
Characterization of Rickets Type II Model Rats to Reveal Functions of Vitamin D and Vitamin D Receptor

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Diets
2.3. HE Staining
2.4. Preparation of Primary Culture Cells
2.5. Immunofluorescence Staining
2.6. Western Blot Analysis
2.7. Computed Tomography
2.8. Measurement of Calcium Metabolism Parameters in Plasma
2.9. Statistical Analysis
3. Results
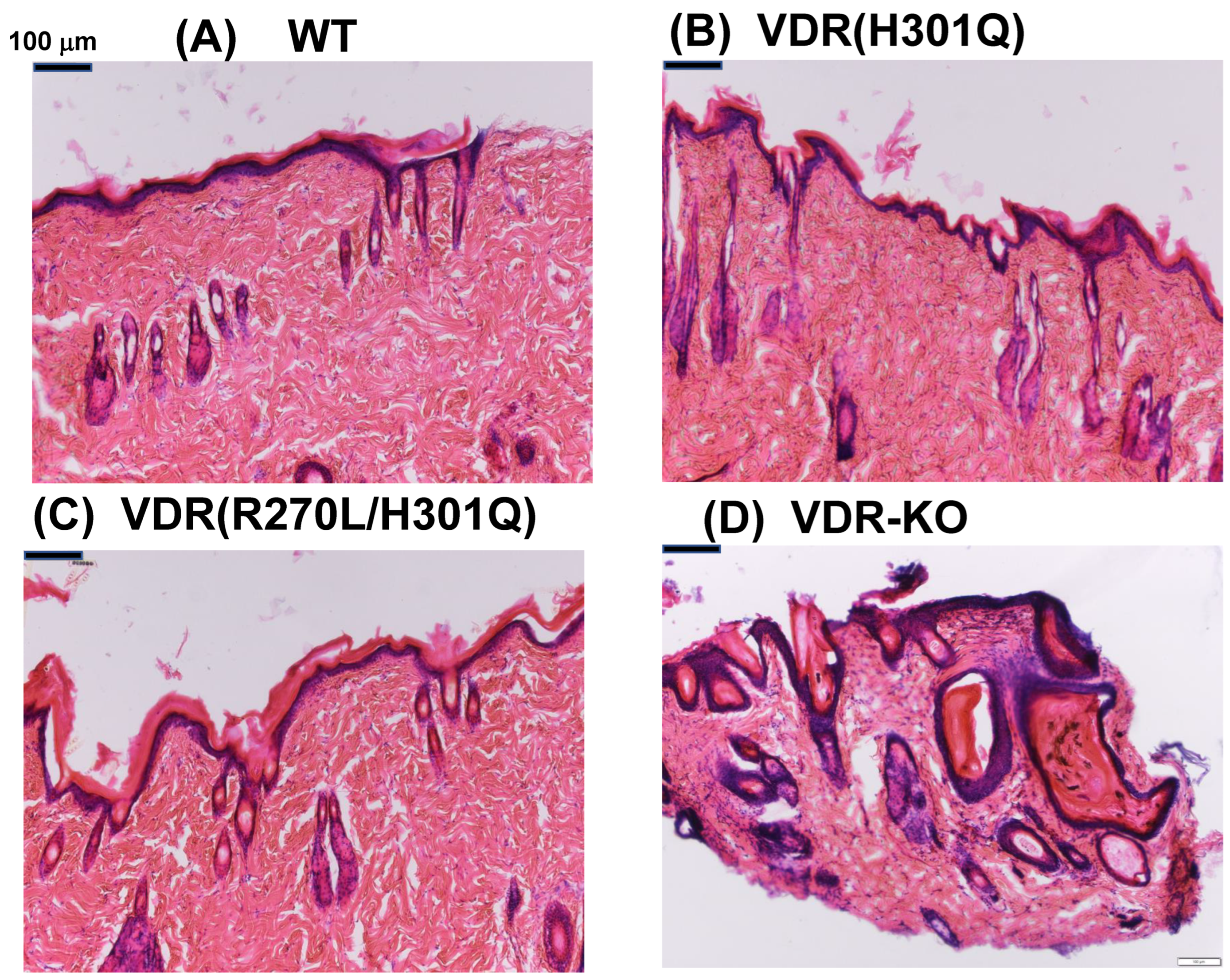
3.1. HE Staining of Dorsal Skin
3.2. Western Blot Analysis
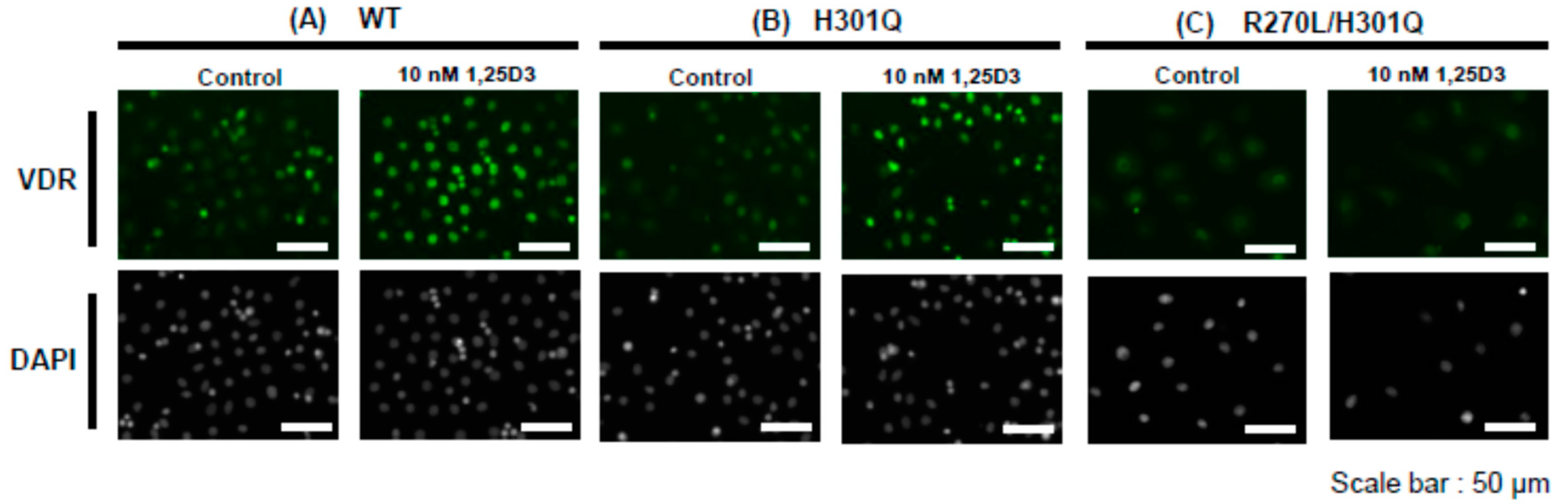
3.3. Nuclear Translocation of VDR in the Presence of 10 nM of 1,25D3
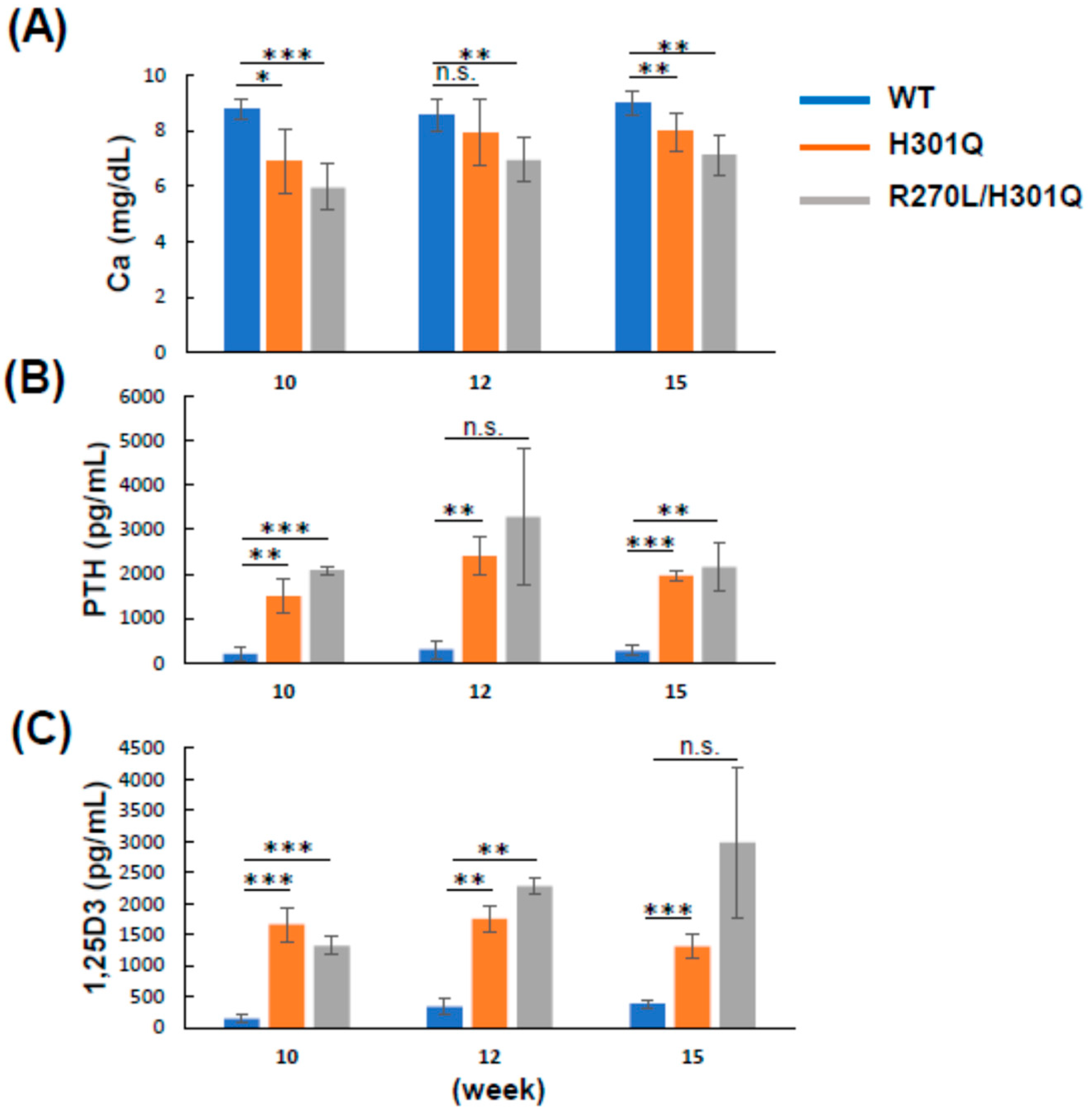
3.4. Plasma Ca, PTH, and 1,25D3 Levels in WT and GM Rats
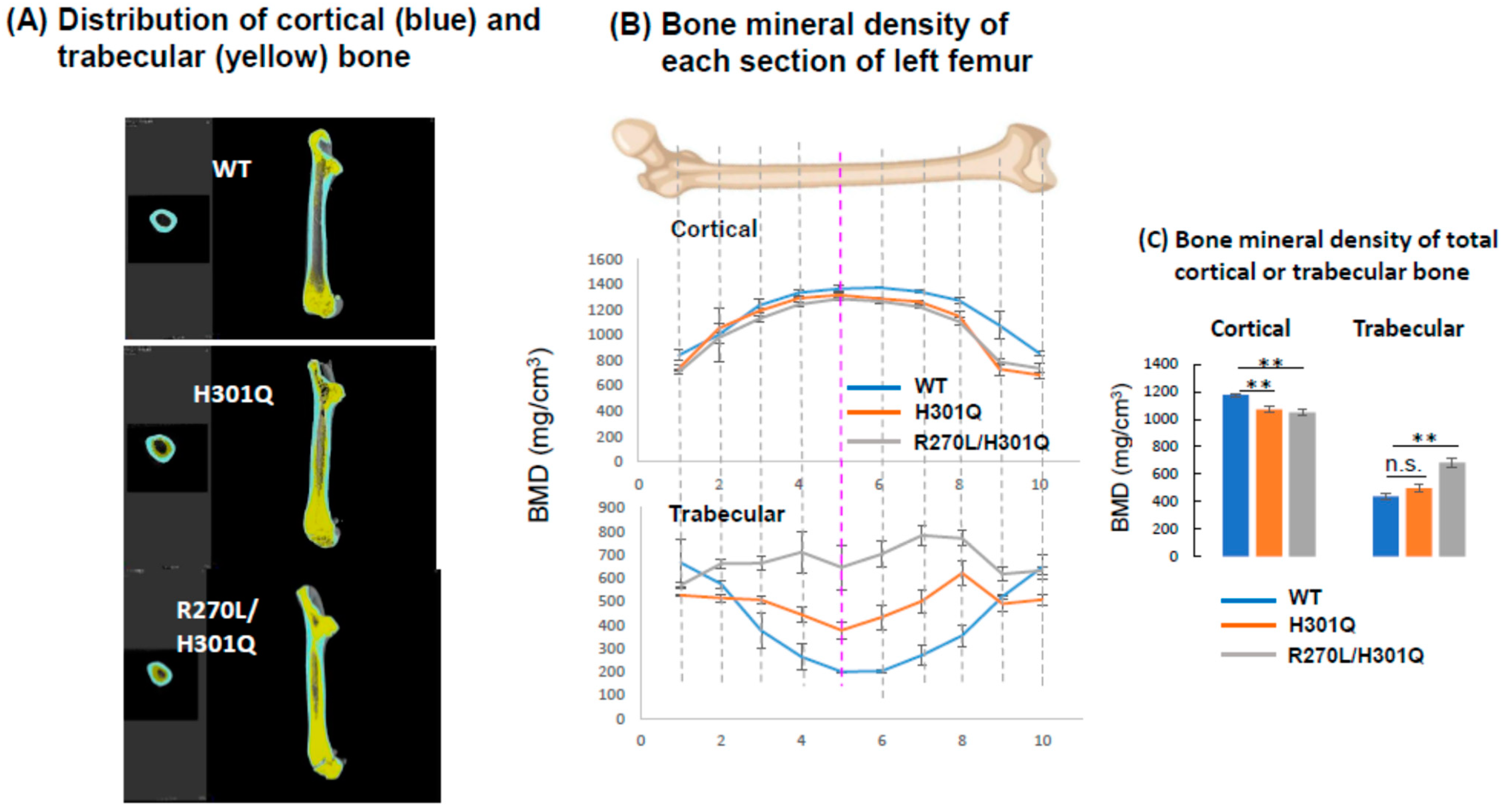
3.5. Bone Formation and Bone Mineral Density in WT and GM Rats
4. Discussion
4.1. Hair and Skin
4.2. Bone Formation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malloy, P.J.; Feldman, D. The role of vitamin D receptor mutations in the development of alopecia. Mol. Cell. Endocrinol. 2011, 347, 90–96. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Malloy, P.J. Mutations in the vitamin D receptor and hereditary vitamin D-resistant rickets. Bonekey Rep. 2014, 3, 510. [Google Scholar] [CrossRef] [PubMed]
- Arita, K.; Nanda, A.; Wessagowit, V.; Akiyama, M.; Alsaleh, Q.A.; McGrath, J.A. A novel mutation in the VDR gene in hereditary vitamin D-resistant rickets. Br. J. Dermatol. 2008, 158, 168–171. [Google Scholar] [CrossRef] [PubMed]
