
A Lysine Residue at the C-Terminus of MHC Class I Ligands Correlates with Low C-Terminal Proteasomal Cleavage Probability
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Line
2.2. MHC Class I Immunoprecipitation and Peptide Elution
2.3. LC-MS/MS and Quantitative Analysis
2.4. Cleavage Probability Prediction
2.5. Statistical Analysis
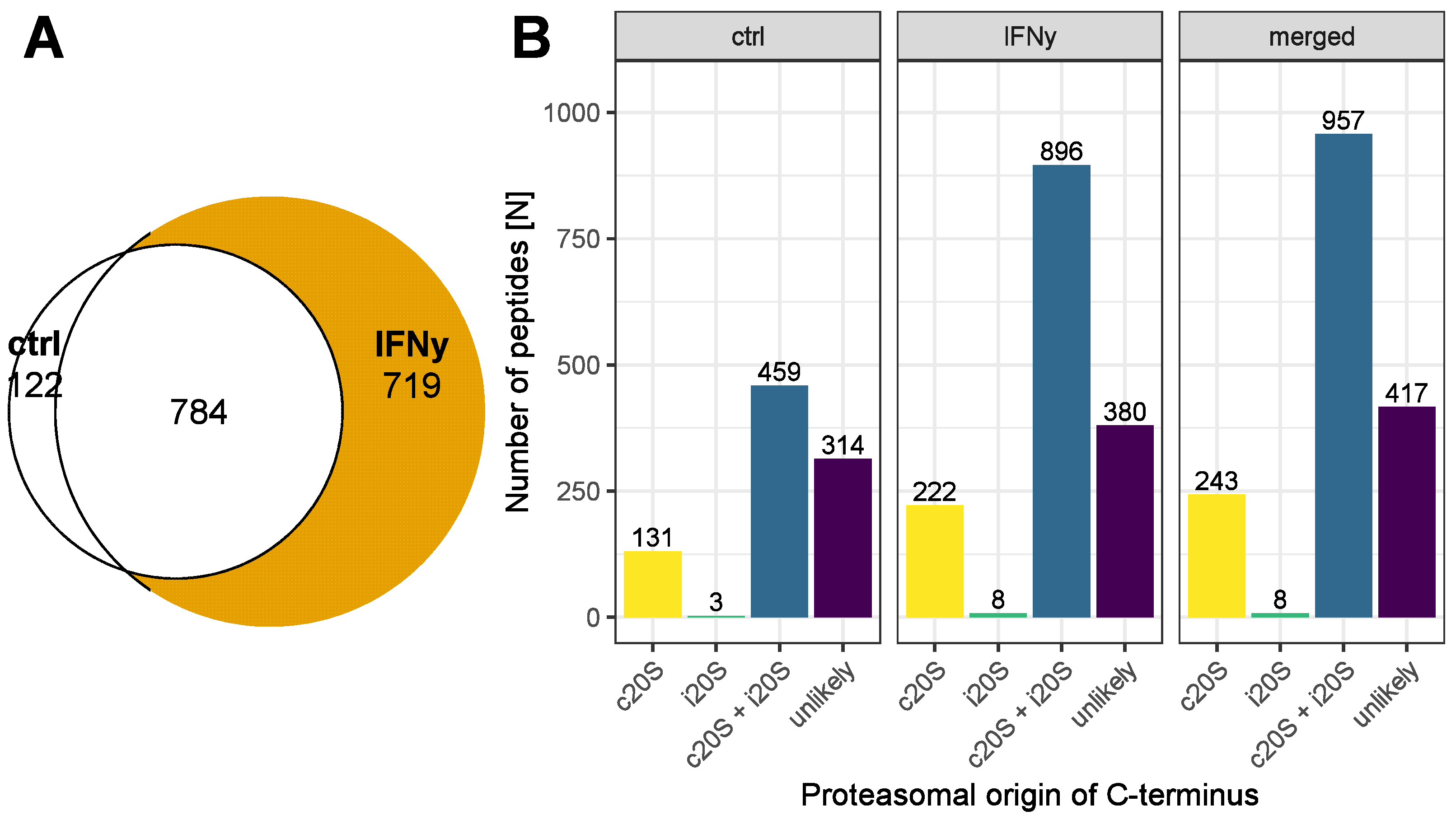
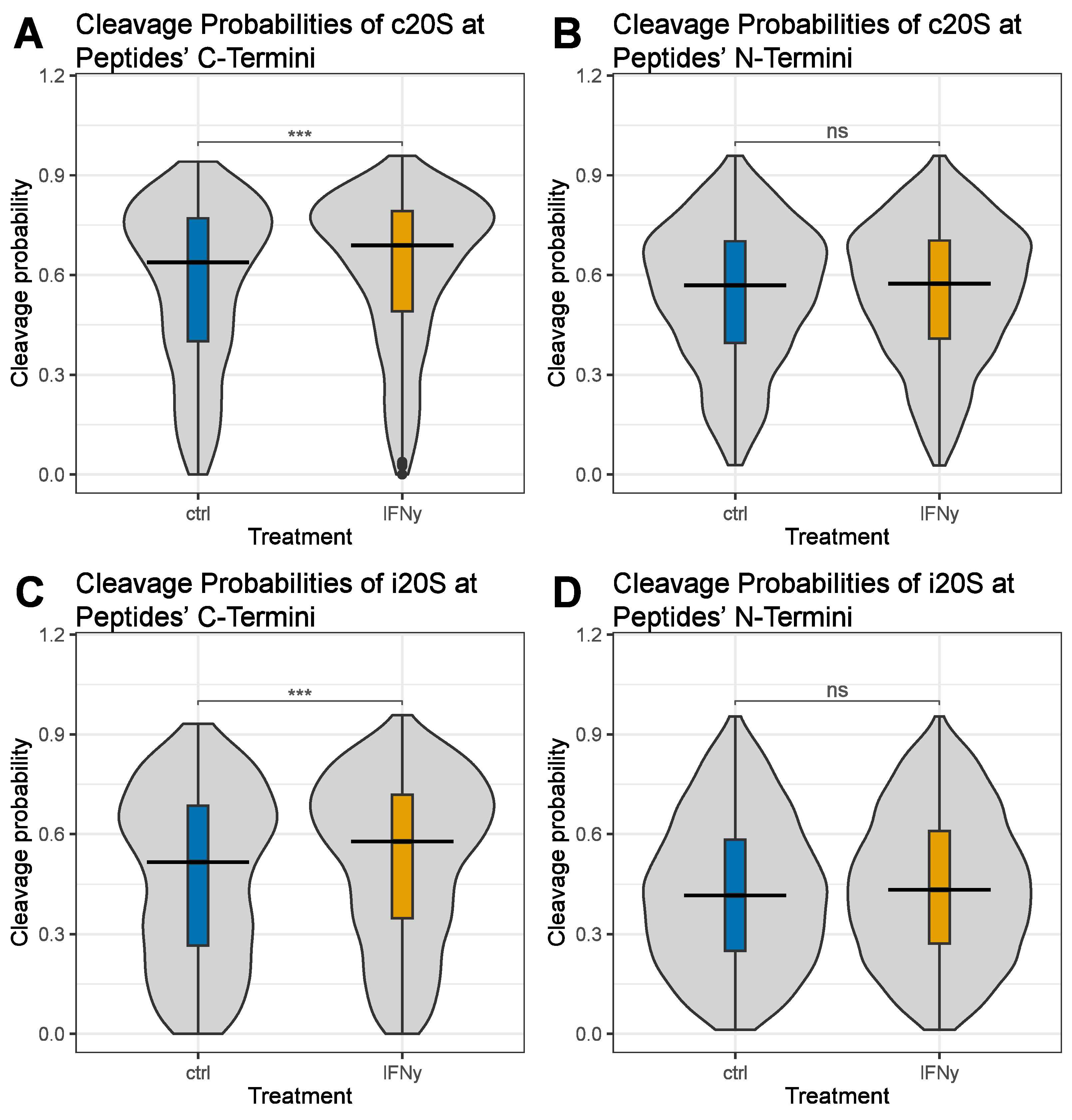
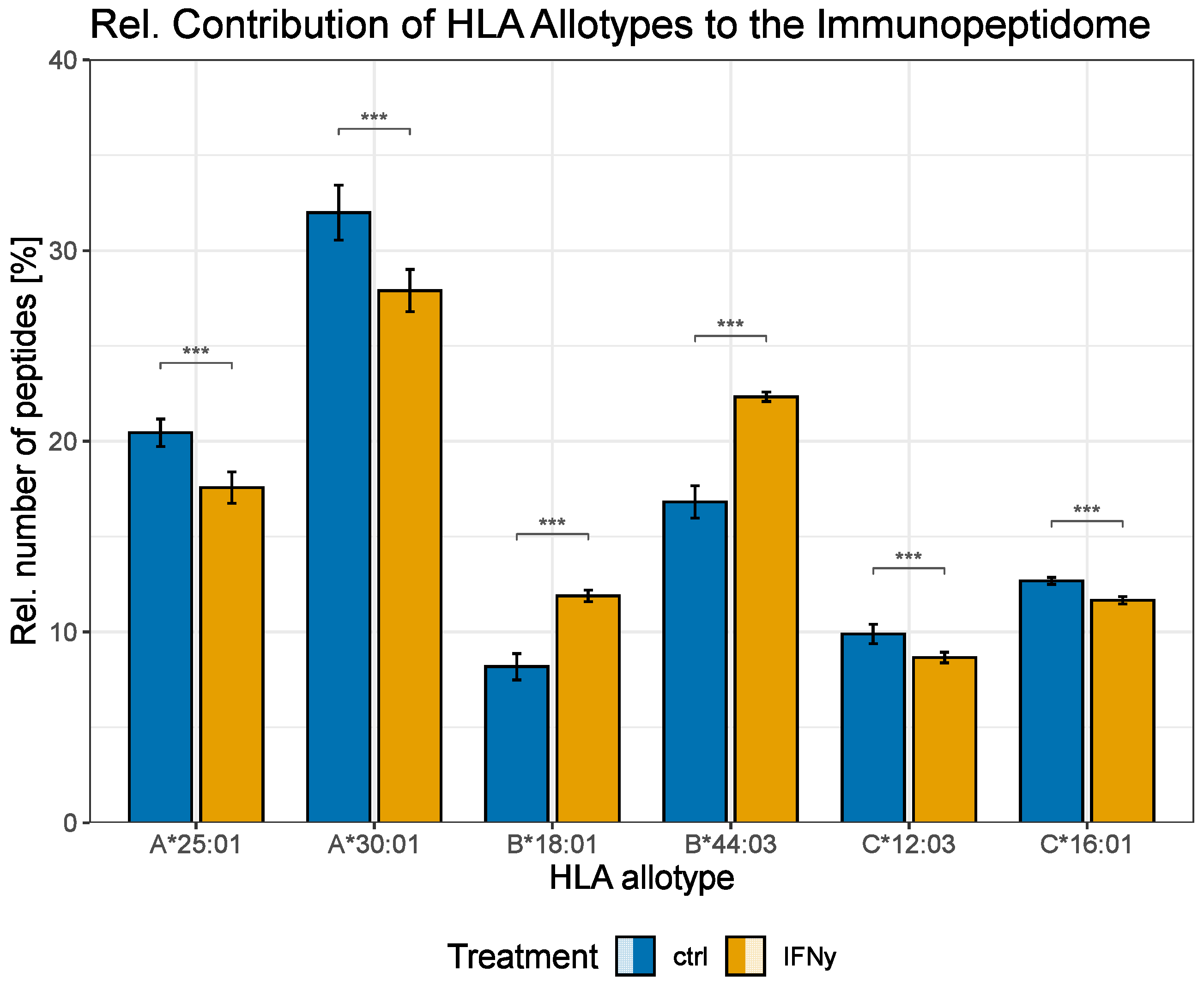
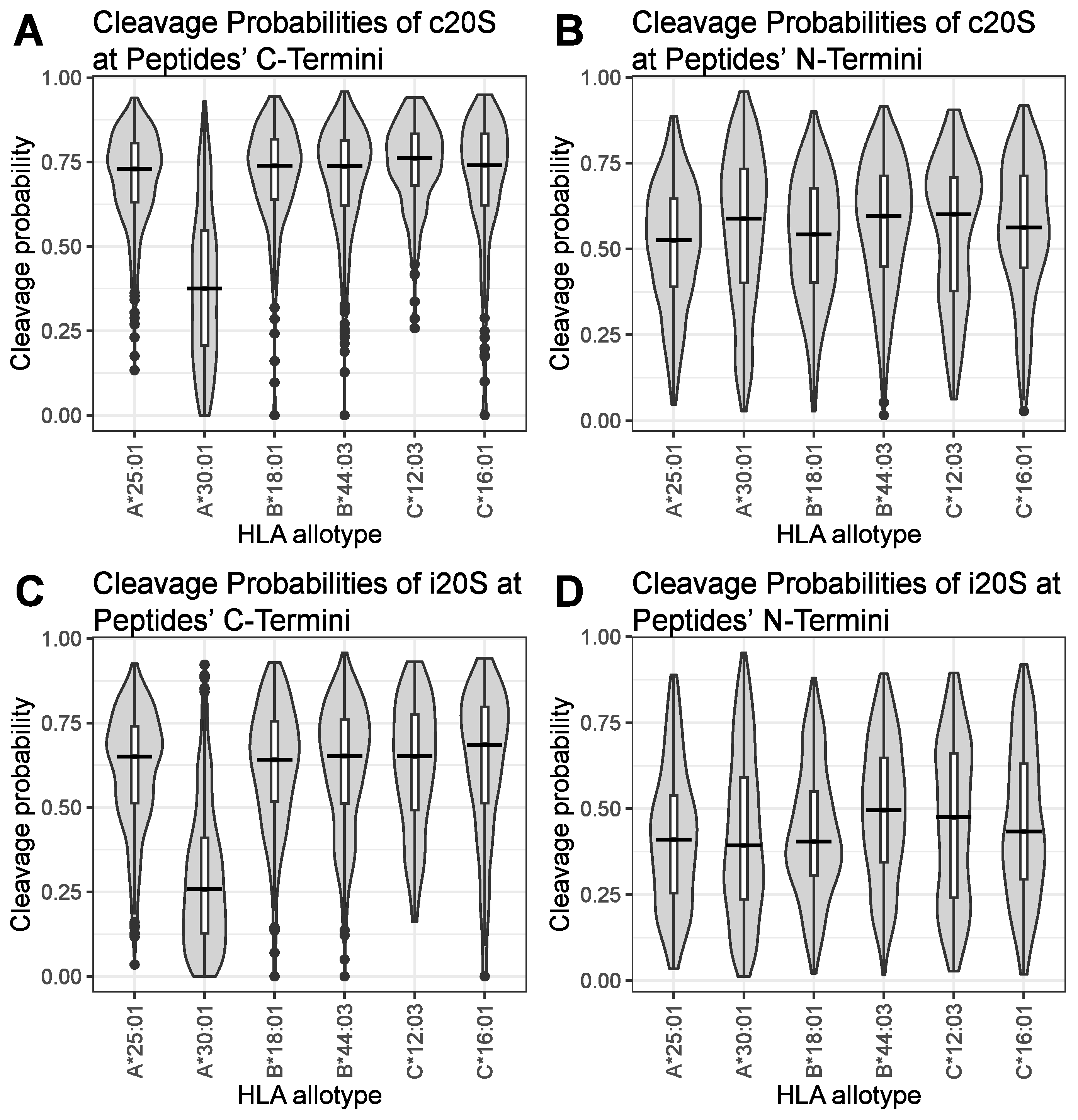
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rock, K.L.; Gramm, C.; Rothstein, L.; Clark, K.; Stein, R.; Dick, L.; Hwang, D.; Goldberg, A.L. Inhibitors of the proteasome block the degradation of most cell proteins and the generation of peptides presented on MHC class I molecules. Cell 1994, 78, 761–771. [Google Scholar] [CrossRef] [PubMed]
- Abildgaard, A.B.; Gersing, S.K.; Larsen-Ledet, S.; Nielsen, S.V.; Stein, A.; Lindorff-Larsen, K.; Hartmann-Petersen, R. Co-Chaperones in Targeting and Delivery of Misfolded Proteins to the 26S Proteasome. Biomolecules 2020, 10, 1141. [Google Scholar] [CrossRef] [PubMed]
- Erhard, F.; Halenius, A.; Zimmermann, C.; L'Hernault, A.; Kowalewski, D.J.; Weekes, M.P.; Stevanovic, S.; Zimmer, R.; Dolken, L. Improved Ribo-seq enables identification of cryptic translation events. Nat. Methods 2018, 15, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Bjorkman, P.J.; Saper, M.A.; Samraoui, B.; Bennett, W.S.; Strominger, J.L.; Wiley, D.C. Structure of the human class I histocompatibility antigen, HLA-A2. Nature 1987, 329, 506–512. [Google Scholar] [CrossRef]
- Babbitt, B.P.; Allen, P.M.; Matsueda, G.; Haber, E.; Unanue, E.R. Binding of immunogenic peptides to Ia histocompatibility molecules. Nature 1985, 317, 359–361. [Google Scholar] [CrossRef]
- Anjanappa, R.; Garcia-Alai, M.; Kopicki, J.D.; Lockhauserbaumer, J.; Aboelmagd, M.; Hinrichs, J.; Nemtanu, I.M.; Uetrecht, C.; Zacharias, M.; Springer, S.; et al. Structures of peptide-free and partially loaded MHC class I molecules reveal mechanisms of peptide selection. Nat. Commun. 2020, 11, 1314. [Google Scholar] [CrossRef]
- Dockree, T.; Holland, C.J.; Clement, M.; Ladell, K.; McLaren, J.E.; van den Berg, H.A.; Gostick, E.; Miners, K.L.; Llewellyn-Lacey, S.; Bridgeman, J.S.; et al. CD8(+) T-cell specificity is compromised at a defined MHCI/CD8 affinity threshold. Immunol. Cell Biol. 2017, 95, 68–76. [Google Scholar] [CrossRef]
