
Distribution of α2-Adrenergic Receptors in the Living Human Brain Using [11C]yohimbine PET
 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. PET Procedures
2.3. Image Processing
2.4. Statistical Analysis
3. Results
3.1. Regional ROI Analysis
3.2. Subregional ROI Analyses
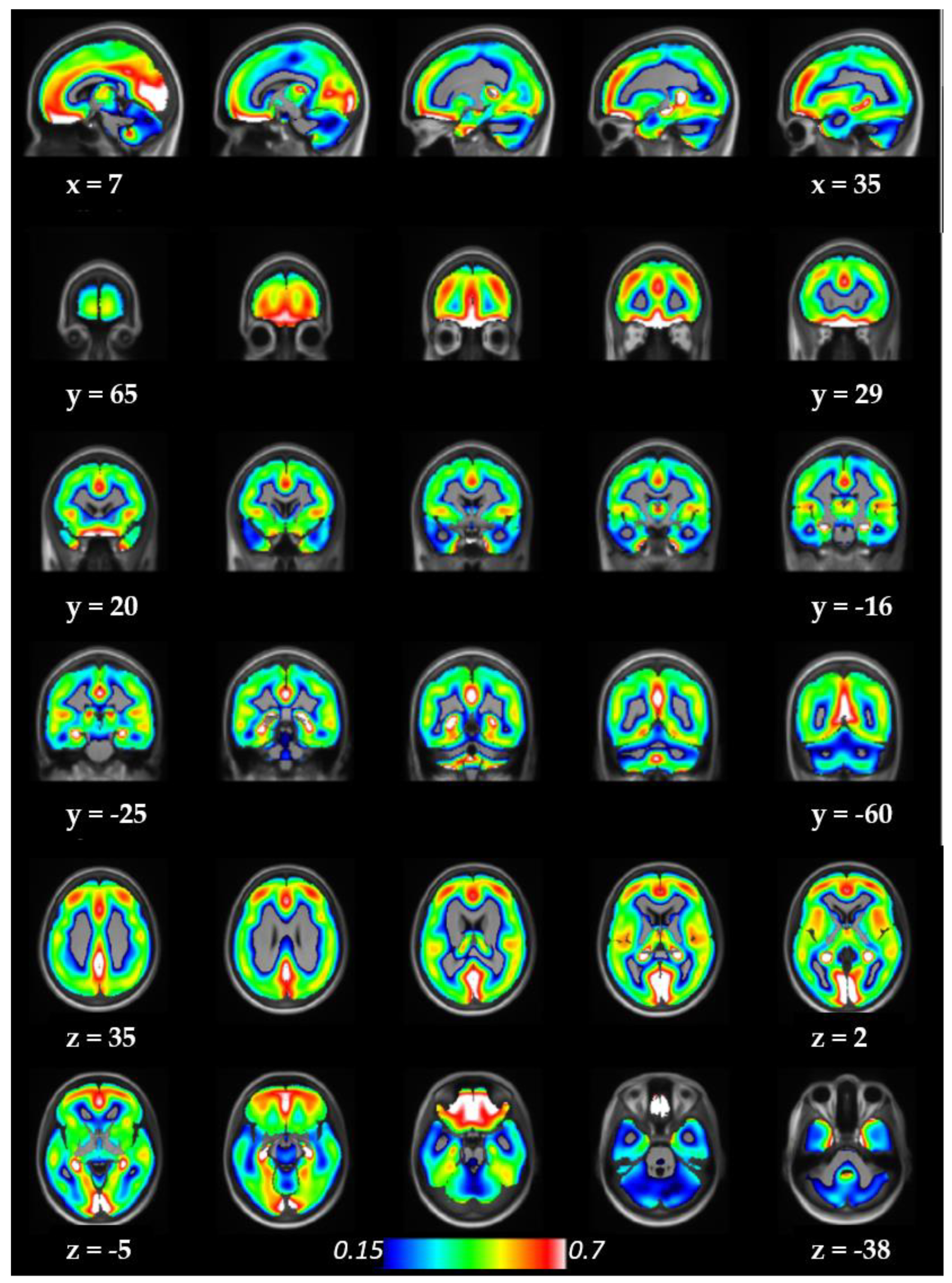
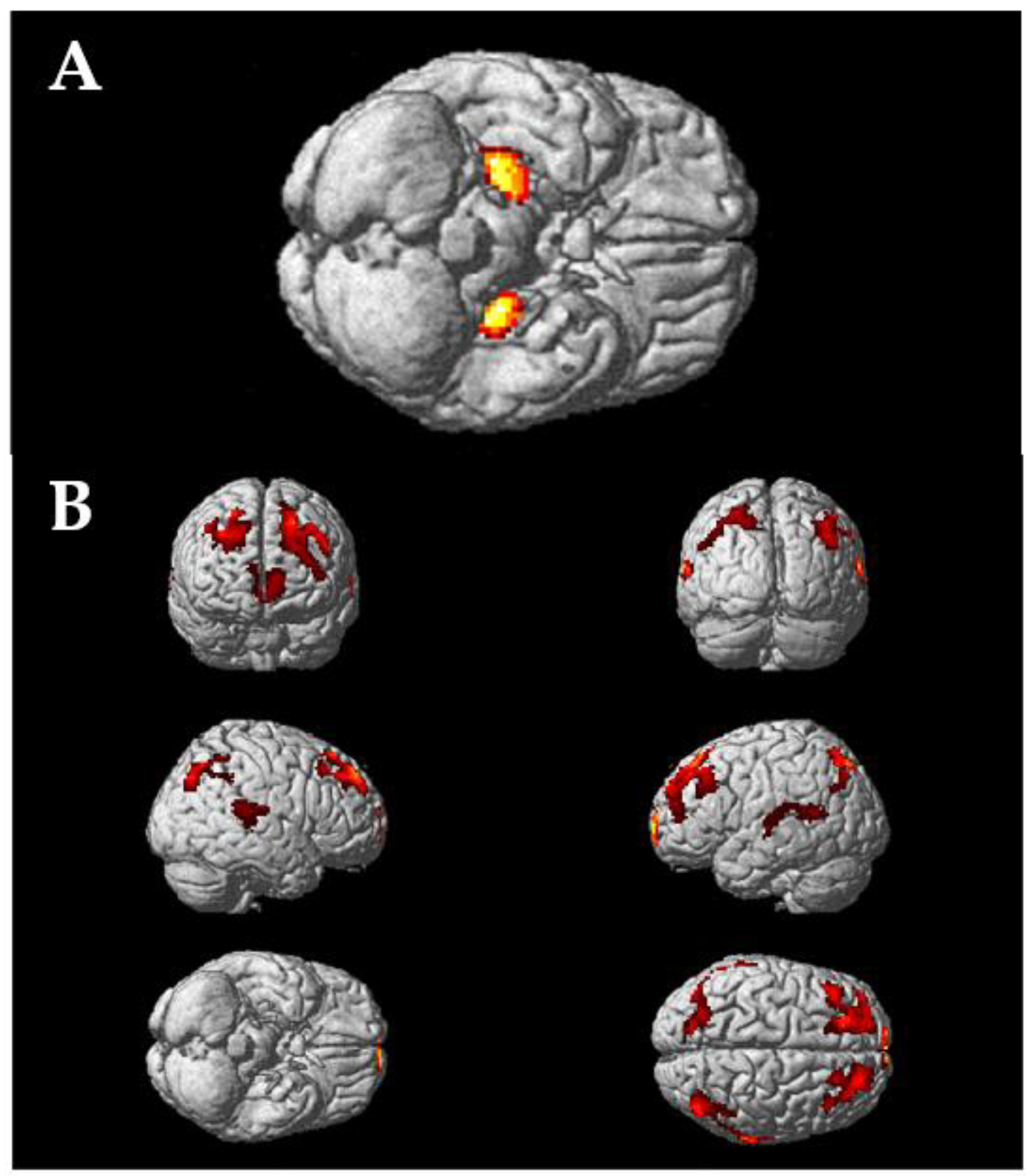
3.3. Whole Brain Illustrations
4. Discussion
4.1. Regional Distribution of [11C]yohimbine
4.2. Subregional Variations in α2-ARs Availability
4.3. New Issues
4.4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Holland, N.; Robbins, T.W.; Rowe, J.B. The Role of Noradrenaline in Cognition and Cognitive Disorders. Brain 2021, 144, 2243–2256. [Google Scholar] [CrossRef] [PubMed]
- Sara, S.J.; Bouret, S. Orienting and Reorienting: The Locus Coeruleus Mediates Cognition through Arousal. Neuron 2012, 76, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Berridge, C.W.; Waterhouse, B.D. The Locus Coeruleus–Noradrenergic System: Modulation of Behavioral State and State-Dependent Cognitive Processes. Brain Res. Rev. 2003, 42, 33–84. [Google Scholar] [CrossRef] [PubMed]
- Aston-Jones, G.; Cohen, J.D. Adaptive Gain and the Role of the Locus Coeruleus-Norepinephrine System in Optimal Performance. J. Comp. Neurol. 2005, 493, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Singh, S. Noradrenergic Pathways of Locus Coeruleus in Parkinson’s and Alzheimer’s Pathology. Int. J. Neurosci. 2020, 130, 251–261. [Google Scholar] [CrossRef]
- Weinshenker, D. Long Road to Ruin: Noradrenergic Dysfunction in Neurodegenerative Disease. Trends Neurosci. 2018, 41, 211–223. [Google Scholar] [CrossRef]
- Shin, E.; Rogers, J.T.; Devoto, P.; Björklund, A.; Carta, M. Noradrenaline Neuron Degeneration Contributes to Motor Impairments and Development of L-DOPA-Induced Dyskinesia in a Rat Model of Parkinson’s Disease. Exp. Neurol. 2014, 257, 25–38. [Google Scholar] [CrossRef]
- Delaville, C.; Deurwaerdère, P.D.; Benazzouz, A. Noradrenaline and Parkinson’s Disease. Front. Syst. Neurosci. 2011, 5, 31. [Google Scholar] [CrossRef]
- Marien, M.R.; Colpaert, F.C.; Rosenquist, A.C. Noradrenergic Mechanisms in Neurodegenerative Diseases: A Theory. Brain Res. Rev. 2004, 45, 38–78. [Google Scholar] [CrossRef]
- Braak, H.; Tredici, K.D.; Rüb, U.; de Vos, R.A.I.; Jansen Steur, E.N.H.; Braak, E. Staging of Brain Pathology Related to Sporadic Parkinson’s Disease. Neurobiol. Aging 2003, 24, 197–211. [Google Scholar] [CrossRef]