- Tamura, M.; Ishizawa, M.; Isojima, T.; Özen, S.; Oka, A.; Makishima, M.; Kitanaka, S. Functional analyses of a novel missense and other mutations of the vitamin D receptor in association with alopecia. Sci. Rep. 2017, 7, 5102. [Google Scholar] [CrossRef] [PubMed]
- Norman, A.W.; Bishop, J.E.; Collins, E.D.; Seo, E.G.; Satchell, D.P.; Dormanen, M.C.; Zanello, S.B.; Farach-Carson, M.C.; Bouillon, R.; Okamura, W.H. Differing shapes of 1 alpha,25-dihydroxyvitamin D3 function as ligands for the D-binding protein, nuclear receptor and membrane receptor: A status report. J. Steroid Biochem. Mol. Biol. 1996, 56, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Mizwicki, M.T.; Norman, A.W. The vitamin D sterol-vitamin D receptor ensemble model offers unique insights into both genomic and rapid-response signaling. Sci. Signal. 2009, 2, re4. [Google Scholar] [CrossRef]
- Haussler, M.R.; Whitfield, G.K.; Kaneko, I.; Haussler, C.A.; Hsieh, D.; Hsieh, J.C.; Jurutka, P.W. Molecular mechanisms of vitamin D action. Calcif. Tissue Int. 2013, 92, 77–98. [Google Scholar] [CrossRef]
- Nishikawa, M.; Yasuda, K.; Takamatsu, M.; Abe, K.; Okamoto, K.; Horibe, K.; Mano, H.; Nakagawa, K.; Tsugawa, N.; Hirota, Y.; et al. Generation of Novel Genetically Modified Rats to Reveal the Molecular Mechanisms of Vitamin D Actions. Sci. Rep. 2020, 10, 5677. [Google Scholar] [CrossRef]
