
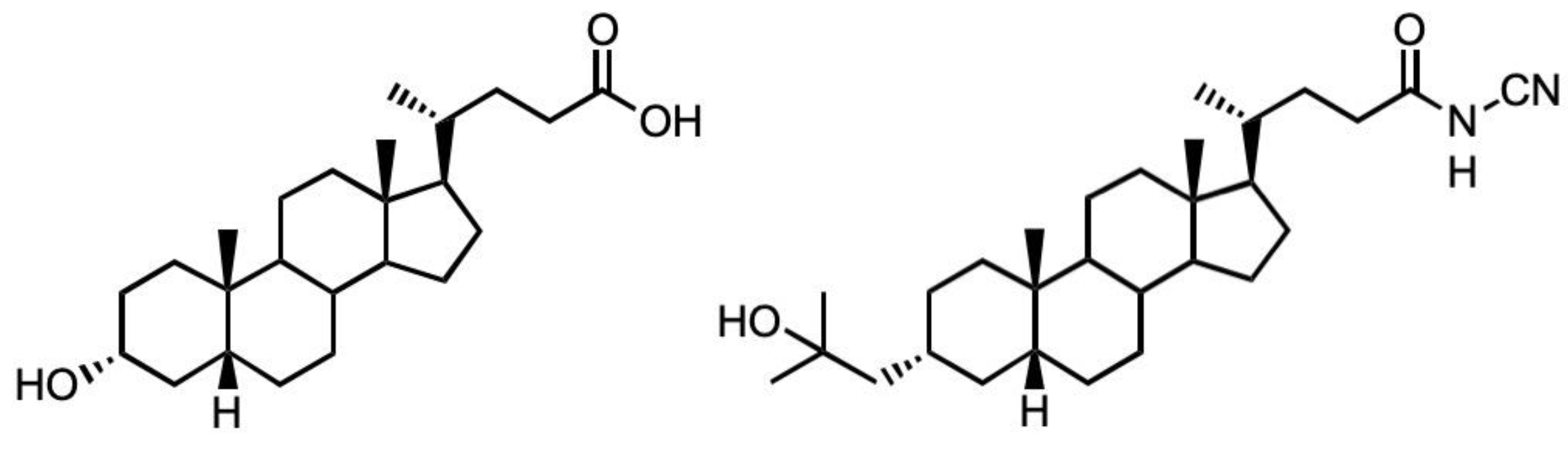
An N-Cyanoamide Derivative of Lithocholic Acid Co-Operates with Lysophosphatidic Acid to Promote Human Osteoblast (MG63) Differentiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Human Osteoblasts
2.3. Osteocalcin Quantification in Conditioned Media
2.4. Cell Number
2.5. ALP Activity
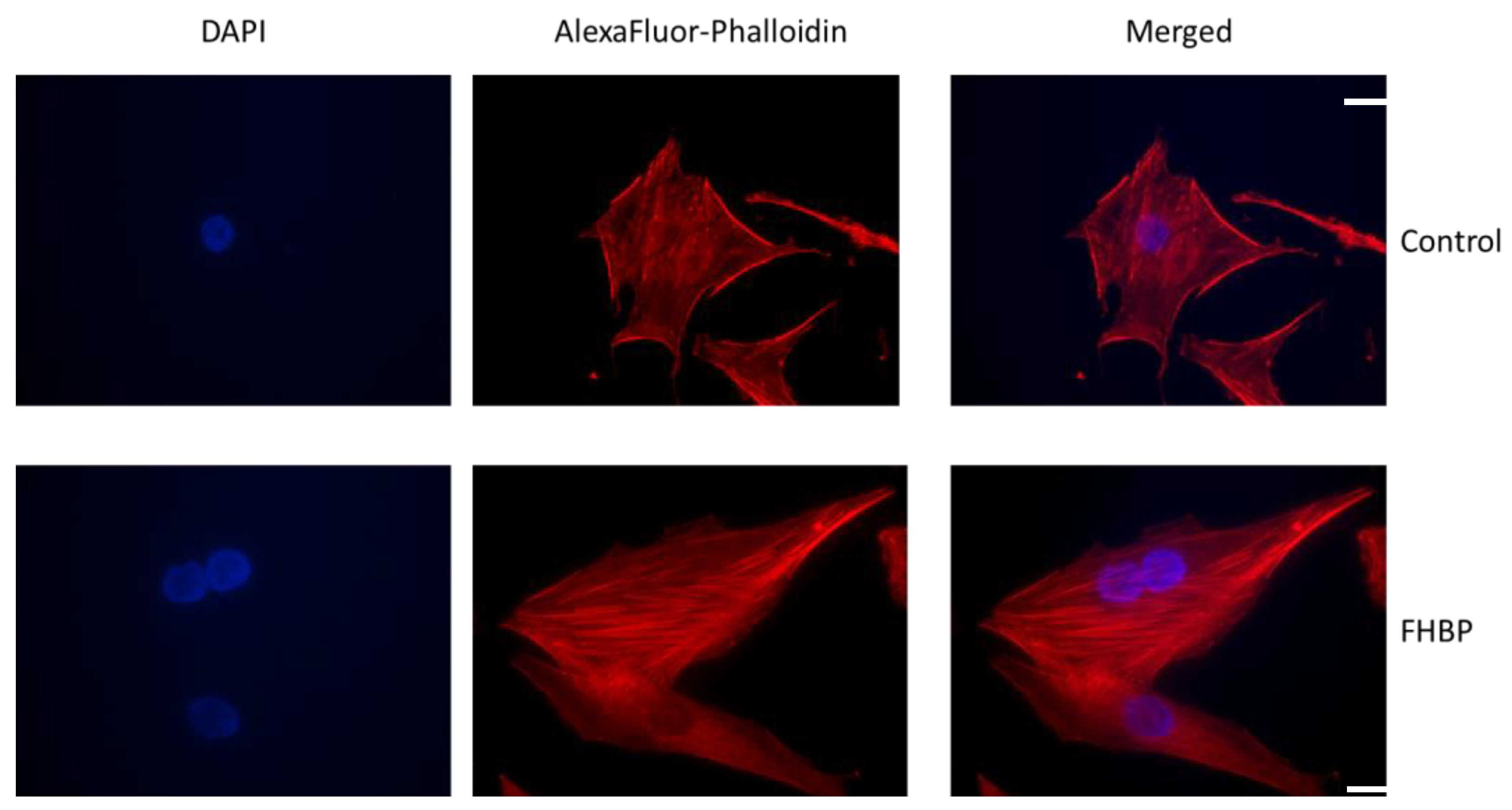
2.6. F-Actin Visualization
2.7. Statistical Analysis
3. Results
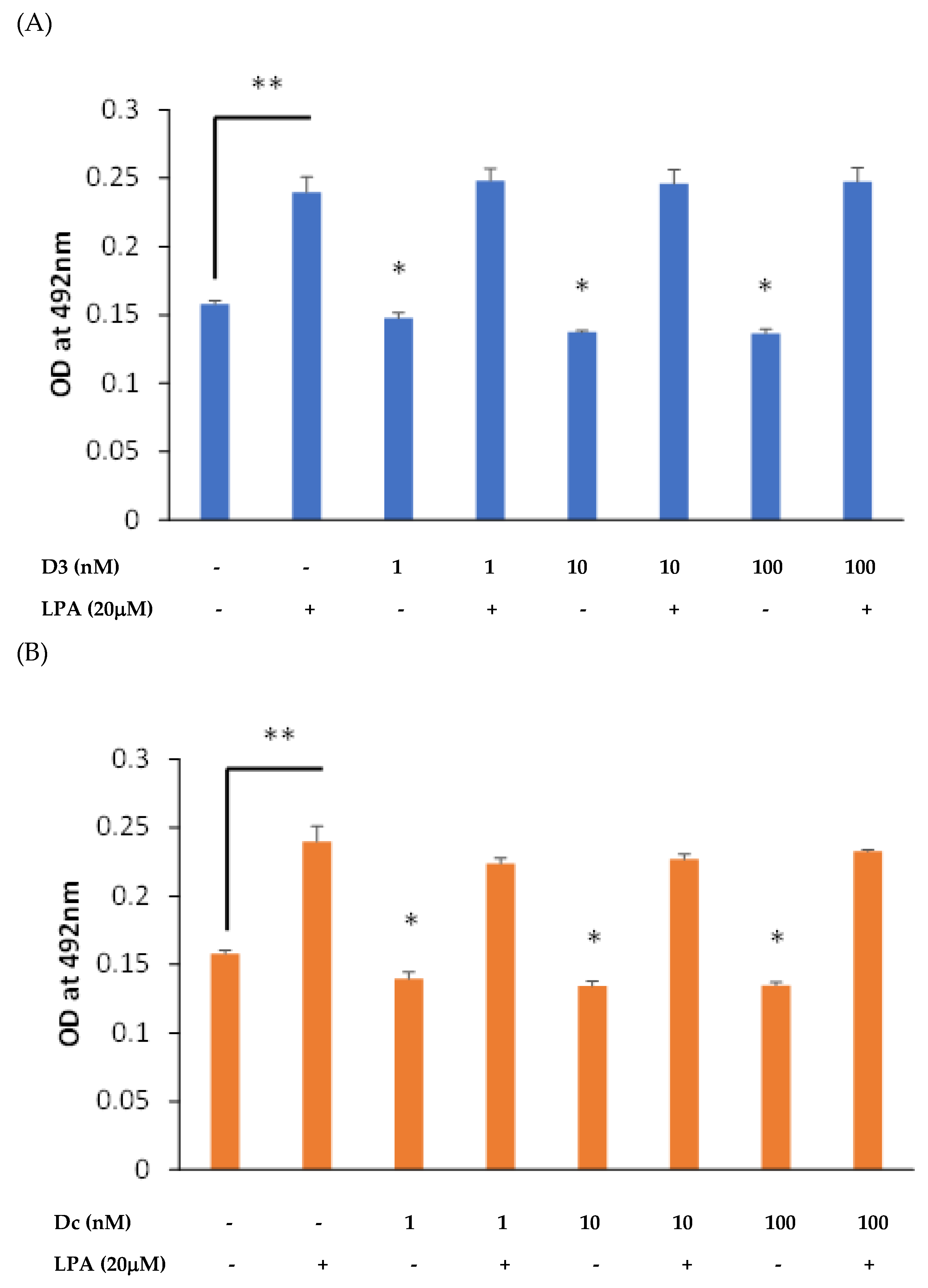
3.1. Both 1,25D and Dcha-150 Modestly Attenuate Osteoblast Growth
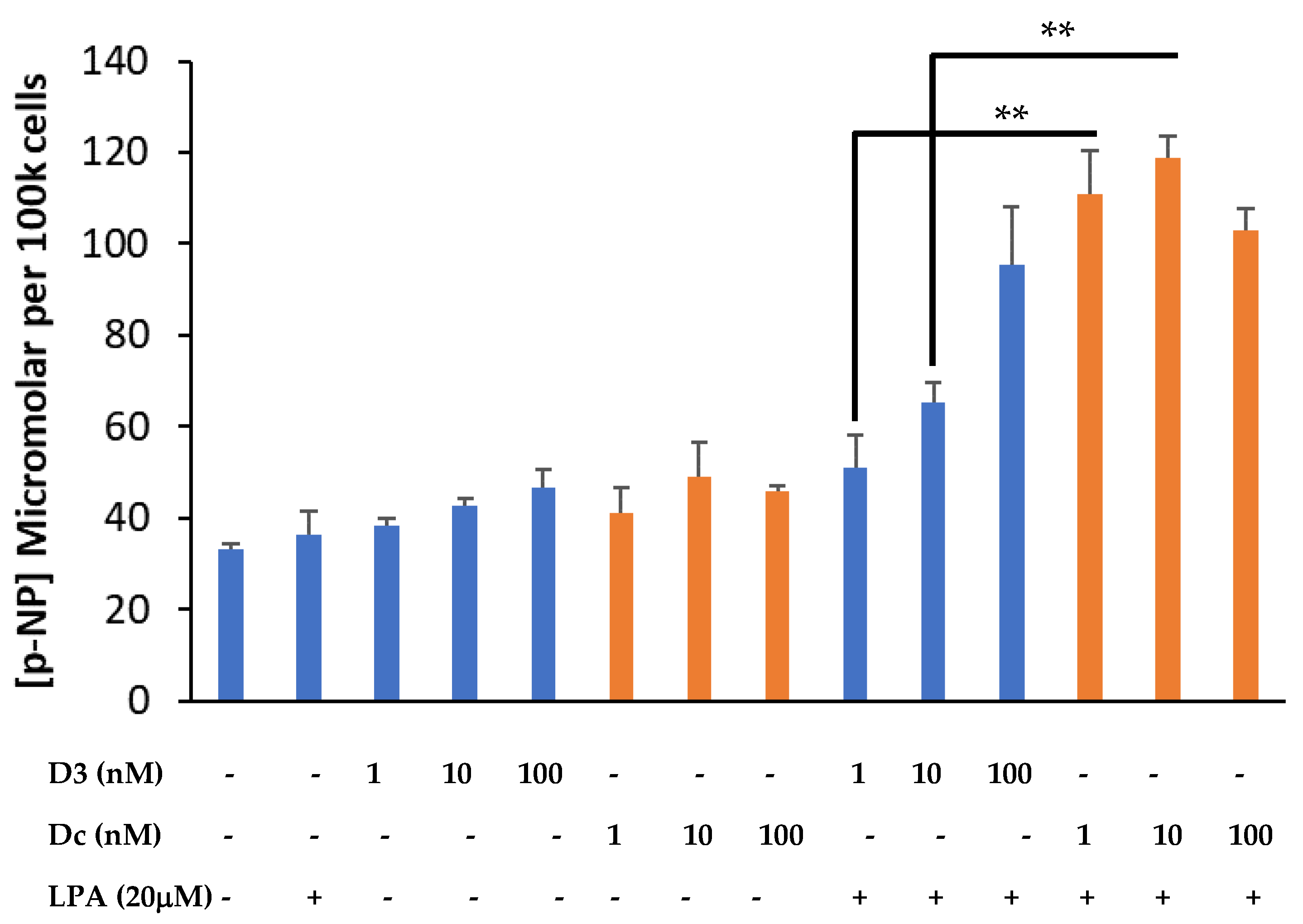
3.2. Dcha-150 Co-Operates with LPA to Enhance Osteoblast Maturation
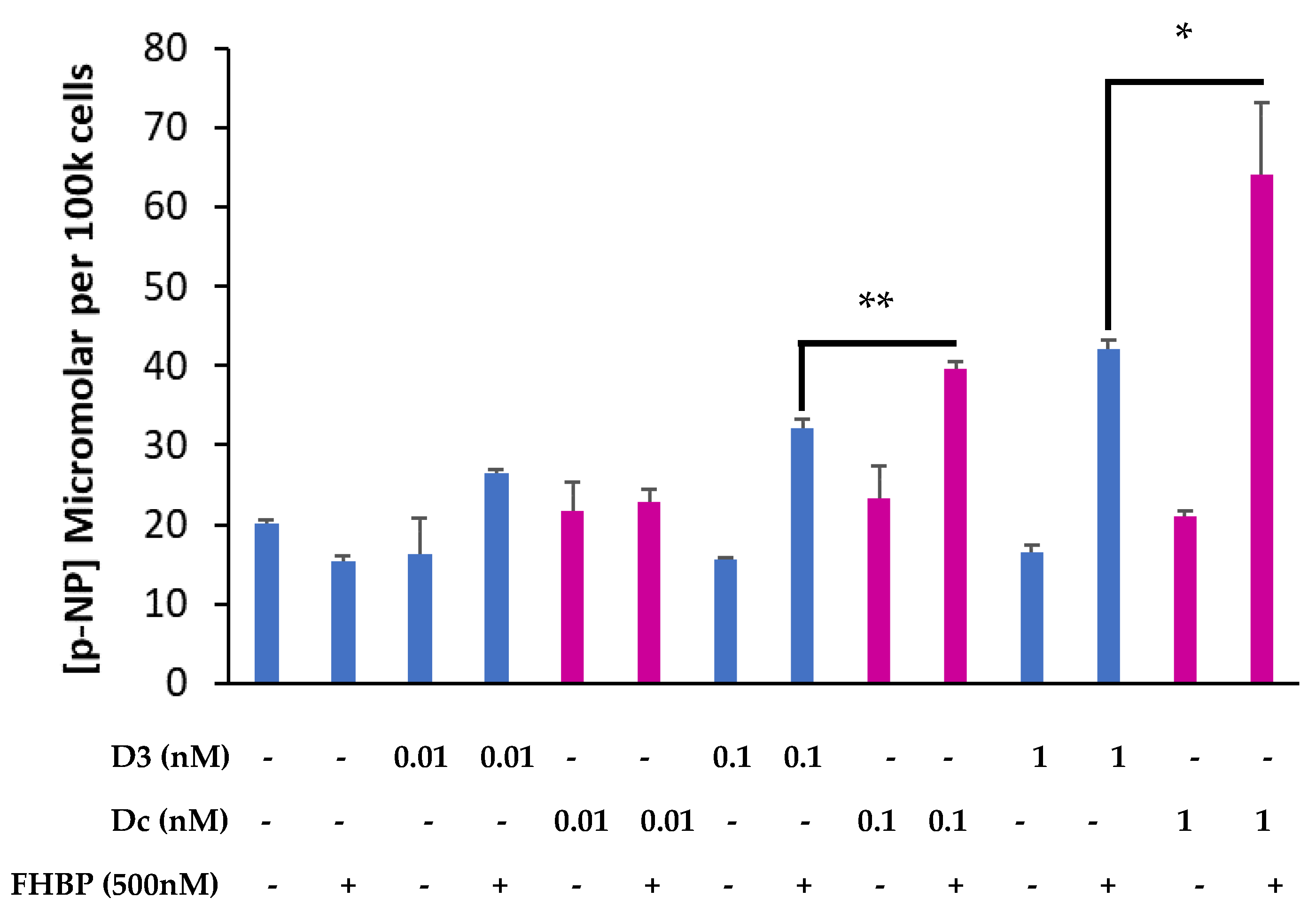
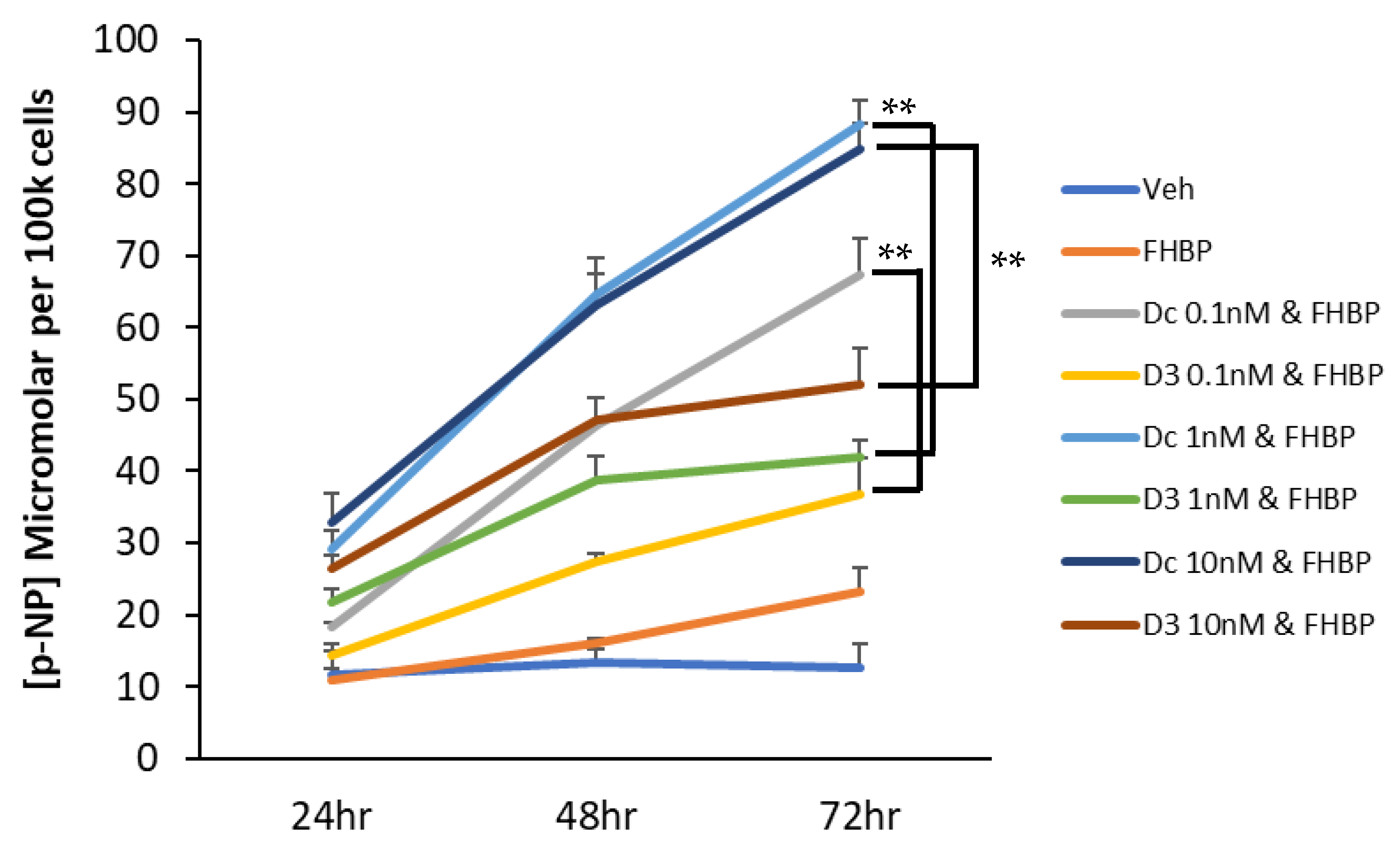
3.3. A Fluoromethylene Analogue of LPA, FHBP, Synergistically Co-Operates with Dcha-150 to Promote Osteoblast Maturation
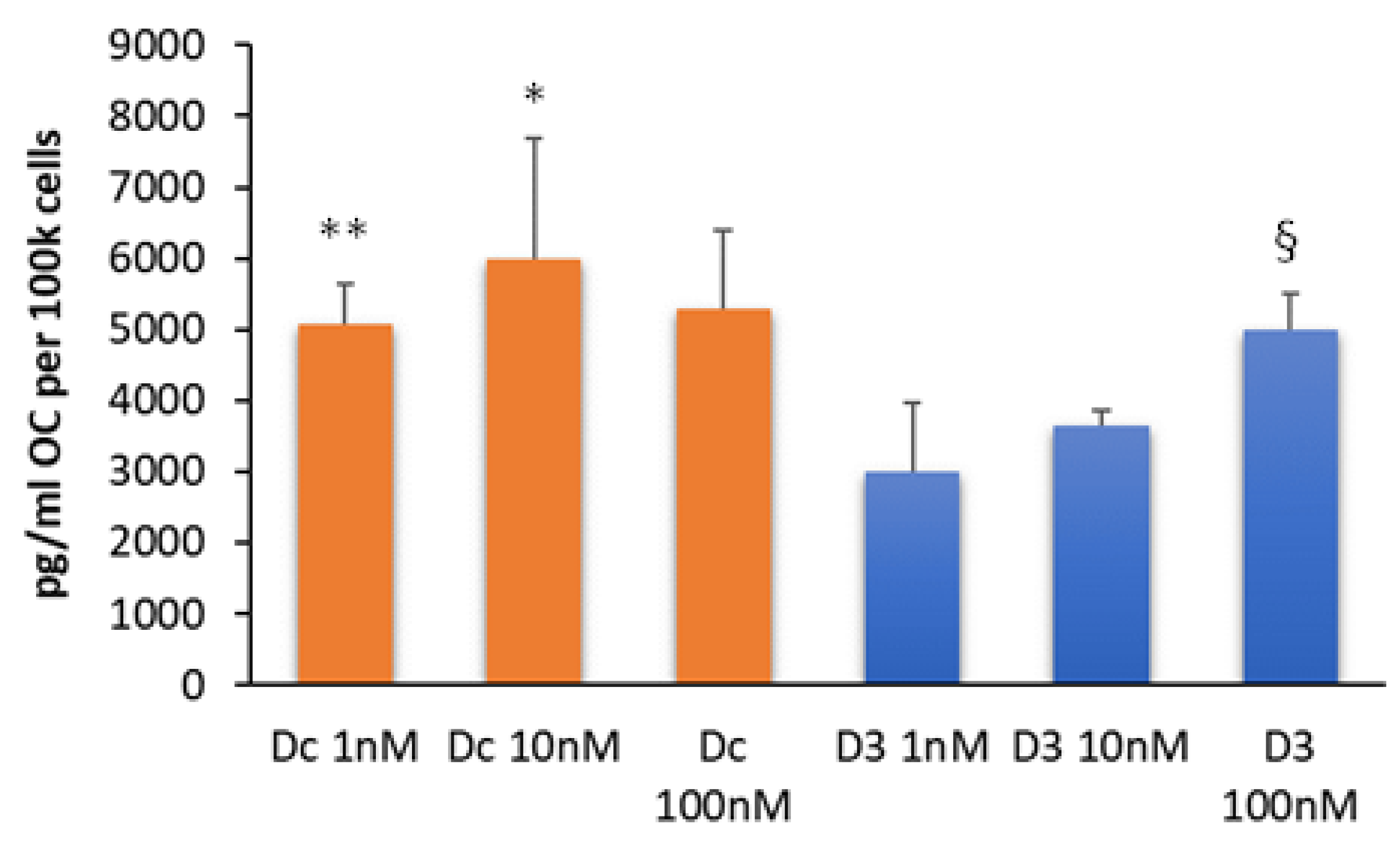
3.4. Dcha-150 Stimulates OC Synthesis
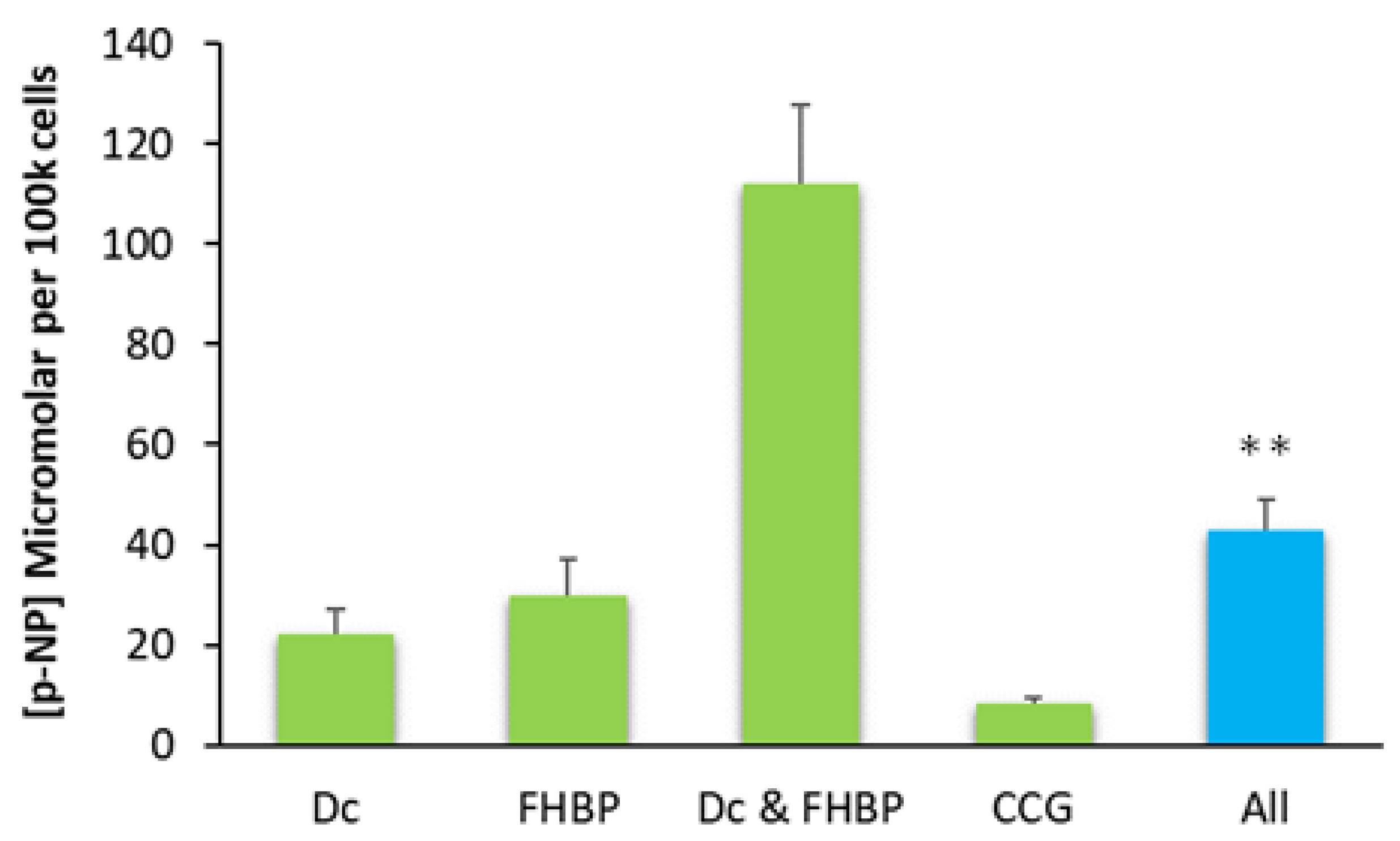
3.5. The Inhibition of MRTF–SRF Prevents MG63 Maturation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berry, J.L.; Davies, M.; Mee, A.P. Vitamin D metabolism, rickets and osteomalacia. Semin. Musculoskelet. Radiol. 2002, 6, 173–182. [Google Scholar] [CrossRef] [PubMed]