- Cui, K.; Yang, F.; Tufan, T.; Raza, M.U.; Zhan, Y.; Fan, Y.; Zeng, F.; Brown, R.W.; Price, J.B.; Jones, T.C.; et al. Restoration of Noradrenergic Function in Parkinson’s Disease Model Mice. ASN Neuro 2021, 13, 175909142110097. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Merlo, S.; Drago, F.; Caruso, G.; Parenti, C.; Sortino, M.A. Rescue of Noradrenergic System as a Novel Pharmacological Strategy in the Treatment of Chronic Pain: Focus on Microglia Activation. Front. Pharmacol. 2019, 10, 1024. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.; Harkin, A. Targeting the Noradrenergic System for Anti-Inflammatory and Neuroprotective Effects: Implications for Parkinson’s Disease. Neural Regen. Res. 2018, 13, 1332. [Google Scholar] [CrossRef] [PubMed]
- Nutt, D.J.; McAllister–Williams, R.H. Noradrenaline—The Forgotten Amine? J. Psychopharmacol. 2013, 27, 657–658. [Google Scholar] [CrossRef]
- Fornai, F.; di Poggio, A.; Pellegrini, A.; Ruggieri, S.; Paparelli, A. Noradrenaline in Parkinsons Disease: From Disease Progression to Current Therapeutics. Curr. Med. Chem. 2007, 14, 2330–2334. [Google Scholar] [CrossRef]
- Probst, A.; Cortés, R.; Palacios, J.M. Distribution of A2-Adrenergic Receptors in the Human Brainstem: An Autoradiographic Study Using [3H]p-Aminoclonidine. Eur. J. Pharmacol. 1984, 106, 477–488. [Google Scholar] [CrossRef]
- Pazos, A. Aa-Adrenoceptors in Human Forebrain: Autoradiographic Visualization and Biochemical Parameters Using the Agonist [AH]UK-14304. Brain Res. 1988, 475, 361–365. [Google Scholar] [CrossRef]
- Jones, C.R.; Palacios, J.M. Autoradiography of Adrenoceptors in Rat and Human Brain: α-Adrenoceptor and Idazoxan Binding Sites. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 1991; Volume 88, pp. 271–291. ISBN 978-0-444-81394-7. [Google Scholar]
- Vos, H.; Vauquelin, G.; Keyser, J.; Backer, J.-P.; Liefde, I. Regional Distribution of α2A- and α2B-Adrenoceptor Subtypes in Postmortem Human Brain. J. Neurochem. 1992, 58, 1555–1560. [Google Scholar] [CrossRef]
- Pascual, J.; Del Arco, C.; Gonzalez, A.; Diaz, A.; del Olmo, E.; Pazos, A. Regionally Specific Age-Dependent Decline in α2-Adrenoceptors: An Autoradiograpic Study in Human Brain. Neurosci. Lett. 1991, 133, 279–283. [Google Scholar] [CrossRef]
- Pascual, J.; del Arco, C.; Gonzalez, A.; Pazos, A. Quantitative Light Microscopic Autoradiographic Localization of A2-Adrenoceptors in the Human Brain. Brain Res. 1992, 582, 116–127. [Google Scholar] [CrossRef]