- Nishikawa, M.; Murose, N.; Mano, H.; Yasuda, K.; Isogai, Y.; Kittaka, A.; Takano, M.; Ikushiro, S.; Sakaki, T. Robust osteogenic efficacy of 2α-heteroarylalkyl vitamin D analogue AH-1 in VDR (R270L) hereditary vitamin D-dependent rickets model rats. Sci. Rep. 2022, 12, 12517. [Google Scholar] [CrossRef]
- Kise, S.; Iijima, A.; Nagao, C.; Okada, T.; Mano, H.; Nishikawa, M.; Ikushiro, S.; Kanemoto, Y.; Kato, S.; Nakanishi, T.; et al. Functional analysis of vitamin D receptor (VDR) using adenovirus vector. J. Steroid Biochem. Mol. Biol. 2023, 230, 106275. [Google Scholar] [CrossRef]
- Mano, H.; Ikushiro, S.; Saito, N.; Kittaka, A.; Sakaki, T. Development of a highly sensitive in vitro system to detect and discriminate between vitamin D receptor agonists and antagonists based on split-luciferase technique. J. Steroid Biochem. Mol. Biol. 2018, 178, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; Etten, E.V.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and Human Health: Lessons from Vitamin D Receptor Null Mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [PubMed]
- Hsieh, J.C.; Sisk, J.M.; Jurutka, P.W.; Haussler, C.A.; Slater, S.A.; Haussler, M.R.; Thompson, C.C. Physical and functional interaction between the vitamin D receptor and hairless corepressor, two proteins required for hair cycling. J. Biol. Chem. 2003, 278, 38665–38674. [Google Scholar] [CrossRef]