- Toes, R.E.; Nussbaum, A.K.; Degermann, S.; Schirle, M.; Emmerich, N.P.; Kraft, M.; Laplace, C.; Zwinderman, A.; Dick, T.P.; Muller, J.; et al. Discrete cleavage motifs of constitutive and immunoproteasomes revealed by quantitative analysis of cleavage products. J. Exp. Med. 2001, 194, 1–12. [Google Scholar] [CrossRef]
- Groll, M.; Ditzel, L.; Lowe, J.; Stock, D.; Bochtler, M.; Bartunik, H.D.; Huber, R. Structure of 20S proteasome from yeast at 2.4 A resolution. Nature 1997, 386, 463–471. [Google Scholar] [CrossRef]
- Kisselev, A.F.; Callard, A.; Goldberg, A.L. Importance of the different proteolytic sites of the proteasome and the efficacy of inhibitors varies with the protein substrate. J. Biol. Chem. 2006, 281, 8582–8590. [Google Scholar] [CrossRef]
- Kessler, J.H.; Khan, S.; Seifert, U.; Le Gall, S.; Chow, K.M.; Paschen, A.; Bres-Vloemans, S.A.; de Ru, A.; van Montfoort, N.; Franken, K.L.; et al. Antigen processing by nardilysin and thimet oligopeptidase generates cytotoxic T cell epitopes. Nat. Immunol. 2011, 12, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Seifert, U.; Maranon, C.; Shmueli, A.; Desoutter, J.F.; Wesoloski, L.; Janek, K.; Henklein, P.; Diescher, S.; Andrieu, M.; de la Salle, H.; et al. An essential role for tripeptidyl peptidase in the generation of an MHC class I epitope. Nat. Immunol. 2003, 4, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, N.; Stroobant, V.; Colau, D.; de Diesbach, P.; Morel, S.; Chapiro, J.; van Endert, P.; Van den Eynde, B.J. Production of an antigenic peptide by insulin-degrading enzyme. Nat. Immunol. 2010, 11, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Aki, M.; Shimbara, N.; Takashina, M.; Akiyama, K.; Kagawa, S.; Tamura, T.; Tanahashi, N.; Yoshimura, T.; Tanaka, K.; Ichihara, A. Interferon-gamma induces different subunit organizations and functional diversity of proteasomes. J. Biochem. 1994, 115, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Matthews, W.; Driscoll, J.; Tanaka, K.; Ichihara, A.; Goldberg, A.L. Involvement of the proteasome in various degradative processes in mammalian cells. Proc. Natl. Acad. Sci. USA 1989, 86, 2597–2601. [Google Scholar] [CrossRef]
