
Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System

Cellular and Molecular Signaling, New York, NY 10022, USA
Biomolecules 2023, 13(5), 816; https://doi.org/10.3390/biom13050816
Submission received: 13 April 2023
/
Revised: 5 May 2023
/
Accepted: 10 May 2023
/
Published: 11 May 2023
Abstract
:It is estimated that, at minimum, 500 million individuals suffer from cellular metabolic dysfunction, such as diabetes mellitus (DM), throughout the world. Even more concerning is the knowledge that metabolic disease is intimately tied to neurodegenerative disorders, affecting both the central and peripheral nervous systems as well as leading to dementia, the seventh leading cause of death. New and innovative therapeutic strategies that address cellular metabolism, apoptosis, autophagy, and pyroptosis, the mechanistic target of rapamycin (mTOR), AMP activated protein kinase (AMPK), growth factor signaling with erythropoietin (EPO), and risk factors such as the apolipoprotein E (APOE-ε4) gene and coronavirus disease 2019 (COVID-19) can offer valuable insights for the clinical care and treatment of neurodegenerative disorders impacted by cellular metabolic disease. Critical insight into and modulation of these complex pathways are required since mTOR signaling pathways, such as AMPK activation, can improve memory retention in Alzheimer’s disease (AD) and DM, promote healthy aging, facilitate clearance of β-amyloid (Aß) and tau in the brain, and control inflammation, but also may lead to cognitive loss and long-COVID syndrome through mechanisms that can include oxidative stress, mitochondrial dysfunction, cytokine release, and APOE-ε4 if pathways such as autophagy and other mechanisms of programmed cell death are left unchecked.
1. Introduction and Overview
Metabolic disorders can affect all systems of the body. In particular, they can be a significant contributor to cognitive loss and dementia and impact both the central nervous system and the peripheral nervous system. Present therapies that may involve pharmacological treatments, immunomodulators, and lifestyle changes for metabolic and neurodegenerative disorders can only address, under most conditions, symptomatic relief and cannot ultimately stop disease progression. Novel investigations into the cellular disease mechanisms of metabolic cellular dysfunction provide exciting and new therapeutic avenues for consideration to treat neurodegenerative disorders with the understanding that these pathways are complex and must be carefully considered to limit any undesirable clinical outcomes.
2. Cellular Metabolism Dysfunction
Cellular metabolism dysfunction, which includes diabetes mellitus (DM), has been rising in prevalence more quickly in low- and middle-income countries than in high-income countries (Table 1). Almost eighty percent of adults with DM are living in low- and middle-income countries [1]. The prevalence of DM has increased from nine and one-half percent during the period of 1999 to 2002 to twelve percent during the period of 2013 to 2016. A number of parameters can affect disease prevalence, such as education, co-morbidities, and socioeconomic status [2,3,4,5]. Almost thirteen percent of adults with less than a high school education have DM compared to ten percent of individuals with a high school education and DM. If one has greater than a high school education, the risk decreases to seven and one-half percent. Additional risk factors for developing complications of DM consist of tobacco consumption, hypertension, exercise, elevated serum cholesterol, and obesity [6,7,8,9,10,11,12,13]. In regard to obesity, increased body weight leads to impaired glucose tolerance that results in DM progression [14,15,16,17,18,19,20,21,22,23,24]. Obesity can especially increase the risk of developing DM in young individuals and can affect stem cell proliferation, aging, inflammation, oxidative stress injury, and mitochondrial function [25,26,27,28,29,30,31,32,33,34].
At least 500 million individuals are believed to suffer from DM throughout the world [2,6,8,35,36,37,38]. Interestingly, approximately forty-five percent of the four million annual deaths that occur in individuals with DM affect those under the age of seventy [39]. In the United States (USA), thirty-five million individuals, approximately ten percent of the population, are diagnosed with DM [40,41]. In addition, greater than four hundred million individuals are believed to either suffer from metabolic disease or be at risk for developing DM [1,14,16,22,26,42,43,44,45,46,47]. Many of these individuals have not received a diagnosis of DM, and it is estimated that at least seven million individuals over the age of 18 are undiagnosed with DM. As an example, it was estimated that almost thirty-five percent of adults in the USA has prediabetes based on their fasting glucose and hemoglobin A1c (HbA1c) levels [3,48].
