
Flexible Attachment and Detachment of Centromeres and Telomeres to and from Chromosomes
Abstract
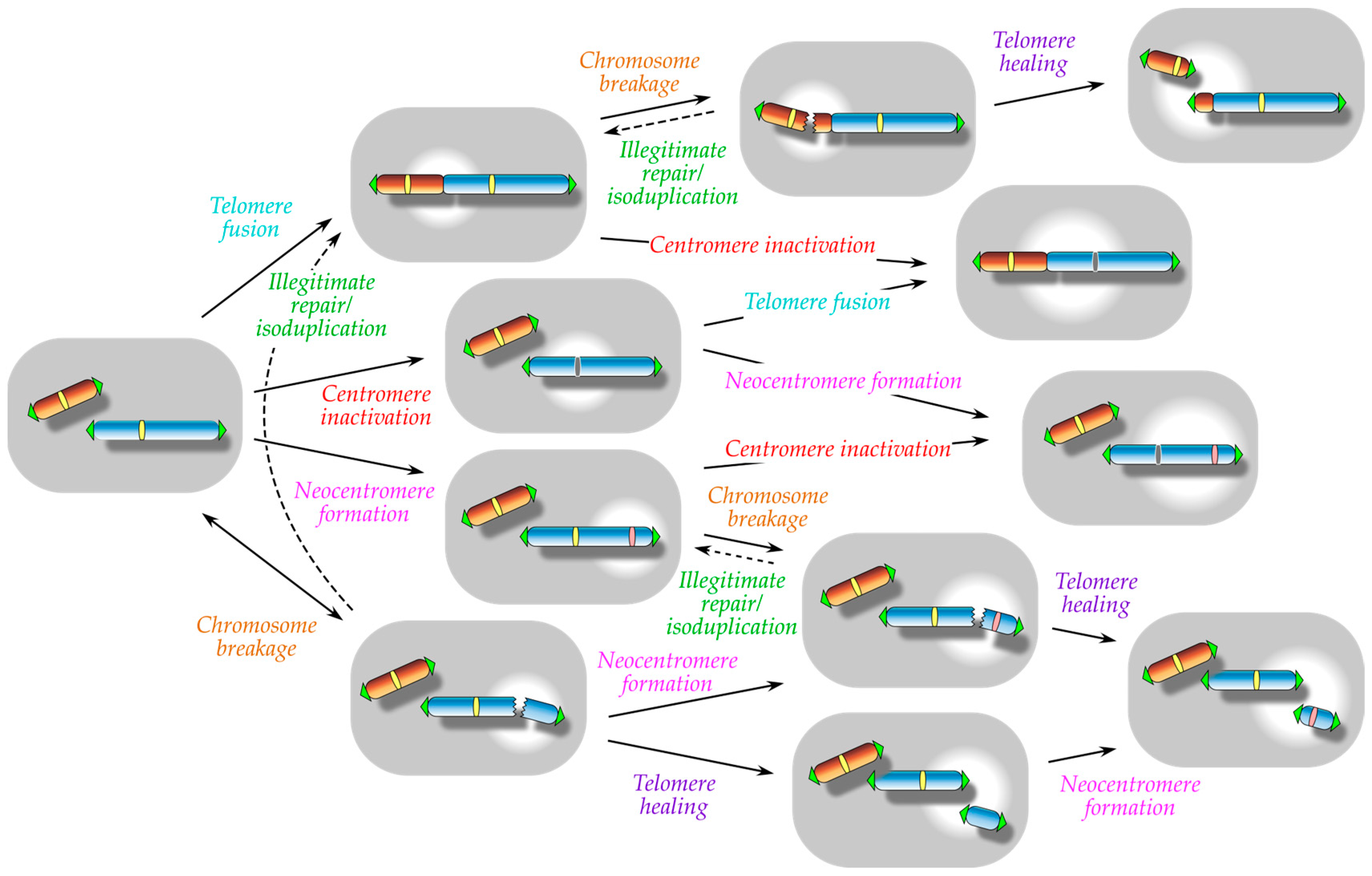
:1. Introduction
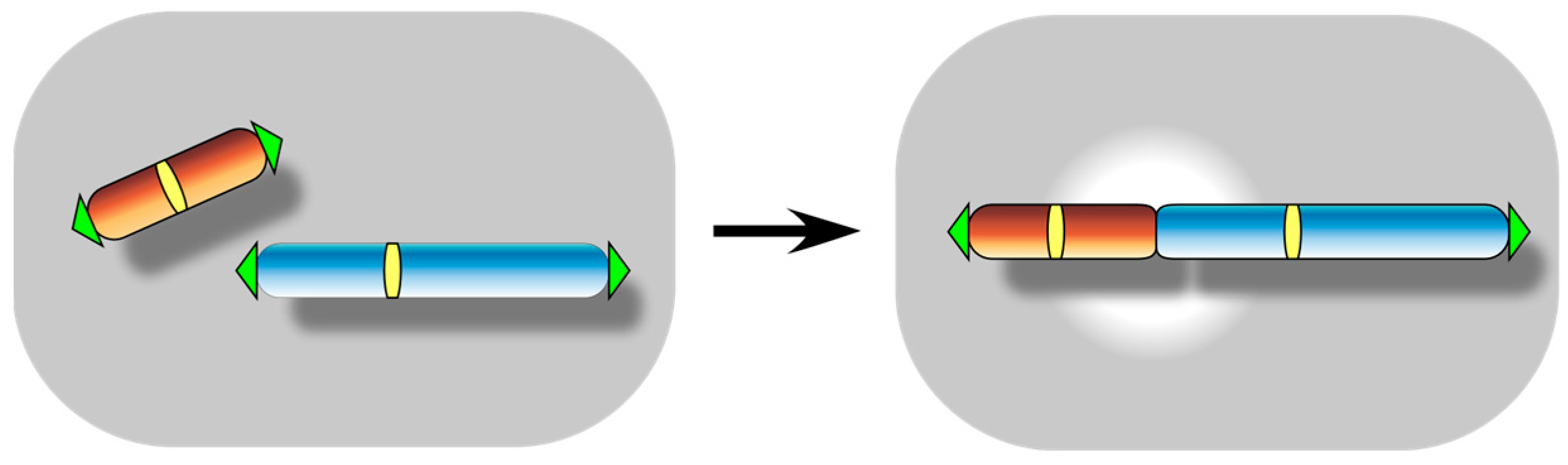
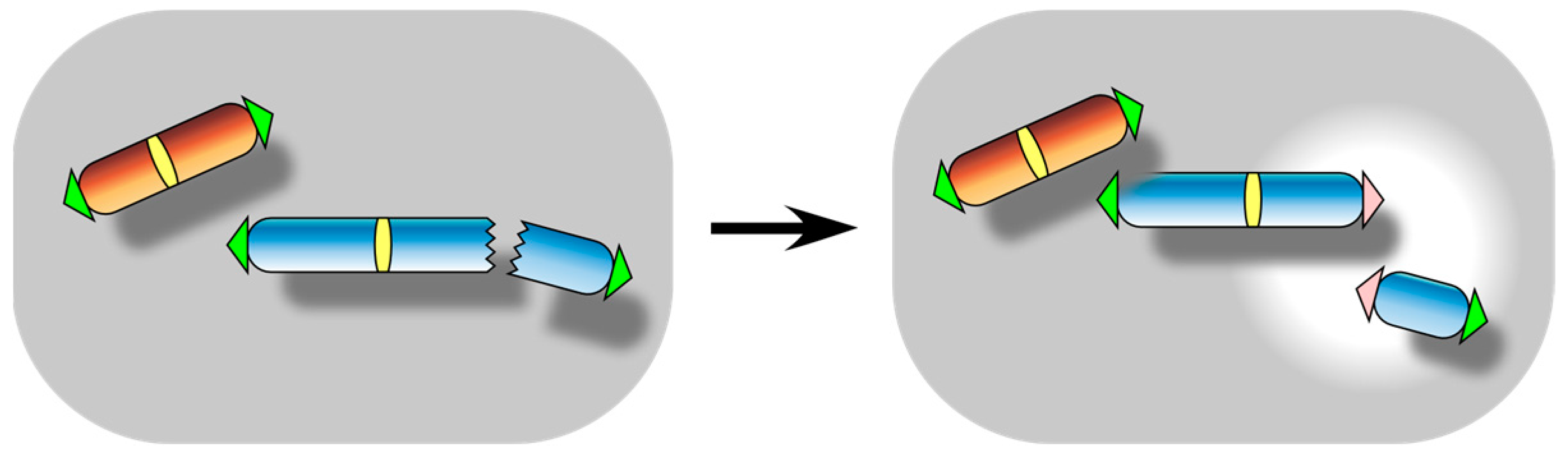
2. Spontaneous Telomere Fusion: A Manifestation of Functional Telomere Detachment
2.1. Estimation of Spontaneous Telomere Fusion Frequency in Mammals
2.2. Spontaneous Telomere Fusion in Fission Yeast (Schizosaccharomyces pombe)
2.3. Spontaneous Telomere Fusion in Budding Yeast (Saccharomyces cerevisiae)
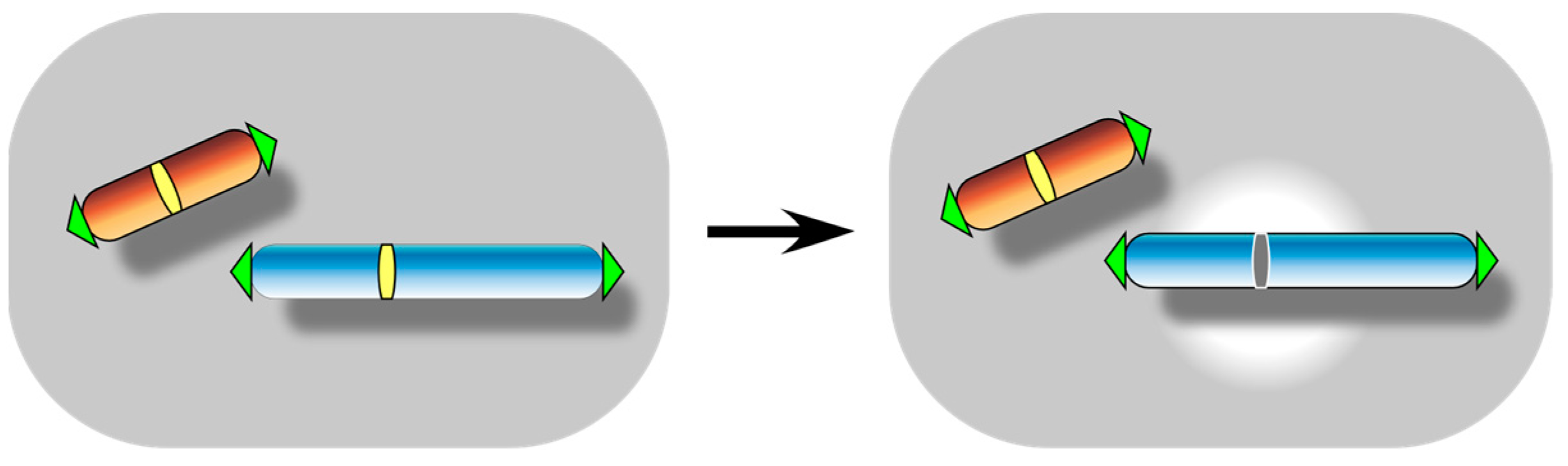
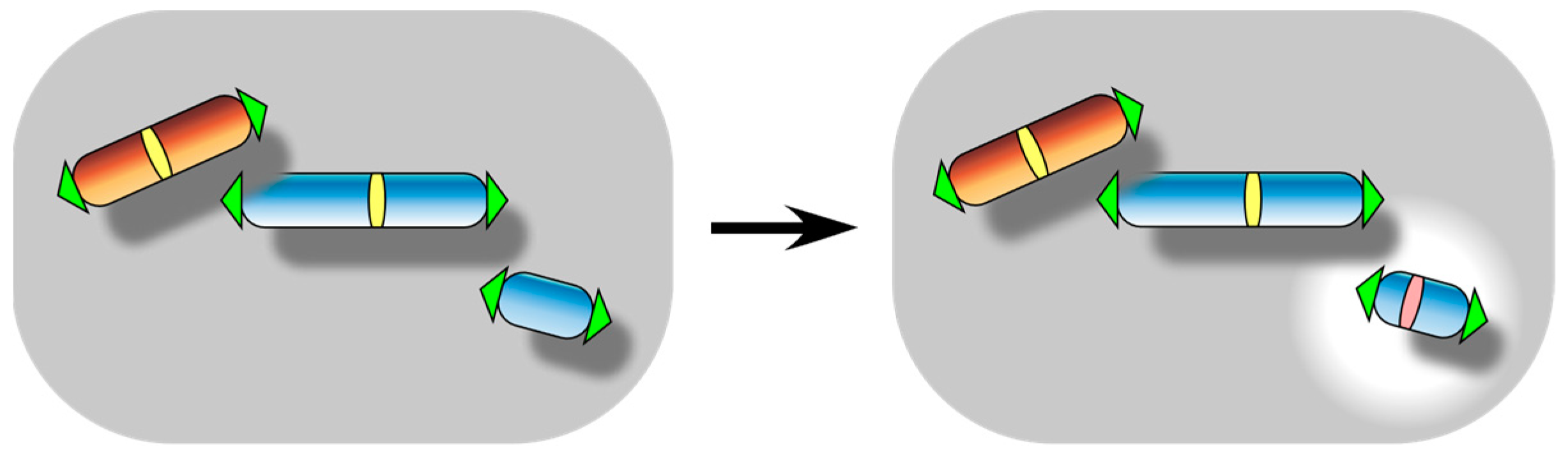
3. Centromere Inactivation: A Manifestation of Functional Centromere Detachment
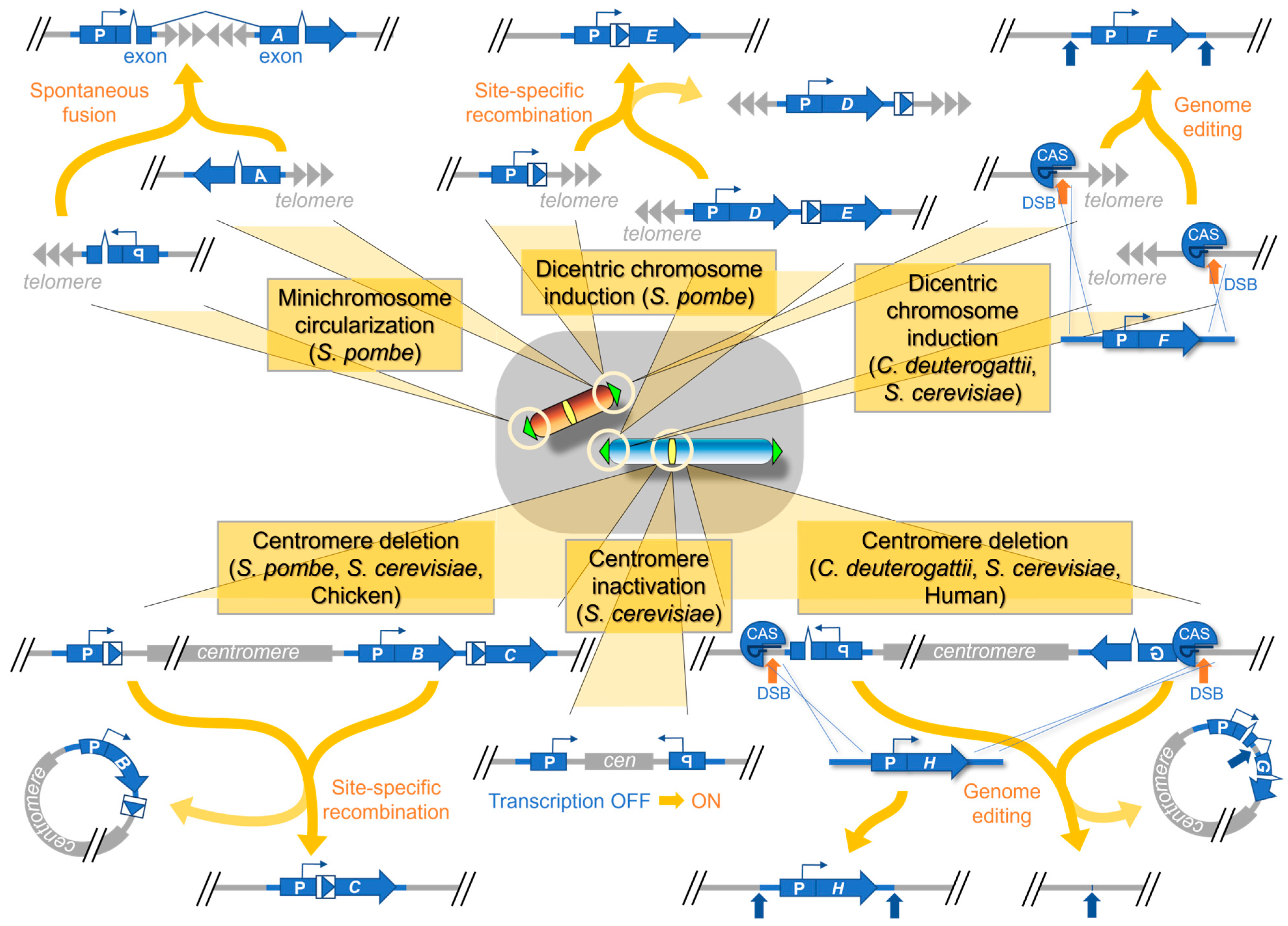
3.1. Centromere Inactivation in the Induced Human Dicentric Chromosomes
3.2. Centromere Inactivation in the Induced Dicentric Chromosomes in S. pombe
3.3. Centromere Inactivation in the Induced Dicentric Chromosomes in Various Yeast Species
4. Telomere Healing: A Manifestation of Functional Telomere Attachment
4.1. Telomere Healing at Site-Specific DSBs in S. cerevisiae
4.2. Telomere Healing during GCRs in S. cerevisiae
4.3. Evolutionary Conservation of Telomere Healing during GCRs
4.4. Site-Specific DSBs, GCRs, and Telomere Healing in Centromeric Minichromosomes in S. pombe
5. Neocentromere Formation: A Manifestation of Functional Centromere Attachment
5.1. Efficiency of Neocentromere Formation in Addition to Telomere Fusion in Various Yeast Species
5.2. Efficiency of Neocentromere Formation in Addition to Telomere Fusion in Vertebrate Cells
5.3. Site Preference for Neocentromere Formation
5.4. Analysis of Ongoing Chromosomal Rearrangements and Maturation
6. Overall Discussion and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, A.W.; Szostak, J.W. Construction of artificial chromosomes in yeast. Nature 1983, 305, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, S.; Fukagawa, T. Kinetochore Architecture Employs Diverse Linker Strategies Across Evolution. Front. Cell Dev. Biol. 2022, 10, 862637. [Google Scholar] [CrossRef] [PubMed]
- Musacchio, A.; Desai, A. A Molecular View of Kinetochore Assembly and Function. Biology 2017, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Pfeiffer, V.; Lingner, J. Replication of telomeres and the regulation of telomerase. Cold Spring Harb. Perspect. Biol. 2013, 5, a010405. [Google Scholar] [CrossRef] [PubMed]
- Nandakumar, J.; Cech, T.R. Finding the end: Recruitment of telomerase to telomeres. Nat. Rev. Mol. Cell Biol. 2013, 14, 69–82. [Google Scholar] [CrossRef]
- Mechali, M. Eukaryotic DNA replication origins: Many choices for appropriate answers. Nat. Rev. Mol. Cell Biol. 2010, 11, 728–738. [Google Scholar] [CrossRef]
- Parker, M.W.; Botchan, M.R.; Berger, J.M. Mechanisms and regulation of DNA replication initiation in eukaryotes. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 107–144. [Google Scholar] [CrossRef] [Green Version]
- Sundararajan, K.; Straight, A.F. Centromere Identity and the Regulation of Chromosome Segregation. Front. Cell Dev. Biol. 2022, 10, 914249. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Almouzni, G. Chromatin dynamics during the cell cycle at centromeres. Nat. Rev. Genet. 2017, 18, 192–208. [Google Scholar] [CrossRef]
- Barra, V.; Fachinetti, D. The dark side of centromeres: Types, causes and consequences of structural abnormalities implicating centromeric DNA. Nat. Commun. 2018, 9, 4340. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, L.L.; Sullivan, B.A. Genomic and functional variation of human centromeres. Exp. Cell Res. 2020, 389, 111896. [Google Scholar] [CrossRef]
- Doksani, Y.; de Lange, T. The role of double-strand break repair pathways at functional and dysfunctional telomeres. Cold Spring Harb. Perspect. Biol. 2014, 6, a016576. [Google Scholar] [CrossRef]
- Cleal, K.; Baird, D.M. Catastrophic Endgames: Emerging Mechanisms of Telomere-Driven Genomic Instability. Trends Genet. 2020, 36, 347–359. [Google Scholar] [CrossRef]