- Tomaru, U.; Ishizu, A.; Murata, S.; Miyatake, Y.; Suzuki, S.; Takahashi, S.; Kazamaki, T.; Ohara, J.; Baba, T.; Iwasaki, S.; et al. Exclusive expression of proteasome subunit beta5t in the human thymic cortex. Blood 2009, 113, 5186–5191. [Google Scholar] [CrossRef]
- Javitt, A.; Barnea, E.; Kramer, M.P.; Wolf-Levy, H.; Levin, Y.; Admon, A.; Merbl, Y. Pro-inflammatory Cytokines Alter the Immunopeptidome Landscape by Modulation of HLA-B Expression. Front. Immunol. 2019, 10, 141. [Google Scholar] [CrossRef]
- Chen, S.; Kammerl, I.E.; Vosyka, O.; Baumann, T.; Yu, Y.; Wu, Y.; Irmler, M.; Overkleeft, H.S.; Beckers, J.; Eickelberg, O.; et al. Immunoproteasome dysfunction augments alternative polarization of alveolar macrophages. Cell Death Differ. 2016, 23, 1026–1037. [Google Scholar] [CrossRef]
- Liepe, J.; Marino, F.; Sidney, J.; Jeko, A.; Bunting, D.E.; Sette, A.; Kloetzel, P.M.; Stumpf, M.P.; Heck, A.J.; Mishto, M. A large fraction of HLA class I ligands are proteasome-generated spliced peptides. Science 2016, 354, 354–358. [Google Scholar] [CrossRef]
- van Endert, P.M.; Tampe, R.; Meyer, T.H.; Tisch, R.; Bach, J.F.; McDevitt, H.O. A sequential model for peptide binding and transport by the transporters associated with antigen processing. Immunity 1994, 1, 491–500. [Google Scholar] [CrossRef]
- Suh, W.K.; Cohen-Doyle, M.F.; Fruh, K.; Wang, K.; Peterson, P.A.; Williams, D.B. Interaction of MHC class I molecules with the transporter associated with antigen processing. Science 1994, 264, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Blees, A.; Januliene, D.; Hofmann, T.; Koller, N.; Schmidt, C.; Trowitzsch, S.; Moeller, A.; Tampe, R. Structure of the human MHC-I peptide-loading complex. Nature 2017, 551, 525–528. [Google Scholar] [CrossRef] [PubMed]
- York, I.A.; Chang, S.C.; Saric, T.; Keys, J.A.; Favreau, J.M.; Goldberg, A.L.; Rock, K.L. The ER aminopeptidase ERAP1 enhances or limits antigen presentation by trimming epitopes to 8-9 residues. Nat. Immunol. 2002, 3, 1177–1184. [Google Scholar] [CrossRef]
- Saveanu, L.; Carroll, O.; Lindo, V.; Del Val, M.; Lopez, D.; Lepelletier, Y.; Greer, F.; Schomburg, L.; Fruci, D.; Niedermann, G.; et al. Concerted peptide trimming by human ERAP1 and ERAP2 aminopeptidase complexes in the endoplasmic reticulum. Nat. Immunol. 2005, 6, 689–697. [Google Scholar] [CrossRef]