The financial expenditures for the care and treatment of individuals with DM are considered to be significant [2,37,39,47,49]. At minimum, USD 20,000 per year is required to care for each individual with DM. The care for patients with DM can now exceed USD 760 billion [1]. Treatment and care for individuals with DM uses more than seventeen percent of the gross domestic product in the USA per the Centers for Medicare and Medicaid Services (CMS) [50]. An additional seventy billion USD is consumed for individuals with DM as a result of disability and functional loss.
3. Cellular Metabolism in the Nervous System
Diseases of the nervous system are a significant component of non-communicable diseases (NCDs) [5,51,52]. At least seventy percent of the annual deaths that occur each year are the result of NCDs [53,54]. An observed rise in NCDs parallels an increase in the life expectancy of the global population [55,56,57,58]. Life expectancy is now approaching eighty years of age [59], with a one percent decrease in the age-adjusted death rate from the years 2000 through 2011 [60]. In addition, India and China, examples of developing nations that are growing, will see their elderly population increase from five to ten percent over future years [7,61]. Increased lifespan may be due to multiple factors that include improved access to healthcare, targeted sanitation measures, and greater public health measures that address populations at the highest risk of disability (Table 1). These measures can lead to earlier and more effective treatment for multiple chronic disorders [56,62,63,64,65,66,67,68,69,70,71,72].
The improvements in healthcare access and a concurrent rise in lifespan for the global population has resulted in an increased prevalence of neurodegenerative disorders [28,41,73,74,75,76,77,78]. Diseases of the nervous system represent over six hundred disorders that lead to death and disability [61,76,77,79,80]. Neurodegenerative disorders affect more than one billion individuals, approximately fifteen percent of the global population, and at least seven million individuals die each year from neurodegenerative disorders [81]. Furthermore, the cost of neurodegenerative disorders is greater than USD 700 billion in the United States (USA) alone. This includes caring for disorders such as cognitive loss, stroke, back pain, epilepsy, trauma to the nervous system, and Parkinson’s disease (PD). Of note, dementia care is the most significant cost factor with more than USD 800 billion a year allotted for cognitive care [54]. These financial projections do not account for the additional expenses necessary to provide adult living care, social outreach programs, and companion care, since almost 60 million new health and social care workers will be needed [53,54,82].
Dementia also is considered to be markedly under-diagnosed and treatment may not begin until twelve to eighteen months after a proper assessment is made [39,83,84]. If one examines the sporadic cases of Alzheimer’s disease (AD), this disorder is expected to increase over time [6,64,81,85,86,87]. Per reports from the World Health Organization [54], dementia affects all countries in the world and it is now considered to be the seventh leading cause of death [41,84,88,89,90]. The sporadic form of AD represents most of the clinical cases for this disorder. It is estimated that at least ten percent of the global population over the age of 65 is affected by sporadic AD [6,47,73,86,87,91,92,93].
In regard to metabolic disorders such as DM, this disorder affects all systems of the body that involve the central nervous system, the peripheral nervous system, the inflammatory system, and the vascular system (Figure 1). In the central nervous system, DM can be a significant contributor to cognitive loss and dementia [6,27,39,47,94,95,96,97,98]. DM can result in insulin resistance and dementia that occurs in patients with AD [84,98,99]. DM can affect impact stem cell proliferation [7,32,33,34,100], cytoprotective pathways [22,29,98,101,102,103], retinal disease [38,104,105,106], and immune-mediated pathways that involve microglia [95,106,107,108,109,110,111]. Furthermore, over seventy percent of diabetic individuals can develop peripheral neuropathy. DM can lead to both autonomic neuropathy [112] and peripheral nerve disorders [39,113,114]. In the immune and vascular systems, DM can result in low-grade and acute inflammation [19,34,115,116,117], endothelial dysfunction [63,118,119], cardiovascular disease [7,24,25,30,31,120,121,122,123,124,125,126,127], and impairment of the neurovascular unit [3,22,41,94,97,101,119,128,129,130].