- Ordway, G.A.; Jaconetta, S.M.; Halaris, A.E. Characterization of Subtypes of Alpha-2 Adrenoceptors in the Human Brain. J. Pharmacol. Exp. Ther. 1993, 264, 967–976. [Google Scholar] [PubMed]
- Fagerholm, V.; Rokka, J.; Nyman, L.; Sallinen, J.; Tiihonen, J.; Tupala, E.; Haaparanta, M.; Hietala, J. Autoradiographic Characterization of A2C-Adrenoceptors in the Human Striatum. Synapse 2008, 62, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Vos, H.D.; Convents, A.; Keyser, J.D.; Backer, J.-P.D.; Megena, V.; Ebinger, G.; Vauquelin, G. Autoradiographic Distribution of A2 Adrenoceptors, NAIBS, and 5-I-IT1A Receptors in Human Brain Using [3H]Idazoxan and [3H]Rauwolscine. Brain Res. 1991, 566, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Sastre, M.; Garcia-Sevilla, J.A. Opposite Age-Dependent Changes of A2A-Adrenoceptors and Nonadrenoceptor [3H]Idazoxan Binding Sites (I2-Imidazoline Sites) in the Human Brain: Strong Correlation of I2 with Monoamine Oxidase-B Sites. J. Neurochem. 1993, 61, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Biegon, A.; Mathis, C.A.; Budinger, T.F. Quantitative in Vitro and Ex Vivo Autoradiography of the A2-Adrenoceptor Antagonist [3H]Atopamezole. Eur. J. Pharmacol. 1992, 224, 27–38. [Google Scholar] [CrossRef]
- Nahimi, A.; Jakobsen, S.; Munk, O.L.; Vang, K.; Phan, J.A.; Rodell, A.; Gjedde, A. Mapping 2 Adrenoceptors of the Human Brain with 11C-Yohimbine. J. Nucl. Med. 2015, 56, 392–398. [Google Scholar] [CrossRef]
- Laurencin, C.; Lancelot, S.; Gobert, F.; Redouté, J.; Mérida, I.; Iecker, T.; Liger, F.; Irace, Z.; Greusard, E.; Lamberet, L.; et al. Modeling [11C]Yohimbine PET Human Brain Kinetics with Test-Retest Reliability, Competition Sensitivity Studies and Search for a Suitable Reference Region. NeuroImage 2021, 140, 118328. [Google Scholar] [CrossRef]
- Jakobsen, S.; Pedersen, K.; Smith, D.F.; Jensen, S.B.; Munk, O.L.; Cumming, P. Detection of A2-Adrenergic Receptors in Brain of Living Pig with 11C-Yohimbine. J. Nucl. Med. 2006, 47, 2008–2015. [Google Scholar]
- Reilhac, A.; Merida, I.; Irace, Z.; Stephenson, M.C.; Weekes, A.A.; Chen, C.; Totman, J.J.; Townsend, D.W.; Fayad, H.; Costes, N. Development of a Dedicated Rebinner with Rigid Motion Correction for the MMR PET/MR Scanner, and Validation in a Large Cohort of 11C-PIB Scans. J. Nucl. Med. 2018, 59, 1761–1767. [Google Scholar] [CrossRef]
- Mérida, I.; Reilhac, A.; Redouté, J.; Heckemann, R.A.; Costes, N.; Hammers, A. Multi-Atlas Attenuation Correction Supports Full Quantification of Static and Dynamic Brain PET Data in PET-MR. Phys. Med. Biol. 2017, 62, 2834–2858. [Google Scholar] [CrossRef]
