
Counting the Toll of Inflammation on Schizophrenia—A Potential Role for Toll-like Receptors

Abstract
:1. Introduction
2. Toll-like Receptors
3. Toll-like Receptors in Central Nervous System (CNS) Immune Regulation
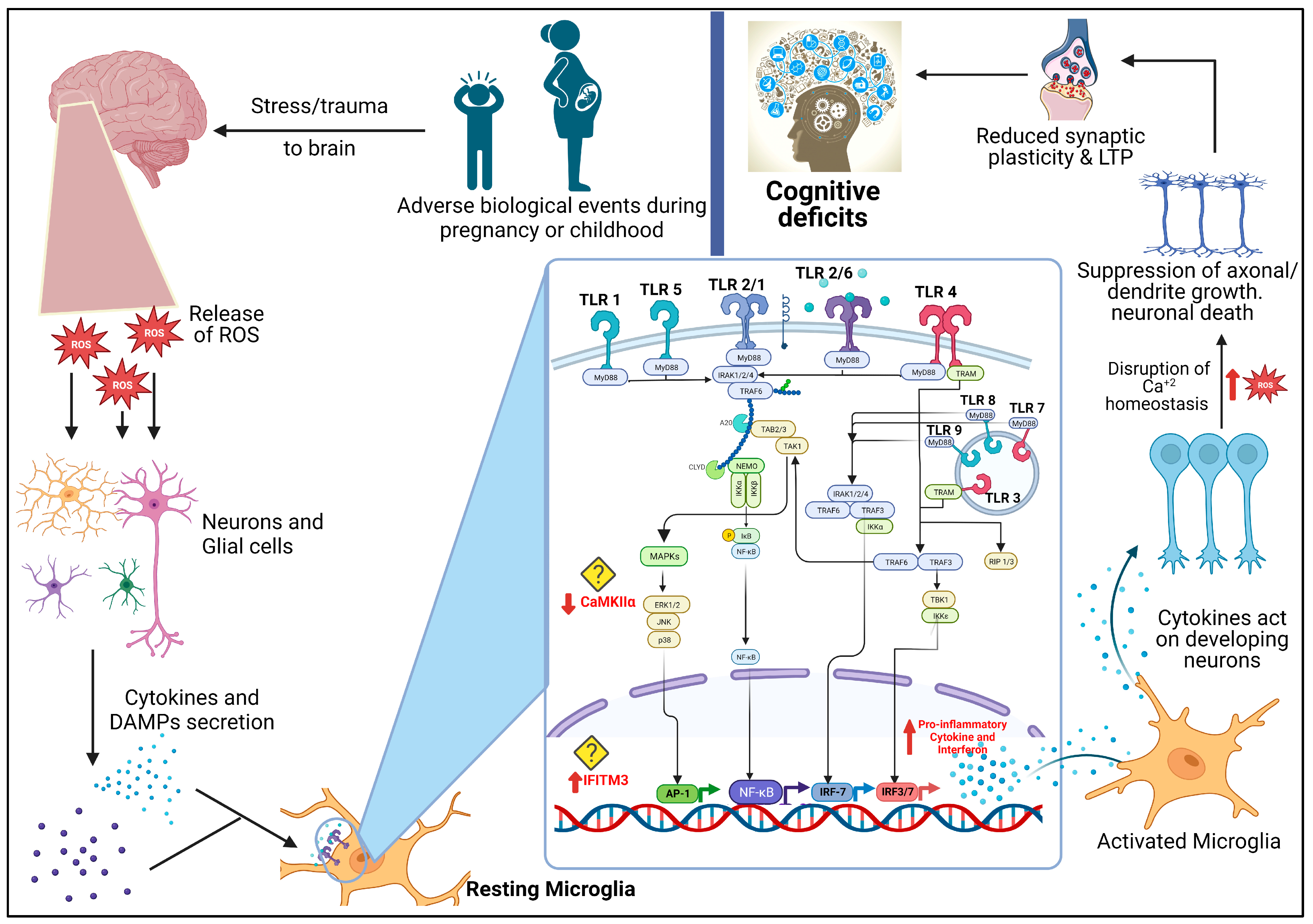
3.1. TLR Signalling in the Brain
3.2. Role of TLR in Neurodevelopment and Pathogenesis
4. The Role of Toll-like Receptors in Schizophrenia
4.1. Maternal Immune Activation (MIA) and Early Life Exposure
4.2. Factors Influencing TLRs in Schizophrenia
4.2.1. Diet and Gut Microbiome
4.2.2. Drugs
4.2.3. Genetics
5. TLRs in Cognitive Impairment in Schizophrenia
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hilker, R.; Helenius, D.; Fagerlund, B.; Skytthe, A.; Christensen, K.; Werge, T.M.; Nordentoft, M.; Glenthøj, B. Heritability of Schizophrenia and Schizophrenia Spectrum Based on the Nationwide Danish Twin Register. Biol. Psychiatry 2018, 83, 492–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramness, J.G.; Gundersen, Ø.H.; Guterstam, J.; Rognli, E.B.; Konstenius, M.; Løberg, E.-M.; Medhus, S.; Tanum, L.; Franck, J. Amphetamine-induced psychosis—A separate diagnostic entity or primary psychosis triggered in the vulnerable? BMC Psychiatry 2012, 12, 221. [Google Scholar] [CrossRef] [Green Version]
- Patterson, P.H. Immune involvement in schizophrenia and autism: Etiology, pathology and animal models. Behav. Brain Res. 2009, 204, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Grohs, M.N.; Reynolds, J.E.; Liu, J.; Martin, J.W.; Pollock, T.; Lebel, C.; Dewey, D.; Kaplan, B.J.; Field, C.J.; Dewey, D.; et al. Prenatal maternal and childhood bisphenol a exposure and brain structure and behavior of young children. Environ. Health 2019, 18, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Wang, G.; Zhang, F. Role of peripheral immune cells-mediated inflammation on the process of neurodegenerative diseases. Front. Immunol. 2020, 11, 582825. [Google Scholar] [CrossRef]
- Mondelli, V.; Dazzan, P.; Pariante, C.M. Immune abnormalities across psychiatric disorders: Clinical relevance. BJPsych Adv. 2015, 21, 150–156. [Google Scholar] [CrossRef] [Green Version]
- McKernan, D.P.; Dennison, U.; Gaszner, G.; Cryan, J.F.; Dinan, T.G. Enhanced peripheral toll-like receptor responses in psychosis: Further evidence of a pro-inflammatory phenotype. Transl. Psychiatry 2011, 1, e36. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-T.; Verma, A.; Hodgkinson, C.P. Toll-like receptors and human disease: Lessons from single nucleotide polymorphisms. Curr. Genom. 2012, 13, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Fiebich, B.L.; Batista, C.R.A.; Saliba, S.W.; Yousif, N.M.; De Oliveira, A.C.P. Role of microglia TLRs in neurodegeneration. Front. Cell. Neurosci. 2018, 12, 329. [Google Scholar] [CrossRef] [Green Version]
- Anderson, K.V.; Jürgens, G.; Nüsslein-Volhard, C. Establishment of dorsal-ventral polarity in the Drosophila embryo: Genetic studies on the role of the Toll gene product. Cell 1985, 42, 779–789. [Google Scholar] [CrossRef]
- Lemaitre, B.; Nicolas, E.; Michaut, L.; Reichhart, J.-M.; Hoffmann, J.A. The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults. Cell 1996, 86, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith, D.; Rapaport, M.; Miller, B. A meta-analysis of blood cytokine network alterations in psychiatric patients: Comparisons between schizophrenia, bipolar disorder and depression. Mol. Psychiatry 2016, 21, 1696–1709. [Google Scholar] [CrossRef]
- Boerrigter, D.; Weickert, T.W.; Lenroot, R.; O’Donnell, M.; Galletly, C.; Liu, D.; Burgess, M.; Cadiz, R.; Jacomb, I.; Catts, V.S. Using blood cytokine measures to define high inflammatory biotype of schizophrenia and schizoaffective disorder. J. Neuroinflamm. 2017, 14, 188. [Google Scholar] [CrossRef] [Green Version]
- Marques, T.R.; Bloomfield, P.; Owen, D.; Gunn, R.; Rabiner, E.; Veronese, M.; Howes, O. 117.4 Pet Imaging of Neuroinflammation in Schizophrenia. Schizophr. Bull. 2017, 43, S64–S65. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.A.; Burgess, S.; Suckling, J.; Lalousis, P.A.; Batool, F.; Griffiths, S.L.; Palmer, E.; Karwath, A.; Barsky, A.; Gkoutos, G.V.; et al. Inflammation and Brain Structure in Schizophrenia and Other Neuropsychiatric Disorders: A Mendelian Randomization Study. JAMA Psychiatry 2022, 79, 498–507. [Google Scholar] [CrossRef]
- Rock, F.L.; Hardiman, G.; Timans, J.C.; Kastelein, R.A.; Bazan, J.F. A family of human receptors structurally related to Drosophila Toll. Proc. Natl. Acad. Sci. USA 1998, 95, 588–593. [Google Scholar] [CrossRef]
- Medzhitov, R.; Preston-Hurlburt, P.; Janeway, C.A. A human homologue of the Drosophila Toll protein signals activation of adaptive immunity. Nature 1997, 388, 394–397. [Google Scholar] [CrossRef]
- Takeuchi, O.; Kawai, T.; Sanjo, H.; Copeland, N.G.; Gilbert, D.; Jenkins, N.A.; Takeda, K.; Akira, S. TLR6: A novel member of an expanding toll-like receptor family. Gene 1999, 231, 59–65. [Google Scholar] [CrossRef]
- Du, X.; Poltorak, A.; Wei, Y.; Beutler, B. Three novel mammalian toll-like receptors: Gene structure, expression, and evolution. Eur. Cytokine Netw. 2000, 11, 362–371. [Google Scholar]
- Chuang, T.-H.; Ulevitch, R.J. Identification of hTLR10: A novel human Toll-like receptor preferentially expressed in immune cells. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2001, 1518, 157–161. [Google Scholar] [CrossRef]
