
Bcl-xL Is Spontaneously Inserted into Preassembled Nanodiscs and Stimulates Bax Insertion in a Cell-Free Protein Synthesis System
 and
and 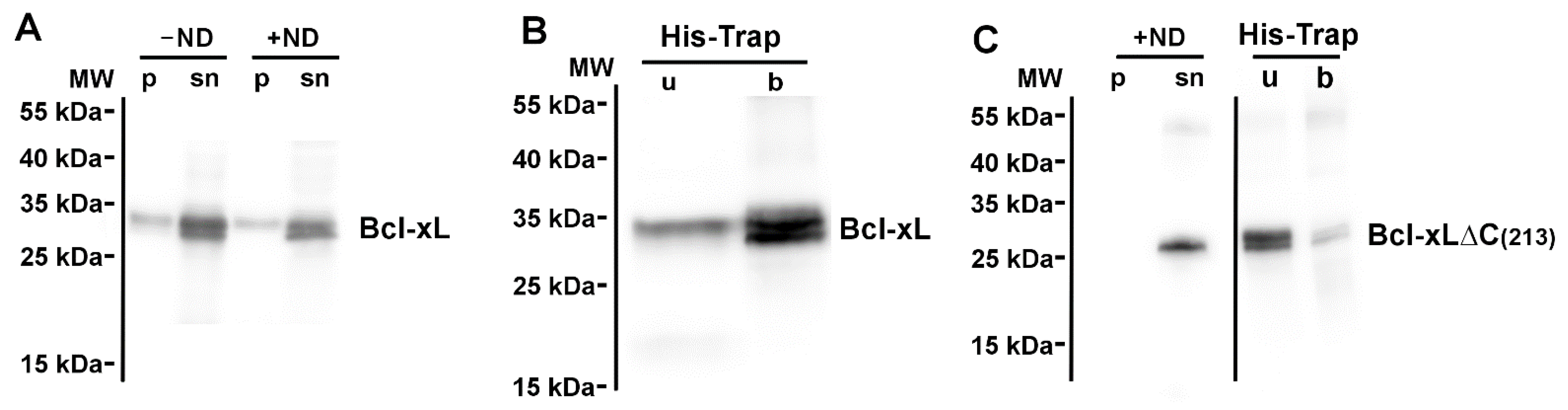{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning of Full-Length Bcl-xL and Bax
2.2. Production of the Scaffold Protein His7-MSP1E3D1
2.3. Nanodisc Formation
2.4. Cell-Free Protein Synthesis and Protein Insertion into Nanodiscs
2.5. MALDI-MS Analysis
2.6. Miscellaneous
3. Results
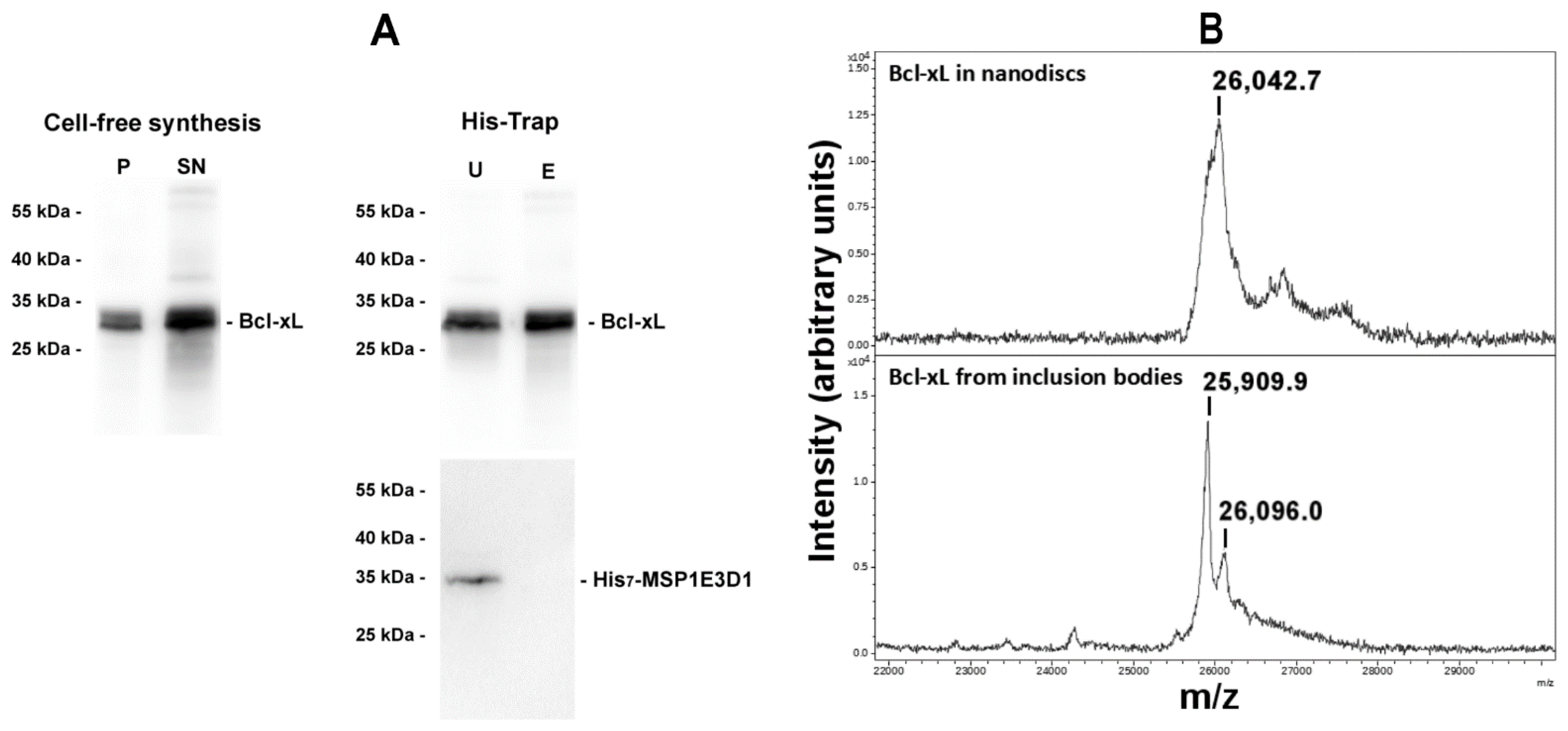
3.1. Neosynthesized Bcl-xL Is Spontaneously Inserted into Nanodiscs
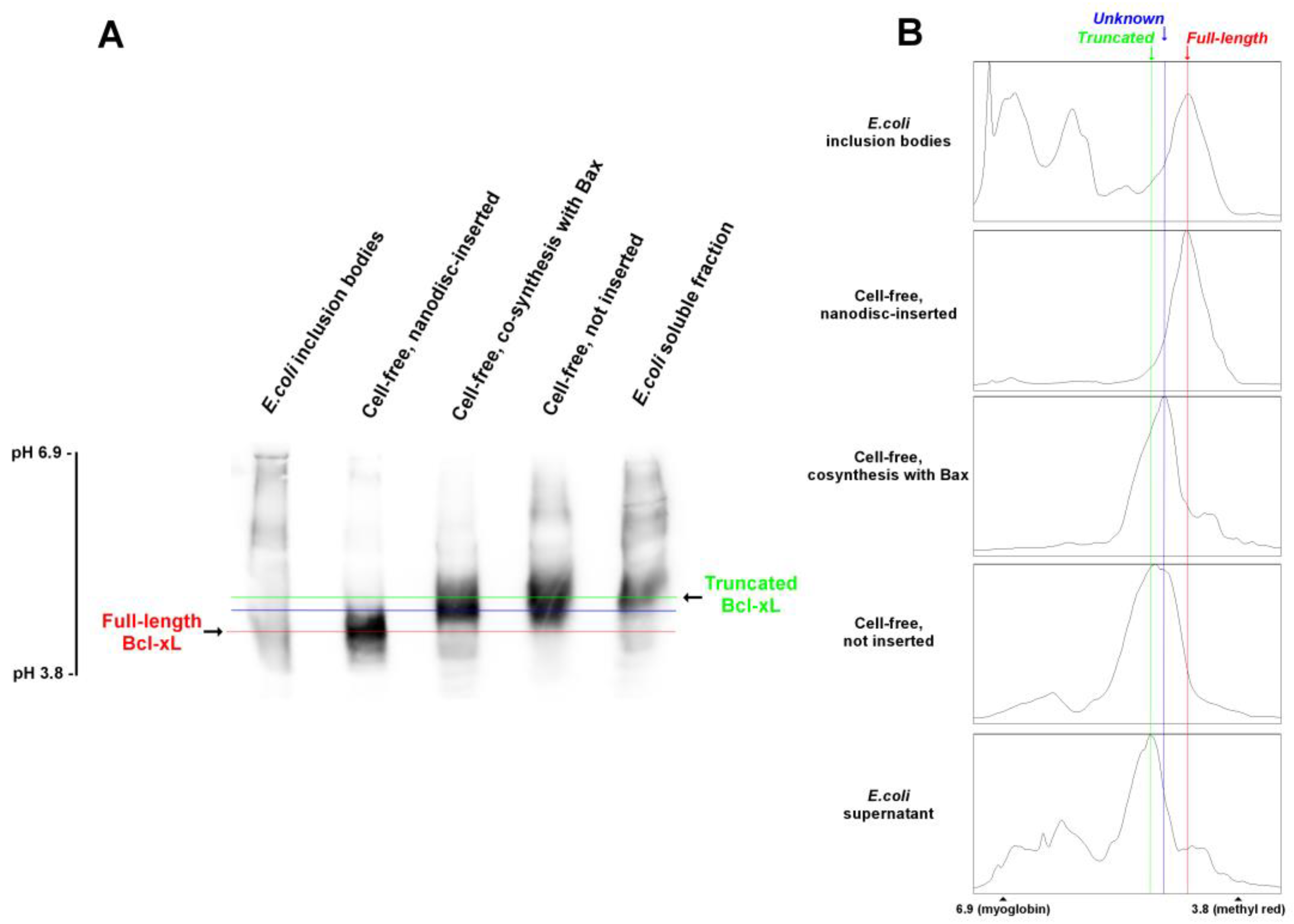
3.2. Nanodisc-Inserted Bcl-xL Is Full-Length
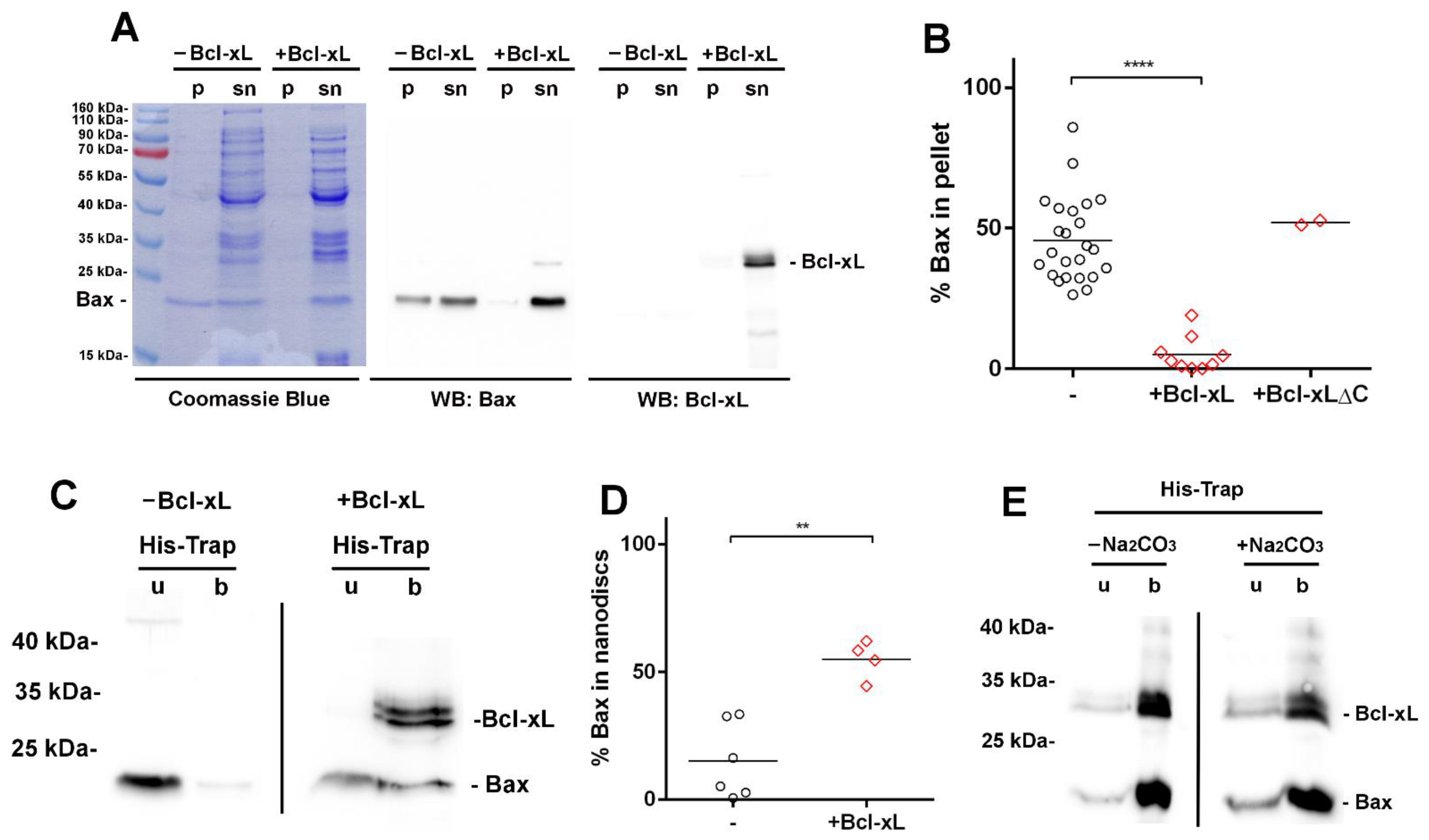
3.3. Bcl-xL Stimulates the Membrane Insertion of Bax into Nanodiscs
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Birkinshaw, R.W.; Czabotar, P.E. The BCL-2 family of proteins and mitochondrial outer membrane permeabilisation. Semin. Cell Dev. Biol. 2017, 72, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Popgeorgiev, N.; Jabbour, L.; Gillet, G. Subcellular Localization and Dynamics of the Bcl-2 Family of Proteins. Front. Cell Dev. Biol. 2018, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Green, D.R. The Mitochondrial Pathway of Apoptosis Part II: The BCL-2 Protein Family. Cold Spring Harb. Perspect. Biol. 2022, 14, a041046. [Google Scholar] [CrossRef]
- Petros, A.M.; Olejniczak, E.T.; Fesik, S.W. Structural biology of the Bcl-2 family of proteins. Biochim. Biophys. Acta 2004, 1644, 83–94. [Google Scholar] [CrossRef]
- Youle, R.J.; Strasser, A. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol. 2008, 9, 47–59. [Google Scholar] [CrossRef]
- Billen, L.P.; Shamas-Din, A.; Andrews, D.W. Bid: A Bax-like BH3 protein. Oncogene 2008, 27 (Suppl. S1), S93–S104. [Google Scholar] [CrossRef]
- Kvansakul, M.; Hinds, M.G. Structural biology of the Bcl-2 family and its mimicry by viral proteins. Cell Death Dis. 2013, 4, e909. [Google Scholar] [CrossRef]