- Cascio, P.; Hilton, C.; Kisselev, A.F.; Rock, K.L.; Goldberg, A.L. 26S proteasomes and immunoproteasomes produce mainly N-extended versions of an antigenic peptide. EMBO J. 2001, 20, 2357–2366. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Akopian, T.N.; Woo, K.M.; Goldberg, A.L. The sizes of peptides generated from protein by mammalian 26 and 20 S proteasomes. Implications for understanding the degradative mechanism and antigen presentation. J. Biol. Chem. 1999, 274, 3363–3371. [Google Scholar] [CrossRef]
- Purcell, A.W.; Ramarathinam, S.H.; Ternette, N. Mass spectrometry-based identification of MHC-bound peptides for immunopeptidomics. Nat. Protoc. 2019, 14, 1687–1707. [Google Scholar] [CrossRef]
- Marino, F.; Chong, C.; Michaux, J.; Bassani-Sternberg, M. High-Throughput, Fast, and Sensitive Immunopeptidomics Sample Processing for Mass Spectrometry. Methods Mol. Biol. 2019, 1913, 67–79. [Google Scholar] [CrossRef]
- Weeder, B.R.; Wood, M.A.; Li, E.; Nellore, A.; Thompson, R.F. pepsickle rapidly and accurately predicts proteasomal cleavage sites for improved neoantigen identification. Bioinformatics 2021, 37, 3723–3733. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Schmalen, A.; Haines, S.; Wolff, M.; Ma, H.; Zhang, H.; Stoleriu, M.G.; Nowak, J.; Nakayama, M.; et al. Antiviral CD8(+) T cell immune responses are impaired by cigarette smoke and in COPD. Eur. Respir. J. 2023, 62, 2201374. [Google Scholar] [CrossRef]
- Zolg, D.P.; Gessulat, S.; Paschke, C.; Graber, M.; Rathke-Kuhnert, M.; Seefried, F.; Fitzemeier, K.; Berg, F.; Lopez-Ferrer, D.; Horn, D.; et al. INFERYS rescoring: Boosting peptide identifications and scoring confidence of database search results. Rapid Commun. Mass Spectrom. 2021, e9128. [Google Scholar] [CrossRef] [PubMed]
- Kall, L.; Canterbury, J.D.; Weston, J.; Noble, W.S.; MacCoss, M.J. Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat. Methods 2007, 4, 923–925. [Google Scholar] [CrossRef] [PubMed]
- Bassani-Sternberg, M.; Chong, C.; Guillaume, P.; Solleder, M.; Pak, H.; Gannon, P.O.; Kandalaft, L.E.; Coukos, G.; Gfeller, D. Deciphering HLA-I motifs across HLA peptidomes improves neo-antigen predictions and identifies allostery regulating HLA specificity. PLoS Comput. Biol. 2017, 13, e1005725. [Google Scholar] [CrossRef]
- Gfeller, D.; Guillaume, P.; Michaux, J.; Pak, H.S.; Daniel, R.T.; Racle, J.; Coukos, G.; Bassani-Sternberg, M. The Length Distribution and Multiple Specificity of Naturally Presented HLA-I Ligands. J. Immunol. 2018, 201, 3705–3716. [Google Scholar] [CrossRef]