4. New Treatment Strategies to Address Cellular Metabolism Dysfunction
Metabolic disorders represent a significant consideration for the treatment of neurodegenerative disease. In some cases, if the disorder is recognized early in the course of the disease, neurodegenerative disability may be limited. Effective and timely treatment may inhibit both the progression of DM and diseases of the nervous system, such as cognitive loss [26,28,33,52,69,70,84,94,104,120,127,131,132,133,134,135,136,137,138,139]. Although the use of current pharmaceuticals and diet modification can assist in the treatment of DM and metabolic disorders to prevent hyperglycemic events, these strategies have potential risks that can decrease organ mass, affect cellular organelles, and lead to neuronal loss through processes that involve autophagy [104,124,140,141]. In regard to limiting cognitive loss, present therapies that may overlap with cellular metabolism pathways, such as the removal of ß-amyloid (Aβ) in the brain [92,142,143] and the use of cholinesterase inhibitors [89,92,144], may provide only symptomatic relief or slow disease progression over a limited period [83,89,145,146,147,148,149,150,151,152,153]. Additional factors that influence cognition involve low education in early life, high blood pressure, vascular disease, and tobacco use [83,84,154]. More recent studies also focus on the role of increased exercise programs for individuals that can assist with metabolic disease and neurodegenerative disorders that involve dementia and PD [8,11,12,13,41,52,155,156,157,158,159,160]. Progressive vascular disease during metabolic disorders also can lead to memory impairment and cognitive loss [55,151,155,161,162,163,164,165]. Given these challenges to overcome cognitive loss in the setting of metabolic disorders, new pathways of discovery are necessary to target novel pathways for disease onset and treatment. Innovative avenues that can address these requirements rely upon our understanding of metabolic homeostasis, programmed cell death pathways, the mechanistic target of rapamycin (mTOR) and its associated pathways of mTOR Complex 1 (mTORC1) and mTOR Complex 2 (mTORC2), AMP activated protein kinase (AMPK), growth factor signaling with erythropoietin (EPO), and underlying risk factors such as the apolipoprotein E (APOE-ε4) gene (Table 1).
5. Cellular Metabolism and the Role of Apoptosis, Autophagy, and Pyroptosis
Programmed cell death pathways during metabolic disorders can affect neuronal cell integrity and lead to the onset of multiple neurodegenerative disorders in the nervous system [94,128,130,149,166,167,168,169,170,171] (Figure 1). Programmed cell death, a biological process that can represent cellular suicide, can be a significant factor that can oversee the activation of inflammatory pathways during metabolic disorders, such as DM [7,19,39,104,119,126,127,135,172,173,174]. Included in these pathways of programmed cell death are apoptosis, autophagy, and pyroptosis [175,176,177,178,179,180].
In regard to apoptosis, this “cell-suicide” pathway has both an early and late phase [85]. The early phase consists of phosphatidylserine (PS) membrane asymmetry loss on the plasma membrane [181,182,183,184,185]. The later apoptotic phase results in deoxyribonucleic acid (DNA) degradation in the genome [186,187,188,189,190,191,192]. Loss of membrane PS asymmetry can lead inflammatory cells, such as microglia, to target, engulf, and remove injured cells [111,181,193,194]. This process is reversible if the engulfment of inflammatory cells can be stopped to allow the remaining functional cells expressing membrane PS residues to be rescued [38,195,196,197]. Apoptotic cell death is mediated through a cascade activation of nucleases and proteases that involve caspases [66,186,187,198,199,200,201,202,203]. As a result, once the destruction of cellular DNA occurs, this process may not be reversible [145]. The modulation of apoptotic pathways can minimize cognitive loss during acute insults [178,191,198,204,205,206]. Fostering anti-inflammatory pathways can improve cognitive performance and limit cellular apoptosis [91,200].
Autophagy processes can recycle cytoplasmic organelles and components for eventual tissue remodeling [37,69,86,87,175,177,207,208,209,210,211,212,213,214,215,216]. There are subtypes of autophagy processing. Macroautophagy recycles organelles and sequesters cytoplasmic proteins and organelles into autophagosomes that combine with lysosomes for degradation and recycling [87,217,218]. Microautophagy refers to the process of invagination of lysosomal membranes for the sequestration and digestion of cytoplasmic components [219]. Chaperone-mediated autophagy relies upon cytosolic chaperones to transport cytoplasmic components across lysosomal membranes [220,221,222].
