano, F.; Falsini, M.; Varani, K.; Vincenzi, F.; Pasquini, S.; di Cesare Mannelli, L.; Ghelardini, C.; Lucarini, E.; et al. Modifications on the Amino-3,5-Dicyanopyridine Core To Obtain Multifaceted Adenosine Receptor Ligands with Antineuropathic Activity. J. Med. Chem. 2019, 62, 6894–6912. [Google Scholar] [CrossRef]
- Falsini, M.; Catarzi, D.; Varano, F.; Ceni, C.; Dal Ben, D.; Marucci, G.; Buccioni, M.; Volpini, R.; Di Cesare Mannelli, L.; Lucarini, E.; et al. Antioxidant-Conjugated 1,2,4-Triazolo[4,3-a]Pyrazin-3-One Derivatives: Highly Potent and Selective Human A2A Adenosine Receptor Antagonists Possessing Protective Efficacy in Neuropathic Pain. J. Med. Chem. 2019, 62, 8511–8531. [Google Scholar] [CrossRef]
- Varano, F.; Catarzi, D.; Vincenzi, F.; Betti, M.; Falsini, M.; Ravani, A.; Borea, P.A.; Colotta, V.; Varani, K. Design, Synthesis, and Pharmacological Characterization of 2-(2-Furanyl)Thiazolo[5,4-d]Pyrimidine-5,7-Diamine Derivatives: New Highly Potent A2A Adenosine Receptor Inverse Agonists with Antinociceptive Activity. J. Med. Chem. 2016, 59, 10564–10576. [Google Scholar] [CrossRef]
- Coppi, E.; Cherchi, F.; Lucarini, E.; Ghelardini, C.; Pedata, F.; Jacobson, K.A.; Di Cesare Mannelli, L.; Pugliese, A.M.; Salvemini, D. Uncovering the Mechanisms of Adenosine Receptor-Mediated Pain Control: Focus on the A3 Receptor Subtype. Int. J. Mol. Sci. 2021, 22, 7952. [Google Scholar] [CrossRef]
- Durante, M.; Squillace, S.; Lauro, F.; Giancotti, L.A.; Coppi, E.; Cherchi, F.; Di Cesare Mannelli, L.; Ghelardini, C.; Kolar, G.; Wahlman, C.; et al. Adenosine A3 Agonists Reverse Neuropathic Pain via T Cell-Mediated Production of IL-10. J. Clin. Investig. 2021, 131, e139299. [Google Scholar] [CrossRef]
- Merighi, S.; Nigro, M.; Travagli, A.; Pasquini, S.; Borea, P.A.; Varani, K.; Vincenzi, F.; Gessi, S. A2A Adenosine Receptor: A Possible Therapeutic Target for Alzheimer’s Disease by Regulating NLRP3 Inflammasome Activity? Int. J. Mol. Sci. 2022, 23, 5056. [Google Scholar] [CrossRef]
- Launay, A.; Nebie, O.; Vijaya Shankara, J.; Lebouvier, T.; Buée, L.; Faivre, E.; Blum, D. The Role of Adenosine A2A Receptors in Alzheimer’s Disease and Tauopathies. Neuropharmacology 2023, 226, 109379. [Google Scholar] [CrossRef]
- Gomez-Castro, F.; Zappettini, S.; Pressey, J.C.; Silva, C.G.; Russeau, M.; Gervasi, N.; Figueiredo, M.; Montmasson, C.; Renner, M.; Canas, P.M.; et al. Convergence of Adenosine and GABA Signaling for Synapse Stabilization during Development. Science 2021, 374, eabk2055. [Google Scholar] [CrossRef]
- Temido-Ferreira, M.; Ferreira, D.G.; Batalha, V.L.; Marques-Morgado, I.; Coelho, J.E.; Pereira, P.; Gomes, R.; Pinto, A.; Carvalho, S.; Canas, P.M.; et al. Age-Related Shift in LTD Is Dependent on Neuronal Adenosine A2A Receptors Interplay with MGluR5 and NMDA Receptors. Mol. Psychiatry 2020, 25, 1876–1900. [Google Scholar] [CrossRef]
- Albasanz, J.L.; Perez, S.; Barrachina, M.; Ferrer, I.; Martín, M. Up-Regulation of Adenosine Receptors in the Frontal Cortex in Alzheimer’s Disease. Brain Pathol. 2008, 18, 211–219. [Google Scholar] [CrossRef]
- Espinosa, J.; Rocha, A.; Nunes, F.; Costa, M.S.; Schein, V.; Kazlauckas, V.; Kalinine, E.; Souza, D.O.; Cunha, R.A.; Porciúncula, L.O. Caffeine Consumption Prevents Memory Impairment, Neuronal Damage, and Adenosine A2A Receptors Upregulation in the Hippocampus of a Rat Model of Sporadic Dementia. J. Alzheimers Dis. 2013, 34, 509–518. [Google Scholar] [CrossRef]
- Viana da Silva, S.; Haberl, M.G.; Zhang, P.; Bethge, P.; Lemos, C.; Gonçalves, N.; Gorlewicz, A.; Malezieux, M.; Gonçalves, F.Q.; Grosjean, N.; et al. Early Synaptic Deficits in the APP/PS1 Mouse Model of Alzheimer’s Disease Involve Neuronal Adenosine A2A Receptors. Nat. Commun. 2016, 7, 11915. [Google Scholar] [CrossRef]
- Orr, A.G.; Hsiao, E.C.; Wang, M.M.; Ho, K.; Kim, D.H.; Wang, X.; Guo, W.; Kang, J.; Yu, G.-Q.; Adame, A.; et al. Astrocytic Adenosine Receptor A2A and Gs-Coupled Signaling Regulate Memory. Nat. Neurosci. 2015, 18, 423–434. [Google Scholar] [CrossRef]
- Merighi, S.; Battistello, E.; Casetta, I.; Gragnaniello, D.; Poloni, T.E.; Medici, V.; Cirrincione, A.; Varani, K.; Vincenzi, F.; Borea, P.A.; et al. Upregulation of Cortical A2A Adenosine Receptors Is Reflected in Platelets of Patients with Alzheimer’s Disease. J. Alzheimers Dis. JAD 2021, 80, 1105–1117. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, K.; Faivre, E.; Pietrowski, M.J.; Marques, X.; Gomez-Murcia, V.; Deleau, A.; Huin, V.; Hansen, J.N.; Kozlov, S.; Danis, C.; et al. Exacerbation of C1q Dysregulation, Synaptic Loss and Memory Deficits in Tau Pathology Linked to Neuronal Adenosine A2A Receptor. Brain J. Neurol. 2019, 142, 3636–3654. [Google Scholar] [CrossRef]
- Silva, A.C.; Lemos, C.; Gonçalves, F.Q.; Pliássova, A.V.; Machado, N.J.; Silva, H.B.; Canas, P.M.; Cunha, R.A.; Lopes, J.P.; Agostinho, P. Blockade of Adenosine A2A Receptors Recovers Early Deficits of Memory and Plasticity in the Triple Transgenic Mouse Model of Alzheimer’s Disease. Neurobiol. Dis. 2018, 117, 72–81. [Google Scholar] [CrossRef]
- Orr, A.G.; Lo, I.; Schumacher, H.; Ho, K.; Gill, M.; Guo, W.; Kim, D.H.; Knox, A.; Saito, T.; Saido, T.C.; et al. Istradefylline Reduces Memory Deficits in Aging Mice with Amyloid Pathology. Neurobiol. Dis. 2018, 110, 29–36. [Google Scholar] [CrossRef]
- Faivre, E.; Coelho, J.E.; Zornbach, K.; Malik, E.; Baqi, Y.; Schneider, M.; Cellai, L.; Carvalho, K.; Sebda, S.; Figeac, M.; et al. Beneficial Effect of a Selective Adenosine A2A Receptor Antagonist in the APPswe/PS1dE9 Mouse Model of Alzheimer’s Disease. Front. Mol. Neurosci. 2018, 11, 235. [Google Scholar] [CrossRef]
- Dias, L.; Madeira, D.; Dias, R.; Tomé, Â.R.; Cunha, R.A.; Agostinho, P. Aβ1-42 Peptides Blunt the Adenosine A2A Receptor-Mediated Control of the Interplay between P2X7 and P2Y1 Receptors Mediated Calcium Responses in Astrocytes. Cell. Mol. Life Sci. CMLS 2022, 79, 457. [Google Scholar] [CrossRef]
- Gonçalves, F.Q.; Lopes, J.P.; Silva, H.B.; Lemos, C.; Silva, A.C.; Gonçalves, N.; Tomé, Â.R.; Ferreira, S.G.; Canas, P.M.; Rial, D.; et al. Synaptic and Memory Dysfunction in a β-Amyloid Model of Early Alzheimer’s Disease Depends on Increased Formation of ATP-Derived Extracellular Adenosine. Neurobiol. Dis. 2019, 132, 104570. [Google Scholar] [CrossRef]
- Merighi, S.; Travagli, A.; Nigro, M.; Pasquini, S.; Cappello, M.; Contri, C.; Varani, K.; Vincenzi, F.; Borea, P.A.; Gessi, S. Caffeine for Prevention of Alzheimer’s Disease: Is the A2A Adenosine Receptor Its Target? Biomolecules 2023, 13, 967. [Google Scholar] [CrossRef]
- Paiva, I.; Cellai, L.; Meriaux, C.; Poncelet, L.; Nebie, O.; Saliou, J.-M.; Lacoste, A.-S.; Papegaey, A.; Drobecq, H.; Gras, S.L.; et al. Caffeine Intake Exerts Dual Genome-Wide Effects on Hippocampal Metabolism and Learning-Dependent Transcription. J. Clin. Investig. 2022, 132, e149371. [Google Scholar] [CrossRef]
- Stazi, M.; Lehmann, S.; Sakib, M.S.; Pena-Centeno, T.; Büschgens, L.; Fischer, A.; Weggen, S.; Wirths, O. Long-Term Caffeine Treatment of Alzheimer Mouse Models Ameliorates Behavioural Deficits and Neuron Loss and Promotes Cellular and Molecular Markers of Neurogenesis. Cell. Mol. Life Sci. CMLS 2021, 79, 55. [Google Scholar] [CrossRef]
- Chen, J.-F.; Cunha, R.A. The Belated US FDA Approval of the Adenosine A2A Receptor Antagonist Istradefylline for Treatment of Parkinson’s Disease. Purinergic Signal. 2020, 16, 167. [Google Scholar] [CrossRef]
- Saini, A.; Patel, R.; Gaba, S.; Singh, G.; Gupta, G.D.; Monga, V. Adenosine Receptor Antagonists: Recent Advances and Therapeutic Perspective. Eur. J. Med. Chem. 2022, 227, 113907. [Google Scholar] [CrossRef]
- Ma, L.; Day-Cooney, J.; Benavides, O.J.; Muniak, M.A.; Qin, M.; Ding, J.B.; Mao, T.; Zhong, H. Locomotion Activates PKA through Dopamine and Adenosine in Striatal Neurons. Nature 2022, 611, 762–768. [Google Scholar] [CrossRef]