- Antonsson, B.; Montessuit, S.; Lauper, S.; Eskes, R.; Martinou, J.C. Bax oligomerization is required for channel-forming activity in liposomes and to trigger cytochrome c release from mitochondria. Biochem. J. 2000, 345 Pt 2, 271–278. [Google Scholar] [CrossRef]
- Antonsson, B.; Montessuit, S.; Sanchez, B.; Martinou, J.C. Bax is present as a high molecular weight oligomer/complex in the mitochondrial membrane of apoptotic cells. J. Biol. Chem. 2001, 276, 11615–11623. [Google Scholar] [CrossRef]
- Annis, M.G.; Soucie, E.L.; Dlugosz, P.J.; Cruz-Aguado, J.A.; Penn, L.Z.; Leber, B.; Andrews, D.W. Bax forms multispanning monomers that oligomerize to permeabilize membranes during apoptosis. EMBO J. 2005, 24, 2096–2103. [Google Scholar] [CrossRef] [PubMed]
- Bleicken, S.; Classen, M.; Padmavathi, P.V.; Ishikawa, T.; Zeth, K.; Steinhoff, H.J.; Bordignon, E. Molecular details of Bax activation, oligomerization, and membrane insertion. J. Biol. Chem. 2010, 285, 6636–6647. [Google Scholar] [CrossRef] [PubMed]
- Pavlov, E.V.; Priault, M.; Pietkiewicz, D.; Cheng, E.H.; Antonsson, B.; Manon, S.; Korsmeyer, S.J.; Mannella, C.A.; Kinnally, K.W. A novel, high conductance channel of mitochondria linked to apoptosis in mammalian cells and Bax expression in yeast. J. Cell Biol. 2001, 155, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Dejean, L.M.; Martinez-Caballero, S.; Guo, L.; Hughes, C.; Teijido, O.; Ducret, T.; Ichas, F.; Korsmeyer, S.J.; Antonsson, B.; Jonas, E.A.; et al. Oligomeric Bax is a component of the putative cytochrome c release channel MAC, mitochondrial apoptosis-induced channel. Mol. Biol. Cell 2005, 16, 2424–2432. [Google Scholar] [CrossRef]
- Hsu, Y.T.; Youle, R.J. Nonionic detergents induce dimerization among members of the Bcl-2 family. J. Biol. Chem. 1997, 272, 13829–13834. [Google Scholar] [CrossRef]
- Peyerl, F.W.; Dai, S.; Murphy, G.A.; Crawford, F.; White, J.; Marrack, P.; Kappler, J.W. Elucidation of some Bax conformational changes through crystallization of an antibody-peptide complex. Cell Death Differ. 2007, 14, 447–452. [Google Scholar] [CrossRef]