- Andreatta, M.; Lund, O.; Nielsen, M. Simultaneous alignment and clustering of peptide data using a Gibbs sampling approach. Bioinformatics 2013, 29, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Andreatta, M.; Alvarez, B.; Nielsen, M. GibbsCluster: Unsupervised clustering and alignment of peptide sequences. Nucleic Acids Res. 2017, 45, W458–W463. [Google Scholar] [CrossRef] [PubMed]
- Wagih, O. ggseqlogo: A versatile R package for drawing sequence logos. Bioinformatics 2017, 33, 3645–3647. [Google Scholar] [CrossRef]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef]
- Benjamini, Y.; Drai, D.; Elmer, G.; Kafkafi, N.; Golani, I. Controlling the false discovery rate in behavior genetics research. Behav. Brain Res. 2001, 125, 279–284. [Google Scholar] [CrossRef]
- Winter, M.B.; La Greca, F.; Arastu-Kapur, S.; Caiazza, F.; Cimermancic, P.; Buchholz, T.J.; Anderl, J.L.; Ravalin, M.; Bohn, M.F.; Sali, A.; et al. Immunoproteasome functions explained by divergence in cleavage specificity and regulation. Elife 2017, 6, e27364. [Google Scholar] [CrossRef]
- Keller, I.E.; Vosyka, O.; Takenaka, S.; Kloss, A.; Dahlmann, B.; Willems, L.I.; Verdoes, M.; Overkleeft, H.S.; Marcos, E.; Adnot, S.; et al. Regulation of immunoproteasome function in the lung. Sci. Rep. 2015, 5, 10230. [Google Scholar] [CrossRef]
- Schatz, M.M.; Peters, B.; Akkad, N.; Ullrich, N.; Martinez, A.N.; Carroll, O.; Bulik, S.; Rammensee, H.G.; van Endert, P.; Holzhutter, H.G.; et al. Characterizing the N-terminal processing motif of MHC class I ligands. J. Immunol. 2008, 180, 3210–3217. [Google Scholar] [CrossRef]
- Tadros, D.M.; Eggenschwiler, S.; Racle, J.; Gfeller, D. The MHC Motif Atlas: A database of MHC binding specificities and ligands. Nucleic Acids Res. 2023, 51, D428–D437. [Google Scholar] [CrossRef]
- Gomez-Perosanz, M.; Ras-Carmona, A.; Lafuente, E.M.; Reche, P.A. Identification of CD8(+) T cell epitopes through proteasome cleavage site predictions. BMC Bioinform. 2020, 21, 484. [Google Scholar] [CrossRef]
- McSparron, H.; Blythe, M.J.; Zygouri, C.; Doytchinova, I.A.; Flower, D.R. JenPep: A novel computational information resource for immunobiology and vaccinology. J. Chem. Inf. Comput. Sci. 2003, 43, 1276–1287. [Google Scholar] [CrossRef]