- Ohno, Y.; Okita, E.; Kawai-Uchida, M.; Fukuda, N.; Shoukei, Y.; Soshiroda, K.; Yamada, K.; Kanda, T.; Uchida, S. Anti-Parkinsonian Activity of the Adenosine A2A Receptor Antagonist/Inverse Agonist KW-6356 as Monotherapy in MPTP-Treated Common Marmosets. Eur. J. Pharmacol. 2023, 950, 175773. [Google Scholar] [CrossRef]
- Ohno, Y.; Okita, E.; Kawai-Uchida, M.; Shoukei, Y.; Soshiroda, K.; Kanda, T.; Uchida, S. The Adenosine A2A Receptor Antagonist/Inverse Agonist, KW-6356 Enhances the Anti-Parkinsonian Activity of L-DOPA with a Low Risk of Dyskinesia in MPTP-Treated Common Marmosets. J. Pharmacol. Sci. 2023, 152, 193–199. [Google Scholar] [CrossRef]
- Carmo, M.; Gonçalves, F.Q.; Canas, P.M.; Oses, J.-P.; Fernandes, F.D.; Duarte, F.V.; Palmeira, C.M.; Tomé, A.R.; Agostinho, P.; Andrade, G.M.; et al. Enhanced ATP Release and CD73-Mediated Adenosine Formation Sustain Adenosine A2A Receptor over-Activation in a Rat Model of Parkinson’s Disease. Br. J. Pharmacol. 2019, 176, 3666–3680. [Google Scholar] [CrossRef]
- Meng, F.; Guo, Z.; Hu, Y.; Mai, W.; Zhang, Z.; Zhang, B.; Ge, Q.; Lou, H.; Guo, F.; Chen, J.; et al. CD73-Derived Adenosine Controls Inflammation and Neurodegeneration by Modulating Dopamine Signalling. Brain J. Neurol. 2019, 142, 700–718. [Google Scholar] [CrossRef]
- Gonçalves, F.Q.; Matheus, F.C.; Silva, H.B.; Real, J.I.; Rial, D.; Rodrigues, R.J.; Oses, J.-P.; Silva, A.C.; Gonçalves, N.; Prediger, R.D.; et al. Increased ATP Release and Higher Impact of Adenosine A2A Receptors on Corticostriatal Plasticity in a Rat Model of Presymptomatic Parkinson’s Disease. Mol. Neurobiol. 2023, 60, 1659–1674. [Google Scholar] [CrossRef]
- Ren, X.; Chen, J.-F. Caffeine and Parkinson’s Disease: Multiple Benefits and Emerging Mechanisms. Front. Neurosci. 2020, 14, 602697. [Google Scholar] [CrossRef]
- Kim, I.Y.; O’Reilly, É.J.; Hughes, K.C.; Gao, X.; Schwarzschild, M.A.; Ascherio, A. Differences in Parkinson’s Disease Risk with Caffeine Intake and Postmenopausal Hormone Use. J. Park. Dis. 2017, 7, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, K.; Miura, Y.; Wagatsuma, K.; Toyohara, J.; Ishiwata, K.; Ishii, K. Adenosine A2A Receptor Occupancy by Caffeine After Coffee Intake in Parkinson’s Disease. Mov. Disord. Off. J. Mov. Disord. Soc. 2022, 37, 853–857. [Google Scholar] [CrossRef] [PubMed]
- Duarte-Silva, E.; Ulrich, H.; Oliveira-Giacomelli, Á.; Hartung, H.-P.; Meuth, S.G.; Peixoto, C.A. The Adenosinergic Signaling in the Pathogenesis and Treatment of Multiple Sclerosis. Front. Immunol. 2022, 13, 946698. [Google Scholar] [CrossRef] [PubMed]
- Loram, L.C.; Strand, K.A.; Taylor, F.R.; Sloane, E.; Van Dam, A.-M.; Rieger, J.; Maier, S.F.; Watkins, L.R. Adenosine 2A Receptor Agonism: A Single Intrathecal Administration Attenuates Motor Paralysis in Experimental Autoimmune Encephalopathy in Rats. Brain. Behav. Immun. 2015, 46, 50–54. [Google Scholar] [CrossRef]
- Vincenzi, F.; Corciulo, C.; Targa, M.; Merighi, S.; Gessi, S.; Casetta, I.; Gentile, M.; Granieri, E.; Borea, P.A.; Varani, K. Multiple Sclerosis Lymphocytes Upregulate A2A Adenosine Receptors That Are Antiinflammatory When Stimulated. Eur. J. Immunol. 2013, 43, 2206–2216. [Google Scholar] [CrossRef]
- Ingwersen, J.; Wingerath, B.; Graf, J.; Lepka, K.; Hofrichter, M.; Schröter, F.; Wedekind, F.; Bauer, A.; Schrader, J.; Hartung, H.-P.; et al. Dual Roles of the Adenosine A2a Receptor in Autoimmune Neuroinflammation. J. Neuroinflamm. 2016, 13, 48. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, Z.-X.; Zheng, L.-P.; Wang, L.; Liu, Y.-F.; Yin, W.-Y.; Chen, Y.-Y.; Wang, X.-S.; Hou, S.-T.; Chen, J.-F.; et al. The Adenosine A2A Receptor Antagonist SCH58261 Reduces Macrophage/Microglia Activation and Protects against Experimental Autoimmune Encephalomyelitis in Mice. Neurochem. Int. 2019, 129, 104490. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, Y.; Zeng, Z.; Ye, M.; Wang, M.; Liu, X.; Tang, P.; Shang, H.; Sun, X.; Lin, X.; et al. Choroid Plexus-Selective Inactivation of Adenosine A2A Receptors Protects against T Cell Infiltration and Experimental Autoimmune Encephalomyelitis. J. Neuroinflamm. 2022, 19, 52. [Google Scholar] [CrossRef]
- Coppi, E.; Cherchi, F.; Fusco, I.; Dettori, I.; Gaviano, L.; Magni, G.; Catarzi, D.; Colotta, V.; Varano, F.; Rossi, F.; et al. Adenosine A2B Receptors Inhibit K+ Currents and Cell Differentiation in Cultured Oligodendrocyte Precursor Cells and Modulate Sphingosine-1-Phosphate Signaling Pathway. Biochem. Pharmacol. 2020, 177, 113956. [Google Scholar] [CrossRef]
- Liu, Y.-J.; Chen, J.; Li, X.; Zhou, X.; Hu, Y.-M.; Chu, S.-F.; Peng, Y.; Chen, N.-H. Research Progress on Adenosine in Central Nervous System Diseases. CNS Neurosci. Ther. 2019, 25, 899–910. [Google Scholar] [CrossRef]
- Schädlich, I.S.; Winzer, R.; Stabernack, J.; Tolosa, E.; Magnus, T.; Rissiek, B. The Role of the ATP-Adenosine Axis in Ischemic Stroke. Semin. Immunopathol. 2023, 45, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Cen, X.-Q.; Li, P.; Wang, B.; Chen, X.; Zhao, Y.; Yang, N.; Peng, Y.; Li, C.-H.; Ning, Y.-L.; Zhou, Y.-G. Knockdown of Adenosine A2A Receptors in Hippocampal Neurons Prevents Post-TBI Fear Memory Retrieval. Exp. Neurol. 2023, 364, 114378. [Google Scholar] [CrossRef]
- Farr, S.A.; Cuzzocrea, S.; Esposito, E.; Campolo, M.; Niehoff, M.L.; Doyle, T.M.; Salvemini, D. Adenosine A3 Receptor as a Novel Therapeutic Target to Reduce Secondary Events and Improve Neurocognitive Functions Following Traumatic Brain Injury. J. Neuroinflamm. 2020, 17, 339. [Google Scholar] [CrossRef] [PubMed]
- Pedata, F.; Dettori, I.; Coppi, E.; Melani, A.; Fusco, I.; Corradetti, R.; Pugliese, A.M. Purinergic Signalling in Brain Ischemia. Neuropharmacology 2016, 104, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Martire, A.; Lambertucci, C.; Pepponi, R.; Ferrante, A.; Benati, N.; Buccioni, M.; Dal Ben, D.; Marucci, G.; Klotz, K.-N.; Volpini, R.; et al. Neuroprotective Potential of Adenosine A1 Receptor Partial Agonists in Experimental Models of Cerebral Ischemia. J. Neurochem. 2019, 149, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Joya, A.; Ardaya, M.; Montilla, A.; Garbizu, M.; Plaza-García, S.; Gómez-Vallejo, V.; Padro, D.; Gutiérrez, J.J.; Rios, X.; Ramos-Cabrer, P.; et al. In Vivo Multimodal Imaging of Adenosine A1 Receptors in Neuroinflammation after Experimental Stroke. Theranostics 2021, 11, 410–425. [Google Scholar] [CrossRef]
- Zhou, Y.; Zeng, X.; Li, G.; Yang, Q.; Xu, J.; Zhang, M.; Mao, X.; Cao, Y.; Wang, L.; Xu, Y.; et al. Inactivation of Endothelial Adenosine A2A Receptors Protects Mice from Cerebral Ischaemia-induced Brain Injury. Br. J. Pharmacol. 2019, 176, 2250–2263. [Google Scholar] [CrossRef]
- Coppi, E.; Gibb, A.J. Selective Block of Adenosine A2A Receptors Prevents Ischaemic-like Effects Induced by Oxygen and Glucose Deprivation in Rat Medium Spiny Neurons. Br. J. Pharmacol. 2022, 179, 4844–4856. [Google Scholar] [CrossRef]
- Dettori, I.; Gaviano, L.; Ugolini, F.; Lana, D.; Bulli, I.; Magni, G.; Rossi, F.; Giovannini, M.G.; Pedata, F. Protective Effect of Adenosine A2B Receptor Agonist, BAY60-6583, Against Transient Focal Brain Ischemia in Rat. Front. Pharmacol. 2021, 11, 588757. [Google Scholar] [CrossRef]
- Cheng, P.; Zhang, J.; Chu, Z.; Liu, W.; Lin, H.; Wu, Y.; Zhu, J. A3 Adenosine Receptor Agonist IB-MECA Reverses Chronic Cerebral Ischemia-Induced Inhibitory Avoidance Memory Deficit. Eur. J. Pharmacol. 2022, 921, 174874. [Google Scholar] [CrossRef]
- Liston, T.E.; Hama, A.; Boltze, J.; Poe, R.B.; Natsume, T.; Hayashi, I.; Takamatsu, H.; Korinek, W.S.; Lechleiter, J.D. Adenosine A1R/A3R (Adenosine A1 and A3 Receptor) Agonist AST-004 Reduces Brain Infarction in a Nonhuman Primate Model of Stroke. Stroke 2022, 53, 238–248. [Google Scholar] [CrossRef]
- Oliveros, A.; Yoo, K.H.; Rashid, M.A.; Corujo-Ramirez, A.; Hur, B.; Sung, J.; Liu, Y.; Hawse, J.R.; Choi, D.-S.; Boison, D.; et al. Adenosine A2A Receptor Blockade Prevents Cisplatin-Induced Impairments in Neurogenesis and Cognitive Function. Proc. Natl. Acad. Sci. USA 2022, 119, e2206415119. [Google Scholar] [CrossRef]