- Luderer, H.F.; Gori, F.; Demay, M.B. Lymphoid enhancer-binding factor-1 (LEF1) interacts with the DNA-binding domain of the vitamin D receptor. J. Biol. Chem. 2011, 286, 18444–18451. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Byrne, C.; Jacobs, J.; Fuchs, E. Lymphoid enhancer factor 1 directs hair follicle patterning and epithelial cell fate. Genes Dev. 1995, 9, 700–713. [Google Scholar] [CrossRef]
- Chen, C.H.; Sakai, Y.; Demay, M.B. Targeting expression of the human vitamin D receptor to the keratinocytes of vitamin D receptor null mice prevents alopecia. Endocrinology 2001, 142, 5386–5389. [Google Scholar] [CrossRef]
- Teichert, A.; Elalieh, H.; Bikle, D. Disruption of the hedgehog signaling pathway contributes to the hair follicle cycling deficiency in Vdr knockout mice. J. Cell. Physiol. 2010, 225, 482–489. [Google Scholar] [CrossRef]
- Joko, Y.; Yamamoto, Y.; Kato, S.; Takemoto, T.; Abe, M.; Matsumoto, T.; Fukumoto, S.; Sawatsubashi, S. VDR is an essential regulator of hair follicle regression through the progression of cell death. Life Sci. Alliance 2023, 6, e202302014. [Google Scholar] [CrossRef]
- Morita, R.; Sanzen, N.; Sasaki, H.; Hayashi, T.; Umeda, M.; Yoshimura, M.; Yamamoto, T.; Shibata, T.; Abe, T.; Kiyonari, H.; et al. Tracing the origin of hair follicle stem cells. Nature 2021, 594, 547–552. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Lieberherr, M.; Fritsch, J.; Guillozo, H.; Alvarez, M.L.; Fitouri, Z.; Jehan, F.; Garabédian, M. The rapid effects of 1,25-dihydroxyvitamin D3 require the vitamin D receptor and influence 24-hydroxylase activity: Studies in human skin fibroblasts bearing vitamin D receptor mutations. J. Biol. Chem. 2004, 279, 7591–7597. [Google Scholar] [CrossRef]