- van Driel, M.; Pols, H.A.P.; van Leeuwen, J.P.T.M. Osteoblast differentiation and control by vitamin D and vitamin D metabolites. Curr. Pharmaceut. Des. 2004, 10, 2535–2555. [Google Scholar] [CrossRef] [PubMed]
- van Driel, M.; Koedam, M.; Buurman, C.J.; Roelse, M.; Weyts, F.; Chiba, H.; Uitterlinden, A.G.; Pols, H.A.P.; van Leeuwen, J.P.T.M. Evidence that both 1α,25-dihydroxyvitamin D3 and 24-hydroxylated D3 enhance human osteoblast differentiation and mineralisation. J. Cell. Biochem. 2006, 99, 922–935. [Google Scholar] [CrossRef] [PubMed]
- Makishima, M.; Lu, T.T.; Xie, W.; Whitfield, G.K.; Domoto, H.; Evans, R.M.; Haussler, M.R.; Mangelsdorf, D.J. Vitamin D receptor as an intestinal bile acid sensor. Science 2002, 296, 1313–1316. [Google Scholar] [CrossRef] [Green Version]
- Nehring, J.A.; Zierold, C.; DeLuca, H.F. Lithocholic acid can carry out in vivo functions of vitamin D. Proc. Natl. Acad. Sci. USA 2007, 104, 10006–10009. [Google Scholar] [CrossRef]
- Ishizawa, M.; Matsunawa, M.; Adachi, R.; Uno, S.; Ikeda, K.; Masuno, H.; Shimizu, M.; Iwasaki, K.-I.; Yamada, S.; Makishima, M. Lithocholic acid derivatives act as selective vitamin D receptor modulators without inducing hypercalcaemia. J. Lipid Res. 2008, 49, 763–772. [Google Scholar] [CrossRef] [Green Version]