- Czabotar, P.E.; Westphal, D.; Dewson, G.; Ma, S.; Hockings, C.; Fairlie, W.D.; Lee, E.F.; Yao, S.; Robin, A.Y.; Smith, B.J.; et al. Bax crystal structures reveal how BH3 domains activate Bax and nucleate its oligomerization to induce apoptosis. Cell 2013, 152, 519–531. [Google Scholar] [CrossRef]
- Bloch, N.B.; Wales, T.E.; Prew, M.S.; Levy, H.R.; Engen, J.R.; Walensky, L.D. The conformational stability of pro-apoptotic BAX is dictated by discrete residues of the protein core. Nat. Commun. 2021, 12, 4932. [Google Scholar] [CrossRef]
- Muchmore, S.W.; Sattler, M.; Liang, H.; Meadows, R.P.; Harlan, J.E.; Yoon, H.S.; Nettesheim, D.; Chang, B.S.; Thompson, C.B.; Wong, S.L.; et al. X-ray and NMR structure of human Bcl-xL, an inhibitor of programmed cell death. Nature 1996, 381, 335–341. [Google Scholar] [CrossRef]
- Rajan, S.; Choi, M.; Baek, K.; Yoon, H.S. Bh3 induced conformational changes in Bcl-Xl revealed by crystal structure and comparative analysis. Proteins 2015, 83, 1262–1272. [Google Scholar] [CrossRef]
- Yao, Y.; Nisan, D.; Fujimoto, L.M.; Antignani, A.; Barnes, A.; Tjandra, N.; Youle, R.J.; Marassi, F.M. Characterization of the membrane-inserted C-terminus of cytoprotective BCL-XL. Protein. Expr. Purif. 2016, 122, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Ryzhov, P.; Tian, Y.; Yao, Y.; Bobkov, A.A.; Im, W.; Marassi, F.M. Conformational States of the Cytoprotective Protein Bcl-xL. Biophys. J. 2020, 119, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Youle, R.J.; Tjandra, N. Structure of Bax: Coregulation of dimer formation and intracellular localization. Cell 2000, 103, 645–654. [Google Scholar] [CrossRef]
- Vasquez-Montes, V.; Vargas-Uribe, M.; Pandey, N.K.; Rodnin, M.V.; Langen, R.; Ladokhin, A.S. Lipid-modulation of membrane insertion and refolding of the apoptotic inhibitor Bcl-xL. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 691–700. [Google Scholar] [CrossRef] [PubMed]