Interestingly, the autophagy–lysosome pathway plays an important role in inflammatory injury during infections, such as with the pathogen severe acute respiratory syndrome coronavirus (SARS-CoV-2) [223,224,225,226,227,228,229,230,231]. Yet, processes involving autophagy hold a significant role in cognitive function [27,95,98,232,233,234,235]. The activation of autophagy can reduce tau and Aß neurotoxicity [236,237,238]. Autophagy can target Aß levels in the brain as one possible component to limit memory loss [98,146,236,239].
In relation to pyroptosis, this form of programmed cell death can affect inflammatory pathways in the nervous system during metabolic disease [3,39,145,191,240,241,242,243]. Pyroptosis begins with the generation of a supramolecular complex, termed the pyroptosome or the inflammasome, that can promote caspase activation to include caspase 1, caspase 4, and caspase 5. In addition, pyroptosis employs permeabilization of the plasma membrane through gasdermin protein family members. Gasdermin proteins contain both an N-terminal domain with intrinsic pore-forming properties and a C-terminal domain that can block the pore-forming properties of the N-terminal domain. Disruption of the linker sequence that binds the N-terminal and the C-terminal domains permits the N-terminal domain fragment to generate pores in the plasma. Cellular membranes are then able to release pro-inflammatory cytokines such as interleukin-1 family members. Inflammatory factors can control a balance in regard to assisting or hampering cell survival. This can be controlled by mechanisms that require gasdermin. For example, interleukin-1 family members lack a signal plasma membrane peptide that would normally allow their cellular release and therefore require gasdermin proteins to generate membrane pores [47,244]. This opening of cell membrane pores can result in the rupture of cell membranes, the release of cytokines, and other damage-associated molecular pattern (DAMP) molecules, which includes DNA and adenosine triphosphate (ATP). DAMPs can lead to the activation of the NLR family pyrin domain containing 3 (NLRP3) inflammasome in the canonical inflammasome pathways. The noncanonical inflammasome pathway is generated by binding of lipopolysaccharide proteins that can be found on Gram-negative bacteria leading to caspase 4 and caspase 5 activation. As a result, pyroptosis, in addition to apoptosis and necroptosis, may lead to pro-inflammatory responses that can cause cytokine storm and cell death [241]. These factors also play a role during DM and diabetic wound healing. Pro-inflammatory mediators, such as the NLRP3 inflammasome, can activate caspase 1 and cytokines, result in metabolic stress, and cause cell death and poor wound healing [242]. During periods of oxidative stress and reactive oxygen species (ROS) release, pyroptosis under inflammatory and toxic environments can have a significant role in affecting cognition, AD, and DM complications, which include neuronal and vascular disease [21,41,56,86,132,204,224,245].
6. A Central Role for the Mechanistic Target of Rapamycin (mTOR) in Cellular Metabolism
The mechanistic target of rapamycin (mTOR) is a 289-kDa serine/threonine protein kinase that is encoded by a single gene FRAP1 [6,81,87,212,246,247,248,249]. mTOR also is known as the mammalian target of rapamycin and the FK506-binding protein 12-rapamycin complex-associated protein 1 [222]. The target of rapamycin (TOR) was initially discovered in Saccharomyces cerevisiae with the genes TOR1 and TOR2 [249]. The agent rapamycin is a macrolide antibiotic in Streptomyces hygroscopicus that blocks TOR and mTOR activity [119].