- Rammensee, H.; Bachmann, J.; Emmerich, N.P.; Bachor, O.A.; Stevanovic, S. SYFPEITHI: Database for MHC ligands and peptide motifs. Immunogenetics 1999, 50, 213–219. [Google Scholar] [CrossRef]
- Rozanov, D.V.; Rozanov, N.D.; Chiotti, K.E.; Reddy, A.; Wilmarth, P.A.; David, L.L.; Cha, S.W.; Woo, S.; Pevzner, P.; Bafna, V.; et al. MHC class I loaded ligands from breast cancer cell lines: A potential HLA-I-typed antigen collection. J. Proteom. 2018, 176, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Bassani-Sternberg, M.; Braunlein, E.; Klar, R.; Engleitner, T.; Sinitcyn, P.; Audehm, S.; Straub, M.; Weber, J.; Slotta-Huspenina, J.; Specht, K.; et al. Direct identification of clinically relevant neoepitopes presented on native human melanoma tissue by mass spectrometry. Nat. Commun. 2016, 7, 13404. [Google Scholar] [CrossRef] [PubMed]
- Zanker, D.; Chen, W. Standard and immunoproteasomes show similar peptide degradation specificities. Eur. J. Immunol. 2014, 44, 3500–3503. [Google Scholar] [CrossRef]
- Basler, M.; Mundt, S.; Groettrup, M. The immunoproteasome subunit LMP7 is required in the murine thymus for filling up a hole in the T cell repertoire. Eur. J. Immunol. 2018, 48, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, G.; Mullan, K.A.; Duscharla, D.; Ayala, R.; Croft, N.P.; Faridi, P.; Purcell, A.W. IFNgamma Modulates the Immunopeptidome of Triple Negative Breast Cancer Cells by Enhancing and Diversifying Antigen Processing and Presentation. Front. Immunol. 2021, 12, 645770. [Google Scholar] [CrossRef]
- Dressler, F.F.; Bragelmann, J.; Reischl, M.; Perner, S. Normics: Proteomic Normalization by Variance and Data-Inherent Correlation Structure. Mol. Cell Proteom. 2022, 21, 100269. [Google Scholar] [CrossRef]
- Lopez-Larrea, C.; Sujirachato, K.; Mehra, N.K.; Chiewsilp, P.; Isarangkura, D.; Kanga, U.; Dominguez, O.; Coto, E.; Pena, M.; Setien, F.; et al. HLA-B27 subtypes in Asian patients with ankylosing spondylitis. Evidence for new associations. Tissue Antigens 1995, 45, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Hermann, E.; Yu, D.T.; Meyer zum Buschenfelde, K.H.; Fleischer, B. HLA-B27-restricted CD8 T cells derived from synovial fluids of patients with reactive arthritis and ankylosing spondylitis. Lancet 1993, 342, 646–650. [Google Scholar] [CrossRef]
- Al-Ani, H.H.; Sims, J.L.; Niederer, R.L. Long term complications and vision loss in HLA-B27 uveitis. Eye 2022, 37, 1673–1677. [Google Scholar] [CrossRef]