- Ashburner, J.; Friston, K.J. Unified Segmentation. NeuroImage 2005, 26, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Heckemann, R.A.; Keihaninejad, S.; Aljabar, P.; Rueckert, D.; Hajnal, J.V.; Hammers, A. Improving Intersubject Image Registration Using Tissue-Class Information Benefits Robustness and Accuracy of Multi-Atlas Based Anatomical Segmentation. NeuroImage 2010, 51, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Hammers, A.; Allom, R.; Koepp, M.J.; Free, S.L.; Myers, R.; Lemieux, L.; Mitchell, T.N.; Brooks, D.J.; Duncan, J.S. Three-Dimensional Maximum Probability Atlas of the Human Brain, with Particular Reference to the Temporal Lobe. Hum. Brain Mapp. 2003, 19, 224–247. [Google Scholar] [CrossRef] [PubMed]
- Heckemann, R.A.; Hajnal, J.V.; Aljabar, P.; Rueckert, D.; Hammers, A. Automatic Anatomical Brain MRI Segmentation Combining Label Propagation and Decision Fusion. NeuroImage 2006, 33, 115–126. [Google Scholar] [CrossRef]
- Javier Meana, J.; Barturen, F.; Asier Garro, M.; García-Sevilla, J.A.; Fontán, A.; Zarranz, J.J. Decreased Density of Presynaptic A2-Adrenoceptors in Postmortem Brains of Patients with Alzheimer’s Disease. J. Neurochem. 1992, 58, 1896–1904. [Google Scholar] [CrossRef]
- Meana, J.J.; Barturen, F. α2-Adrenoceptors in the Brain of Suicide Victims: Increased Receptor Density Associated with Major Depression. Biol. Psychiatry 1992, 31, 471–490. [Google Scholar] [CrossRef]
- González, A.M.; Pascual, J.; Meana, J.J.; Barturen, F.; Del Arco, C.; Pazos, A.; García-Sevilla, J.A. Autoradiographic Demonstration of Increased A2-Adrenoceptor Agonist Binding Sites in the Hippocampus and Frontal Cortex of Depressed Suicide Victims. J. Neurochem. 1994, 63, 256–265. [Google Scholar] [CrossRef]
- Yuan, P.; Raz, N. Prefrontal Cortex and Executive Functions in Healthy Adults: A Meta-Analysis of Structural Neuroimaging Studies. Neurosci. Biobehav. Rev. 2014, 42, 180–192. [Google Scholar] [CrossRef]
- Volz, K.G.; Schubotz, R.I.; von Cramon, D.Y. Decision-Making and the Frontal Lobes. Curr. Opin. Neurol. 2006, 19, 401–406. [Google Scholar] [CrossRef]
- Rolls, E.T. The Functions of the Orbitofrontal Cortex. Brain Cogn. 2004, 55, 11–29. [Google Scholar] [CrossRef]