mTOR is an integral component of the protein complexes mTOR Complex 1 (mTORC1) and mTOR Complex 2 (mTORC2) (Figure 1). mTORC1 contains Raptor, Deptor (DEP domain-containing mTOR interacting protein), the proline-rich Akt substrate 40 kDa (PRAS40), and mammalian lethal with Sec13 protein 8, termed mLST8 (mLST8) [6,81]. mTOR can control Raptor activity, which can be blocked by rapamycin. Rapamycin may block the activity of mTORC1 by binding to immunophilin FK-506-binding protein 12 (FKBP12) that normally attaches to the FKBP12 -rapamycin-binding domain (FRB) at the carboxy (C)-terminal of mTOR and blocks the FRB domain of mTORC1 [85]. However, the mechanism of how rapamycin blocks mTORC1 activity with the interaction of the domain of FRB is unclear. One consideration may involve allosteric changes in the catalytic domain as well as the inhibition of phosphorylation of protein kinase B (Akt) and p70 ribosomal S6 kinase (p70S6K) [250]. mTORC1 appears to be more sensitive to inhibition by rapamycin than mTORC2, but chronic administration of rapamycin can inhibit mTORC2 activity as a result of the disruption of the assembly of mTORC2. Deptor, also an inhibitor, blocks mTORC1 activity by binding to the FAT domain (FKBP12 -rapamycin-associated protein (FRAP), ataxia-telangiectasia (ATM), and the transactivation/transformation domain-associated protein) of mTOR. PRAS40 blocks mTORC1 activity by limiting the association of p70 ribosomal S6 kinase (p70S6K) and the eukaryotic initiation factor 4E (eIF4E)-binding protein 1 (4EBP1) with Raptor [6,85,127,246]. Akt is important in this pathway as a checkpoint since mTORC1 activity is increased once the phosphorylation of PRAS40 occurs by Akt [66,190,251,252]. This releases the binding of PRAS40 and Raptor to localize PRAS40 in the cell cytoplasm with the docking protein 14-3-3 [253,254,255]. mLST8 can promote the activity of mTOR [85]. This activation involves the binding of p70S6K and 4EBP1 to Raptor [256].
In relation to metabolic disease, mLST8 controls insulin signaling through the transcription factor FoxO3 [115,257], is necessary for Akt and protein kinase C-α (PKCα) phosphorylation, and is required for Rictor to associate with mTOR [257]. In addition, mTORC1 is associated with metabolic disorders [26,113,224,258] and dementia [27,39,163,259]. mTORC1 can promote lipogenesis and fat storage [260], improve glucose homeostasis [261], and may increase pancreatic ß-cell mass [262].
mTORC2 has differences from mTOC1 and is composed of Rictor, Deptor, the mammalian stress-activated protein kinase interacting protein (mSIN1), mLST8, and the protein observed with Rictor-1 (Protor-1) [6,85,208,263,264]. mTORC2 oversees cytoskeleton remodeling through PKCα and cell migration through the Rac guanine nucleotide exchange factors P-Rex1 and P-Rex2 and through Rho signaling. mTORC2 can foster the activity of protein kinases, including glucocorticoid induced protein kinase 1 (SGK1), a member of the protein kinase A/protein kinase G/protein kinase C (AGC) family of protein kinases. Protor-1, a Rictor-binding subunit of mTORC2, can result in SGK1 activity [265,266]. mSin1 is necessary for the assembly of mTORC2 and for mTORC2 to phosphorylate Akt [267]. Rictor and mSIN1 phosphorylate Akt at serine473 and promote threonine308 phosphorylation by phosphoinositide-dependent kinase 1 (PDK1) to increase cell survival. mTORC2 also has a role during cellular metabolism [26,47]. mTORC2 signaling is required for the maintenance of pancreatic β-cell proliferation and mass [268]. Loss of mTORC2 signaling results in insulin resistance, oxidative damage [269], and severe hyperglycemia [270].
7. Metabolic Cell Death and mTOR
During cellular metabolic dysfunction and neurodegenerative processes, mTOR has an intimate relationship with programmed cell death pathways (Table 1). The activation of mTOR can limit apoptotic cell death in the nervous system [271,272,273,274]. During mTOR activation, retinal ganglion cell regeneration can be fostered [275], microglia can be protected during oxidative stress exposure [276], and diabetic peripheral neuropathy can be reduced [277,278]. In addition, during the activation of mTOR, Aß toxicity can be blocked [205,237,254,279,280,281,282], vascular cell death is prevented [215,283], increased neuroplasticity can occur [284], loss of mitochondria during oxidative stress is blocked [285], neuronal differentiation can be fostered [286], neonatal central nervous system hypoxic injury is prevented [287], and reduced stroke volume with decreased apoptotic cell death dependent is promoted [198].