- Moen, S.H.; Ehrnström, B.; Kojen, J.F.; Yurchenko, M.; Beckwith, K.S.; Afset, J.E.; Damås, J.K.; Hu, Z.; Yin, H.; Espevik, T. Human Toll-like receptor 8 (TLR8) is an important sensor of pyogenic bacteria, and is attenuated by cell surface TLR signaling. Front. Immunol. 2019, 10, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, M.; Funami, K.; Tanabe, M.; Oshiumi, H.; Shingai, M.; Seto, Y.; Yamamoto, A.; Seya, T. Subcellular Localization of Toll-Like Receptor 3 in Human Dendritic Cells. J. Immunol. 2003, 171, 3154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A.; Ulevitch, R.J. Toll-like receptors in the induction of the innate immune response. Nature 2000, 406, 782–787. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef]
- Underhill, D.M.; Ozinsky, A. Toll-like receptors: Key mediators of microbe detection. Curr. Opin. Immunol. 2002, 14, 103–110. [Google Scholar] [CrossRef]
- Roh, J.S.; Sohn, D.H. Damage-Associated Molecular Patterns in Inflammatory Diseases. Immune Netw. 2018, 18, e27. [Google Scholar] [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nat. Rev. Immunol. 2020, 20, 95–112. [Google Scholar] [CrossRef]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar]
- Sameer, A.S.; Nissar, S. Toll-Like Receptors (TLRs): Structure, Functions, Signaling, and Role of Their Polymorphisms in Colorectal Cancer Susceptibility. BioMed Res. Int. 2021, 2021, 1157023. [Google Scholar] [CrossRef]
- Akira, S.; Takeda, K. Toll-like receptor signalling. Nat. Rev. Immunol. 2004, 4, 499–511. [Google Scholar] [CrossRef]
- El-Zayat, S.R.; Sibaii, H.; Mannaa, F.A. Toll-like receptors activation, signaling, and targeting: An overview. Bull. Natl. Res. Cent. 2019, 43, 187. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Muldoon, L.L.; Alvarez, J.I.; Begley, D.J.; Boado, R.J.; Del Zoppo, G.J.; Doolittle, N.D.; Engelhardt, B.; Hallenbeck, J.M.; Lonser, R.R.; Ohlfest, J.R.; et al. Immunologic privilege in the central nervous system and the blood-brain barrier. J. Cereb. Blood Flow. Metab. 2013, 33, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Dong, Y.; Benveniste, E.N. Immune function of astrocytes. Glia 2001, 36, 180–190. [Google Scholar] [CrossRef]
- Bsibsi, M.; Ravid, R.; Gveric, D.; van Noort, J.M. Broad Expression of Toll-Like Receptors in the Human Central Nervous System. J. Neuropathol. Exp. Neurol. 2002, 61, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, M.; Naito, S. Tissue-specific mRNA expression profiles of human toll-like receptors and related genes. Biol. Pharm. Bull. 2005, 28, 886–892. [Google Scholar] [CrossRef] [Green Version]
- Jack, C.S.; Arbour, N.; Manusow, J.; Montgrain, V.; Blain, M.; McCrea, E.; Shapiro, A.; Antel, J.P. TLR Signaling Tailors Innate Immune Responses in Human Microglia and Astrocytes. J. Immunol. 2005, 175, 4320–4330. [Google Scholar] [CrossRef] [Green Version]
- Madar, R.; Rotter, A.; Ben-Asher, H.W.; Mughal, M.R.; Arumugam, T.V.; Wood, W.H.; Becker, K.G.; Mattson, M.P.; Okun, E. Postnatal TLR2 activation impairs learning and memory in adulthood. Brain Behav. Immun. 2015, 48, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.-C.; Arumugam, T.V.; Xu, X.; Cheng, A.; Mughal, M.R.; Jo, D.G.; Lathia, J.D.; Siler, D.A.; Chigurupati, S.; Ouyang, X. Pivotal role for neuronal Toll-like receptors in ischemic brain injury and functional deficits. Proc. Natl. Acad. Sci. USA 2007, 104, 13798–13803. [Google Scholar] [CrossRef] [PubMed]
- Lafon, M.; Megret, F.; Lafage, M.; Prehaud, C. The innate immune facet of brain. J. Mol. Neurosci. 2006, 29, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.M.; Rosenberger, K.; Krüger, C.; Habbel, P.; Derkow, K.; Kaul, D.; Rybak, A.; Brandt, C.; Schott, E.; Wulczyn, F.G.; et al. Extracellularly Delivered Single-Stranded Viral RNA Causes Neurodegeneration Dependent on TLR7. J. Immunol. 2012, 189, 1448. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Li, J.; Chiu, I.; Wang, Y.; Sloane, J.A.; Lü, J.; Kosaras, B.; Sidman, R.L.; Volpe, J.J.; Vartanian, T. Toll-like receptor 8 functions as a negative regulator of neurite outgrowth and inducer of neuronal apoptosis. J. Cell Biol. 2006, 175, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Acioglu, C.; Mirabelli, E.; Baykal, A.T.; Ni, L.; Ratnayake, A.; Heary, R.F.; Elkabes, S. Toll like receptor 9 antagonism modulates spinal cord neuronal function and survival: Direct versus astrocyte-mediated mechanisms. Brain Behav. Immun. 2016, 56, 310–324. [Google Scholar] [CrossRef]
- Farina, C.; Krumbholz, M.; Giese, T.; Hartmann, G.; Aloisi, F.; Meinl, E. Preferential expression and function of Toll-like receptor 3 in human astrocytes. J. Neuroimmunol. 2005, 159, 12–19. [Google Scholar] [CrossRef]
- Rivieccio, M.A.; John, G.R.; Song, X.; Suh, H.-S.; Zhao, Y.; Lee, S.C.; Brosnan, C.F. The Cytokine IL-1β Activates IFN Response Factor 3 in Human Fetal Astrocytes in Culture. J. Immunol. 2005, 174, 3719. [Google Scholar] [CrossRef]
- Echchannaoui, H.; Frei, K.; Schnell, C.; Leib, S.L.; Zimmerli, W.; Landmann, R. Toll-like receptor 2–deficient mice are highly susceptible to Streptococcus pneumoniae meningitis because of reduced bacterial clearing and enhanced inflammation. J. Infect. Dis. 2002, 186, 798–806. [Google Scholar]
- Préhaud, C.; Mégret, F.; Lafage, M.; Lafon, M. Virus Infection Switches TLR-3-Positive Human Neurons to Become Strong Producers of Beta Interferon. J. Virol. 2005, 79, 12893–12904. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Shih, Y.-C.; Hung, Y.-F.; Hsueh, Y.-P. Beyond defense: Regulation of neuronal morphogenesis and brain functions via Toll-like receptors. J. Biomed. Sci. 2019, 26, 90. [Google Scholar] [CrossRef] [Green Version]