- Schall, J.D.; Stuphorn, V.; Brown, J.W. Monitoring and Control of Action by the Frontal Lobes. Neuron 2002, 36, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Chayer, C.; Freedman, M. Frontal Lobe Functions. Curr. Neurol. Neurosci. Rep. 2001, 1, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Talley, E.M.; Rosin, D.L.; Lee, A.; Guyenet, P.G.; Lynch, K.R. Distribution of α2A-Adrenergic Receptor-like Immunoreactivity in the Rat Central Nervous System. J. Comp. Neurol. 1996, 372, 111–134. [Google Scholar] [CrossRef]
- Strazielle, C.; Lalonde, R.; Hébert, C.; Reader, T.A. Regional Brain Distribution of Noradrenaline Uptake Sites, and of α1-α2- and Beta-Adrenergic Receptors in PCD Mutant Mice: A Quantitative Autoradiographic Study. Neuroscience 1999, 94, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Santos, I.; Palomero-Gallagher, N.; Zilles, K.; Cavada, C. Distribution of the Noradrenaline Innervation and Adrenoceptors in the Macaque Monkey Thalamus. Cereb. Cortex 2021, 31, 4115–4139. [Google Scholar] [CrossRef]
- Landau, A.M.; Doudet, D.J.; Jakobsen, S. Amphetamine Challenge Decreases Yohimbine Binding to A2 Adrenoceptors in Landrace Pig Brain. Psychopharmacology 2012, 222, 155–163. [Google Scholar] [CrossRef]
- Wilkinson, M.; Herdon, H.; Pearce, M.; Wilson, C. Radioligand Binding Studies on Hypothalamic Noradrenergic Receptors during the Estrous Cycle or after Steroid Injection in Ovariectomized Rats. Brain Res. 1979, 168, 652–655. [Google Scholar] [CrossRef]
- Wilkinson, M.; Herdon, H.J. Diethylstilbestrol Regulates the Number of A- and -Adrenergic Binding Sites in Incubated Hypothalamus and Amygdala. Brain Res. 1982, 248, 79–85. [Google Scholar] [CrossRef]
- Karkanias, G.B.; Etgen, A.M. Estradiol Reduction of the Agonist High Affinity Form of the Alpha 2-Adrenoceptor in the Hypothalamus of Female Rats: Identification as the Alpha 2D Subtype. Mol. Pharmacol. 1994, 45, 509–516. [Google Scholar]
- Karkanias, G.B.; Etgen, A.M. Estradiol Attenuates Alpha 2-Adrenoceptor-Mediated Inhibition of Hypothalamic Norepinephrine Release. J. Neurosci. 1993, 13, 3448–3455. [Google Scholar] [CrossRef]
- Eaton, W.W.; Kessler, R.C.; Wittchen, H.U.; Magee, W.J. Panic and Panic Disorder in the United States. Am. J. Psychiatry 1994, 151, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Oquendo, M.A.; Ellis, S.P.; Greenwald, S.; Malone, K.M.; Weissman, M.M.; Mann, J.J. Ethnic and Sex Differences in Suicide Rates Relative to Major Depression in the United States. Am. J. Psychiatry 2001, 158, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Bylund, D.B. Pharmacological Characteristics of Alpha-2 Adrenergic Receptor Subtypes. Ann. N. Y. Acad. Sci. 1995, 763, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Altman, J.D.; Trendelenburg, A.U.; Macmillan, L.; Bernstein, D.; Limbird, L.; Starke, K.; Kobilka, B.K.; Hein, L. Abnormal Regulation of the Sympathetic Nervous System in α2A-Adrenergic Receptor Knockout Mice. Mol. Pharmacol. 