- Mansell, J.P.; Shorez, D.; Farrar, D.; Nowghani, M. Lithocholate—A promising non-calcaemic calcitriol surrogate for promoting human osteoblast maturation upon biomaterials. Steroids 2009, 74, 963–970. [Google Scholar] [CrossRef]
- Whyte, M.P. Physiological role of alkaline phosphatase explored in hypophosphatasia. Ann. N. Y. Acad. Sci. 2010, 1192, 190–200. [Google Scholar] [CrossRef]
- Wagner, E.F. Functions of AP1 (Fos/Jun) in bone development. Ann. Rheum. Dis. 2002, 61, 40–42. [Google Scholar] [CrossRef]
- Marie, P.J. Transcription factors controlling osteoblastogenesis. Arch. Biochem. Biophys. 2008, 473, 98–105. [Google Scholar] [CrossRef]
- Yoshihara, A.; Kawasaki, H.; Masuno, H.; Takada, K.; Numoto, N.; Ito, N.; Hirata, N.; Kanda, Y.; Ishizawa, M.; Makishima, M.; et al. Lithocholic Acid Amides as Potent Vitamin D Receptor Agonists. Biomolecules 2022, 12, 130. [Google Scholar] [CrossRef]
- Ayre, W.N.; Scott, T.; Hallam, K.; Blom, A.W.; Denyer, S.; Bone, H.K.; Mansell, J.P. Fluorophosphonate-functionalised titanium via a pre-adsorbed alkane phosphonic acid: A novel dual action surface finish for bone regenerative applications. J. Mater. Sci. Mater. Med. 2016, 27, 36. [Google Scholar] [CrossRef]
- Neary, G.; Blom, A.W.; Shiel, A.I.; Wheway, G.; Mansell, J.P. Development and biological evaluation of fluorophosphonate-modified hydroxyapatite for orthopaedic applications. J. Mater. Med. Mater. Med. 2018, 29, 122. [Google Scholar] [CrossRef] [Green Version]
- Shiel, A.I.; Ayre, W.N.; Blom, A.W.; Hallam, K.R.; Heard, P.J.; Payton, O.; Picco, L.; Mansell, J.P. Development of a facile fluorophosphonate-functionalised titanium surface for potential orthopaedic applications. J. Orthop. Trans. 2020, 23, 140–151. [Google Scholar] [CrossRef]
- Baldwin, F.; Craig, T.J.; Shiel, A.I.; Cox, T.; Lee, K.; Mansell, J.P. Polydopamine-Lysophosphatidate-Functionalised Titanium: A Novel Hybrid Surface Finish for Bone Regenerative Applications. Molecules 2020, 25, 1583. [Google Scholar] [CrossRef] [Green Version]
- Heremans, H.; Billiau, A.; Caissiman, J.J.; Mulier, J.C.; de Somer, P. In vitro cultivation of human tumour tissues II. Morphological and virological characterisation of three cell lines. Oncology 1978, 35, 246–252. [Google Scholar] [CrossRef]