- Miranda, J.D.; Yaddanapudi, K.; Hornig, M.; Villar, G.; Serge, R.; Lipkin, W.I.; Rall, G.; Biron, C.A. Induction of Toll-Like Receptor 3-Mediated Immunity during Gestation Inhibits Cortical Neurogenesis and Causes Behavioral Disturbances. mBio 2010, 1, e00110–e00176. [Google Scholar] [CrossRef] [Green Version]
- Arrode-Brusés, G.; Brusés, J.L. Maternal immune activation by poly(I:C) induces expression of cytokines IL-1β and IL-13, chemokine MCP-1 and colony stimulating factor VEGF in fetal mouse brain. J. Neuroinflamm. 2012, 9, 83. [Google Scholar] [CrossRef] [Green Version]
- Oh-Nishi, A.; Obayashi, S.; Sugihara, I.; Minamimoto, T.; Suhara, T. Maternal immune activation by polyriboinosinic-polyribocytidilic acid injection produces synaptic dysfunction but not neuronal loss in the hippocampus of juvenile rat offspring. Brain Res. 2010, 1363, 170–179. [Google Scholar] [CrossRef]
- Forrest, C.M.; Khalil, O.S.; Pisar, M.; Smith, R.A.; Darlington, L.G.; Stone, T.W. Prenatal activation of Toll-like receptors-3 by administration of the viral mimetic poly(I:C) changes synaptic proteins, N-methyl-D-aspartate receptors and neurogenesis markers in offspring. Mol. Brain 2012, 5, 22. [Google Scholar] [CrossRef] [Green Version]
- Carpentier, P.A.; Dingman, A.L.; Palmer, T.D. Placental TNF-alpha Signaling in Illness-Induced Complications of Pregnancy. Am. J. Pathol. 2011, 178, 2802–2810. [Google Scholar] [CrossRef] [Green Version]
- Lehnardt, S.; Lachance, C.; Patrizi, S.; Lefebvre, S.; Follett, P.L.; Jensen, F.E.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. The toll-like receptor TLR4 is necessary for lipopolysaccharide-induced oligodendrocyte injury in the CNS. J. Neurosci. 2002, 22, 2478–2486. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.F.; Zhou, X.; Chang, D.; Bhuyan, D.J.; Zhang, J.P.; Yu, W.Z.; Jiang, X.S.; Seto, S.W.; Yeon, S.Y.; Li, J. A novel tri-culture model for neuroinflammation. J. Neurochem. 2021, 156, 249–261. [Google Scholar] [CrossRef]
- WHO. Schizophrenia. Available online: https://www.who.int/news-room/fact-sheets/detail/schizophrenia#:~:text=Schizophrenia%20affects%20approximately%2024%20million,%25)%20among%20adults%20(2) (accessed on 10 August 2022).
- Saloni, D.; Hannah, R.; Max, R. Mental Health. Available online: https://ourworldindata.org/mental-health#schizophrenia (accessed on 11 August 2022).
- Ermakov, E.A.; Melamud, M.M.; Buneva, V.N.; Ivanova, S.A. Immune System Abnormalities in Schizophrenia: An Integrative View and Translational Perspectives. Front. Psychiatry 2022, 13, 880568. [Google Scholar]
- Kozłowska, E.; Agier, J.; Wysokiński, A.; Łucka, A.; Sobierajska, K.; Brzezińska-Błaszczyk, E. The expression of toll-like receptors in peripheral blood mononuclear cells is altered in schizophrenia. Psychiatry Res. 2019, 272, 540–550. [Google Scholar]
- Chase, K.A.; Melbourne, J.K.; Rosen, C.; McCarthy-Jones, S.; Jones, N.; Feiner, B.M.; Sharma, R.P. Traumagenics: At the intersect of childhood trauma, immunity and psychosis. Psychiatry Res. 2019, 273, 369–377. [Google Scholar] [CrossRef]
- Juncal-Ruiz, M.; Riesco-Davila, L.; Vazquez-Bourgon, J.; Ortiz-Garcia de la Foz, V.; Mayoral-Van Son, J.; Ayesa-Arriola, R.; Setien-Suero, E.; Leza, J.C.; Lopez-Hoyos, M.; Crespo-Facorro, B. Expression and functionality study of 9 toll-like receptors in 33 drug-naive non-affective first episode psychosis individuals: A 3-month study. Int. J. Mol. Sci. 2020, 21, 6106. [Google Scholar]
- MacDowell, K.S.; Pinacho, R.; Leza, J.C.; Costa, J.; Ramos, B.; García-Bueno, B. Differential regulation of the TLR4 signalling pathway in post-mortem prefrontal cortex and cerebellum in chronic schizophrenia: Relationship with SP transcription factors. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 481–492. [Google Scholar] [CrossRef] [PubMed]
- López-González, I.; Pinacho, R.; Vila, È.; Escanilla, A.; Ferrer, I.; Ramos, B. Neuroinflammation in the dorsolateral prefrontal cortex in elderly chronic schizophrenia. Eur. Neuropsychopharmacol. 2019, 29, 384–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Bueno, B.; Gassó, P.; MacDowell, K.S.; Callado, L.F.; Mas, S.; Bernardo, M.; Lafuente, A.; Meana, J.J.; Leza, J.C. Evidence of activation of the Toll-like receptor-4 proinflammatory pathway in patients with schizophrenia. J. Psychiatry Neurosci. 2016, 41, E46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.-H.; Chiang, S.-Y.; Chiu, C.-C.; Tsai, C.-C.; Tsai, H.-H.; Huang, C.-Y.; Hsu, T.-C.; Tzang, B.-S. Expression of anti-cardiolipin antibodies and inflammatory associated factors in patients with schizophrenia. Psychiatry Res. 2011, 187, 341–346. [Google Scholar] [CrossRef]
- Müller, N.; Wagner, J.K.; Krause, D.; Weidinger, E.; Wildenauer, A.; Obermeier, M.; Dehning, S.; Gruber, R.; Schwarz, M.J. Impaired monocyte activation in schizophrenia. Psychiatry Res. 2012, 198, 341–346. [Google Scholar] [CrossRef]
- Kéri, S.; Szabó, C.; Kelemen, O. Uniting the neurodevelopmental and immunological hypotheses: Neuregulin 1 receptor ErbB and Toll-like receptor activation in first-episode schizophrenia. Sci. Rep. 2017, 7, 4147. [Google Scholar] [CrossRef] [Green Version]
- Kéri, S.; Szabó, C.; Kelemen, O. Antipsychotics influence Toll-like receptor (TLR) expression and its relationship with cognitive functions in schizophrenia. Brain Behav. Immun. 2017, 62, 256–264. [Google Scholar]