- Singh, A.K.; Mahalingam, R.; Squillace, S.; Jacobson, K.A.; Tosh, D.K.; Dharmaraj, S.; Farr, S.A.; Kavelaars, A.; Salvemini, D.; Heijnen, C.J. Targeting the A3 Adenosine Receptor to Prevent and Reverse Chemotherapy-Induced Neurotoxicities in Mice. Acta Neuropathol. Commun. 2022, 10, 11. [Google Scholar] [CrossRef] [PubMed]
- Hargus, N.J.; Jennings, C.; Perez-Reyes, E.; Bertram, E.H.; Patel, M.K. Enhanced Actions of Adenosine in Medial Entorhinal Cortex Layer II Stellate Neurons in Temporal Lobe Epilepsy Are Mediated via A(1)-Receptor Activation. Epilepsia 2012, 53, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Lovatt, D.; Xu, Q.; Liu, W.; Takano, T.; Smith, N.A.; Schnermann, J.; Tieu, K.; Nedergaard, M. Neuronal Adenosine Release, and Not Astrocytic ATP Release, Mediates Feedback Inhibition of Excitatory Activity. Proc. Natl. Acad. Sci. USA 2012, 109, 6265–6270. [Google Scholar] [CrossRef]
- Beamer, E.; Kuchukulla, M.; Boison, D.; Engel, T. ATP and Adenosine—Two Players in the Control of Seizures and Epilepsy Development. Prog. Neurobiol. 2021, 204, 102105. [Google Scholar] [CrossRef]
- Saggu, S.; Chen, Y.; Chen, L.; Pizarro, D.; Pati, S.; Law, W.J.; McMahon, L.; Jiao, K.; Wang, Q. A Peptide Blocking the ADORA1-Neurabin Interaction Is Anticonvulsant and Inhibits Epilepsy in an Alzheimer’s Model. JCI Insight 2022, 7, e155002. [Google Scholar] [CrossRef] [PubMed]
- Xie, P.; Liu, S.; Huang, Q.; Xiong, Z.; Deng, J.; Tang, C.; Xu, K.; Zhang, B.; He, B.; Wang, X.; et al. Deep Brain Stimulation Suppresses Epileptic Seizures in Rats via Inhibition of Adenosine Kinase and Activation of Adenosine A1 Receptors. CNS Neurosci. Ther. 2023, 29, 2597–2607. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.-Y.; Baer, S.B.; Gesese, R.; Cook, J.M.; Weltha, L.; Coffman, S.Q.; Wu, J.; Chen, J.-F.; Gao, M.; Ji, T. Adenosine-A2A Receptor Signaling Plays a Crucial Role in Sudden Unexpected Death in Epilepsy. Front. Pharmacol. 2022, 13, 910535. [Google Scholar] [CrossRef]
- Domenici, M.R.; Ferrante, A.; Martire, A.; Chiodi, V.; Pepponi, R.; Tebano, M.T.; Popoli, P. Adenosine A2A Receptor as Potential Therapeutic Target in Neuropsychiatric Disorders. Pharmacol. Res. 2019, 147, 104338. [Google Scholar] [CrossRef]
- Pasquini, S.; Contri, C.; Merighi, S.; Gessi, S.; Borea, P.A.; Varani, K.; Vincenzi, F. Adenosine Receptors in Neuropsychiatric Disorders: Fine Regulators of Neurotransmission and Potential Therapeutic Targets. Int. J. Mol. Sci. 2022, 23, 1219. [Google Scholar] [CrossRef]
- Wang, M.; Li, P.; Li, Z.; da Silva, B.S.; Zheng, W.; Xiang, Z.; He, Y.; Xu, T.; Cordeiro, C.; Deng, L.; et al. Lateral Septum Adenosine A2A Receptors Control Stress-Induced Depressive-like Behaviors via Signaling to the Hypothalamus and Habenula. Nat. Commun. 2023, 14, 1880. [Google Scholar] [CrossRef]
- Yu, W.; Wu, Z.; Li, X.; Ding, M.; Xu, Y.; Zhao, P. Ketamine Counteracts Sevoflurane-Induced Depressive-like Behavior and Synaptic Plasticity Impairments through the Adenosine A2A Receptor/ERK Pathway in Rats. Mol. Neurobiol. 2023. ahead-of-print. [Google Scholar] [CrossRef]
- Xu, Y.; Ning, Y.; Zhao, Y.; Peng, Y.; Luo, F.; Zhou, Y.; Li, P. Caffeine Functions by Inhibiting Dorsal and Ventral Hippocampal Adenosine 2A Receptors to Modulate Memory and Anxiety, Respectively. Front. Pharmacol. 2022, 13, 807330. [Google Scholar] [CrossRef] [PubMed]
- Florén Lind, S.; Stam, F.; Zelleroth, S.; Meurling, E.; Frick, A.; Grönbladh, A. Acute Caffeine Differently Affects Risk-Taking and the Expression of BDNF and of Adenosine and Opioid Receptors in Rats with High or Low Anxiety-like Behavior. Pharmacol. Biochem. Behav. 2023, 227–228, 173573. [Google Scholar] [CrossRef]
- Singer, P.; Yee, B.K. The Adenosine Hypothesis of Schizophrenia into Its Third Decade: From Neurochemical Imbalance to Early Life Etiological Risks. Front. Cell. Neurosci. 2023, 17, 1120532. [Google Scholar] [CrossRef] [PubMed]
- Boison, D.; Singer, P.; Shen, H.-Y.; Feldon, J.; Yee, B.K. Adenosine Hypothesis of Schizophrenia--Opportunities for Pharmacotherapy. Neuropharmacology 2012, 62, 1527–1543. [Google Scholar] [CrossRef] [PubMed]
- Valle-León, M.; Callado, L.F.; Aso, E.; Cajiao-Manrique, M.M.; Sahlholm, K.; López-Cano, M.; Soler, C.; Altafaj, X.; Watanabe, M.; Ferré, S.; et al. Decreased Striatal Adenosine A2A-Dopamine D2 Receptor Heteromerization in Schizophrenia. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2021, 46, 665–672. [Google Scholar] [CrossRef]
- Valle-León, M.; Casajuana-Martin, N.; Del Torrent, C.L.; Argerich, J.; Gómez-Acero, L.; Sahlholm, K.; Ferré, S.; Pardo, L.; Ciruela, F. Unique Effect of Clozapine on Adenosine A2A-Dopamine D2 Receptor Heteromerization. Biomed. Pharmacother. Biomed. Pharmacother. 2023, 160, 114327. [Google Scholar] [CrossRef]
- Shalaby, H.N.; Zaki, H.F.; Ain-Shoka, A.A.A.; Mohammed, R.A. Adenosine A2A Receptor Blockade Ameliorates Mania Like Symptoms in Rats: Signaling to PKC-α and Akt/GSK-3β/β-Catenin. Mol. Neurobiol. 2022, 59, 6397–6410. [Google Scholar] [CrossRef]
- Ma, W.-X.; Yuan, P.-C.; Zhang, H.; Kong, L.-X.; Lazarus, M.; Qu, W.-M.; Wang, Y.-Q.; Huang, Z.-L. Adenosine and P1 Receptors: Key Targets in the Regulation of Sleep, Torpor, and Hibernation. Front. Pharmacol. 2023, 14, 1098976. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Wu, Z.; Song, K.; Zhang, S.; Li, Y.; Xu, M. Regulation of Sleep Homeostasis Mediator Adenosine by Basal Forebrain Glutamatergic Neurons. Science 2020, 369, eabb0556. [Google Scholar] [CrossRef] [PubMed]
- Jagannath, A.; Varga, N.; Dallmann, R.; Rando, G.; Gosselin, P.; Ebrahimjee, F.; Taylor, L.; Mosneagu, D.; Stefaniak, J.; Walsh, S.; et al. Adenosine Integrates Light and Sleep Signalling for the Regulation of Circadian Timing in Mice. Nat. Commun. 2021, 12, 2113. [Google Scholar] [CrossRef]
- Kim, T.-H.; Bormate, K.J.; Custodio, R.J.P.; Cheong, J.H.; Lee, B.K.; Kim, H.J.; Jung, Y.-S. Involvement of the Adenosine A1 Receptor in the Hypnotic Effect of Rosmarinic Acid. Biomed. Pharmacother. Biomed. Pharmacother. 2022, 146, 112483. [Google Scholar] [CrossRef]
- Lin, Y.; Roy, K.; Ioka, S.; Otani, R.; Amezawa, M.; Ishikawa, Y.; Cherasse, Y.; Kaushik, M.K.; Klewe-Nebenius, D.; Zhou, L.; et al. Positive Allosteric Adenosine A2A Receptor Modulation Suppresses Insomnia Associated with Mania- and Schizophrenia-like Behaviors in Mice. Front. Pharmacol. 2023, 14, 1072. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, Y.-Q.; Liu, W.-Y.; Zhang, M.-Q.; Li, L.; Cherasse, Y.; Schiffmann, S.N.; de Kerchove d’Exaerde, A.; Lazarus, M.; Qu, W.-M.; et al. Activation of Adenosine A2A Receptors in the Olfactory Tubercle Promotes Sleep in Rodents. Neuropharmacology 2020, 168, 107923. [Google Scholar] [CrossRef]
- Choi, I.-S.; Kim, J.-H.; Jeong, J.-Y.; Lee, M.-G.; Suk, K.; Jang, I.-S. Astrocyte-Derived Adenosine Excites Sleep-Promoting Neurons in the Ventrolateral Preoptic Nucleus: Astrocyte-Neuron Interactions in the Regulation of Sleep. Glia 2022, 70, 1864–1885. [Google Scholar] [CrossRef]
- Ye, S.-S.; Tang, Y.; Song, J.-T. ATP and Adenosine in the Retina and Retinal Diseases. Front. Pharmacol. 2021, 12, 654445. [Google Scholar] [CrossRef]
- Boia, R.; Salinas-Navarro, M.; Gallego-Ortega, A.; Galindo-Romero, C.; Aires, I.D.; Agudo-Barriuso, M.; Ambrósio, A.F.; Vidal-Sanz, M.; Santiago, A.R. Activation of Adenosine A3 Receptor Protects Retinal Ganglion Cells from Degeneration Induced by Ocular Hypertension. Cell Death Dis. 2020, 11, 401. [Google Scholar] [CrossRef]
- Boia, R.; Dias, P.A.N.; Galindo-Romero, C.; Ferreira, H.; Aires, I.D.; Vidal-Sanz, M.; Agudo-Barriuso, M.; Bernardes, R.; Santos, P.F.; de Sousa, H.C.; et al. Intraocular Implants Loaded with A3R Agonist Rescue Retinal Ganglion Cells from Ischemic Damage. J. Control. Release 2022, 343, 469–481. [Google Scholar] [CrossRef]