- Bowness, P. HLA B27 in health and disease: A double-edged sword? Rheumatology 2002, 41, 857–868. [Google Scholar] [CrossRef]
- Urban, R.G.; Chicz, R.M.; Lane, W.S.; Strominger, J.L.; Rehm, A.; Kenter, M.J.; UytdeHaag, F.G.; Ploegh, H.; Uchanska-Ziegler, B.; Ziegler, A. A subset of HLA-B27 molecules contains peptides much longer than nonamers. Proc. Natl. Acad. Sci. USA 1994, 91, 1534–1538. [Google Scholar] [CrossRef]
- Mobbs, J.I.; Illing, P.T.; Dudek, N.L.; Brooks, A.G.; Baker, D.G.; Purcell, A.W.; Rossjohn, J.; Vivian, J.P. The molecular basis for peptide repertoire selection in the human leucocyte antigen (HLA) C*06:02 molecule. J. Biol. Chem. 2017, 292, 17203–17215. [Google Scholar] [CrossRef] [PubMed]
- Trolle, T.; McMurtrey, C.P.; Sidney, J.; Bardet, W.; Osborn, S.C.; Kaever, T.; Sette, A.; Hildebrand, W.H.; Nielsen, M.; Peters, B. The Length Distribution of Class I-Restricted T Cell Epitopes Is Determined by Both Peptide Supply and MHC Allele-Specific Binding Preference. J. Immunol. 2016, 196, 1480–1487. [Google Scholar] [CrossRef] [PubMed]
- Grice, G.L.; Nathan, J.A. The recognition of ubiquitinated proteins by the proteasome. Cell. Mol. Life Sci. 2016, 73, 3497–3506. [Google Scholar] [CrossRef] [PubMed]
- Zecha, J.; Gabriel, W.; Spallek, R.; Chang, Y.C.; Mergner, J.; Wilhelm, M.; Bassermann, F.; Kuster, B. Linking post-translational modifications and protein turnover by site-resolved protein turnover profiling. Nat. Commun. 2022, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Harris, J.L.; Alper, P.B.; Li, J.; Rechsteiner, M.; Backes, B.J. Substrate specificity of the human proteasome. Chem. Biol. 2001, 8, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Boulpicante, M.; Darrigrand, R.; Pierson, A.; Salgues, V.; Rouillon, M.; Gaudineau, B.; Khaled, M.; Cattaneo, A.; Bachi, A.; Cascio, P.; et al. Tumors escape immunosurveillance by overexpressing the proteasome activator PSME3. Oncoimmunology 2020, 9, 1761205. [Google Scholar] [CrossRef] [PubMed]
- Huber, E.M.; Groll, M. The Mammalian Proteasome Activator PA28 Forms an Asymmetric alpha(4)beta(3) Complex. Structure 2017, 25, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Niewerth, D.; Kaspers, G.J.; Assaraf, Y.G.; van Meerloo, J.; Kirk, C.J.; Anderl, J.; Blank, J.L.; van de Ven, P.M.; Zweegman, S.; Jansen, G.; et al. Interferon-gamma-induced upregulation of immunoproteasome subunit assembly overcomes bortezomib resistance in human hematological cell lines. J. Hematol. Oncol. 2014, 7, 7. [Google Scholar] [CrossRef]