- Webb, C.J.; Wu, Y.; Zakian, V.A. DNA repair at telomeres: Keeping the ends intact. Cold Spring Harb. Perspect. Biol. 2013, 5, a012666. [Google Scholar] [CrossRef] [PubMed]
- Marcomini, I.; Gasser, S.M. Nuclear organization in DNA end processing: Telomeres vs double-strand breaks. DNA Repair. (Amst.) 2015, 32, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Tuduri, S.; Tourriere, H.; Pasero, P. Defining replication origin efficiency using DNA fiber assays. Chromosome Res. 2010, 18, 91–102. [Google Scholar] [CrossRef]
- Marchal, C.; Sima, J.; Gilbert, D.M. Control of DNA replication timing in the 3D genome. Nat. Rev. Mol. Cell Biol. 2019, 20, 721–737. [Google Scholar] [CrossRef] [PubMed]
- Putnam, C.D.; Kolodner, R.D. Pathways and Mechanisms that Prevent Genome Instability in Saccharomyces cerevisiae. Genetics 2017, 206, 1187–1225. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, R.; Hu, Q.; Ly, P. Mechanistic origins of diverse genome rearrangements in cancer. Semin. Cell Dev. Biol. 2022, 123, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Bolzan, A.D. Chromosomal aberrations involving telomeres and interstitial telomeric sequences. Mutagenesis 2012, 27, 1–15. [Google Scholar] [CrossRef] [Green Version]
- McClintock, B. The Stability of Broken Ends of Chromosomes in Zea Mays. Genetics 1941, 26, 234–282. [Google Scholar] [CrossRef]
- Stanyon, R.; Rocchi, M.; Capozzi, O.; Roberto, R.; Misceo, D.; Ventura, M.; Cardone, M.F.; Bigoni, F.; Archidiacono, N. Primate chromosome evolution: Ancestral karyotypes, marker order and neocentromeres. Chromosome Res. 2008, 16, 17–39. [Google Scholar] [CrossRef] [PubMed]
- Bracewell, R.; Chatla, K.; Nalley, M.J.; Bachtrog, D. Dynamic turnover of centromeres drives karyotype evolution in Drosophila. Elife 2019, 8, e49002. [Google Scholar] [CrossRef]
- Sankaranarayanan, S.R.; Ianiri, G.; Coelho, M.A.; Reza, M.H.; Thimmappa, B.C.; Ganguly, P.; Vadnala, R.N.; Sun, S.; Siddharthan, R.; Tellgren-Roth, C.; et al. Loss of centromere function drives karyotype evolution in closely related Malassezia species. Elife 2020, 9, e53944. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Xu, Z.; Bai, H.; Huang, Y.; Kang, N.; Ding, X.; Liu, J.; Luo, H.; Yang, C.; Chen, W.; et al. Evolutionary analysis of a complete chicken genome. Proc. Natl. Acad. Sci. USA 2023, 120, e2216641120. [Google Scholar] [CrossRef] [PubMed]
- Murnane, J.P. Telomere dysfunction and chromosome instability. Mutat. Res. 2012, 730, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Casari, E.; Gnugnoli, M.; Rinaldi, C.; Pizzul, P.; Colombo, C.V.; Bonetti, D.; Longhese, M.P. To Fix or Not to Fix: Maintenance of Chromosome Ends Versus Repair of DNA Double-Strand Breaks. Cells 2022, 11, 3224. [Google Scholar] [CrossRef]
- Mackinnon, R.N.; Campbell, L.J. The role of dicentric chromosome formation and secondary centromere deletion in the evolution of myeloid malignancy. Genet. Res. Int. 2011, 2011, 643628. [Google Scholar] [CrossRef] [Green Version]
- Stimpson, K.M.; Matheny, J.E.; Sullivan, B.A. Dicentric chromosomes: Unique models to study centromere function and inactivation. Chromosome Res. 2012, 20, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Saitoh, S. Switching the centromeres on and off: Epigenetic chromatin alterations provide plasticity in centromere activity stabilizing aberrant dicentric chromosomes. Biochem. Soc. Trans. 2013, 41, 1648–1653. [Google Scholar] [CrossRef] [Green Version]
- Ribeyre, C.; Shore, D. Regulation of telomere addition at DNA double-strand breaks. Chromosoma 2013, 122, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Hoerr, R.E.; Ngo, K.; Friedman, K.L. When the Ends Justify the Means: Regulation of Telomere Addition at Double-Strand Breaks in Yeast. Front. Cell Dev. Biol. 2021, 9, 655377. [Google Scholar] [CrossRef]
- Hori, T.; Fukagawa, T. Artificial generation of centromeres and kinetochores to understand their structure and function. Exp. Cell Res. 2020, 389, 111898. [Google Scholar] [CrossRef] [PubMed]
- Murillo-Pineda, M.; Jansen, L.E.T. Genetics, epigenetics and back again: Lessons learned from neocentromeres. Exp. Cell Res. 2020, 389, 111909. [Google Scholar] [CrossRef]