1999, 56, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Hein, L.; Altman, J.D.; Kobilka, B.K. Two Functionally Distinct A2-Adrenergic Receptors Regulate Sympathetic Neurotransmission. Nature 1999, 402, 181. [Google Scholar] [CrossRef]
- Vonend, O.; Habbel, S.; Stegbauer, J.; Roth, J.; Hein, L.; Rump, L.C. α2A-Adrenoceptors Regulate Sympathetic Transmitter Release in Mice Kidneys. Br. J. Pharmacol. 2007, 150, 121–127. [Google Scholar] [CrossRef]
- Bücheler, M.M.; Hadamek, K.; Hein, L. Two A2-Adrenergic Receptor Subtypes, A2A and A2C, Inhibit Transmitter Release in the Brain of Gene-Targeted Mice. Neuroscience 2002, 109, 819–826. [Google Scholar] [CrossRef]
- Scheibner, J.; Trendelenburg, A.U.; Hein, L.; Starke, K. α2-Adrenoceptors Modulating Neuronal Serotonin Release: A Study in α2-Adrenoceptor Subtype-Deficient Mice. Br. J. Pharmacol. 2001, 132, 925–933. [Google Scholar] [CrossRef]
- Langer, S.Z. Presynaptic Regulation of the Release of Catecholamines. Pharmacol. Rev. 1980, 32, 337–362. [Google Scholar] [CrossRef]
- Heal, D.J.; Butler, S.A.; Prow, M.R.; Buckett, W.R. Quantification of Presynaptic α2-Adrenoceptors in Rat Brain after Short-Term DSP-4 Lesioning. Eur. J. Pharmacol. 1993, 249, 37–41. [Google Scholar] [CrossRef]
- Szabadi, E. Functional Neuroanatomy of the Central Noradrenergic System. J. Psychopharmacol. 2013, 27, 659–693. [Google Scholar] [CrossRef] [PubMed]
- Langer, S.Z. Presynaptic Receptors and Modulation of Neurotransmission: Pharmacological Implications and Therapeutic Relevance. Trends Neurosci. 1980, 3, 110–112. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| [11C]yohimbine BPND | |||
|---|---|---|---|
| Females | Males | All | |
| Hippocampus | 0.67 ± 0.23 | 0.55 ± 0.17 | 0.61 ± 0.21 |
| Occipital Lobe | 0.57 ± 0.21 | 0.60 ± 0.20 | 0.58 ± 0.20 |
| Cingulate Gyrus | 0.55 ± 0.14 | 0.56 ± 0.15 | 0.56 ± 0.14 |
| Frontal Lobe | 0.50 ± 0.17 | 0.54 ± 0.16 | 0.52 ± 0.17 |
| Parietal Lobe | 0.45 ± 0.13 | 0.51 ± 0.14 | 0.48 ± 0.14 |
| Thalamus | 0.46 ± 0.16 | 0.49 ± 0.13 | 0.48 ± 0.15 |
| Parahippocampus | 0.42 ± 0.12 | 0.44 ± 0.12 | 0.43 ± 0.12 |
| Insula | 0.42 ± 0.13 | 0.43 ± 0.15 | 0.42 ± 0.14 |
| Temporal Lobe | 0.36 ± 0.14 | 0.39 ± 0.15 | 0.38 ± 0.14 |
| Basal Ganglia | 0.30 ± 0.18 | 0.32 ± 0.18 | 0.31 ± 0.18 |