- Benham, A.M.; Gromme, M.; Neefjes, J. Allelic differences in the relationship between proteasome activity and MHC class I peptide loading. J. Immunol. 1998, 161, 83–89. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
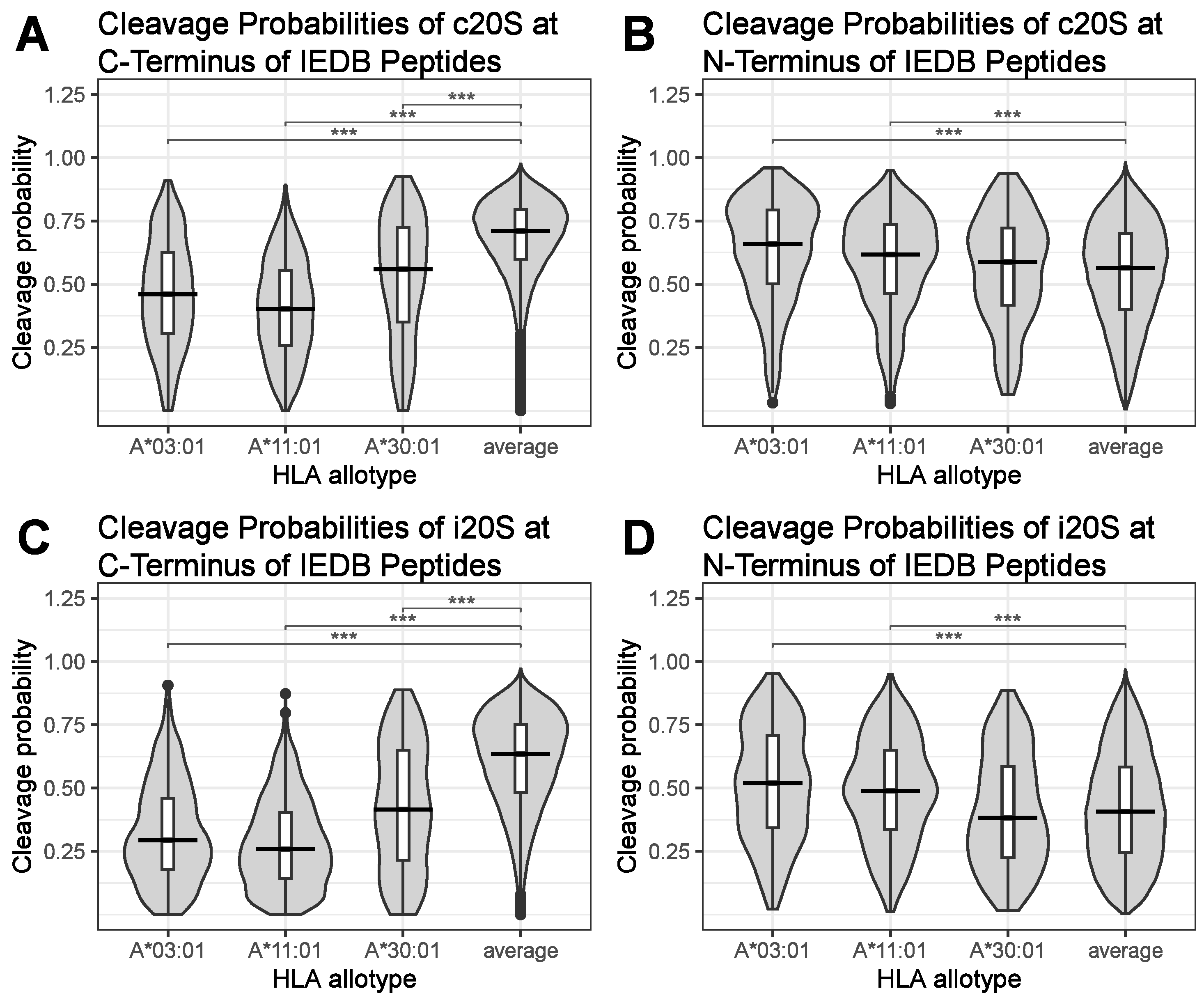{kind=link}
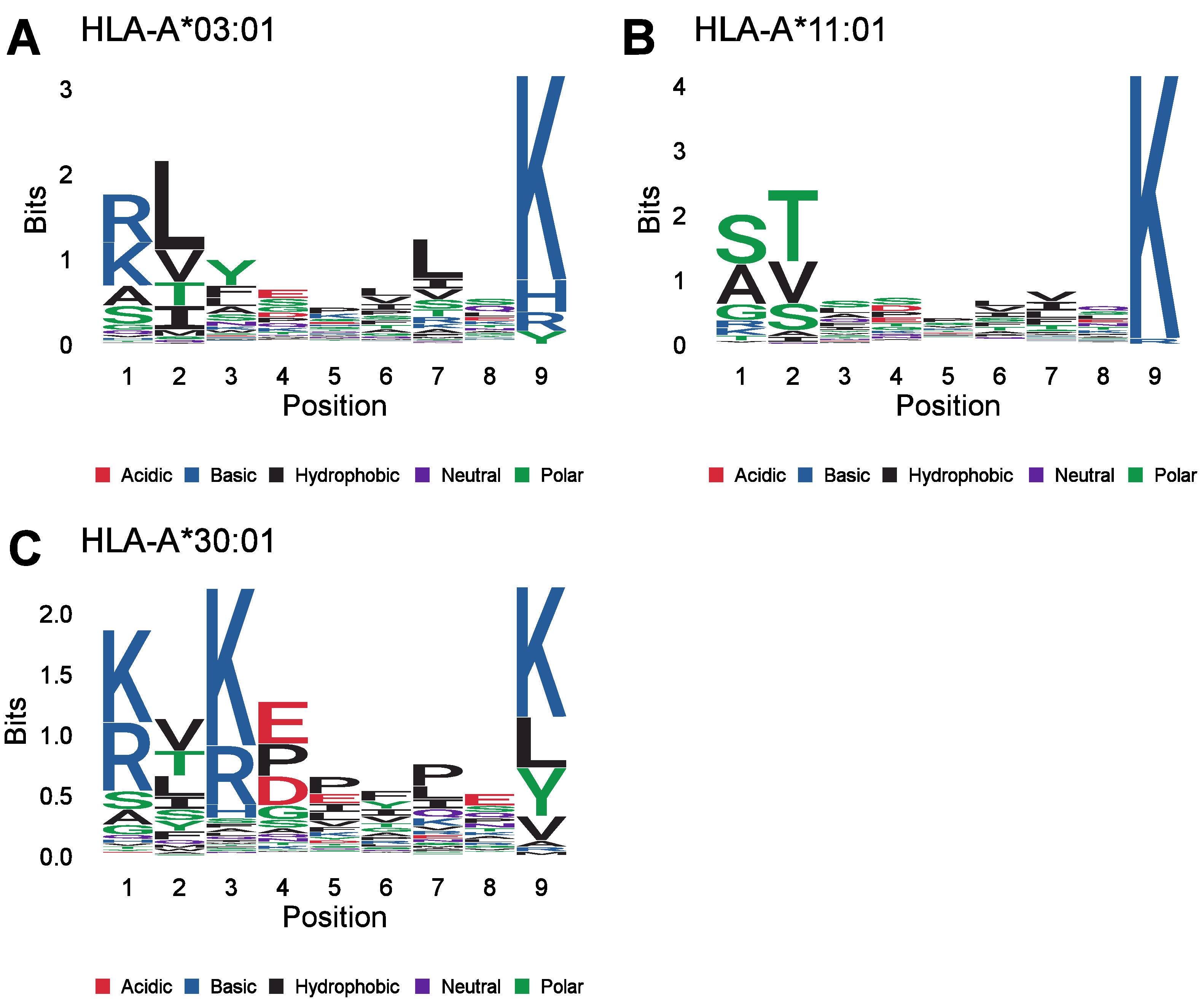{kind=link}
| Gene Symbol | Description | Log2 Abundance Ratio: IFNΓ/Ctrl | Adj. p-Value: IFNΓ/CTRL |
|---|---|---|---|
| PSMB8 | Proteasome subunit beta type-8 | 1.97 | 2.41 × 10−6 |
| PSMB9 | Proteasome subunit beta type-9 | 2.56 | 5.13 × 10−11 |
| PSMB10 | Proteasome subunit beta type-10 | 2.89 | 2.70 × 10−12 |
| TAP1 | Antigen peptide transporter 1 | 3.31 | 3.65 × 10−16 |
| TAP2 | Antigen peptide transporter 2 | 3.00 | 9.66 × 10−14 |
| HLA-A | HLA class I histocompatibility antigen, A alpha chain | 1.36 | 7.47 × 10−3 |
| HLA-B | HLA class I histocompatibility antigen, B alpha chain | 1.68 | 4.72 × 10−6 |
| HLA-C | HLA class I histocompatibility antigen, C alpha chain | 1.80 | 1.95 × 10−5 |
| HLA-E | HLA class I histocompatibility antigen, alpha chain E | 1.99 | 6.32 × 10−6 |
| B2M | Beta-2-microglobulin | 2.40 | 1.09 × 10−9 |
| TAPBP | Tapasin | 2.00 | 6.69 × 10−6 |
| ERAP1 | Endoplasmic reticulum aminopeptidase 1 | 1.16 | 4.63 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmalen, A.; Kammerl, I.E.; Meiners, S.; Noessner, E.; Deeg, C.A.; Hauck, S.M. A Lysine Residue at the C-Terminus of MHC Class I Ligands Correlates with Low C-Terminal Proteasomal Cleavage Probability. Biomolecules 2023, 13, 1300. https://doi.org/10.3390/biom13091300
Schmalen A, Kammerl IE, Meiners S, Noessner E, Deeg CA, Hauck SM. A Lysine Residue at the C-Terminus of MHC Class I Ligands Correlates with Low C-Terminal Proteasomal Cleavage Probability. Biomolecules. 2023; 13(9):1300. https://doi.org/10.3390/biom13091300
Chicago/Turabian StyleSchmalen, Adrian, Ilona E. Kammerl, Silke Meiners, Elfriede Noessner, Cornelia A. Deeg, and Stefanie M. Hauck. 2023. "A Lysine Residue at the C-Terminus of MHC Class I Ligands Correlates with Low C-Terminal Proteasomal Cleavage Probability" Biomolecules 13, no. 9: 1300. https://doi.org/10.3390/biom13091300