- Fang, G.; Zhou, Y.; Zhou, X.; Zhou, H.; Ge, Y.-Y.; Luo, S.; Chen, J.-F.; Zhang, L. The Adenosine A2A Receptor Antagonist Protects against Retinal Mitochondrial Injury in Association with an Altered Network of Competing Endogenous RNAs. Neuropharmacology 2022, 208, 108981. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Li, Y.; Zhang, Y.; Shi, R.; Tang, P.; Zhang, D.; Kuang, X.; Chen, J.; Qu, J.; Gao, Y. The Adenosine A2A Receptor Antagonist KW6002 Distinctly Regulates Retinal Ganglion Cell Morphology during Postnatal Development and Neonatal Inflammation. Front. Pharmacol. 2022, 13, 1082997. [Google Scholar] [CrossRef] [PubMed]
- Headrick, J.P.; Ashton, K.J.; Rose’Meyer, R.B.; Peart, J.N. Cardiovascular Adenosine Receptors: Expression, Actions and Interactions. Pharmacol. Ther. 2013, 140, 92–111. [Google Scholar] [CrossRef]
- Sanni, O.; Terre’Blanche, G. Therapeutic Potentials of Agonist and Antagonist of Adenosine Receptors in Type 2 Diabetes. Rev. Endocr. Metab. Disord. 2021, 22, 1073–1090. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Chen, X.; Yi, B.; Li, J.; Wen, Z. Pathophysiology Roles for Adenosine 2A Receptor in Obesity and Related Diseases. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2022, 23, e13490. [Google Scholar] [CrossRef]
- Ruan, W.; Ma, X.; Bang, I.H.; Liang, Y.; Muehlschlegel, J.D.; Tsai, K.-L.; Mills, T.W.; Yuan, X.; Eltzschig, H.K. The Hypoxia-Adenosine Link during Myocardial Ischemia-Reperfusion Injury. Biomedicines 2022, 10, 1939. [Google Scholar] [CrossRef]
- Xu, S.; Gu, R.; Bian, X.; Xu, X.; Xia, X.; Liu, Y.; Jia, C.; Gu, Y.; Zhang, H. Remote Conditioning by Rhythmic Compression of Limbs Ameliorated Myocardial Infarction by Downregulation of Inflammation via A2 Adenosine Receptors. Front. Cardiovasc. Med. 2022, 8, 723332. [Google Scholar] [CrossRef]
- Mehaffey, J.H.; Money, D.; Charles, E.J.; Schubert, S.; Piñeros, A.F.; Wu, D.; Bontha, S.V.; Hawkins, R.; Teman, N.R.; Laubach, V.E.; et al. Adenosine 2A Receptor Activation Attenuates Ischemia Reperfusion Injury During Extracorporeal Cardiopulmonary Resuscitation. Ann. Surg. 2019, 269, 1176–1183. [Google Scholar] [CrossRef]
- Ruan, W.; Li, J.; Choi, S.; Ma, X.; Liang, Y.; Nair, R.; Yuan, X.; Mills, T.W.; Eltzschig, H.K. Targeting Myocardial Equilibrative Nucleoside Transporter ENT1 Provides Cardioprotection by Enhancing Myeloid Adora2b Signaling. JCI Insight 2023, 8, e166011. [Google Scholar] [CrossRef]
- Reichert, K.P.; Castro, M.F.V.; Assmann, C.E.; Bottari, N.B.; Miron, V.V.; Cardoso, A.; Stefanello, N.; Morsch, V.M.M.; Schetinger, M.R.C. Diabetes and Hypertension: Pivotal Involvement of Purinergic Signaling. Biomed. Pharmacother. Biomed. Pharmacother. 2021, 137, 111273. [Google Scholar] [CrossRef]
- Yadav, V.R.; Teng, B.; Mustafa, S.J. Enhanced A1 Adenosine Receptor-Induced Vascular Contractions in Mesenteric Artery and Aorta of in L-NAME Mouse Model of Hypertension. Eur. J. Pharmacol. 2019, 842, 111–117. [Google Scholar] [CrossRef]
- Jackson, E.K.; Gillespie, D.G.; Mi, Z.; Cheng, D. Adenosine Receptors Influence Hypertension in Dahl Salt-Sensitive Rats: Dependence on Receptor Subtype, Salt Diet, and Sex. Hypertens. Dallas Tex 1979 2018, 72, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.-C.; Kong, L.-R.; Chen, X.-H.; Ma, Y.; Pan, X.-X.; Zhang, Z.-B.; Gao, P.-J. A2A Receptor Activation Attenuates Hypertensive Cardiac Remodeling via Promoting Brown Adipose Tissue-Derived FGF21. Cell Metab. 2018, 28, 476–489.e5. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahmad, S.; Glover, L.; Miller, S.M.; Shannon, J.M.; Guo, X.; Franklin, W.A.; Bridges, J.P.; Schaack, J.B.; Colgan, S.P.; et al. Adenosine A2A Receptor Is a Unique Angiogenic Target of HIF-2α in Pulmonary Endothelial Cells. Proc. Natl. Acad. Sci. USA 2009, 106, 10684–10689. [Google Scholar] [CrossRef]
- Angioni, R.; Liboni, C.; Herkenne, S.; Sánchez-Rodríguez, R.; Borile, G.; Marcuzzi, E.; Calì, B.; Muraca, M.; Viola, A. CD73+ Extracellular Vesicles Inhibit Angiogenesis through Adenosine A2B Receptor Signalling. J. Extracell. Vesicles 2020, 9, 1757900. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xiang, Q.; Feng, L.; Wu, D.; Huang, S.; Zhang, L.; Rao, S.; Luo, J.; Xiong, W.; Deng, J.; et al. Adenosine-ADORA2A Promotes Ang-Induced Angiogenesis in Intrauterine Growth Restriction Placenta via the Stat3/Akt Pathway. Arterioscler. Thromb. Vasc. Biol. 2023, 43, e190–e209. [Google Scholar] [CrossRef]
- Wu, Z.; Nie, J.; Wu, D.; Huang, S.; Chen, J.; Liang, H.; Hao, X.; Feng, L.; Luo, H.; Tan, C. Dietary Adenosine Supplementation Improves Placental Angiogenesis in IUGR Piglets by Up-Regulating Adenosine A2a Receptor. Anim. Nutr. Zhongguo Xu Mu Shou Yi Xue Hui 2023, 13, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Gnad, T.; Navarro, G.; Lahesmaa, M.; Reverte-Salisa, L.; Copperi, F.; Cordomi, A.; Naumann, J.; Hochhäuser, A.; Haufs-Brusberg, S.; Wenzel, D.; et al. Adenosine/A2B Receptor Signaling Ameliorates the Effects of Aging and Counteracts Obesity. Cell Metab. 2020, 32, 56–70.e7. [Google Scholar] [CrossRef]
- Kong, L.-R.; Chen, X.-H.; Sun, Q.; Zhang, K.-Y.; Xu, L.; Ding, L.; Zhou, Y.-P.; Zhang, Z.-B.; Lin, J.-R.; Gao, P.-J. Loss of C3a and C5a Receptors Promotes Adipocyte Browning and Attenuates Diet-Induced Obesity via Activating Inosine/A2aR Pathway. Cell Rep. 2023, 42, 112078. [Google Scholar] [CrossRef]
- Sacramento, J.F.; Martins, F.O.; Rodrigues, T.; Matafome, P.; Ribeiro, M.J.; Olea, E.; Conde, S.V. A2 Adenosine Receptors Mediate Whole-Body Insulin Sensitivity in a Prediabetes Animal Model: Primary Effects on Skeletal Muscle. Front. Endocrinol. 2020, 11, 262. [Google Scholar] [CrossRef]
- Antonioli, L.; Fornai, M.; Blandizzi, C.; Pacher, P.; Haskó, G. Adenosine Signaling and the Immune System: When a Lot Could Be Too Much. Immunol. Lett. 2019, 205, 9–15. [Google Scholar] [CrossRef]
- Boncler, M.; Bartczak, K.; Rozalski, M. Potential for Modulation of Platelet Function via Adenosine Receptors during Inflammation. Br. J. Pharmacol. 2023. ahead-of-print. [Google Scholar] [CrossRef] [PubMed]
- Cronstein, B.N.; Sitkovsky, M. Adenosine and Adenosine Receptors in the Pathogenesis and Treatment of Rheumatic Diseases. Nat. Rev. Rheumatol. 2017, 13, 41–51. [Google Scholar] [CrossRef]
- Varani, K.; Vincenzi, F.; Tosi, A.; Targa, M.; Masieri, F.F.; Ongaro, A.; De Mattei, M.; Massari, L.; Borea, P.A. Expression and Functional Role of Adenosine Receptors in Regulating Inflammatory Responses in Human Synoviocytes. Br. J. Pharmacol. 2010, 160, 101–115. [Google Scholar] [CrossRef]
- Varani, K.; Padovan, M.; Vincenzi, F.; Targa, M.; Trotta, F.; Govoni, M.; Borea, P.A. A2A and A3 Adenosine Receptor Expression in Rheumatoid Arthritis: Upregulation, Inverse Correlation with Disease Activity Score and Suppression of Inflammatory Cytokine and Metalloproteinase Release. Arthritis Res. Ther. 2011, 13, R197. [Google Scholar] [CrossRef] [PubMed]
- Vincenzi, F.; Padovan, M.; Targa, M.; Corciulo, C.; Giacuzzo, S.; Merighi, S.; Gessi, S.; Govoni, M.; Borea, P.A.; Varani, K. A(2A) Adenosine Receptors Are Differentially Modulated by Pharmacological Treatments in Rheumatoid Arthritis Patients and Their Stimulation Ameliorates Adjuvant-Induced Arthritis in Rats. PLoS ONE 2013, 8, e54195. [Google Scholar] [CrossRef]
- Bortoluzzi, A.; Vincenzi, F.; Govoni, M.; Padovan, M.; Ravani, A.; Borea, P.A.; Varani, K. A2A Adenosine Receptor Upregulation Correlates with Disease Activity in Patients with Systemic Lupus Erythematosus. Arthritis Res. Ther. 2016, 18, 192. [Google Scholar] [CrossRef]
- Ravani, A.; Vincenzi, F.; Bortoluzzi, A.; Padovan, M.; Pasquini, S.; Gessi, S.; Merighi, S.; Borea, P.A.; Govoni, M.; Varani, K. Role and Function of A2A and A₃ Adenosine Receptors in Patients with Ankylosing Spondylitis, Psoriatic Arthritis and Rheumatoid Arthritis. Int. J. Mol. Sci. 2017, 18, 697. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.; Barer, F.; Bar-Yehuda, S.; IJzerman, A.P.; Jacobson, K.A.; Fishman, P. A₃ Adenosine Receptor Allosteric Modulator Induces an Anti-Inflammatory Effect: In Vivo Studies and Molecular Mechanism of Action. Mediat. Inflamm. 2014, 2014, 708746. [Google Scholar] [CrossRef]