- DeBose-Scarlett, E.M.; Sullivan, B.A. Genomic and Epigenetic Foundations of Neocentromere Formation. Annu. Rev. Genet. 2021, 55, 331–348. [Google Scholar] [CrossRef] [PubMed]
- van Steensel, B.; Smogorzewska, A.; de Lange, T. TRF2 protects human telomeres from end-to-end fusions. Cell 1998, 92, 401–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, J.A.; Feldser, D.M.; Greider, C.W. Telomere dysfunction increases mutation rate and genomic instability. Cell 2001, 106, 275–286. [Google Scholar] [CrossRef] [Green Version]
- Mieczkowski, P.A.; Mieczkowska, J.O.; Dominska, M.; Petes, T.D. Genetic regulation of telomere-telomere fusions in the yeast Saccharomyces cerevisae. Proc. Natl. Acad. Sci. USA 2003, 100, 10854–10859. [Google Scholar] [CrossRef] [Green Version]
- Pennaneach, V.; Kolodner, R.D. Recombination and the Tel1 and Mec1 checkpoints differentially effect genome rearrangements driven by telomere dysfunction in yeast. Nat. Genet. 2004, 36, 612–617. [Google Scholar] [CrossRef]
- Naito, T.; Matsuura, A.; Ishikawa, F. Circular chromosome formation in a fission yeast mutant defective in two ATM homologues. Nat. Genet. 1998, 20, 203–206. [Google Scholar] [CrossRef]
- Nakamura, T.M.; Cooper, J.P.; Cech, T.R. Two modes of survival of fission yeast without telomerase. Science 1998, 282, 493–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, P.; Cech, T.R. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 2001, 292, 1171–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, M.G.; Cooper, J.P. The fission yeast Taz1 protein protects chromosomes from Ku-dependent end-to-end fusions. Mol. Cell 2001, 7, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.; McEachern, M.J.; Dandjinou, A.T.; Tzfati, Y.; Orr, E.; Blackburn, E.H.; Lue, N.F. Telomerase core components protect Candida telomeres from aberrant overhang accumulation. Proc. Natl. Acad. Sci. USA 2007, 104, 11682–11687. [Google Scholar] [CrossRef] [Green Version]
- Capper, R.; Britt-Compton, B.; Tankimanova, M.; Rowson, J.; Letsolo, B.; Man, S.; Haughton, M.; Baird, D.M. The nature of telomere fusion and a definition of the critical telomere length in human cells. Genes. Dev. 2007, 21, 2495–2508. [Google Scholar] [CrossRef] [Green Version]
- Letsolo, B.T.; Rowson, J.; Baird, D.M. Fusion of short telomeres in human cells is characterized by extensive deletion and microhomology, and can result in complex rearrangements. Nucleic Acids Res. 2010, 38, 1841–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hemann, M.T.; Strong, M.A.; Hao, L.Y.; Greider, C.W. The shortest telomere, not average telomere length, is critical for cell viability and chromosome stability. Cell 2001, 107, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.R.; Taylor, M.R.; Boulton, S.J. Playing the end game: DNA double-strand break repair pathway choice. Mol. Cell 2012, 47, 497–510. [Google Scholar] [CrossRef] [Green Version]
- Scully, R.; Panday, A.; Elango, R.; Willis, N.A. DNA double-strand break repair-pathway choice in somatic mammalian cells. Nat. Rev. Mol. Cell Biol. 2019, 20, 698–714. [Google Scholar] [CrossRef]
- Cleal, K.; Jones, R.E.; Grimstead, J.W.; Hendrickson, E.A.; Baird, D.M. Chromothripsis during telomere crisis is independent of NHEJ, and consistent with a replicative origin. Genome Res. 2019, 29, 737–749. [Google Scholar] [CrossRef] [Green Version]
- Martinez, A.C.; van Wely, K.H. Centromere fission, not telomere erosion, triggers chromosomal instability in human carcinomas. Carcinogenesis 2011, 32, 796–803. [Google Scholar] [CrossRef] [Green Version]
- Dewhurst, S.M.; Yao, X.; Rosiene, J.; Tian, H.; Behr, J.; Bosco, N.; Takai, K.K.; de Lange, T.; Imielinski, M. Structural variant evolution after telomere crisis. Nat. Commun. 2021, 12, 2093. [Google Scholar] [CrossRef]
- Stimpson, K.M.; Song, I.Y.; Jauch, A.; Holtgreve-Grez, H.; Hayden, K.E.; Bridger, J.M.; Sullivan, B.A. Telomere disruption results in non-random formation of de novo dicentric chromosomes involving acrocentric human chromosomes. PLoS Genet. 2010, 6. [Google Scholar] [CrossRef] [Green Version]