- Todt, F.; Cakir, Z.; Reichenbach, F.; Youle, R.J.; Edlich, F. The C-terminal helix of Bcl-x(L) mediates Bax retrotranslocation from the mitochondria. Cell Death Differ. 2013, 20, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Renault, T.T.; Teijido, O.; Missire, F.; Ganesan, Y.T.; Velours, G.; Arokium, H.; Beaumatin, F.; Llanos, R.; Athané, A.; Camougrand, N.; et al. Bcl-xL stimulates Bax relocation to mitochondria and primes cells to ABT-737. Int. J. Biochem. Cell Biol. 2015, 64, 136–146. [Google Scholar] [CrossRef]
- Upreti, M.; Galitovskaya, E.N.; Chu, R.; Tackett, A.J.; Terrano, D.T.; Granell, S.; Chambers, T.C. Identification of the major phosphorylation site in Bcl-xL induced by microtubule inhibitors and analysis of its functional significance. J. Biol. Chem. 2008, 283, 35517–35525. [Google Scholar] [CrossRef]
- Baruah, P.S.; Beauchemin, M.; Hébert, J.; Bertrand, R. Dynamic Bcl-xL (S49) and (S62) Phosphorylation/Dephosphorylation during Mitosis Prevents Chromosome Instability and Aneuploidy in Normal Human Diploid Fibroblasts. PLoS ONE 2016, 11, e0159091. [Google Scholar] [CrossRef]
- Deverman, B.E.; Cook, B.L.; Manson, S.R.; Niederhoff, R.A.; Langer, E.M.; Rosová, I.; Kulans, L.A.; Fu, X.; Weinberg, J.S.; Heinecke, J.W.; et al. Bcl-xL deamidation is a critical switch in the regulation of the response to DNA damage. Cell 2002, 111, 51–62. [Google Scholar] [CrossRef]
- Beaumatin, F.; El Dhaybi, M.; Lasserre, J.P.; Salin, B.; Moyer, M.P.; Verdier, M.; Manon, S.; Priault, M. N52 monodeamidated Bcl-xL shows impaired oncogenic properties in vivo and in vitro. Oncotarget 2016, 7, 17129–17143. [Google Scholar] [CrossRef]
- Rues, R.B.; Gräwe, A.; Henrich, E.; Bernhard, F. Membrane Protein Production in E. coli Lysates in Presence of Preassembled Nanodiscs. Methods Mol. Biol. 2017, 1586, 291–312. [Google Scholar] [CrossRef] [PubMed]
- Köck, Z.; Ermel, U.; Martin, J.; Morgner, N.; Frangakis, A.S.; Dötsch, V.; Hilger, D.; Bernhard, F. Biochemical Characterization of Cell-free Synthesized Human β1 Adrenergic Receptor Cotranslationally Inserted into Nanodiscs. J. Mol. Biol. 2022, 434, 167687. [Google Scholar] [CrossRef] [PubMed]