- Ochaion, A.; Bar-Yehuda, S.; Cohen, S.; Amital, H.; Jacobson, K.A.; Joshi, B.V.; Gao, Z.G.; Barer, F.; Patoka, R.; Del Valle, L.; et al. The A3 Adenosine Receptor Agonist CF502 Inhibits the PI3K, PKB/Akt and NF-KappaB Signaling Pathway in Synoviocytes from Rheumatoid Arthritis Patients and in Adjuvant-Induced Arthritis Rats. Biochem. Pharmacol. 2008, 76, 482–494. [Google Scholar] [CrossRef]
- Flögel, U.; Burghoff, S.; van Lent, P.L.E.M.; Temme, S.; Galbarz, L.; Ding, Z.; El-Tayeb, A.; Huels, S.; Bönner, F.; Borg, N.; et al. Selective Activation of Adenosine A2A Receptors on Immune Cells by a CD73-Dependent Prodrug Suppresses Joint Inflammation in Experimental Rheumatoid Arthritis. Sci. Transl. Med. 2012, 4, 146ra108. [Google Scholar] [CrossRef]
- Schmiel, S.E.; Kalekar, L.A.; Zhang, N.; Blankespoor, T.W.; Robinson, L.J.; Mueller, D.L. Adenosine 2a Receptor Signals Block Autoimmune Arthritis by Inhibiting Pathogenic Germinal Center T Follicular Helper Cells. Arthritis Rheumatol. 2019, 71, 773–783. [Google Scholar] [CrossRef]
- Sadatpour, O.; Ebrahimi, M.T.; Akhtari, M.; Ahmadzadeh, N.; Vojdanian, M.; Jamshidi, A.; Farhadi, E.; Mahmoudi, M. A2A Adenosine Receptor Agonist Reduced MMP8 Expression in Healthy M2-like Macrophages but Not in Macrophages from Ankylosing Spondylitis Patients. BMC Musculoskelet. Disord. 2022, 23, 908. [Google Scholar] [CrossRef]
- Winslow, G.M.; Papillion, A.M.; Kenderes, K.J.; Levack, R.C. CD11c+ T-Bet+ Memory B Cells: Immune Maintenance during Chronic Infection and Inflammation? Cell. Immunol. 2017, 321, 8–17. [Google Scholar] [CrossRef]
- Levack, R.C.; Newell, K.L.; Cabrera-Martinez, B.; Cox, J.; Perl, A.; Bastacky, S.I.; Winslow, G.M. Adenosine Receptor 2a Agonists Target Mouse CD11c+T-Bet+ B Cells in Infection and Autoimmunity. Nat. Commun. 2022, 13, 452. [Google Scholar] [CrossRef]
- López-Cano, M.; Filgaira, I.; Nolen, E.G.; Cabré, G.; Hernando, J.; Tosh, D.K.; Jacobson, K.A.; Soler, C.; Ciruela, F. Optical Control of Adenosine A3 Receptor Function in Psoriasis. Pharmacol. Res. 2021, 170, 105731. [Google Scholar] [CrossRef]
- Fishman, P. Drugs Targeting the A3 Adenosine Receptor: Human Clinical Study Data. Molecules 2022, 27, 3680. [Google Scholar] [CrossRef]
- Carmona-Rivera, C.; Khaznadar, S.S.; Shwin, K.W.; Irizarry-Caro, J.A.; O’Neil, L.J.; Liu, Y.; Jacobson, K.A.; Ombrello, A.K.; Stone, D.L.; Tsai, W.L.; et al. Deficiency of Adenosine Deaminase 2 Triggers Adenosine-Mediated NETosis and TNF Production in Patients with DADA2. Blood 2019, 134, 395–406. [Google Scholar] [CrossRef]
- Bekisz, J.M.; Lopez, C.D.; Corciulo, C.; Mediero, A.; Coelho, P.G.; Witek, L.; Flores, R.L.; Cronstein, B.N. The Role of Adenosine Receptor Activation in Attenuating Cartilaginous Inflammation. Inflammation 2018, 41, 1135–1141. [Google Scholar] [CrossRef]
- Corciulo, C.; Lendhey, M.; Wilder, T.; Schoen, H.; Cornelissen, A.S.; Chang, G.; Kennedy, O.D.; Cronstein, B.N. Endogenous Adenosine Maintains Cartilage Homeostasis and Exogenous Adenosine Inhibits Osteoarthritis Progression. Nat. Commun. 2017, 8, 15019. [Google Scholar] [CrossRef]
- Friedman, B.; Larranaga-Vera, A.; Castro, C.M.; Corciulo, C.; Rabbani, P.; Cronstein, B.N. Adenosine A2A Receptor Activation Reduces Chondrocyte Senescence. FASEB J. 2023, 37, e22838. [Google Scholar] [CrossRef] [PubMed]
- Castro, C.M.; Corciulo, C.; Solesio, M.E.; Liang, F.; Pavlov, E.V.; Cronstein, B.N. Adenosine A2A Receptor (A2AR) Stimulation Enhances Mitochondrial Metabolism and Mitigates Reactive Oxygen Species-Mediated Mitochondrial Injury. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 5027–5045. [Google Scholar] [CrossRef]
- Friedman, B.; Corciulo, C.; Castro, C.M.; Cronstein, B.N. Adenosine A2A Receptor Signaling Promotes FoxO Associated Autophagy in Chondrocytes. Sci. Rep. 2021, 11, 968. [Google Scholar] [CrossRef]
- Bai, H.; Zhang, Z.; Liu, L.; Wang, X.; Song, X.; Gao, L. Activation of Adenosine A3 Receptor Attenuates Progression of Osteoarthritis through Inhibiting the NLRP3/Caspase-1/GSDMD Induced Signalling. J. Cell. Mol. Med. 2022, 26, 4230–4243. [Google Scholar] [CrossRef] [PubMed]
- Le, T.-T.T.; Berg, N.K.; Harting, M.T.; Li, X.; Eltzschig, H.K.; Yuan, X. Purinergic Signaling in Pulmonary Inflammation. Front. Immunol. 2019, 10, 1633. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Schneider, D.J.; Blackburn, M.R. Adenosine Signaling and the Regulation of Chronic Lung Disease. Pharmacol. Ther. 2009, 123, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Wendell, S.G.; Fan, H.; Zhang, C. G Protein–Coupled Receptors in Asthma Therapy: Pharmacology and Drug Action. Pharmacol. Rev. 2020, 72, 1–49. [Google Scholar] [CrossRef]
- Basu, S.; Barawkar, D.A.; Ramdas, V.; Patel, M.; Waman, Y.; Panmand, A.; Kumar, S.; Thorat, S.; Naykodi, M.; Goswami, A.; et al. Design and Synthesis of Novel Xanthine Derivatives as Potent and Selective A2B Adenosine Receptor Antagonists for the Treatment of Chronic Inflammatory Airway Diseases. Eur. J. Med. Chem. 2017, 134, 218–229. [Google Scholar] [CrossRef]
- Ramos-Barbon, D.; Brienza, N.S.; Rodríguez, T.B.; Medina, É.F.M.; Saladich, I.G.; Rodríguez, M.P.; Arbos, R.M.A.; Gancedo, S.Q.; Laria, N.C.P.; Laria, J.C.P. PBF-680, an Oral A1 Adenosine Receptor Antagonist, Inhibits the Late Allergic Response (LAR) in Mild-to-Moderate Atopic Asthmatics: A Phase-IIa Trial. Eur. Respir. J. 2020, 56, 4784. [Google Scholar] [CrossRef]
- Ramos-Barbon, D.; Brienza, N.S.; Rodríguez, T.B.; Saladich, I.G.; Rodríguez, M.P.; Arbos, R.M.A.; Laria, N.C.P.; Laria, J.C.P. PBF-680, an Oral A1 Adenosine Receptor Antagonist, Inhibits Adenosine Monophosphate (AMP) Airway Hyperresponsiveness (AHR) in Mild-to-Moderate Asthma: A Phase-IIa Proof-of-Concept Trial. Eur. Respir. J. 2020, 56, 2279. [Google Scholar] [CrossRef]
- Xiao, Q.; Han, X.; Liu, G.; Zhou, D.; Zhang, L.; He, J.; Xu, H.; Zhou, P.; Yang, Q.; Chen, J.; et al. Adenosine Restrains ILC2-Driven Allergic Airway Inflammation via A2A Receptor. Mucosal Immunol. 2022, 15, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Ko, I.-G.; Hwang, J.J.; Chang, B.S.; Kim, S.-H.; Jin, J.-J.; Hwang, L.; Kim, C.-J.; Choi, C.W. Polydeoxyribonucleotide Ameliorates Lipopolysaccharide-Induced Acute Lung Injury via Modulation of the MAPK/NF-ΚB Signaling Pathway in Rats. Int. Immunopharmacol. 2020, 83, 106444. [Google Scholar] [CrossRef]
- Li, X.; Berg, N.K.; Mills, T.; Zhang, K.; Eltzschig, H.K.; Yuan, X. Adenosine at the Interphase of Hypoxia and Inflammation in Lung Injury. Front. Immunol. 2020, 11, 604944. [Google Scholar] [CrossRef] [PubMed]
- Korb, V.G.; Schultz, I.C.; Beckenkamp, L.R.; Wink, M.R. A Systematic Review of the Role of Purinergic Signalling Pathway in the Treatment of COVID-19. Int. J. Mol. Sci. 2023, 24, 7865. [Google Scholar] [CrossRef] [PubMed]
- Dorneles, G.P.; Teixeira, P.C.; da Silva, I.M.; Schipper, L.L.; Santana Filho, P.C.; Rodrigues Junior, L.C.; Bonorino, C.; Peres, A.; Fonseca, S.G.; Monteiro, M.C.; et al. Alterations in CD39/CD73 Axis of T Cells Associated with COVID-19 Severity. J. Cell. Physiol. 2022, 237, 3394–3407. [Google Scholar] [CrossRef]
- Halpin-Veszeleiova, K.; Hatfield, S.M. Therapeutic Targeting of Hypoxia-A2-Adenosinergic Pathway in COVID-19 Patients. Physiology 2022, 37, 46–52. [Google Scholar] [CrossRef]
- Correale, P.; Caracciolo, M.; Bilotta, F.; Conte, M.; Cuzzola, M.; Falcone, C.; Mangano, C.; Falzea, A.C.; Iuliano, E.; Morabito, A.; et al. Therapeutic Effects of Adenosine in High Flow 21% Oxygen Aereosol in Patients with Covid19-Pneumonia. PLoS ONE 2020, 15, e0239692. [Google Scholar] [CrossRef]
- Ngamsri, K.-C.; Fuhr, A.; Schindler, K.; Simelitidis, M.; Hagen, M.; Zhang, Y.; Gamper-Tsigaras, J.; Konrad, F.M. Sevoflurane Dampens Acute Pulmonary Inflammation via the Adenosine Receptor A2B and Heme Oxygenase-1. Cells 2022, 11, 1094. [Google Scholar] [CrossRef]