- Clover, J.; Gowen, M. Are MG-63 and HOS TE85 human osteosarcoma cell lines representative models of the osteoblastic phenotype? Bone 1994, 15, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Gidley, J.; Openshaw, S.; Pring, E.T.; Sale, S.; Mansell, J.P. Lysophosphatidic acid cooperates with 1α,25(OH)2D3 in stimulating human MG63 osteoblast maturation. Prostaglandins Other Lipid Mediat. 2006, 80, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Mansell, J.P.; Farrar, D.; Jones, S.; Nowghani, M. Cytoskeletal reorganisation, 1α,25-dihydroxy vitamin D3 and human MG63 osteoblast maturation. Mol. Cell. Endocrinol. 2009, 305, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, R.; Honma, Y.; Masuno, H.; Kawana, K.; Shimomura, I.; Yamada, S.; Makishima, M. Selective activation of vitamin D receptor by lithocholic acid acetate, a bile acid derivative. J. Lipid. Res. 2005, 46, 46–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komori, T. Functions of Osteocalcin in Bone, Pancreas, Testis, and Muscle. Int. J. Mol. Sci. 2020, 21, 7513. [Google Scholar] [CrossRef] [PubMed]
- Ozono, K.; Liao, J.; Kerner, S.A.; Scott, R.A.; Pike, J.W. The Vitamin D-responsive Element in the Human Osteocalcin Gene. Association With a Nuclear Proto-Oncogene Enhancer. J. Biol. Chem. 1990, 265, 21881–21888. [Google Scholar] [CrossRef]
- Orimo, H.; Shimada, T. Posttranscriptional modulation of the human tissue–nonspecific alkaline phosphatase gene expression by 1,25-dihydroxyvitamin D3 in MG-63 osteoblastic osteosarcoma cells. Nut. Res. 2006, 26, 227–234. [Google Scholar] [CrossRef]
- Mansell, J.P. Convergence of vitamin D and lysophosphatidic acid signaling in stimulating human osteoblast maturation. Front. Physiol. 2014, 5, 263. [Google Scholar] [CrossRef] [Green Version]
- Ridley, A.J.; Hall, A. The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef]
- Ridley, A.J.; Hall, A. Signal transduction pathways regulating Rho-mediated stress fibre formation: Requirement for a tyrosine kinase. EMBO J. 1994, 13, 2600–2610. [Google Scholar] [CrossRef]
- Ridley, A.J. Rho family proteins: Coordinating cell responses. Trends Cell Biol. 2001, 11, 471–477. [Google Scholar] [CrossRef]
- Gineitis, D.; Treisman, R. Differential usage of signal transduction pathways defines two types of serum response factor target gene. J. Biol. Chem. 2001, 276, 24531–24539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miralles, F.; Posern, G.; Zaromytidou, A.