- Li, H.; Chen, W.; Gou, M.; Li, W.; Tong, J.; Zhou, Y.; Xie, T.; Yu, T.; Feng, W.; Li, Y. The relationship between TLR4/NF-κB/IL-1β signaling, cognitive impairment, and white-matter integrity in patients with stable chronic schizophrenia. Front. Psychiatry 2022, 13, 966657. [Google Scholar] [CrossRef]
- Li, N.; Li, Y.; Huang, J.; Zhang, P.; Tong, J.; Chen, S.; Cui, Y.; Tan, S.; Wang, Z.; Yang, F. Dysfunctional monocytic toll-like receptor 4 signaling pathway and cognitive deficits in chronic schizophrenia patients with tardive dyskinesia. Neurosci. Lett. 2022, 777, 136581. [Google Scholar] [CrossRef]
- Menninger, K.A. Influenza and schizophrenia: An analysis of post-influenzal “dementia precox”, as of 1918, and five years later. Am. J. Psychiatry 1926, 82, 469–529. [Google Scholar] [CrossRef]
- Kępińska, A.P.; Iyegbe, C.O.; Vernon, A.C.; Yolken, R.; Murray, R.M.; Pollak, T.A. Schizophrenia and Influenza at the Centenary of the 1918-1919 Spanish Influenza Pandemic: Mechanisms of Psychosis Risk. Front. Psychiatry 2020, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Buka, S.L.; Tsuang, M.T.; Torrey, E.F.; Klebanoff, M.A.; Bernstein, D.; Yolken, R.H. Maternal Infections and Subsequent Psychosis Among Offspring. Arch. Gen. Psychiatry 2001, 58, 1032–1037. [Google Scholar] [CrossRef] [Green Version]
- Dickerson, F.; Jones-Brando, L.; Ford, G.; Genovese, G.; Stallings, C.; Origoni, A.; O’Dushlaine, C.; Katsafanas, E.; Sweeney, K.; Khushalani, S.; et al. Schizophrenia is Associated with an Aberrant Immune Response to Epstein–Barr Virus. Schizophr. Bull. 2018, 45, 1112–1119. [Google Scholar] [CrossRef] [Green Version]
- Burgdorf, K.S.; Trabjerg, B.B.; Pedersen, M.G.; Nissen, J.; Banasik, K.; Pedersen, O.B.; Sørensen, E.; Nielsen, K.R.; Larsen, M.H.; Erikstrup, C.; et al. Large-scale study of Toxoplasma and Cytomegalovirus shows an association between infection and serious psychiatric disorders. Brain Behav. Immun. 2019, 79, 152–158. [Google Scholar] [CrossRef]
- Choudhury, Z.; Lennox, B. Maternal Immune Activation and Schizophrenia-Evidence for an Immune Priming Disorder. Front. Psychiatry 2021, 12, 585742. [Google Scholar] [CrossRef]
- Ashdown, H.; Dumont, Y.; Ng, M.; Poole, S.; Boksa, P.; Luheshi, G.N. The role of cytokines in mediating effects of prenatal infection on the fetus: Implications for schizophrenia. Mol. Psychiatry 2006, 11, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Meyer, U.; Feldon, J.; Yee, B.K. A Review of the Fetal Brain Cytokine Imbalance Hypothesis of Schizophrenia. Schizophr. Bull. 2008, 35, 959–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, S.T.; Scott, G.; Naing, Z.; Iwasenko, J.; Hall, B.; Graf, N.; Arbuckle, S.; Craig, M.E.; Rawlinson, W.D. Human Cytomegalovirus-Induces Cytokine Changes in the Placenta with Implications for Adverse Pregnancy Outcomes. PLoS ONE 2013, 7, e52899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.-R.; Wei, X.-W.; Lin, Y.; Wang, Y.-H. Toll-Like Receptor-Dependent Antiviral Responses at the Maternal-Fetal Interface. Reprod. Dev. Med. 2020, 04, 251–256. [Google Scholar] [CrossRef]
- Yockey, L.J.; Iwasaki, A. Interferons and Proinflammatory Cytokines in Pregnancy and Fetal Development. Immunity 2018, 49, 397–412. [Google Scholar] [CrossRef] [Green Version]
- Kuzmicki, M.; Telejko, B.; Wawrusiewicz-Kurylonek, N.; Lipinska, D.; Pliszka, J.; Wilk, J.; Zielinska, A.; Skibicka, J.; Szamatowicz, J.; Kretowski, A. The expression of genes involved in NF-kB activation in peripheral blood mononuclear cells of patients with gestational diabetes. Eur. J. Endocrinol. 2013, 168, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Yu, X.; Liu, J.; Wang, J. Pro-inflammatory cytokines are elevated in pregnant women with systemic lupus erythematosus in association with the activation of TLR4. Clin. Lab. 2016, 62, 535–544. [Google Scholar] [CrossRef]
- Yang, X.; Li, M.; Haghiac, M.; Catalano, P.M.; O’Tierney-Ginn, P.; Hauguel-de Mouzon, S. Causal relationship between obesity-related traits and TLR4-driven responses at the maternal–fetal interface. Diabetologia 2016, 59, 2459–2466. [Google Scholar] [CrossRef] [Green Version]
- Kendell, R.E.; Kemp, I.W. Maternal Influenza in the Etiology of Schizophrenia. Arch. Gen. Psychiatry 1989, 46, 878–882. [Google Scholar] [CrossRef]
- Buka, S.L.; Tsuang, M.T.; Torrey, E.F.; Klebanoff, M.A.; Wagner, R.L.; Yolken, R.H. Maternal cytokine levels during pregnancy and adult psychosis. Brain Behav. Immun. 2001, 15, 411–420. [Google Scholar] [CrossRef] [Green Version]
- Melnik, A.; Tauber, S.; Dumrese, C.; Ullrich, O.; Wolf, S.A. Murine adult neural progenitor cells alter their proliferative behavior and gene expression after the activation of toll-like-receptor 3. Eur. J. Microbiol. Immunol. 2012, 2, 239–248. [Google Scholar] [CrossRef] [Green Version]
- Ni, L.; Acevedo, G.; Muralidharan, B.; Padala, N.; To, J.; Jonakait, G.M. Toll-like receptor ligands and CD154 stimulate microglia to produce a factor(s) that promotes excess cholinergic differentiation in the developing rat basal forebrain: Implications for neurodevelopmental disorders. Pediatr. Res. 2007, 61, 15–20. [Google Scholar]
- Hennessy, C.; McKernan, D.P. Anti-Viral Pattern Recognition Receptors as Therapeutic Targets. Cells 2021, 10, 2258. [Google Scholar] [CrossRef]
- Xia, Y.; Qi, F.; Zou, J.; Yang, J.; Yao, Z. Influenza vaccination during early pregnancy contributes to neurogenesis and behavioral function in offspring. Brain Behav. Immun. 2014, 42, 212–221. [Google Scholar] [CrossRef]