- Ren, Y.; Qi, L.; Zhang, L.; Xu, J.; Ma, J.; Lv, Y.; Zhang, Y.; Wu, R. Cupping Alleviates Lung Injury through the Adenosine/A2BAR Pathway. Heliyon 2022, 8, e12141. [Google Scholar] [CrossRef]
- Boo, H.-J.; Park, S.J.; Noh, M.; Min, H.-Y.; Jeong, L.S.; Lee, H.-Y. LJ-529, a Partial Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) Agonist and Adenosine A3 Receptor Agonist, Ameliorates Elastase-Induced Pulmonary Emphysema in Mice. Arch. Pharm. Res. 2020, 43, 540–552. [Google Scholar] [CrossRef]
- Sgambellone, S.; Marri, S.; Catarinicchia, S.; Pini, A.; Tosh, D.K.; Jacobson, K.A.; Masini, E.; Salvemini, D.; Lucarini, L. Adenosine A3 Receptor (A3AR) Agonist for the Treatment of Bleomycin-Induced Lung Fibrosis in Mice. Int. J. Mol. Sci. 2022, 23, 13300. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wu, Q.; Zhong, D.; Li, C.; Du, L. Caffeine Prevents Hyperoxia-Induced Lung Injury in Neonatal Mice through NLRP3 Inflammasome and NF-ΚB Pathway. Respir. Res. 2020, 21, 140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yu-Jing, L.; Ma, T. The Immunomodulatory Function of Adenosine in Sepsis. Front. Immunol. 2022, 13, 936547. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, D.C.; Viacava, P.R.; Ferreira, R.G.; Damaceno, M.A.; Piñeros, A.R.; Melo, P.H.; Donate, P.B.; Toller-Kawahisa, J.E.; Zoppi, D.; Veras, F.P.; et al. Sepsis Expands a CD39+ Plasmablast Population That Promotes Immunosuppression via Adenosine-Mediated Inhibition of Macrophage Antimicrobial Activity. Immunity 2021, 54, 2024–2041. [Google Scholar] [CrossRef] [PubMed]
- Ngamsri, K.-C.; Putri, R.A.; Jans, C.; Schindler, K.; Fuhr, A.; Zhang, Y.; Gamper-Tsigaras, J.; Ehnert, S.; Konrad, F.M. CXCR4 and CXCR7 Inhibition Ameliorates the Formation of Platelet-Neutrophil Complexes and Neutrophil Extracellular Traps through Adora2b Signaling. Int. J. Mol. Sci. 2021, 22, 13576. [Google Scholar] [CrossRef]
- Ngamsri, K.-C.; Fabian, F.; Fuhr, A.; Gamper-Tsigaras, J.; Straub, A.; Fecher, D.; Steinke, M.; Walles, H.; Reutershan, J.; Konrad, F.M. Sevoflurane Exerts Protective Effects in Murine Peritonitis-Induced Sepsis via Hypoxia-Inducible Factor 1α/Adenosine A2B Receptor Signaling. Anesthesiology 2021, 135, 136–150. [Google Scholar] [CrossRef]
- Yegutkin, G.G.; Boison, D. ATP and Adenosine Metabolism in Cancer: Exploitation for Therapeutic Gain. Pharmacol. Rev. 2022, 74, 797–822. [Google Scholar] [CrossRef]
- Zahavi, D.; Hodge, J.W. Targeting Immunosuppressive Adenosine Signaling: A Review of Potential Immunotherapy Combination Strategies. Int. J. Mol. Sci. 2023, 24, 8871. [Google Scholar] [CrossRef]
- Kang, C.; Liu, L.; Wu, C.; Li, L.; Jia, X.; Xie, W.; Chen, S.; Wu, X.; Zheng, H.; Liu, J.; et al. The Adenosinergic Machinery in Cancer: In-Tandem Insights from Basic Mechanisms to Therapy. Front. Immunol. 2023, 14, 1111369. [Google Scholar] [CrossRef]
- Xia, C.; Yin, S.; To, K.K.W.; Fu, L. CD39/CD73/A2AR Pathway and Cancer Immunotherapy. Mol. Cancer 2023, 22, 44. [Google Scholar] [CrossRef]
- Slaats, J.; Wagena, E.; Smits, D.; Berends, A.A.; Peters, E.; Bakker, G.-J.; van Erp, M.; Weigelin, B.; Adema, G.J.; Friedl, P. Adenosine A2a Receptor Antagonism Restores Additive Cytotoxicity by Cytotoxic T Cells in Metabolically Perturbed Tumors. Cancer Immunol. Res. 2022, 10, 1462–1474. [Google Scholar] [CrossRef] [PubMed]
- Mastelic-Gavillet, B.; Navarro Rodrigo, B.; Décombaz, L.; Wang, H.; Ercolano, G.; Ahmed, R.; Lozano, L.E.; Ianaro, A.; Derré, L.; Valerio, M.; et al. Adenosine Mediates Functional and Metabolic Suppression of Peripheral and Tumor-Infiltrating CD8+ T Cells. J. Immunother. Cancer 2019, 7, 257. [Google Scholar] [CrossRef]
- Arruga, F.; Serra, S.; Vitale, N.; Guerra, G.; Papait, A.; Gyau, B.B.; Tito, F.; Efremov, D.; Vaisitti, T.; Deaglio, S. Targeting the A2A Adenosine Receptor Counteracts Immunosuppression in Vivo in a Mouse Model of Chronic Lymphocytic Leukemia. Haematologica 2020, 106, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Y.; Chu, Y.; Li, Z.; Yu, X.; Huang, Z.; Xu, J.; Zheng, L. Tumor-Derived Adenosine Promotes Macrophage Proliferation in Human Hepatocellular Carcinoma. J. Hepatol. 2021, 74, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Borodovsky, A.; Barbon, C.M.; Wang, Y.; Ye, M.; Prickett, L.; Chandra, D.; Shaw, J.; Deng, N.; Sachsenmeier, K.; Clarke, J.D.; et al. Small Molecule AZD4635 Inhibitor of A2AR Signaling Rescues Immune Cell Function Including CD103+ Dendritic Cells Enhancing Anti-Tumor Immunity. J. Immunother. Cancer 2020, 8, e000417. [Google Scholar] [CrossRef]
- Nakamura, K.; Casey, M.; Oey, H.; Vari, F.; Stagg, J.; Gandhi, M.K.; Smyth, M.J. Targeting an Adenosine-Mediated “Don’t Eat Me Signal” Augments Anti-Lymphoma Immunity by Anti-CD20 Monoclonal Antibody. Leukemia 2020, 34, 2708–2721. [Google Scholar] [CrossRef]
- Allard, B.; Cousineau, I.; Allard, D.; Buisseret, L.; Pommey, S.; Chrobak, P.; Stagg, J. Adenosine A2a Receptor Promotes Lymphangiogenesis and Lymph Node Metastasis. Oncoimmunology 2019, 8, 1601481. [Google Scholar] [CrossRef]
- Sidders, B.; Zhang, P.; Goodwin, K.; O’Connor, G.; Russell, D.L.; Borodovsky, A.; Armenia, J.; McEwen, R.; Linghu, B.; Bendell, J.C.; et al. Adenosine Signaling Is Prognostic for Cancer Outcome and Has Predictive Utility for Immunotherapeutic Response. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2020, 26, 2176–2187. [Google Scholar] [CrossRef]
- Kamai, T.; Kijima, T.; Tsuzuki, T.; Nukui, A.; Abe, H.; Arai, K.; Yoshida, K.-I. Increased Expression of Adenosine 2A Receptors in Metastatic Renal Cell Carcinoma Is Associated with Poorer Response to Anti-Vascular Endothelial Growth Factor Agents and Anti-PD-1/Anti-CTLA4 Antibodies and Shorter Survival. Cancer Immunol. Immunother. 2021, 70, 2009–2021. [Google Scholar] [CrossRef]
- Bai, Y.; Zhang, X.; Zheng, J.; Liu, Z.; Yang, Z.; Zhang, X. Overcoming High Level Adenosine-Mediated Immunosuppression by DZD2269, a Potent and Selective A2aR Antagonist. J. Exp. Clin. Cancer Res. 2022, 41, 302. [Google Scholar] [CrossRef]
- Willingham, S.B.; Ho, P.Y.; Hotson, A.; Hill, C.; Piccione, E.C.; Hsieh, J.; Liu, L.; Buggy, J.J.; McCaffery, I.; Miller, R.A. A2AR Antagonism with CPI-444 Induces Antitumor Responses and Augments Efficacy to Anti–PD-(L)1 and Anti–CTLA-4 in Preclinical Models. Cancer Immunol. Res. 2018, 6, 1136–1149. [Google Scholar] [CrossRef] [PubMed]
- Fong, L.; Hotson, A.; Powderly, J.D.; Sznol, M.; Heist, R.S.; Choueiri, T.K.; George, S.; Hughes, B.G.M.; Hellmann, M.D.; Shepard, D.R.; et al. Adenosine A2A Receptor Blockade as an Immunotherapy for Treatment-Refractory Renal Cell Cancer. Cancer Discov. 2020, 10, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.A.; Bendell, J.C.; Falchook, G.S.; Bauer, T.M.; Drake, C.G.; Choe, J.H.; George, D.J.; Karlix, J.L.; Ulahannan, S.; Sachsenmeier, K.F.; et al. Phase Ia/b, Open-Label, Multicenter Study of AZD4635 (an Adenosine A2A Receptor Antagonist) as Monotherapy or Combined with Durvalumab, in Patients with Solid Tumors. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2022, 28, 4871–4884. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Ze, S.; Zhou, R.; Yang, X.; Wang, H.; Chai, X.; Fang, M.; Liu, M.; Wang, Y.; Lu, W.; et al. Discovery of Pyridinone Derivatives as Potent, Selective, and Orally Bioavailable Adenosine A2A Receptor Antagonists for Cancer Immunotherapy. J. Med. Chem. 2023, 66, 4734–4754. [Google Scholar] [CrossRef]
- Yu, F.; Zhu, C.; Ze, S.; Wang, H.; Yang, X.; Liu, M.; Xie, Q.; Lu, W.; Wang, Y. Design, Synthesis, and Bioevaluation of 2-Aminopteridin-7(8H)-One Derivatives as Novel Potent Adenosine A2A Receptor Antagonists for Cancer Immunotherapy. J. Med. Chem. 2022, 65, 4367–4386. [Google Scholar] [CrossRef]
- Zhang, J.; Luo, Z.; Duan, W.; Yang, K.; Ling, L.; Yan, W.; Liu, R.; Wüthrich, K.; Jiang, H.; Xie, C.; et al. Dual-Acting Antitumor Agents Targeting the A2A Adenosine Receptor and Histone Deacetylases: Design and Synthesis of 4-(Furan-2-Yl)-1H-Pyrazolo[3,4-d]Pyrimidin-6-Amine Derivatives. Eur. J. Med. Chem. 2022, 236, 114326. [Google Scholar] [CrossRef]