- Levin, R.; Köck, Z.; Martin, J.; Zangl, R.; Gewering, T.; Schüler, L.; Moeller, A.; Dötsch, V.; Morgner, N.; Bernhard, F. Cotranslational assembly of membrane protein/nanoparticles in cell-free systems. Biochim. Biophys. Acta Biomembr. 2022, 1864, 184017. [Google Scholar] [CrossRef]
- Rouchidane Eyitayo, A.; Giraud, M.F.; Daury, L.; Lambert, O.; Gonzalez, C.; Manon, S. Cell-free synthesis and reconstitution of Bax in nanodiscs: Comparison between wild-type Bax and a constitutively active mutant. Biochim. Biophys. Acta Biomembr. 2023, 1865, 184075. [Google Scholar] [CrossRef]
- Simonyan, L.; Légiot, A.; Lascu, I.; Durand, G.; Giraud, M.F.; Gonzalez, C.; Manon, S. The substitution of Proline 168 favors Bax oligomerization and stimulates its interaction with LUVs and mitochondria. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1144–1155. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, D.; Junge, F.; Durst, F.; Frölich, N.; Schneider, B.; Reckel, S.; Sobhanifar, S.; Dötsch, V.; Bernhard, F. Preparative scale expression of membrane proteins in Escherichia coli-based continuous exchange cell-free systems. Nat. Protoc. 2007, 2, 2945–2957. [Google Scholar] [CrossRef]
- Boudier-Lemosquet, A.; Mahler, A.; Bobo, C.; Moreau, V.; Priault, M. 1D continuous gel electrophoresis composition for the separation of deamidated proteins. Methods 2022, 200, 23–30. [Google Scholar] [CrossRef]
- Bobo, C.; Céré, C.; Dufossée, M.; Dautant, A.; Moreau, V.; Manon, S.; Beaumatin, F.; Priault, M. Improved Electrophoretic Separation to Assist the Monitoring of Bcl-xL Post-Translational Modifications. Int. J. Mol. Sci. 2019, 20, 5571. [Google Scholar] [CrossRef]
- Kaufmann, T.; Schlipf, S.; Sanz, J.; Neubert, K.; Stein, R.; Borner, C. Characterization of the signal that directs Bcl-x(L), but not Bcl-2, to the mitochondrial outer membrane. J. Cell Biol. 2003, 160, 53–64. [Google Scholar] [CrossRef]
- Minn, A.J.; Vélez, P.; Schendel, S.L.; Liang, H.; Muchmore, S.W.; Fesik, S.W.; Fill, M.; Thompson, C.B. Bcl-x(L) forms an ion channel in synthetic lipid membranes. Nature 1997, 385, 353–357. [Google Scholar] [CrossRef]