- Fouladi, B.; Sabatier, L.; Miller, D.; Pottier, G.; Murnane, J.P. The relationship between spontaneous telomere loss and chromosome instability in a human tumor cell line. Neoplasia 2000, 2, 540–554. [Google Scholar] [CrossRef] [Green Version]
- Murillo-Pineda, M.; Valente, L.P.; Dumont, M.; Mata, J.F.; Fachinetti, D.; Jansen, L.E.T. Induction of spontaneous human neocentromere formation and long-term maturation. J. Cell Biol. 2021, 220, e202007210. [Google Scholar] [CrossRef] [PubMed]
- Almeida, H.; Godinho Ferreira, M. Spontaneous telomere to telomere fusions occur in unperturbed fission yeast cells. Nucleic Acids Res. 2013, 41, 3056–3067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, H.; Masuda, F.; Takayama, Y.; Takahashi, K.; Saitoh, S. Epigenetic inactivation and subsequent heterochromatinization of a centromere stabilize dicentric chromosomes. Curr. Biol. 2012, 22, 658–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogiyama, Y.; Ohno, Y.; Kubota, Y.; Ishii, K. Epigenetically induced paucity of histone H2A.Z stabilizes fission-yeast ectopic centromeres. Nat. Struct. Mol. Biol. 2013, 20, 1397–1406. [Google Scholar] [CrossRef]
- Irie, H.; Yamamoto, I.; Tarumoto, Y.; Tashiro, S.; Runge, K.W.; Ishikawa, F. Telomere-binding proteins Taz1 and Rap1 regulate DSB repair and suppress gross chromosomal rearrangements in fission yeast. PLoS Genet. 2019, 15, e1008335. [Google Scholar] [CrossRef] [Green Version]
- Mangahas, J.L.; Alexander, M.K.; Sandell, L.L.; Zakian, V.A. Repair of chromosome ends after telomere loss in Saccharomyces. Mol. Biol. Cell 2001, 12, 4078–4089. [Google Scholar] [CrossRef] [Green Version]
- Pobiega, S.; Alibert, O.; Marcand, S. A new assay capturing chromosome fusions shows a protection trade-off at telomeres and NHEJ vulnerability to low-density ionizing radiation. Nucleic Acids Res. 2021, 49, 6817–6831. [Google Scholar] [CrossRef]
- Surosky, R.T.; Tye, B.K. Resolution of dicentric chromosomes by Ty-mediated recombination in yeast. Genetics 1985, 110, 397–419. [Google Scholar] [CrossRef]
- Jager, D.; Philippsen, P. Stabilization of dicentric chromosomes in Saccharomyces cerevisiae by telomere addition to broken ends or by centromere deletion. EMBO J. 1989, 8, 247–254. [Google Scholar] [CrossRef]
- Diede, S.J.; Gottschling, D.E. Telomerase-mediated telomere addition in vivo requires DNA primase and DNA polymerases alpha and delta. Cell 1999, 99, 723–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Kolodner, R.D. Gross chromosomal rearrangements in Saccharomyces cerevisiae replication and recombination defective mutants. Nat. Genet. 1999, 23, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Myung, K.; Datta, A.; Kolodner, R.D. Suppression of spontaneous chromosomal rearrangements by S phase checkpoint functions in Saccharomyces cerevisiae. Cell 2001, 104, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotanus, K.; Heitman, J. Centromere deletion in Cryptococcus deuterogattii leads to neocentromere formation and chromosome fusions. Elife 2020, 9, e56026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotanus, K.; Yadav, V.; Heitman, J. Epigenetic dynamics of centromeres and neocentromeres in Cryptococcus deuterogattii. PLoS Genet. 2021, 17, e1009743. [Google Scholar] [CrossRef] [PubMed]
- Ketel, C.; Wang, H.S.; McClellan, M.; Bouchonville, K.; Selmecki, A.; Lahav, T.; Gerami-Nejad, M.; Berman, J. Neocentromeres form efficiently at multiple possible loci in Candida albicans. PLoS Genet. 2009, 5, e1000400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, J.; Sanyal, K. Efficient neocentromere formation is suppressed by gene conversion to maintain centromere function at native physical chromosomal loci in Candida albicans. Genome Res. 2013, 23, 638–652. [Google Scholar] [CrossRef] [Green Version]