| Amygdala | 0.29 ± 0.10 | 0.30 ± 0.10 | 0.30 ± 0.10 |
| Cerebellum | 0.23 ± 0.18 | 0.31 ± 0.12 | 0.27 ± 0.16 |
| Raphe | 0.22 ± 0.16 | 0.26 ± 0.18 | 0.24 ± 0.17 |
| Hippocampus | Occipital Lobe | Cingulate Gyrus | Frontal Lobe | Parietal Lobe | Thalamus | Parahippocampus | Insula | Temporal Lobe | Basal Ganglia | Amygdala | Cerebellum | Raphe | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hippocampus | |||||||||||||
| Occipital Lobe | ns | ||||||||||||
| Cingulate Gyrus | ns | ns | |||||||||||
| Frontal Lobe | *** | *** | ns | ||||||||||
| Parietal Lobe | *** | *** | *** | *** | |||||||||
| Thalamus | *** | *** | * | ns | ns | ||||||||
| Parahippocampus | *** | *** | *** | *** | ns | ns | |||||||
| Insula | *** | *** | *** | *** | *** | ns | ns | ||||||
| Temporal Lobe | *** | *** | *** | *** | *** | *** | ns | *** | |||||
| Basal Ganglia | *** | *** | *** | *** | *** | *** | *** | *** | *** | ||||
| Amygdala | *** | *** | *** | *** | *** | *** | *** | *** | ** | ns | |||
| Cerebellum | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns | ns | ||
| Raphe | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | ns | ns |
| Inferior Frontal Gyrus | Middle Frontal Gyrus | Orbital Gyrus | Precentral Gyrus | Presubgenual ACC | Straight Gyrus | Subcallosal Area | Subgenual ACC | Superior Frontal Gyrus | |
|---|---|---|---|---|---|---|---|---|---|
| Inferior Frontal Gyrus | |||||||||
| Middle Frontal Gyrus | ns | ||||||||
| Orbital Gyrus | **** | ns | |||||||
| Precentral Gyrus | ns | **** | **** | ||||||
| Presubgenual ACC | * | ns | ns | **** | |||||
| Straight Gyrus | **** | **** | *** | **** | ** | ||||
| Subcallosal Area | **** | **** | **** | * | **** | **** | |||
| Subgenual ACC | ns | **** | **** | ns | **** | **** | ns | ||
| Superior Frontal Gyrus | ns | ns | ** | * | ns | **** | **** | *** |
| [11C]yohimbine BPND | |||
|---|---|---|---|
| Females | Males | All | |
| Cuneus | 0.67 ± 0.17 | 0.71 ± 0.16 | 0.68 ± 0.17 |
| Lingual Gyrus | 0.65 ± 0.18 | 0.67 ± 0.16 | 0.66 ± 0.17 |
| Straight Gyrus | 0.65 ± 0.15 | 0.65 ± 0.14 | 0.65 ± 0.14 |
| Posterior Cingulate Cortex | 0.60 ± 0.15 | 0.62 ± 0.14 | 0.61 ± 0.14 |
| Hippocampus | 0.67 ± 0.23 | 0.55 ± 0.17 | 0.61 ± 0.21 |
| Orbital Gyrus | 0.55 ± 0.17 | 0.60 ± 0.14 | 0.58 ± 0.16 |
| Presubgenual ACC | 0.56 ± 0.18 | 0.57 ± 0.15 | 0.57 ± 0.16 |
| Middle Frontal Gyrus | 0.49 ± 0.14 | 0.58 ± 0.16 | 0.54 ± 0.15 |
| Anterior Cingulate Cortex | 0.51 ± 0.12 | 0.51 ± 0.14 | 0.51 ± 0.13 |