- Yan, W.; Ling, L.; Wu, Y.; Yang, K.; Liu, R.; Zhang, J.; Zhao, S.; Zhong, G.; Zhao, S.; Jiang, H.; et al. Structure-Based Design of Dual-Acting Compounds Targeting Adenosine A2A Receptor and Histone Deacetylase as Novel Tumor Immunotherapeutic Agents. J. Med. Chem. 2021, 64, 16573–16597. [Google Scholar] [CrossRef]
- Varano, F.; Catarzi, D.; Vincenzi, F.; Pasquini, S.; Pelletier, J.; Lopes Rangel Fietto, J.; Espindola Gelsleichter, N.; Sarlandie, M.; Guilbaud, A.; Sévigny, J.; et al. Structural Investigation on Thiazolo[5,4-d]Pyrimidines to Obtain Dual-Acting Blockers of CD73 and Adenosine A2A Receptor as Potential Antitumor Agents. Bioorg. Med. Chem. Lett. 2020, 30, 127067. [Google Scholar] [CrossRef]
- Yu, W.; Sun, J.; Wang, X.; Yu, S.; Yan, M.; Wang, F.; Liu, X. Boosting Cancer Immunotherapy via the Convenient A2AR Inhibition Using a Tunable Nanocatalyst with Light-Enhanced Activity. Adv. Mater. Deerfield Beach Fla 2022, 34, e2106967. [Google Scholar] [CrossRef]
- Wen, X.; Xiong, X.; Yang, G.; Xiao, W.; Hou, J.; Pan, T.; Hu, Y.; Zhou, S. A Macrophage Membrane-Coated Mesoporous Silica Nanoplatform Inhibiting Adenosine A2AR via in Situ Oxygen Supply for Immunotherapy. J. Control. Release 2023, 353, 535–548. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Xu, D.; Zang, J.; Zheng, X.; Zhao, Y.; Li, Y.; He, R.; Ruan, S.; Dong, H.; et al. Targeting the Negative Feedback of Adenosine-A2AR Metabolic Pathway by a Tailored Nanoinhibitor for Photothermal Immunotherapy. Adv. Sci. 2022, 9, 2104182. [Google Scholar] [CrossRef]
- Labanieh, L.; Mackall, C.L. CAR Immune Cells: Design Principles, Resistance and the next Generation. Nature 2023, 614, 635–648. [Google Scholar] [CrossRef]
- Li, N.; Tang, N.; Cheng, C.; Hu, T.; Wei, X.; Han, W.; Wang, H. Improving the Anti-Solid Tumor Efficacy of CAR-T Cells by Inhibiting Adenosine Signaling Pathway. Oncoimmunology 2020, 9, 1824643. [Google Scholar] [CrossRef] [PubMed]
- Giuffrida, L.; Sek, K.; Henderson, M.A.; Lai, J.; Chen, A.X.Y.; Meyran, D.; Todd, K.L.; Petley, E.V.; Mardiana, S.; Mølck, C.; et al. CRISPR/Cas9 Mediated Deletion of the Adenosine A2A Receptor Enhances CAR T Cell Efficacy. Nat. Commun. 2021, 12, 3236. [Google Scholar] [CrossRef] [PubMed]
- Masoumi, E.; Jafarzadeh, L.; Mirzaei, H.R.; Alishah, K.; Fallah-Mehrjardi, K.; Rostamian, H.; Khakpoor-Koosheh, M.; Meshkani, R.; Noorbakhsh, F.; Hadjati, J. Genetic and Pharmacological Targeting of A2a Receptor Improves Function of Anti-Mesothelin CAR T Cells. J. Exp. Clin. Cancer Res. 2020, 39, 49. [Google Scholar] [CrossRef] [PubMed]
- Seifert, M.; Benmebarek, M.-R.; Briukhovetska, D.; Märkl, F.; Dörr, J.; Cadilha, B.L.; Jobst, J.; Stock, S.; Andreu-Sanz, D.; Lorenzini, T.; et al. Impact of the Selective A2AR and A2BR Dual Antagonist AB928/Etrumadenant on CAR T Cell Function. Br. J. Cancer 2022, 127, 2175–2185. [Google Scholar] [CrossRef]
- Vecchio, E.A.; White, P.J.; May, L.T. The Adenosine A2B G Protein-Coupled Receptor: Recent Advances and Therapeutic Implications. Pharmacol. Ther. 2019, 198, 20–33. [Google Scholar] [CrossRef]
- Chen, S.; Akdemir, I.; Fan, J.; Linden, J.; Zhang, B.; Cekic, C. The Expression of Adenosine A2B Receptor on Antigen Presenting Cells Suppresses CD8+ T Cell Responses and Promotes Tumor Growth. Cancer Immunol. Res. 2020, 8, 1064–1074. [Google Scholar] [CrossRef]
- Lan, J.; Lu, H.; Samanta, D.; Salman, S.; Lu, Y.; Semenza, G.L. Hypoxia-Inducible Factor 1-Dependent Expression of Adenosine Receptor 2B Promotes Breast Cancer Stem Cell Enrichment. Proc. Natl. Acad. Sci. USA 2018, 115, E9640–E9648. [Google Scholar] [CrossRef]
- Lan, J.; Wei, G.; Liu, J.; Yang, F.; Sun, R.; Lu, H. Chemotherapy-Induced Adenosine A2B Receptor Expression Mediates Epigenetic Regulation of Pluripotency Factors and Promotes Breast Cancer Stemness. Theranostics 2022, 12, 2598–2612. [Google Scholar] [CrossRef]
- Tay, A.H.M.; Prieto-Díaz, R.; Neo, S.; Tong, L.; Chen, X.; Carannante, V.; Önfelt, B.; Hartman, J.; Haglund, F.; Majellaro, M.; et al. A2B Adenosine Receptor Antagonists Rescue Lymphocyte Activity in Adenosine-Producing Patient-Derived Cancer Models. J. Immunother. Cancer 2022, 10, e004592. [Google Scholar] [CrossRef]
- Yu, M.; Guo, G.; Huang, L.; Deng, L.; Chang, C.-S.; Achyut, B.R.; Canning, M.; Xu, N.; Arbab, A.S.; Bollag, R.J.; et al. CD73 on Cancer-Associated Fibroblasts Enhanced by the A2B-Mediated Feedforward Circuit Enforces an Immune Checkpoint. Nat. Commun. 2020, 11, 515. [Google Scholar] [CrossRef]
- Faraoni, E.Y.; Singh, K.; Chandra, V.; Le Roux, O.; Dai, Y.; Sahin, I.; O’Brien, B.J.; Strickland, L.N.; Li, L.; Vucic, E.; et al. CD73-Dependent Adenosine Signaling through Adora2b Drives Immunosuppression in Ductal Pancreatic Cancer. Cancer Res. 2023, 83, 1111–1127. [Google Scholar] [CrossRef]
- Ludwig, N.; Yerneni, S.S.; Azambuja, J.H.; Gillespie, D.G.; Menshikova, E.V.; Jackson, E.K.; Whiteside, T.L. Tumor-Derived Exosomes Promote Angiogenesis via Adenosine A2B Receptor Signaling. Angiogenesis 2020, 23, 599–610. [Google Scholar] [CrossRef]
- Zhou, Y.; Tong, L.; Chu, X.; Deng, F.; Tang, J.; Tang, Y.; Dai, Y. The Adenosine A1 Receptor Antagonist DPCPX Inhibits Tumor Progression via the ERK/JNK Pathway in Renal Cell Carcinoma. Cell. Physiol. Biochem. 2017, 43, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Mirza, A.; Basso, A.; Black, S.; Malkowski, M.; Kwee, L.; Patcher, J.A.; Lachowicz, J.E.; Wang, Y.; Liu, S. RNA Interference Targeting of A1 Receptor-Overexpressing Breast Carcinoma Cells Leads to Diminished Rates of Cell Proliferation and Induction of Apoptosis. Cancer Biol. Ther. 2005, 4, 1355–1360. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Kuang, X.; Zhang, Y.; Ye, Y.; Li, J.; Liang, L.; Xie, Z.; Weng, L.; Guo, J.; Li, H.; et al. ADORA1 Inhibition Promotes Tumor Immune Evasion by Regulating the ATF3-PD-L1 Axis. Cancer Cell 2020, 37, 324–339.e8. [Google Scholar] [CrossRef]
- Le, X.; Negrao, M.V.; Reuben, A.; Federico, L.; Diao, L.; McGrail, D.; Nilsson, M.; Robichaux, J.; Munoz, I.G.; Patel, S.; et al. Characterization of the Immune Landscape of EGFR-Mutant NSCLC Identifies CD73/Adenosine Pathway as a Potential Therapeutic Target. J. Thorac. Oncol. 2021, 16, 583–600. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Ng, K.T.-P.; Liu, J.; Yeung, W.-H.O.; Zhu, J.; Chiu, T.-L.S.; Liu, H.; Chen, Z.; Lo, C.-M.; Man, K. Plasmacytoid Dendritic Cells Recruited by HIF-1α/EADO/ADORA1 Signaling Induce Immunosuppression in Hepatocellular Carcinoma. Cancer Lett. 2021, 522, 80–92. [Google Scholar] [CrossRef]
- Merighi, S.; Battistello, E.; Giacomelli, L.; Varani, K.; Vincenzi, F.; Borea, P.A.; Gessi, S. Targeting A3 and A2A Adenosine Receptors in the Fight against Cancer. Expert Opin. Ther. Targets 2019, 23, 669–678. [Google Scholar] [CrossRef]
- Bi, J.; Zheng, C.; Zheng, X. Increased Expression of Adenosine A3 Receptor in Tumor-Infiltrating Natural Killer Cells. Cell. Mol. Immunol. 2021, 18, 496–497. [Google Scholar] [CrossRef] [PubMed]
- Stemmer, S.M.; Manojlovic, N.S.; Marinca, M.V.; Petrov, P.; Cherciu, N.; Ganea, D.; Ciuleanu, T.E.; Pusca, I.A.; Beg, M.S.; Purcell, W.T.; et al. Namodenoson in Advanced Hepatocellular Carcinoma and Child–Pugh B Cirrhosis: Randomized Placebo-Controlled Clinical Trial. Cancers 2021, 13, 187. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Pharmacological Action | Compound | Experimental Model | Species | Effects |
|---|---|---|---|---|
| A1AR agonists | benzyloxy-cyclopentyladenosine | Neuropathic pain | rat | Analgesia [16] |
| ENBA | Cerebral ischemia | rat | Microglia/macrophages proliferation reduction [66] | |
| Dual A1/A3 AR agonists | AST-004 | Cerebral ischemia | nonhuman primate | Ischemic damage reduction [71] |
| A1AR antagonists | DPCPX | Epilepsy | rat | Reversion of DBS impact on interictal epileptic discharges [78] |