- Shang, W.H.; Hori, T.; Martins, N.M.; Toyoda, A.; Misu, S.; Monma, N.; Hiratani, I.; Maeshima, K.; Ikeo, K.; Fujiyama, A.; et al. Chromosome engineering allows the efficient isolation of vertebrate neocentromeres. Dev. Cell 2013, 24, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Gisselsson, D. Mitotic instability in cancer: Is there method in the madness? Cell Cycle 2005, 4, 1007–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisselsson, D.; Hoglund, M. Connecting mitotic instability and chromosome aberrations in cancer--can telomeres bridge the gap? Semin. Cancer Biol. 2005, 15, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Nimmo, E.R.; Cranston, G.; Allshire, R.C. Telomere-associated chromosome breakage in fission yeast results in variegated expression of adjacent genes. EMBO J. 1994, 13, 3801–3811. [Google Scholar] [CrossRef] [PubMed]
- Smogorzewska, A.; Karlseder, J.; Holtgreve-Grez, H.; Jauch, A.; de Lange, T. DNA ligase IV-dependent NHEJ of deprotected mammalian telomeres in G1 and G2. Curr. Biol. 2002, 12, 1635–1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oizumi, Y.; Kaji, T.; Tashiro, S.; Takeshita, Y.; Date, Y.; Kanoh, J. Complete sequences of Schizosaccharomyces pombe subtelomeres reveal multiple patterns of genome variation. Nat. Commun. 2021, 12, 611. [Google Scholar] [CrossRef]
- Ishii, K.; Ogiyama, Y.; Chikashige, Y.; Soejima, S.; Masuda, F.; Kakuma, T.; Hiraoka, Y.; Takahashi, K. Heterochromatin integrity affects chromosome reorganization after centromere dysfunction. Science 2008, 321, 1088–1091. [Google Scholar] [CrossRef]
- Ohno, Y.; Ogiyama, Y.; Kubota, Y.; Kubo, T.; Ishii, K. Acentric chromosome ends are prone to fusion with functional chromosome ends through a homology-directed rearrangement. Nucleic Acids Res. 2016, 44, 232–244. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Baumann, P. Chromosome fusions following telomere loss are mediated by single-strand annealing. Mol. Cell 2008, 31, 463–473. [Google Scholar] [CrossRef]
- Torres, E.M.; Sokolsky, T.; Tucker, C.M.; Chan, L.Y.; Boselli, M.; Dunham, M.J.; Amon, A. Effects of aneuploidy on cellular physiology and cell division in haploid yeast. Science 2007, 317, 916–924. [Google Scholar] [CrossRef]
- Pobiega, S.; Marcand, S. Dicentric breakage at telomere fusions. Genes. Dev. 2010, 24, 720–733. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.; Bloom, K. Genetic manipulation of centromere function. Mol. Cell Biol. 1987, 7, 2397–2405. [Google Scholar] [CrossRef]
- Lopez, V.; Barinova, N.; Onishi, M.; Pobiega, S.; Pringle, J.R.; Dubrana, K.; Marcand, S. Cytokinesis breaks dicentric chromosomes preferentially at pericentromeric regions and telomere fusions. Genes. Dev. 2015, 29, 322–336. [Google Scholar] [CrossRef] [Green Version]
- Stimpson, K.M.; Sullivan, L.L.; Kuo, M.E.; Sullivan, B.A. Nucleolar organization, ribosomal DNA array stability, and acrocentric chromosome integrity are linked to telomere function. PLoS ONE 2014, 9, e92432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maraschio, P.; Zuffardi, O.; Caiulo, A.; Dainotti, E.; Piantanida, M.; Rivera, H.; Tupler, R. Deletion of specific sequences or modification of centromeric chromatin are responsible for Y chromosome centromere inactivation. Hum. Genet. 1990, 85, 491–494. [Google Scholar] [CrossRef] [PubMed]
- Fisher, A.M.; Al-Gazali, L.; Pramathan, T.; Quaife, R.; Cockwell, A.E.; Barber, J.C.; Earnshaw, W.C.; Axelman, J.; Migeon, B.R.; Tyler-Smith, C. Centromeric inactivation in a dicentric human Y;21 translocation chromosome. Chromosoma 1997, 106, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Hasson, D.; Alonso, A.; Cheung, F.; Tepperberg, J.H.; Papenhausen, P.R.; Engelen, J.J.; Warburton, P.E. Formation of novel CENP-A domains on tandem repetitive DNA and across chromosome breakpoints on human chromosome 8q21 neocentromeres. Chromosoma 2011, 120, 621–632. [Google Scholar] [CrossRef]
- Warburton, P.E.; Cooke, C.A.; Bourassa, S.; Vafa, O.; Sullivan, B.A.; Stetten, G.; Gimelli, G.; Warburton, D.; Tyler-Smith, C.; Sullivan, K.F.; et al. Immunolocalization of CENP-A suggests a distinct nucleosome structure at the inner kinetochore plate of active centromeres. Curr. Biol. 1997, 7, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Aagaard, L.; Schmid, M.; Warburton, P.; Jenuwein, T. Mitotic phosphorylation of SUV39H1, a novel component of active centromeres, coincides with transient accumulation at mammalian centromeres. J. Cell Sci. 2000, 113 Pt 5, 817–829. [Google Scholar] [CrossRef]