- Bossy-Wetzel, E.; Green, D.R. Caspases induce cytochrome c release from mitochondria by activating cytosolic factors. J. Biol. Chem. 1999, 274, 17484–17490. [Google Scholar] [CrossRef] [PubMed]
- Degterev, A.; Lugovskoy, A.; Cardone, M.; Mulley, B.; Wagner, G.; Mitchison, T.; Yuan, J. Identification of small-molecule inhibitors of interaction between the BH3 domain and Bcl-xL. Nat. Cell Biol. 2001, 3, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Jonas, E.A.; Hickman, J.A.; Chachar, M.; Polster, B.M.; Brandt, T.A.; Fannjiang, Y.; Ivanovska, I.; Basañez, G.; Kinnally, K.W.; Zimmerberg, J.; et al. Proapoptotic N-truncated BCL-xL protein activates endogenous mitochondrial channels in living synaptic terminals. Proc. Natl. Acad. Sci. USA 2004, 101, 13590–13595. [Google Scholar] [CrossRef]
- Thai, K.; Choi, J.; Franzin, C.M.; Marassi, F.M. Bcl-XL as a fusion protein for the high-level expression of membrane-associated proteins. Protein Sci. 2005, 14, 948–955. [Google Scholar] [CrossRef]
- Chipuk, J.E.; Bouchier-Hayes, L.; Kuwana, T.; Newmeyer, D.D.; Green, D.R. PUMA couples the nuclear and cytoplasmic proapoptotic function of p53. Science 2005, 309, 1732–1735. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.W.; Manion, M.K.; Maguire, B.; Hockenbery, D.M. BCL-XL dimerization by three-dimensional domain swapping. J. Mol. Biol. 2006, 356, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Malia, T.J.; Wagner, G. NMR structural investigation of the mitochondrial outer membrane protein VDAC and its interaction with antiapoptotic Bcl-xL. Biochemistry 2007, 46, 514–525. [Google Scholar] [CrossRef]
- Gautier, F.; Guillemin, Y.; Cartron, P.F.; Gallenne, T.; Cauquil, N.; Le Diguarher, T.; Casara, P.; Vallette, F.M.; Manon, S.; Hickman, J.A.; et al. Bax activation by engagement with, then release from, the BH3 binding site of Bcl-xL. Mol. Cell Biol. 2011, 31, 832–844. [Google Scholar] [CrossRef]