- Nemere, I.; Garbi, N.; Hämmerling, G.J.; Khanal, R.C. Intestinal cell calcium uptake and the targeted knockout of the 1,25D3-MARRS (membrane-associated, rapid response steroid-binding) receptor/PDIA3/Erp57. J. Biol. Chem. 2010, 285, 31859–31866. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rat Strain | VDR-Dependent 1,25D3 | VDR-Independent 1,25D3 | VDR-Dependent 25D3 | VDR-Independent 25D3 | Unliganded VDR |
|---|---|---|---|---|---|
| WT | ◯ | ◯ | ◯ | ◯ | ◯ |
| Vdr(R270L) | × | ◯ * | ◯ | ◯ | ◯ |
| Cyp27b1-KO | × | × | ◯ | ◯ | ◯ |
| Vdr(H301Q) | △ | ◯ * | △ | ◯ | ◯ |
| Vdr(R270L/H301Q) | × | ◯ * | × | ◯ | ◯ |
| Vdr-KO | × | ◯ * | × | ◯ | × |
| VDR | Relative Affinity for 1,25D3 | Relative Affinity for 25D3 | Nuclear Transfer 10 nM 1,25D3 | Nuclear Transfer 100 nM 25D3 | Induction of Cyp24a1 10 nM 1,25D3 |
|---|---|---|---|---|---|
| WT | 100 | 100 | Yes | Yes | Yes |
| H301Q | 0.2 | 0.2 | Yes | partially Yes | Yes |
| R270L/H301Q | <0.001 | <0.1 | No | No | No |
| Rat Strain. | Ca (mg/dL) | PTH (pg/mL) | 1,25D3 (pg/mL) |
|---|---|---|---|
| WT | 8.56 ± 0.57 | 301.5 ± 191.3 | 344.0 ± 130.0 |
| Vdr(H301Q) | 7.93 ± 1.17 n.s. | 2397.0 ± 419.5 ** | 1750.7 ± 210.9 ** |
| Vdr(R270L/H301Q) | 6.93 ± 0.79 ** | 3282.5 ± 1539.1 n.s. | 2272.7 ± 127.2 ** |