- Agudo, M.; Abad, J.P.; Molina, I.; Losada, A.; Ripoll, P.; Villasante, A. A dicentric chromosome of Drosophila melanogaster showing alternate centromere inactivation. Chromosoma 2000, 109, 190–196. [Google Scholar] [CrossRef]
- Haber, J.E.; Thorburn, P.C.; Rogers, D. Meiotic and mitotic behavior of dicentric chromosomes in Saccharomyces cerevisiae. Genetics 1984, 106, 185–205. [Google Scholar] [CrossRef] [PubMed]
- Haber, J.E.; Thorburn, P.C. Healing of broken linear dicentric chromosomes in yeast. Genetics 1984, 106, 207–226. [Google Scholar] [CrossRef] [PubMed]
- Pennaneach, V.; Kolodner, R.D. Stabilization of dicentric translocations through secondary rearrangements mediated by multiple mechanisms in S. cerevisiae. PLoS ONE 2009, 4, e6389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, A.; Bloom, K. Acquisition and processing of a conditional dicentric chromosome in Saccharomyces cerevisiae. Mol. Cell Biol. 1989, 9, 1368–1370. [Google Scholar] [CrossRef]
- Kramer, K.M.; Brock, J.A.; Bloom, K.; Moore, J.K.; Haber, J.E. Two different types of double-strand breaks in Saccharomyces cerevisiae are repaired by similar RAD52-independent, nonhomologous recombination events. Mol. Cell Biol. 1994, 14, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Long, S.; Stanton, J.; Cusick, P.; Lawrimore, C.; Yeh, E.; Grant, S.; Bloom, K. Behavior of dicentric chromosomes in budding yeast. PLoS Genet. 2021, 17, e1009442. [Google Scholar] [CrossRef]
- Niwa, O.; Matsumoto, T.; Yanagida, M. Construction of a mini-chromosome by deletion and its mitotic and meiotic behaviour in fission yeast. Mol. Gen. Genet. 1986, 203, 397–405. [Google Scholar] [CrossRef]
- Matsumoto, T.; Fukui, K.; Niwa, O.; Sugawara, N.; Szostak, J.W.; Yanagida, M. Identification of healed terminal DNA fragments in linear minichromosomes of Schizosaccharomyces pombe. Mol. Cell Biol. 1987, 7, 4424–4430. [Google Scholar] [CrossRef]
- Szostak, J.W.; Blackburn, E.H. Cloning yeast telomeres on linear plasmid vectors. Cell 1982, 29, 245–255. [Google Scholar] [CrossRef]
- Kramer, K.M.; Haber, J.E. New telomeres in yeast are initiated with a highly selected subset of TG1-3 repeats. Genes. Dev. 1993, 7, 2345–2356. [Google Scholar] [CrossRef] [Green Version]
- Schulz, V.P.; Zakian, V.A. The saccharomyces PIF1 DNA helicase inhibits telomere elongation and de novo telomere formation. Cell 1994, 76, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Lydeard, J.R.; Lipkin-Moore, Z.; Jain, S.; Eapen, V.V.; Haber, J.E. Sgs1 and exo1 redundantly inhibit break-induced replication and de novo telomere addition at broken chromosome ends. PLoS Genet. 2010, 6, e1000973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, W.H.; Zhu, Z.; Papusha, A.; Malkova, A.; Ira, G. Defective resection at DNA double-strand breaks leads to de novo telomere formation and enhances gene targeting. PLoS Genet. 2010, 6, e1000948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negrini, S.; Ribaud, V.; Bianchi, A.; Shore, D. DNA breaks are masked by multiple Rap1 binding in yeast: Implications for telomere capping and telomerase regulation. Genes. Dev. 2007, 21, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Hirano, Y.; Sugimoto, K. Cdc13 telomere capping decreases Mec1 association but does not affect Tel1 association with DNA ends. Mol. Biol. Cell 2007, 18, 2026–2036. [Google Scholar] [CrossRef] [Green Version]
- Hirano, Y.; Fukunaga, K.; Sugimoto, K. Rif1 and rif2 inhibit localization of tel1 to DNA ends. Mol. Cell 2009, 33, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Durocher, D. De novo telomere formation is suppressed by the Mec1-dependent inhibition of Cdc13 accumulation at DNA breaks. Genes. Dev. 2010, 24, 502–515. [Google Scholar] [CrossRef] [Green Version]