- Tan, Y.J.; Beerheide, W.; Ting, A.E. Biophysical characterization of the oligomeric state of Bax and its complex formation with Bcl-XL. Biochem. Biophys. Res. Commun. 1999, 255, 334–339. [Google Scholar] [CrossRef]
- Vander Heiden, M.G.; Li, X.X.; Gottleib, E.; Hill, R.B.; Thompson, C.B.; Colombini, M. Bcl-xL promotes the open configuration of the voltage-dependent anion channel and metabolite passage through the outer mitochondrial membrane. J. Biol. Chem. 2001, 276, 19414–19419. [Google Scholar] [CrossRef]
- Bleicken, S.; Hofhaus, G.; Ugarte-Uribe, B.; Schröder, R.; García-Sáez, A.J. cBid, Bax and Bcl-xL exhibit opposite membrane remodeling activities. Cell Death Dis. 2016, 7, e2121. [Google Scholar] [CrossRef]
- Denisov, I.G.; Grinkova, Y.V.; Lazarides, A.A.; Sligar, S.G. Directed self-assembly of monodisperse phospholipid bilayer Nanodiscs with controlled size. J. Am. Chem. Soc. 2004, 126, 3477–3487. [Google Scholar] [CrossRef] [PubMed]
- Denisov, I.G.; Sligar, S.G. Nanodiscs for structural and functional studies of membrane proteins. Nat. Struct. Mol. Biol. 2016, 23, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Sligar, S.G.; Denisov, I.G. Nanodiscs: A toolkit for membrane protein science. Protein Sci. 2021, 30, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Fujimoto, L.M.; Hirshman, N.; Bobkov, A.A.; Antignani, A.; Youle, R.J.; Marassi, F.M. Conformation of BCL-XL upon Membrane Integration. J. Mol. Biol. 2015, 427, 2262–2270. [Google Scholar] [CrossRef]
- Follis, A.V.; Llambi, F.; Kalkavan, H.; Yao, Y.; Phillips, A.H.; Park, C.G.; Marassi, F.M.; Green, D.R.; Kriwacki, R.W. Regulation of apoptosis by an intrinsically disordered region of Bcl-xL. Nat. Chem. Biol. 2018, 14, 458–465. [Google Scholar] [CrossRef]
- Yethon, J.A.; Epand, R.F.; Leber, B.; Epand, R.M.; Andrews, D.W. Interaction with a membrane surface triggers a reversible conformational change in Bax normally associated with induction of apoptosis. J. Biol. Chem. 2003, 278, 48935–48941. [Google Scholar] [CrossRef]