| Superior Frontal Gyrus | 0.49 ± 0.15 | 0.53 ± 0.15 | 0.51 ± 0.15 |
| Superior Parietal Gyrus | 0.47 ± 0.13 | 0.52 ± 0.14 | 0.50 ± 0.14 |
| Inferior Frontal Gyrus | 0.46 ± 0.11 | 0.51 ± 0.14 | 0.49 ± 0.13 |
| Supramarginal Gyrus | 0.46 ± 0.13 | 0.50 ± 0.14 | 0.48 ± 0.14 |
| Angular Gyrus | 0.44 ± 0.14 | 0.51 ± 0.15 | 0.48 ± 0.15 |
| Thalamus | 0.46 ± 0.16 | 0.49 ± 0.13 | 0.48 ± 0.15 |
| Posterior Temporal Lobe | 0.46 ± 0.13 | 0.48 ± 0.13 | 0.47 ± 0.13 |
| Postcentral Gyrus | 0.43 ± 0.13 | 0.49 ± 0.14 | 0.46 ± 0.14 |
| Parahippocampus | 0.42 ± 0.12 | 0.44 ± 0.12 | 0.43 ± 0.12 |
| Precentral Gyrus | 0.40 ± 0.13 | 0.46 ± 0.14 | 0.43 ± 0.14 |
| Subgenual ACC | 0.42 ± 0.13 | 0.41 ± 0.12 | 0.42 ± 0.13 |
| Insula | 0.42 ± 0.13 | 0.43 ± 0.15 | 0.42 ± 0.14 |
| Fusiform gyrus | 0.41 ± 0.12 | 0.41 ± 014 | 0.41 ± 0.13 |
| Nucleus Accumbens | 0.40 ± 0.16 | 0.41 ± 0.13 | 0.41 ± 0.14 |
| Lateral Occipital Lobe | 0.39 ± 0.13 | 0.42 ± 0.14 | 0.40 ± 0.14 |
| Putamen | 0.37 ± 0.18 | 0.43 ± 0.17 | 0.40 ± 0.17 |
| Pallidum | 0.37 ± 0.14 | 0.41 ± 0.16 | 0.39 ± 0.15 |
| Superior Temporal Gyrus | 0.36 ± 0.14 | 0.39 ± 0.16 | 0.38 ± 0.15 |
| Middle and Inferior Temporal Gyrus | 0.37 ± 0.12 | 0.36 ± 0.13 | 0.37 ± 0.12 |
| Subcallosal Area | 0.36 ± 0.14 | 0.35 ± 0.12 | 0.35 ± 0.13 |
| Anterior Temporal Lobe | 0.30 ± 0.13 | 0.33 ± 0.13 | 0.32 ± 0.13 |
| Amygdala | 0.29 ± 0.10 | 0.30 ± 0.10 | 0.30 ± 0.10 |
| Cerebellum | 0.23 ± 0.18 | 0.31 ± 0.12 | 0.27 ± 0.16 |
| Raphe | 0.22 ± 0.16 | 0.26 ± 0.18 | 0.24 ± 0.17 |
| Substantia Nigra | 0.20 ± 0.14 | 0.19 ± 0.14 | 0.19 ± 0.14 |
| Caudate Nucleus | 0.15 ± 0.14 | 0.19 ± 0.09 | 0.17 ± 0.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laurencin, C.; Lancelot, S.; Merida, I.; Costes, N.; Redouté, J.; Le Bars, D.; Boulinguez, P.; Ballanger, B. Distribution of α2-Adrenergic Receptors in the Living Human Brain Using [11C]yohimbine PET. Biomolecules 2023, 13, 843. https://doi.org/10.3390/biom13050843
Laurencin C, Lancelot S, Merida I, Costes N, Redouté J, Le Bars D, Boulinguez P, Ballanger B. Distribution of α2-Adrenergic Receptors in the Living Human Brain Using [11C]yohimbine PET. Biomolecules. 2023; 13(5):843. https://doi.org/10.3390/biom13050843
Chicago/Turabian StyleLaurencin, Chloé, Sophie Lancelot, Inès Merida, Nicolas Costes, Jérôme Redouté, Didier Le Bars, Philippe Boulinguez, and Bénédicte Ballanger. 2023. "Distribution of α2-Adrenergic Receptors in the Living Human Brain Using [11C]yohimbine PET" Biomolecules 13, no. 5: 843. https://doi.org/10.3390/biom13050843