- Putnam, C.D.; Pennaneach, V.; Kolodner, R.D. Chromosome healing through terminal deletions generated by de novo telomere additions in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2004, 101, 13262–13267. [Google Scholar] [CrossRef] [Green Version]
- Putnam, C.D.; Hayes, T.K.; Kolodner, R.D. Specific pathways prevent duplication-mediated genome rearrangements. Nature 2009, 460, 984–989. [Google Scholar] [CrossRef] [Green Version]
- Myung, K.; Kolodner, R.D. Induction of genome instability by DNA damage in Saccharomyces cerevisiae. DNA Repair. (Amst.) 2003, 2, 243–258. [Google Scholar] [CrossRef]
- Myung, K.; Kolodner, R.D. Suppression of genome instability by redundant S-phase checkpoint pathways in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 4500–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zschenker, O.; Kulkarni, A.; Miller, D.; Reynolds, G.E.; Granger-Locatelli, M.; Pottier, G.; Sabatier, L.; Murnane, J.P. Increased sensitivity of subtelomeric regions to DNA double-strand breaks in a human cancer cell line. DNA Repair. (Amst.) 2009, 8, 886–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prudden, J.; Evans, J.S.; Hussey, S.P.; Deans, B.; O’Neill, P.; Thacker, J.; Humphrey, T. Pathway utilization in response to a site-specific DNA double-strand break in fission yeast. EMBO J. 2003, 22, 1419–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cullen, J.K.; Hussey, S.P.; Walker, C.; Prudden, J.; Wee, B.Y.; Dave, A.; Findlay, J.S.; Savory, A.P.; Humphrey, T.C. Break-induced loss of heterozygosity in fission yeast: Dual roles for homologous recombination in promoting translocations and preventing de novo telomere addition. Mol. Cell Biol. 2007, 27, 7745–7757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tinline-Purvis, H.; Savory, A.P.; Cullen, J.K.; Dave, A.; Moss, J.; Bridge, W.L.; Marguerat, S.; Bahler, J.; Ragoussis, J.; Mott, R.; et al. Failed gene conversion leads to extensive end processing and chromosomal rearrangements in fission yeast. EMBO J. 2009, 28, 3400–3412. [Google Scholar] [CrossRef] [Green Version]
- Blaikley, E.J.; Tinline-Purvis, H.; Kasparek, T.R.; Marguerat, S.; Sarkar, S.; Hulme, L.; Hussey, S.; Wee, B.Y.; Deegan, R.S.; Walker, C.A.; et al. The DNA damage checkpoint pathway promotes extensive resection and nucleotide synthesis to facilitate homologous recombination repair and genome stability in fission yeast. Nucleic Acids Res. 2014, 42, 5644–5656. [Google Scholar] [CrossRef] [Green Version]
- Dave, A.; Pai, C.C.; Durley, S.C.; Hulme, L.; Sarkar, S.; Wee, B.Y.; Prudden, J.; Tinline-Purvis, H.; Cullen, J.K.; Walker, C.; et al. Homologous recombination repair intermediates promote efficient de novo telomere addition at DNA double-strand breaks. Nucleic Acids Res. 2020, 48, 1271–1284. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Okamoto, A.; Katou, Y.; Yadani, C.; Shitanda, T.; Kaweeteerawat, C.; Takahashi, T.S.; Itoh, T.; Shirahige, K.; Masukata, H.; et al. Rad51 suppresses gross chromosomal rearrangement at centromere in Schizosaccharomyces pombe. EMBO J. 2008, 27, 3036–3046. [Google Scholar] [CrossRef] [Green Version]
- Onaka, A.T.; Su, J.; Katahira, Y.; Tang, C.; Zafar, F.; Aoki, K.; Kagawa, W.; Niki, H.; Iwasaki, H.; Nakagawa, T. DNA replication machinery prevents Rad52-dependent single-strand annealing that leads to gross chromosomal rearrangements at centromeres. Commun. Biol. 2020, 3, 202. [Google Scholar] [CrossRef]
- Su, J.; Xu, R.; Mongia, P.; Toyofuku, N.; Nakagawa, T. Fission yeast Rad8/HLTF facilitates Rad52-dependent chromosomal rearrangements through PCNA lysine 107 ubiquitination. PLoS Genet. 2021, 17, e1009671. [Google Scholar] [CrossRef]
- Marshall, O.J.; Chueh, A.C.; Wong, L.H.; Choo, K.H. Neocentromeres: New insights into centromere structure, disease development, and karyotype evolution. Am. J. Hum. Genet. 2008, 82, 261–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mythreye, K.; Bloom, K.S. Differential kinetochore protein requirements for establishment versus propagation of centromere activity in Saccharomyces cerevisiae. J. Cell Biol. 2003, 160, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diotti, R.; Esposito, M.; Shen, C.H. Telomeric and Sub-Telomeric Structure and Implications in Fungal Opportunistic Pathogens. Microorganisms 2021, 9, 1405. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.C.; Xu, Z.; Karari, K.; Williams, W.; Hauf, S.; Brown, W.R.A. Mutation and selection explain why many eukaryotic centromeric DNA sequences are often A + T rich. Nucleic Acids Res. 2022, 50, 579–596. [Google Scholar] [CrossRef]