- Billen, L.P.; Kokoski, C.L.; Lovell, J.F.; Leber, B.; Andrews, D.W. Bcl-XL inhibits membrane permeabilization by competing with Bax. PLoS Biol. 2008, 6, e147. [Google Scholar] [CrossRef]
- Xu, X.P.; Zhai, D.; Kim, E.; Swift, M.; Reed, J.C.; Volkmann, N.; Hanein, D. Three-dimensional structure of Bax-mediated pores in membrane bilayers. Cell Death Dis. 2013, 4, e683. [Google Scholar] [CrossRef]
- López, C.A.; Swift, M.F.; Xu, X.P.; Hanein, D.; Volkmann, N.; Gnanakaran, S. Biophysical Characterization of a Nanodisc with and without BAX: An Integrative Study Using Molecular Dynamics Simulations and Cryo-EM. Structure 2019, 27, 988–999.e4. [Google Scholar] [CrossRef]
- Sedlak, T.W.; Oltvai, Z.N.; Yang, E.; Wang, K.; Boise, L.H.; Thompson, C.B.; Korsmeyer, S.J. Multiple Bcl-2 family members demonstrate selective dimerizations with Bax. Proc. Natl. Acad. Sci. USA 1995, 92, 7834–7838. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, J.P.; Smetsers, T.F.; Slöetjes, A.W.; Linders, E.H.; Mensink, E.J. Bax mutations in cell lines derived from hematological malignancies. Leukemia 1995, 9, 1828–1832. [Google Scholar] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rouchidane Eyitayo, A.; Boudier-Lemosquet, A.; Chaignepain, S.; Priault, M.; Manon, S. Bcl-xL Is Spontaneously Inserted into Preassembled Nanodiscs and Stimulates Bax Insertion in a Cell-Free Protein Synthesis System. Biomolecules 2023, 13, 876. https://doi.org/10.3390/biom13060876
Rouchidane Eyitayo A, Boudier-Lemosquet A, Chaignepain S, Priault M, Manon S. Bcl-xL Is Spontaneously Inserted into Preassembled Nanodiscs and Stimulates Bax Insertion in a Cell-Free Protein Synthesis System. Biomolecules. 2023; 13(6):876. https://doi.org/10.3390/biom13060876
Chicago/Turabian StyleRouchidane Eyitayo, Akandé, Axel Boudier-Lemosquet, Stéphane Chaignepain, Muriel Priault, and Stéphen Manon. 2023. "Bcl-xL Is Spontaneously Inserted into Preassembled Nanodiscs and Stimulates Bax Insertion in a Cell-Free Protein Synthesis System" Biomolecules 13, no. 6: 876. https://doi.org/10.3390/biom13060876