- Talukdar, P.M.; Abdul, F.; Maes, M.; Berk, M.; Venkatasubramanian, G.; Kutty, B.M.; Debnath, M. A proof-of-concept study of maternal immune activation mediated induction of Toll-like receptor (TLR) and inflammasome pathways leading to neuroprogressive changes and schizophrenia-like behaviours in offspring. Eur. Neuropsychopharmacol. 2021, 52, 48–61. [Google Scholar] [CrossRef] [PubMed]
- O’Loughlin, E.; Pakan, J.M.; Yilmazer-Hanke, D.; McDermott, K.W. Acute in utero exposure to lipopolysaccharide induces inflammation in the pre-and postnatal brain and alters the glial cytoarchitecture in the developing amygdala. J. Neuroinflamm. 2017, 14, 212. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jeon, S.J.; Choi, J.W.; Ryu, J.H. Activation of maternal toll-like receptor-2 causes social deficits and memory impairment in female offspring. NeuroReport 2016, 27, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Connolly, M.G.; Potter, O.V.; Sexton, A.R.; Kohman, R.A. Effects of Toll-like receptor 4 inhibition on spatial memory and cell proliferation in male and female adult and aged mice. Brain Behav. Immun. 2021, 97, 383–393. [Google Scholar] [CrossRef]
- Drouin-Ouellet, J.; LeBel, M.; Filali, M.; Cicchetti, F. MyD88 deficiency results in both cognitive and motor impairments in mice. Brain Behav. Immun. 2012, 26, 880–885. [Google Scholar] [CrossRef]
- Ibi, D.; Nagai, T.; Kitahara, Y.; Mizoguchi, H.; Koike, H.; Shiraki, A.; Takuma, K.; Kamei, H.; Noda, Y.; Nitta, A.; et al. Neonatal polyI:C treatment in mice results in schizophrenia-like behavioral and neurochemical abnormalities in adulthood. Neurosci. Res. 2009, 64, 297–305. [Google Scholar] [CrossRef]
- Liu, H.-Y.; Hong, Y.-F.; Huang, C.-M.; Chen, C.-Y.; Huang, T.-N.; Hsueh, Y.-P. TLR7 negatively regulates dendrite outgrowth through the Myd88–c-Fos–IL-6 pathway. J. Neurosci. 2013, 33, 11479–11493. [Google Scholar] [CrossRef]
- Nakajima, A.; Ibi, D.; Nagai, T.; Yamada, S.; Nabeshima, T.; Yamada, K. Induction of interferon-induced transmembrane protein 3 gene expression by lipopolysaccharide in astrocytes. Eur. J. Pharmacol. 2014, 745, 166–175. [Google Scholar] [CrossRef]
- Zhou, X.Y.; Liu, J.; Xu, Z.P.; Fu, Q.; Wang, P.Q.; Wang, J.H.; Zhang, H. Dexmedetomidine ameliorates postoperative cognitive dysfunction by inhibiting Toll-like receptor 4 signaling in aged mice. Kaohsiung J. Med. Sci. 2020, 36, 721–731. [Google Scholar] [CrossRef]
- Manly, J.T.; Kim, J.E.; Rogosch, F.A.; Cicchetti, D. Dimensions of child maltreatment and children’s adjustment: Contributions of developmental timing and subtype. Dev. Psychopathol. 2001, 13, 759–782. [Google Scholar]
- Jurewicz, J.; Polanska, K.; Hanke, W. Chemical exposure early in life and the neurodevelopment of children–an overview of current epidemiological evidence. Ann. Agric. Environ. Med. 2013, 20, 3. [Google Scholar]
- Khandaker, G.M.; Zimbron, J.; Dalman, C.; Lewis, G.; Jones, P.B. Childhood infection and adult schizophrenia: A meta-analysis of population-based studies. Schizophr. Res. 2012, 139, 161–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghel, M.S.; Singh, B.; Dhuriya, Y.K.; Shukla, R.K.; Patro, N.; Khanna, V.K.; Patro, I.K.; Thakur, M.K. Postnatal exposure to poly (I:C) impairs learning and memory through changes in synaptic plasticity gene expression in developing rat brain. Neurobiol. Learn. Mem. 2018, 155, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Dhuriya, Y.K.; Patro, N.; Thakur, M.K.; Khanna, V.K.; Patro, I.K. Early life exposure to poly I: C impairs striatal DA-D2 receptor binding, myelination and associated behavioural abilities in rats. J. Chem. Neuroanat. 2021, 118, 102035. [Google Scholar] [CrossRef] [PubMed]
- King, S.; Holleran, L.; Mothersill, D.; Patlola, S.; Rokita, K.; McManus, R.; Kenyon, M.; McDonald, C.; Hallahan, B.; Corvin, A.; et al. Early life Adversity, functional connectivity and cognitive performance in Schizophrenia: The mediating role of IL-6. Brain Behav. Immun. 2021, 98, 388–396. [Google Scholar] [CrossRef]
- Flouri, E.; Francesconi, M.; Papachristou, E.; Midouhas, E.; Lewis, G. Stressful life events, inflammation and emotional and behavioural problems in children: A population-based study. Brain Behav. Immun. 2019, 80, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Song, R.; Gao, Y.; Dozmorov, I.; Malladi, V.; Saha, I.; McDaniel, M.M.; Parameswaran, S.; Liang, C.; Arana, C.; Zhang, B.; et al. IRF1 governs the differential interferon-stimulated gene responses in human monocytes and macrophages by regulating chromatin accessibility. Cell Rep. 2021, 34, 108891. [Google Scholar] [CrossRef]
- Lumey, L.H.; Stein, A.D.; Susser, E. Prenatal famine and adult health. Annu. Rev. Public. Health 2011, 32, 237–262. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Phillips, M.R.; Zhang, Y.; Zhang, J.; Shi, Q.; Song, Z.; Ding, Z.; Pang, S.; Martorell, R. Malnutrition in early life and adult mental health: Evidence from a natural experiment. Soc. Sci. Med. 2013, 97, 259–266. [Google Scholar] [CrossRef] [Green Version]
- Selhub, E. Nutritional psychiatry: Your brain on food. Harv. Health Blog 2020, 16, 11. [Google Scholar]