| Rat Strain | Femur Length (cm) | Femur Max Width (cm) | Bone Volume (cm3) | BMD Cortical (mg/cm3) | BMD Trabecular (mg/cm3) |
|---|---|---|---|---|---|
| WT | 3.9 ± 1.8 | 7.9 ± 0.10 | 0.55 ± 0.06 | 1178.6 ± 11.0 | 438.5 ± 19.0 |
| Vdr(H301Q) | 3.3 ± 0.8 ** | 8.3 ± 0.03 n.s. | 0.47 ± 0.03 n.s. | 1071.4 ± 19.4 ** | 496.9 ± 24.2 n.s. |
| Vdr(R270L/H301Q) | 3.2 ± 1.1 ** | 8.6 ± 0.01 * | 0.54 ± 0.01 n.s. | 1048.7 ± 21.5 ** | 683.5 ± 34.7 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwai, Y.; Iijima, A.; Kise, S.; Nagao, C.; Senda, Y.; Yabu, K.; Mano, H.; Nishikawa, M.; Ikushiro, S.; Yasuda, K.; et al. Characterization of Rickets Type II Model Rats to Reveal Functions of Vitamin D and Vitamin D Receptor. Biomolecules 2023, 13, 1666. https://doi.org/10.3390/biom13111666
Iwai Y, Iijima A, Kise S, Nagao C, Senda Y, Yabu K, Mano H, Nishikawa M, Ikushiro S, Yasuda K, et al. Characterization of Rickets Type II Model Rats to Reveal Functions of Vitamin D and Vitamin D Receptor. Biomolecules. 2023; 13(11):1666. https://doi.org/10.3390/biom13111666
Chicago/Turabian StyleIwai, Yuichiro, Ayano Iijima, Satoko Kise, Chika Nagao, Yuto Senda, Kana Yabu, Hiroki Mano, Miyu Nishikawa, Shinichi Ikushiro, Kaori Yasuda, and et al. 2023. "Characterization of Rickets Type II Model Rats to Reveal Functions of Vitamin D and Vitamin D Receptor" Biomolecules 13, no. 11: 1666. https://doi.org/10.3390/biom13111666