-I.; Treisman, R. Actin dynamics control SRF activity by regulation of its coactivator MAL. Cell 2003, 113, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuwahara, K.; Barrientos, G.C.; Pipes, G.C.; Li, S.; Olson, E.N. Muscle-specific signaling mechanism that links actin dynamics to serum response factor. Mol. Cell. Biol. 2005, 25, 3173–3181. [Google Scholar] [CrossRef] [Green Version]
- Pipes, G.C.; Creemers, E.E.; Olson, E.N. The myocardin family of transcriptional coactivators: Versatile regulators of cell growth, migration, and myogenesis. Genes Dev. 2006, 20, 1545–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, E.N.; Nordheim, A. Linking actin dynamics and gene transcription to drive cellular motile functions. Nat. Rev. Mol. Cell. Biol. 2010, 11, 353–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchings, K.M.; Lisabeth, E.M.; Rajeswaran, W.; Wilson, M.W.; Sorenson, R.J.; Campbell, P.L.; Ruth, J.H.; Amin, A.; Tsou, P.-S.; Leipprandt, J.R.; et al. Pharmacokinetic Optimization of CCG-203971: Novel Inhibitors of the Rho/MRTF/SRF Transcriptional Pathway as Potential Antifibrotic Therapeutics for Systemic Scleroderma. Bioorg. Med. Chem. Lett. 2017, 27, 1744–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyväri, L.; Vanhatupa, S.; Halonen, H.T.; Kääriäinen, M.; Miettinen, S. Myocardin-Related Transcription Factor A (MRTF-A) Regulates the Balance between Adipogenesis and Osteogenesis of Human Adipose Stem Cells. Stem Cells Int. 2020, 2020, 8853541. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mansell, J.P.; Tanatani, A.; Kagechika, H. An N-Cyanoamide Derivative of Lithocholic Acid Co-Operates with Lysophosphatidic Acid to Promote Human Osteoblast (MG63) Differentiation. Biomolecules 2023, 13, 1113. https://doi.org/10.3390/biom13071113
Mansell JP, Tanatani A, Kagechika H. An N-Cyanoamide Derivative of Lithocholic Acid Co-Operates with Lysophosphatidic Acid to Promote Human Osteoblast (MG63) Differentiation. Biomolecules. 2023; 13(7):1113. https://doi.org/10.3390/biom13071113
Chicago/Turabian StyleMansell, Jason P., Aya Tanatani, and Hiroyuki Kagechika. 2023. "An N-Cyanoamide Derivative of Lithocholic Acid Co-Operates with Lysophosphatidic Acid to Promote Human Osteoblast (MG63) Differentiation" Biomolecules 13, no. 7: 1113. https://doi.org/10.3390/biom13071113