- Maggert, K.A.; Karpen, G.H. The activation of a neocentromere in Drosophila requires proximity to an endogenous centromere. Genetics 2001, 158, 1615–1628. [Google Scholar] [CrossRef] [PubMed]
- Burrack, L.S.; Hutton, H.F.; Matter, K.J.; Clancey, S.A.; Liachko, I.; Plemmons, A.E.; Saha, A.; Power, E.A.; Turman, B.; Thevandavakkam, M.A.; et al. Neocentromeres Provide Chromosome Segregation Accuracy and Centromere Clustering to Multiple Loci along a Candida albicans Chromosome. PLoS Genet. 2016, 12, e1006317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreekumar, L.; Jaitly, P.; Chen, Y.; Thimmappa, B.C.; Sanyal, A.; Sanyal, K. Cis- and Trans-chromosomal Interactions Define Pericentric Boundaries in the Absence of Conventional Heterochromatin. Genetics 2019, 212, 1121–1132. [Google Scholar] [CrossRef]
- Nishimura, K.; Komiya, M.; Hori, T.; Itoh, T.; Fukagawa, T. 3D genomic architecture reveals that neocentromeres associate with heterochromatin regions. J. Cell Biol. 2019, 218, 134–149. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; McHugh, T.; Kelly, D.A.; Pidoux, A.L.; Allshire, R.C. Establishment of centromere identity is dependent on nuclear spatial organization. Curr. Biol. 2022, 32, 3121–3136 e3126. [Google Scholar] [CrossRef]
- Luo, J.; Sun, X.; Cormack, B.P.; Boeke, J.D. Karyotype engineering by chromosome fusion leads to reproductive isolation in yeast. Nature 2018, 560, 392–396. [Google Scholar] [CrossRef]
- Shao, Y.; Lu, N.; Wu, Z.; Cai, C.; Wang, S.; Zhang, L.L.; Zhou, F.; Xiao, S.; Liu, L.; Zeng, X.; et al. Creating a functional single-chromosome yeast. Nature 2018, 560, 331–335. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Lu, N.; Cai, C.; Zhou, F.; Wang, S.; Zhao, Z.; Zhao, G.; Zhou, J.Q.; Xue, X.; Qin, Z. A single circular chromosome yeast. Cell Res. 2019, 29, 87–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, X.; Ye, T.; Zhang, X.R.; Nie, L.; Wang, H.; Li, W.; Lu, R.; Fu, C.; Du, L.L.; Zhou, J.Q. Single-chromosome fission yeast models reveal the configuration robustness of a functional genome. Cell Rep. 2022, 40, 111237. [Google Scholar] [CrossRef]
- Wu, Z.J.; Liu, J.C.; Man, X.; Gu, X.; Li, T.Y.; Cai, C.; He, M.H.; Shao, Y.; Lu, N.; Xue, X.; et al. Cdc13 is predominant over Stn1 and Ten1 in preventing chromosome end fusions. Elife 2020, 9, e53144. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Telomere Fusion | Centromere Inactivation | Telomere Healing | Neocentromere Formation | ||
|---|---|---|---|---|---|
| (DNA Deletion) | (Epigenetic Loss) | ||||
| Human *1 | ~4 × 10−6 [45] | <50% [53] | — | ~1 × 10−7 [54] | ~8 × 10−6 [55] |
| S. pombe | ~1 × 10−4 *2 [56] | ~7 × 10−4 [57] | ~3 × 10−3 [57] | ~8 × 10−4 *5 [57] | ~2 × 10−4 [58] |
| ~6 × 10−5 *3 [58] | ~2 × 10−9 *6 [59] | ||||
| S. cerevisiae | <1 × 10−3 *7 [60] | ||||
| ~1 × 10−7 [61] | ~2% [62,63] | — | >90% *8 [64] | <1 × 10−7 [61] | |
| ~3 × 10−10 *6 [65,66] | |||||
| C. deuterogattii | u.f. *4 [67] | — | u.f. *4 [68] | — | <1 × 10−3 [67] |
| C. albicans | — | — | — | — | ~100% [69,70] |
| Chicken | ~3 × 10−7 [71] | — | — | — | ~3 × 10−6 [71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuse, R.; Ishii, K. Flexible Attachment and Detachment of Centromeres and Telomeres to and from Chromosomes. Biomolecules 2023, 13, 1016. https://doi.org/10.3390/biom13061016
Kuse R, Ishii K. Flexible Attachment and Detachment of Centromeres and Telomeres to and from Chromosomes. Biomolecules. 2023; 13(6):1016. https://doi.org/10.3390/biom13061016
Chicago/Turabian StyleKuse, Riku, and Kojiro Ishii. 2023. "Flexible Attachment and Detachment of Centromeres and Telomeres to and from Chromosomes" Biomolecules 13, no. 6: 1016. https://doi.org/10.3390/biom13061016