- Lee, J.Y.; Zhao, L.; Youn, H.S.; Weatherill, A.R.; Tapping, R.; Feng, L.; Lee, W.H.; Fitzgerald, K.A.; Hwang, D.H. Saturated fatty acid activates but polyunsaturated fatty acid inhibits Toll-like receptor 2 dimerized with Toll-like receptor 6 or 1. J. Biol. Chem. 2004, 279, 16971–16979. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, D.; Wang, F.; Liu, S.; Zhao, S.; Ling, E.-A.; Hao, A. Saturated fatty acids activate microglia via Toll-like receptor 4/NF-κB signalling. Br. J. Nutr. 2012, 107, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Plakidas, A.; Lee, W.H.; Heikkinen, A.; Chanmugam, P.; Bray, G.; Hwang, D.H. Differential modulation of Toll-like receptors by fatty acids: Preferential inhibition by n-3 polyunsaturated fatty acids. J. Lipid Res. 2003, 44, 479–486. [Google Scholar] [CrossRef] [Green Version]
- Fessler, M.B.; Rudel, L.L.; Brown, J.M. Toll-like receptor signaling links dietary fatty acids to the metabolic syndrome. Curr. Opin. Lipidol. 2009, 20, 379–385. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Wang, H.; Zhang, B.; Popkin, B.M.; Du, S. Elevated Fat Intake Increases Body Weight and the Risk of Overweight and Obesity among Chinese Adults: 1991–2015 Trends. Nutrients 2020, 12, 3272. [Google Scholar] [CrossRef]
- Ding, S.; Chi, M.M.; Scull, B.P.; Rigby, R.; Schwerbrock, N.M.J.; Magness, S.; Jobin, C.; Lund, P.K. High-fat diet: Bacteria interactions promote intestinal inflammation which precedes and correlates with obesity and insulin resistance in mouse. PLoS ONE 2010, 5, e12191. [Google Scholar] [CrossRef] [Green Version]
- Jialal, I.; Kaur, H.; Devaraj, S. Toll-like Receptor Status in Obesity and Metabolic Syndrome: A Translational Perspective. J. Clin. Endocrinol. Metab. 2014, 99, 39–48. [Google Scholar] [CrossRef]
- Marrone, M.C.; Coccurello, R. Dietary Fatty Acids and Microbiota-Brain Communication in Neuropsychiatric Diseases. Biomolecules 2019, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Arentsen, T.; Qian, Y.; Gkotzis, S.; Femenia, T.; Wang, T.; Udekwu, K.; Forssberg, H.; Diaz Heijtz, R. The bacterial peptidoglycan-sensing molecule Pglyrp2 modulates brain development and behavior. Mol. Psychiatry 2017, 22, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Severance, E.G.; Yolken, R.H.; Eaton, W.W. Autoimmune diseases, gastrointestinal disorders and the microbiome in schizophrenia: More than a gut feeling. Schizophr. Res. 2016, 176, 23–35. [Google Scholar] [CrossRef] [Green Version]
- R Caso, J.; Balanza-Martinez, V.; Palomo, T.; Garcia-Bueno, B. The microbiota and gut-brain axis: Contributions to the immunopathogenesis of schizophrenia. Curr. Pharm. Des. 2016, 22, 6122–6133. [Google Scholar] [CrossRef] [PubMed]
- Severance, E.G.; Alaedini, A.; Yang, S.; Halling, M.; Gressitt, K.L.; Stallings, C.R.; Origoni, A.E.; Vaughan, C.; Khushalani, S.; Leweke, F.M. Gastrointestinal inflammation and associated immune activation in schizophrenia. Schizophr. Res. 2012, 138, 48–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadgyas-Stanculete, M.; Buga, A.-M.; Popa-Wagner, A.; Dumitrascu, D.L. The relationship between irritable bowel syndrome and psychiatric disorders: From molecular changes to clinical manifestations. J. Mol. Psychiatry 2014, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.J.; Buckley, P.; Seabolt, W.; Mellor, A.; Kirkpatrick, B. Meta-analysis of cytokine alterations in schizophrenia: Clinical status and antipsychotic effects. Biol. Psychiatry 2011, 70, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Tourjman, V.; Kouassi, E.; Koue, M.-E.; Rocchetti, M.; Fortin-Fournier, S.; Fusar-Poli, P.; Potvin, S. Antipsychotics’ effects on blood levels of cytokines in schizophrenia: A meta-analysis. Schizophr. Res. 2013, 151, 43–47. [Google Scholar]
- Marcinowicz, P.; Więdłocha, M.; Zborowska, N.; Dębowska, W.; Podwalski, P.; Misiak, B.; Tyburski, E.; Szulc, A. A meta-analysis of the influence of antipsychotics on cytokines levels in first episode psychosis. J. Clin. Med. 2021, 10, 2488. [Google Scholar] [CrossRef]
- Patlola, S.R.; Donohoe, G.; McKernan, D.P. Anti-inflammatory effects of 2nd generation antipsychotics in patients with schizophrenia: A systematic review and meta-analysis. J. Psychiatr. Res. 2023, 160, 126–136. [Google Scholar] [CrossRef]
- MacDowell, K.; Caso, J.; Martin-Hernandez, D.; Madrigal, J.; Leza, J.; Garcia-Bueno, B. Paliperidone prevents brain toll-like receptor 4 pathway activation and neuroinflammation in rat models of acute and chronic restraint stress. Int. J. Neuropsychopharmacol. 2015, 18. [Google Scholar]
- Jeon, S.; Kim, S.H.; Shin, S.Y.; Lee, Y.H. Clozapine reduces Toll-like receptor 4/NF-κB-mediated inflammatory responses through inhibition of calcium/calmodulin-dependent Akt activation in microglia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2018, 81, 477–487. [Google Scholar] [CrossRef]
- MacDowell, K.S.; Munarriz-Cuezva, E.; Caso, J.R.; Madrigal, J.L.; Zabala, A.; Meana, J.J.; García-Bueno, B.; Leza, J.C. Paliperidone reverts Toll-like receptor 3 signaling pathway activation and cognitive deficits in a maternal immune activation mouse model of schizophrenia. Neuropharmacology 2016, 116, 196–207. [Google Scholar] [CrossRef]
- Li, W.-T.; Huang, X.-F.; Deng, C.; Zhang, B.-H.; Qian, K.; He, M.; Sun, T.-L. Olanzapine Induces inflammation and immune response via activating ER stress in the rat prefrontal cortex. Curr. Med. Sci. 2021, 41, 788–802. [Google Scholar] [CrossRef]
- Balaji, R.; Subbanna, M.; Shivakumar, V.; Abdul, F.; Venkatasubramanian, G.; Debnath, M. Pattern of expression of toll like receptor (TLR)-3 and-4 genes in drug-naïve and antipsychotic treated patients diagnosed with schizophrenia. Psychiatry Res. 2020, 285, 112727. [Google Scholar]
- Yang, Z.; Xiong, G.L. Alcohol-Related Psychosis. Available online: https://emedicine.medscape.com/article/289848-overview (accessed on 11 August 2022).