| A1AR PAMs | MIPS521 | Neuropathic pain | rat | Analgesia [14] |
| A2AAR agonists | ATL313 | Neuropathic pain | rat | Anti-allodynia and anti-inflammation [18] |
| A2AAR antagonists | KW-6002 (istradefylline) | EAE model | mouse | Protection against T Cell Infiltration [58] |
| Chemotherapy-induced cognitive impairment | mouse | Reversion of cisplatin-induced neurotoxicity [72] | ||
| Retinal injury | mouse | Reduction of apoptosis of retinal ganglia cells [101] | ||
| mouse | Regulation of retinal ganglion cell morphology [102] | |||
| KW-6356 | PD | marmoset | Motor disability reversion [45] | |
| SCH58261 | SUDEP | mouse | Increase in theta and beta oscillations [79] | |
| Mania-like behavior | rat | Reduction of locomotor hyperactivity and risk-taking behavior [90] | ||
| ZM241385 | Craniocerebral trauma | mouse | Reduction of fear memory [62] | |
| A2AAR PAMs | A2ARPAM-1 | Mania-like behavior | mouse | Insomnia reduction [95] |
| A2BAR agonists | BAY60-6583 | Focal ischemia | rat | Reduced brain damage [69] |
| A3AR agonists | Cl-IB-MECA | Glaucoma | rat | Preservation of retinal ganglion cell [99] |
| IB-MECA | Chronic cerebral ischemia | mouse | Improved memory deficits [70] | |
| MRS5980 | Traumatic brain injury | mouse | Reduction of tissue injury and cognitive impairment [63] | |
| Chemotherapy-induced cognitive impairment | mouse | Prevention of mitochondrial dysfunction and oxidative stress [73] | ||
| Neuropathic pain | mouse | Anti-allodynia [23] | ||
| Non-selective antagonists | Caffeine | AD | mouse | Neuroprotection [41] |
| Pharmacological Action | Compound | Experimental Model | Species | Effects |
|---|---|---|---|---|
| A2AAR agonists | ATL1223 | Circulatory arrest and ECPR | pig | Reduction of systemic ischemia-reperfusion injury and ameliorated renal, hepatic, and cardiac injury [108] |
| A2AAR antagonists | SCH58261 | High-sucrose diet | rat | Improved insulin response [120] |
| A2BAR agonists | BAY 60–6583 | Obesity | mouse | Increased whole body energy expenditure [118] |
| A2BAR antagonists | MRS1754 | High-sucrose diet | rat | Improved insulin response [120] |
| Pharmacological Action | Compound | Experimental Model | Species | Effects |
|---|---|---|---|---|
| A2AAR agonists | CGS21680 | Arthritis | mouse | Arthritis development inhibition [132] |
| LES | mouse | Depletion of pathogenic lymphocytes and anti-nuclear antibodies reduction [135] | ||
| Obesity-induced OA | mouse | Senescence markers reduction and improvement of mitochondrial stability and function [141,142] | ||
| Increased autophagy and improved metabolic function in chondrocytes [143] | ||||
| PDRN | Acute lung injury | rat | Decrease in lung tissue damage, pro-inflammatory cytokines, apoptotic factors, and MAPK/NF-κB activation [152] | |
| A2AAR antagonists | ZM241385 | Sepsis | mouse | Enhanced survival by improving bacterial clearance [164] |
| A3AR agonists | CF101 | OA | rat | Reduction of cartilage damage and pain [144] |
| MRS5980 | Lung fibrosis | mouse | Attenuation of lung stiffness and inflammatory/oxidative stress markers [161] | |
| MRS7344 | Psoriasis | mouse | Hinder development of psoriatic-like characteristics [126] | |
| A3AR/partial PPARγ agonist | LJ-529 | Pulmonary Emphysema | mouse | Pulmonary function restoration, reduction of MMP activity and apoptosis [160] |
| Non-selective agonists | NECA | Airway inflammation | mouse | Reduction of innate lymphoid cells responses and inflammation [151] |
| Non-selective antagonists | Caffeine | Bronchopulmonary dysplasia | mouse | Decrease in lung injury, oxidative stress, and inflammatory infiltration [162] |
| Pharmacological Action | Compound | Experimental Model | Species985 | Effects |
|---|---|---|---|---|
| A1AR antagonists | DPCPX | HCC | mouse | Suppression of tumor growth [209] |
| A2AAR antagonists | AZD4635 | Melanoma, colorectal carcinoma, fibrosarcoma | mouse | Decrease in tumor volume with enhanced T cell response [175] |
| CPI-444 (ciforadenant) | Colorectal carcinoma, kidney tumor, melanoma | mouse | Diminution of tumor growth intensifying the effectiveness of checkpoint inhibitors [181] | |
| DZD2269 | Melanoma, prostate cancer, and pancreatic cancer | mouse | Antitumor effects particularly when used together with immune checkpoint inhibitors, radiotherapy, or chemotherapy [180] | |
| SCH58261 | CLL | mouse | Immune competence restoration by inhibiting the accumulation and differentiation of Treg cells [173] | |
| Breast cancer | mouse | Regression of primary tumor and inhibition of metastasis [191] | ||
| ZM241385 | Melanoma | mouse | Restoration of the functionality of tumor-infiltrating cytotoxic T cells [171] | |
| Dual A2A/A2B AR antagonists | AB928 (etrumadenant) | Colon carcinoma | mouse | Enhancement of CAR T cell activation [196] |
| A2BAR antagonists | Alloxazine | Breast cancer | mouse | Delay of tumor recurrence after discontinuation of chemotherapy [200] |
| Pharmacological Action | Compound | Condition | Phase | NTC Number |
|---|---|---|---|---|
| A1 antagonists | PBF-680 | Asthma | II | 03774290 |
| COPD | IIa | 05262218 | ||
| A2A agonists | Regadenoson | COVID-19 | I/II | 04606069 |
| Myocardial ischemia | I/II | 04604782 | ||
| High grade gliomas | I | 03971734 | ||
| Lung transplant | I | 04521569 | ||
| A2A antagonists | Istradefylline | ALS | I/II | 05377424 |
| PD | IV | 05885360 | ||
| Cognitive impairment in PD | II | 05333549 | ||
| Apathy in PD | Observational | 05182151 | ||
| Ciforadenant (CPI-444) | Renal cell carcinoma | Ib/II | 05501054 | |
| Multiple myeloma | I | 04280328 | ||
| TT-10 | Solid cancers | I/II | 04969315 | |
| Inupadenant (EOS100850) | NSCLC | II | 05403385 | |
| Solid cancers | I | 05117177 | ||
| AZD4635 | Solid cancers | I | 03980821 | |
| NSCLC | I/II | 03381274 | ||
| Prostate cancer | II | 04089553 | ||
| CRPC | II | 04495179 | ||
| DZD2269 | CRPC | I | 04634344 | |
| Dual A2A/A2B antagonists | Etrumadenant (AB928) | Head and neck cancers | I | 04892875 |
| Prostate cancer | II | 05915442 | ||
| Liposarcoma | II | 05886634 | ||
| Rectal cancer | II | 05024097 | ||
| Urothelial carcinoma | II | 05335941 | ||
| Gastrointestinal cancers | I | 03720678 | ||
| Colorectal cancer | I/II | 04660812 | ||
| CRPC | Ib/II | 04381832 | ||
| CRPC | II | 05177770 | ||
| M1069 | Solid cancers | I | 05198349 | |
| A2B antagonists | PBF-1129 | NSCLC | I | 03274479 |
| NSCLC | I | 05234307 | ||
| TT-702 (prodrug of TT-478) | Solid cancers | I/II | 05272709 | |
| TT-4 | Solid cancers | I/II | 04976660 | |
| A3 agonists | Piclidenoson (CF101, IB-MECA) | Ocular hypertension | I/II | 04585100 |
| COVID-19 | II | 04333472 | ||
| Plaque psoriasis | III | 03168256 | ||
| Namodenoson (CF102, Cl-IB-MECA) | NASH | II | 04697810 | |
| HCC/Cirrhosis | III | 05201404 | ||
| FM-101 | Ocular hypertension | I/II | 04585100 | |
| NASH | II | 04710524 | ||
| A3 antagonists | PBF-1650 | Psoriasis | I | 03798236 |
| PBF-677 | Ulcerative colitis | II | 03773952 | |
| Non-selective antagonists | Caffeine | Hypoxic-ischemic encephalopathy | I | 03913221 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vincenzi, F.; Pasquini, S.; Contri, C.; Cappello, M.; Nigro, M.; Travagli, A.; Merighi, S.; Gessi, S.; Borea, P.A.; Varani, K. Pharmacology of Adenosine Receptors: Recent Advancements. Biomolecules 2023, 13, 1387. https://doi.org/10.3390/biom13091387
Vincenzi F, Pasquini S, Contri C, Cappello M, Nigro M, Travagli A, Merighi S, Gessi S, Borea PA, Varani K. Pharmacology of Adenosine Receptors: Recent Advancements. Biomolecules. 2023; 13(9):1387. https://doi.org/10.3390/biom13091387
Chicago/Turabian StyleVincenzi, Fabrizio, Silvia Pasquini, Chiara Contri, Martina Cappello, Manuela Nigro, Alessia Travagli, Stefania Merighi, Stefania Gessi, Pier Andrea Borea, and Katia Varani. 2023. "Pharmacology of Adenosine Receptors: Recent Advancements" Biomolecules 13, no. 9: 1387. https://doi.org/10.3390/biom13091387