- Du, S.-H.; Qiao, D.-F.; Chen, C.-X.; Chen, S.; Liu, C.; Lin, Z.; Wang, H.; Xie, W.-B. Toll-Like Receptor 4 Mediates Methamphetamine-Induced Neuroinflammation through Caspase-11 Signaling Pathway in Astrocytes. Front. Mol. Neurosci. 2017, 10, 409. [Google Scholar] [CrossRef] [Green Version]
- Medhus, S.; Rognli, E.B.; Gossop, M.; Holm, B.; Mørland, J.; Bramness, J.G. Amphetamine-induced psychosis: Transition to schizophrenia and mortality in a small prospective sample. Am. J. Addict. 2015, 24, 586–589. [Google Scholar]
- Sukhumsirichart, W. Polymorphisms. In Genetic Diversity and Disease Susceptibility; IntechOpen: London, UK, 2018. [Google Scholar]
- Medvedev, A.E. Toll-like receptor polymorphisms, inflammatory and infectious diseases, allergies, and cancer. J. Interferon Cytokine Res. 2013, 33, 467–484. [Google Scholar] [CrossRef] [Green Version]
- Crisafulli, C.; Drago, A.; Calabrò, M.; Spina, E.; Serretti, A. A molecular pathway analysis informs the genetic background at risk for schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2015, 59, 21–30. [Google Scholar]
- Aflouk, Y.; Inoubli, O.; Saoud, H.; Zaafrane, F.; Gaha, L.; Bel Hadj Jrad, B. Association between TLR2 polymorphisms (− 196–174 Ins/Del, R677W, R753Q, and P631H) and schizophrenia in a Tunisian population. Immunol. Res. 2021, 69, 541–552. [Google Scholar] [CrossRef]
- Kang, W.S.; Park, J.K.; Lee, S.M.; Kim, S.K.; Park, H.J.; Kim, J.W. Association between genetic polymorphisms of Toll-like receptor 2 (TLR2) and schizophrenia in the Korean population. Gene 2013, 526, 182–186. [Google Scholar] [CrossRef]
- Sharma, I.; Priya, I.; Sharma, S.; Gupta, S.; Arora, M.; Mahajan, R.; Kapoor, N. Association of toll-like receptor 2 gene polymorphism (rs3804099) with susceptibility to Schizophrenia risk in the Dogra population of Jammu region, North India. Eur. J. Psychiatry 2022, 36, 106–113. [Google Scholar] [CrossRef]
- Mostafa, M.; Elwasify, M.; Fathy, A.A.; Abdelsalam, M. Toll-Like Receptor 4 Gene Polymorphisms and Susceptibility to Schizophrenia: A Case-Control Study. Immunol. Investig. 2022, 51, 2009–2024. [Google Scholar] [CrossRef] [PubMed]
- Cowman, M.; Holleran, L.; Lonergan, E.; O’Connor, K.; Birchwood, M.; Donohoe, G. Cognitive Predictors of Social and Occupational Functioning in Early Psychosis: A Systematic Review and Meta-analysis of Cross-Sectional and Longitudinal Data. Schizophr. Bull. 2021, 47, 1243–1253. [Google Scholar] [CrossRef] [PubMed]
- Fairweather-Schmidt, A.; Batterham, P.; Butterworth, P.; Nada-Raja, S. The impact of suicidality on health-related quality of life: A latent growth curve analysis of community-based data. J. Affect. Disord. 2016, 203, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa, J.; Choi, J.; Kangas, J.L.; Kaufmann, C.N.; Harvey, P.D.; Depp, C.A. Associations of suicidality with cognitive ability and cognitive insight in outpatients with Schizophrenia. Schizophr. Res. 2018, 192, 340–344. [Google Scholar] [CrossRef]
- Patlola, S.R.; Donohoe, G.; McKernan, D.P. The relationship between inflammatory biomarkers and cognitive dysfunction in patients with schizophrenia: A systematic review and meta-analysis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2023, 121, 110668. [Google Scholar] [CrossRef]
- Park, S.; Lee, J.; Kim, S.; Choi, S.; Yune, T.; Ryu, J. Toll-like receptor 2 deficiency induces schizophrenia-like behaviors in mice. Sci. Rep. 2015, 5, 8502. [Google Scholar]
- Lin, F.; Shan, W.R.; Zheng, Y.X.; Pan, L.H.; Zuo, Z.Y. Toll-like receptor 2 activation and up-regulation by high mobility group box-1 contribute to post-operative neuroinflammation and cognitive dysfunction in mice. J. Neurochem. 2021, 158, 328–341. [Google Scholar] [CrossRef]
- Ihara, F.; Tanaka, S.; Fereig, R.M.; Nishimura, M.; Nishikawa, Y. Involvement of Toll-like receptor 2 in the cerebral immune response and behavioral changes caused by latent Toxoplasma infection in mice. PLoS ONE 2019, 14, e0220560. [Google Scholar] [CrossRef] [Green Version]
- Bae, H.J.; Kim, J.; Bae, H.J.; Park, K.; Yang, X.; Cho, Y.-j.; Jung, S.Y.; Park, S.J.; Ryu, J.H. Effects of repetitive training on learning and memory performance of TLR2 KO mice. Behav. Brain Res. 2022, 426, 113836. [Google Scholar] [CrossRef]
- Chen, S.; Tian, L.; Chen, N.; Xiu, M.-H.; Wang, Z.-R.; Wang, Y.-C.; Tan, Y.-L.; Wang, C.-Y. More dampened monocytic Toll-like receptor 4 response to lipopolysaccharide and its association with cognitive function in Chinese Han first-episode patients with schizophrenia. Schizophr. Res. 2019, 206, 300–306. [Google Scholar] [CrossRef]
- Yamada, S.; Itoh, N.; Nagai, T.; Nakai, T.; Ibi, D.; Nakajima, A.; Nabeshima, T.; Yamada, K. Innate immune activation of astrocytes impairs neurodevelopment via upregulation of follistatin-like 1 and interferon-induced transmembrane protein 3. J. Neuroinflamm. 2018, 15, 295. [Google Scholar] [CrossRef]
- Arion, D.; Unger, T.; Lewis, D.A.; Levitt, P.; Mirnics, K. Molecular evidence for increased expression of genes related to immune and chaperone function in the prefrontal cortex in schizophrenia. Biol. Psychiatry 2007, 62, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Ogundele, O.; Lee, C. CaMKIIα expression in a mouse model of NMDAR hypofunction schizophrenia: Putative roles for IGF-1R and TLR4. Brain Res. Bull. 2018, 137, 53–70. [Google Scholar]
- O’Neill, L.A.J.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors—redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Liu, H.-Y.; Chen, C.-Y.; Hsueh, Y.-P. Neuronally-expressed Sarm1 regulates expression of inflammatory and antiviral cytokines in brains. Innate Immun. 2013, 20, 161–172. [Google Scholar] [CrossRef]
- Lin, C.-W.; Hsueh, Y.-P. Sarm1, a neuronal inflammatory regulator, controls social interaction, associative memory and cognitive flexibility in mice. Brain Behav. Immun. 2014, 37, 142–151. [Google Scholar] [CrossRef]
- Venkatasubramanian, G.; Debnath, M. The TRIPS (Toll-like receptors in immuno-inflammatory pathogenesis) Hypothesis: A novel postulate to understand schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 44, 301–311. [Google Scholar]


{kind=link}
| Study | Diagnosis of Schizophrenia | Sample Type | TLR Analysed | Analysis Result |
|---|---|---|---|---|
| García-Bueno et al., 2016 [66] | DSM-IV. Absence of other neuropsychiatric disorders and drug abuse. | Postmortem human brain tissue (PFC). | TLR4 and MyD88 protein and mRNA expression. | Significant higher TLR4, MyD88 not IkBa and NFkB protein expression in patients. No change in MyD88 mRNA expression. Significant higher TLR4, MyD88 not IkBa protein expression and MyD88 mRNA expression in patients treated with antipsychotics vs. controls. Significant higher NFkB protein expression in antipsychotic free patients vs. controls. |
| López-González et al., 2019 [65] | DSM-IV and ICD-10. No history of neurological diseases. | Postmortem humanbrain tissue (PFC). | TLR4 and TLR7 mRNA expression. | Significant downregulation of TLR4 not TLR7 in patients. |
| MacDowell et al., 2017 [64] | DSM-IV and ICD-10. No history of neurological diseases. | Postmortem humanbrain tissue (PFC and CB). | TLR4 and MyD88 protein expression. | Significant lower TLR4, MyD88, and IkBa, not NFkB in patients (PFC). Significant higher TLR4, MyD88, and IkBa, not NFkB in patients (CB). |
| Chase et al., 2019 [62] | SCID-IV. No current or previous psychiatric history. | PBMC. | TLR4 mRNA expression. | Adverse childhood events were associated with IL-6 expression, but not TLR4. |
| Juncal-Ruiz et al., 2020 [63] | DSM-IV. Absence of other neuropsychiatric disorders and drug abuse. | PBMC. | TLR1–9 protein expression. | FEP patients showed significantly higher TLR1, but not TLR 5 and 8 in PBMCs. Three months of antipsychotic treatment led to increase in TLR6 monocyte expression, and a similar trend for TLR8 and 9 in PBMCs. |
| Kozłowska et al., 2019 [61] | DSM-IV and ICD-10. No history of neurological diseases. | PBMC. | TLR1–9 mRNA. | TLR1, 2, 4, 6, and 9 expressions were downregulated in patients. TLR3 and 7 expressions were upregulated in patients. TLR5 and 8 no change. |
| Chang et al., 2011 [67] | Patients were recruited if they experienced psychosis (hallucinations or delusions). | Monocytes. | TLR1–6 and 9 mRNA expression. | Downregulation of TLR3 and 5 mRNA in patients. No change with other TLRs. |
| Kéri et al., 2017 [70] | SCID-CV. No current or previous psychiatric history. | Monocytes. | TLR1, 2, 4, 5, and 6 protein expression. | Significant higher TLR4 and 5 expression in patients. At baseline, TLR4 and 5 expression correlates with RBANS scores, but not postmedication. |
| Li et al., 2022 [71] | DSM-IV. Absence of other neuropsychiatric disorders and drug abuse. | Monocytes. | TLR4 protein expression. | Elevated TLR4 was associated with decreased processing speed and social cognition. |
| Li et al., 2022 [72] | DSM-IV. Absence of other neuropsychiatric disorders and drug abuse. | Monocytes. | TLR4 protein expression. | Elevated TLR4 was associated with decreased reasoning/problem-solving and MCCB scores in schizophrenia patients with tardive dyskinesia, but not with no tardive dyskinesia or controls. |
| Müller et al., 2012 [68] | DSM-IV. | Monocytes stimulated with poly (IC), LPS. | TLR2, 3, and 4 protein expression. | No change with TLR2. Significant higher TLR3 in patients vs. controls (unstimulated and PolyIC); no changes in LPS stimulated. Significant higher TLR4 in patients vs. controls (unstimulated); no changes in LPS and polyIC stimulated. |
| Study | Animal (Type of Cells/Culture) | Model | TLR Analysed | Analysis Results |
|---|---|---|---|---|
| Talukdar et al., 2021 [93] | Rats | MIA (Poly IC and LPS) | TLR2, 3, and 4 mRNA | Elevated TLR genes were highly associated with a decrease in sociability and normal behaviour in the offspring. |
| Lee et al., 2016 [95] | Mice | MIA (Zymosan) | TLR2 | TLR2 stimulation showed decreased cognitive and behavioural changes only in female offspring. |
| O’Loughlin et al., 2017 [94] | Mice | MIA (LPS) | TLR2 and 4 mRNA | Elevated TLR genes in pre/postnatal stages after stimulation. These were associated with morphological changes in microglia, suggesting activation. |
| Connolly et al., 2021 [96] | Mice | TLR 4 antagonism | TLR4 | TLR4 antagonism showed beneficial cognitive effects (enhanced spatial learning and memory) only in young female mice. |
| Drouin-Ouellet et al., 2012 [97] | Mice | MyD88 KO | TLR | The absence of MyD88 in mice led to spatial learning and working memory deficits, and reduced motor activity. |
| Ibi et al., 2009 [98] | Mice | Poly IC stimulation | TLR3 | TLR3 stimulation showed decreased social interaction, cognitive, and behavioural changes. |
| Liu et al., 2013 [99] | Mice | TLR7 KO | TLR7 | TLR 7 activation reduced dendrite growth mediated by IL-6 in mice. TLR 7 KO reduced dendrite growth and reduced behavioural activity. |
| Madar et al., 2015 | Mice | TLR2 KO and TLR2+ | TLR2 | TLR2 KO decreased mice activity and impaired spatial learning. Neonatal TLR2/1 activation led to impaired spatial learning in adulthood. |
| Park et al., 2015 | Mice | TLR2 KO | TLR2 | TLR2 KO mice showed cognitive and social recognition impairment. |
| Melnik et al., 2012 | Mice | In utero (poly IC stimulation) | TLR3 mRNA | TLR3 stimulation leads to decreased proliferation after Poly IC treatment unlike, in vitro studies. |
| Mice (DG NPC) | Poly IC treatment | TLR3 mRNA | TLR3 stimulation leads to increased proliferation after 24 and 48 h of poly IC treatment. | |
| Liu et al., 2013 [99] | Mice (cortical neurons) | TLR 7 | TLR7 activation produced cytokines and hindered dendrite growth. TLR7 KO promoted axon and dendrite growth. | |
| Nakajima et al., 2014 [100] | Mice (astrocyte cell culture) | LPS stimulated | TLR3 and 4 mRNA | LPS stimulation showed significantly elevated TLR3 expression up to 24 h and dropped until 72 h. No significant change was seen with TLR4 expression, but a trend of upregulation was observed from 3 to 72 h. |
| Zhou et al., 2020 [101] | Mice (BV2 microglial cells) | LPS treatment | TLR4 protein | TLR4 and NF-kB upregulation post-treatment. |
| Study | Population | Gene (SNPs) | Location of SNP | Region of SNP | SNP Function | Analysis Results |
|---|---|---|---|---|---|---|
| Aflouk et al., 2021 [142] | Tunisian | tlr2 (rs111200466) | Chr4. −196 to −174 Insertion/Deletion | Promoter region | Decreased activation and expression of TLR2 and cytokines | Protective action in females, as this polymorphism was not observed in the schizophrenia group (females). |
| tlr2 (rs5743708) | Chr4. G > A Missense variant | Third exon (functional) | Increases the risk and susceptibility to schizophrenia. | |||
| tlr2 (rs121917864) | Chr4. C > T Missense variant | |||||
| Kang et al., 2013 [143] | Korean | tlr2 (rs3804099) | Chr4. T > C Synonymous Variant | Third exon (functional) | Unknown function | C’ allele associated with cognition (poor concentration). |
| tlr2 (rs3804100) | Chr4. T > A/T > C Missense variant | |||||
| Sharma et al., 2022 [144] | Indian | tlr2 (rs3804099) | Chr4. T > C Synonymous Variant | Third exon (functional) | Unknown function | Increases the risk of schizophrenia. |
| Mostafa et al., 2022 [145] | Egyptian | tlr4 (rs11536889) | Chr9. G > A/G > C UTR Variant | 3′UTR | Interferes with the translation process and increase the expression of TLR4 | A strong association between these genes and schizophrenia. |
| tlr4 (rs1927911) | Chr9. A > C/G/T | Intron | - | |||
| tlr4 (rs1927914) | Chr9. G > A | 5′UTR | Regulation of gene expression and protein levels | |||
| García-Bueno et al., 2016 [66] | Spanish | myd88 (rs7744) | Chr3. A > G/A > T Noncoding Transcript Variant | 3′UTR | Previously associated with autoimmune or inflammatory processes | These two polymorphisms were found to be crucial in the schizophrenia predictive model for this population. |
| il6 (rs1800795) | Chr7. C > G/C > T | Promoter region | Increases IL-6 levels |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patlola, S.R.; Donohoe, G.; McKernan, D.P. Counting the Toll of Inflammation on Schizophrenia—A Potential Role for Toll-like Receptors. Biomolecules 2023, 13, 1188. https://doi.org/10.3390/biom13081188
Patlola SR, Donohoe G, McKernan DP. Counting the Toll of Inflammation on Schizophrenia—A Potential Role for Toll-like Receptors. Biomolecules. 2023; 13(8):1188. https://doi.org/10.3390/biom13081188
Chicago/Turabian StylePatlola, Saahithh Redddi, Gary Donohoe, and Declan P. McKernan. 2023. "Counting the Toll of Inflammation on Schizophrenia—A Potential Role for Toll-like Receptors" Biomolecules 13, no. 8: 1188. https://doi.org/10.3390/biom13081188