In some cases, the activation of autophagy with the inhibition of mTOR may be neuroprotective [37,47,61,86,87,205,207,235]. Cognitive function may be improved with low-calorie diets that promote autophagy and limit mTOR activity [288]. The activation of autophagy with decreased mTOR function may result in improved memory and more robust insulin signaling that can increase Aß clearance [289]. Additional work suggests that cognition may improve with tau clearance during autophagy activation and reduced mTOR function [238]. Autophagy activation in the setting of decreased mTOR activity also can prevent mitochondrial dysfunction [290], limit injury to dopamine-dependent cells [291], decrease reactive oxygen species release [292], and modulate neuroprotection with glutamine-dependent mechanisms [293]. During DM, mTOR inhibition can offer protection such as during cerebral ischemia [294] and is necessary for maintaining a balance between pancreatic β-cell proliferation and cell size [268]. Additional studies indicate that the dysregulation of autophagy as a central pathway can result in the progression of cognitive loss with AD and the induction of DM [99]. Studies also have shown that autophagy haploinsufficiency with deletion of the Atg7 gene in mouse models of obesity leads to increased insulin resistance with elevated lipids and inflammation [295]. A loss of autophagic proteins Atg7, Atg5, and LC3 also can be responsible for diabetic nephropathy [296]. Autophagy offers protection by removing misfolded proteins and eliminating non-functioning mitochondria to maintain β-cell function and prevent the onset of DM [297]. Exercise in mice also has been shown to foster autophagy and regulate glucose homeostasis [298], possibly through improved insulin sensitivity [299] and supporting microglial function during acute glucose fluctuations [95].
It is important to note though that the activity of mTOR may be required to some degree, since studies suggest that dysfunction in mTOR signaling can result in cognitive impairment and synaptic dysfunction [64,85,87,235,300,301]. Autophagy activation can injure endothelial progenitor cells, lead to mitochondrial oxidative stress, and prevent new blood vessel formation during elevated glucose exposure [302]. At times, autophagy can be a component that leads to neuronal cell death [239]. Increased activity of autophagy can result in a significant loss of cardiac and liver tissue in diabetic rats during attempts to achieve glycemic control through diet modification [140]. During periods of elevated glucose, advanced glycation end products (AGEs), agents that can result in complications during DM, have been shown to lead to the induction of autophagy and vascular smooth muscle proliferation that can result in atherosclerosis [303] as well as cardiomyopathy [304]. Autophagy during elevated glucose exposure can lead to mitochondrial oxidative stress [305], impair endothelial progenitor cells, and block angiogenesis [302]. Growth factors that promote mTOR activation and limit autophagy can offer neuronal and vascular protection in the nervous system, such as during application with EPO [186,252,306]. EPO can modulate several mTOR pathways, such as PRAS40 and Akt, to lead to increased neuronal survival [253,307,308,309]. The activity of mTOR also is necessary for interneuron progenitor growth in the brain during autophagy inhibition [310].
In addition to its role in the nervous system, mTOR is necessary for the proper function of several metabolic pathways [6,26,27,37,113,117,127,130,226,311,312]. mTOR activation with the pathways of p70S6K and 4EBP1 can promote insulin secretion in pancreatic β-cells and increase resistance to β-cell streptozotocin toxicity and obesity in mice [262]. In contrast, the loss of p70S6K activity results in hypo-insulinemia, insulin insensitivity to glucose secretion, glucose intolerance, and decreased pancreatic β-cell size [313]. mTOR activation in patients with metabolic syndrome has also been found to be diminished and possibly responsible for insulin resistance with an increased risk of vascular thrombosis [314]. The activation of mTOR pathways has been tied to pancreatic β- cell protection against cholesterol-induced apoptosis [315], increased neuronal cell survival in cell models of DM [316], and decreased glucolipotoxicity [317]. Yet, mTOR inhibition leads to reduced β-cell function, insulin resistance, and limited insulin secretion associated with the progression of DM [318]. Loss of mTOR activity can also increase mortality in a mouse model of DM [319]. Furthermore, the translocation of glucose transporters to the plasma membrane in skeletal muscle is impacted during mTOR inhibition [320].
mTOR can, under some scenarios, control cellular metabolism to preserve cognition. mTOR activity can oversee insulin signaling in experimental models of AD and maintain astrocyte viability [321], prevent endothelial cell dysfunction during periods of hyperglycemia [322], and maintain glucose homeostasis [261]. mTOR may offer additional protection as a component of the Mediterranean diet and nutrition. mTOR may reduce Aβ toxicity in astrocytes via enhanced Akt activity through the consumption of polyphenols in olives and olive oil, which may be linked to the onset of AD [321].
8. Metabolic Cell Death and AMP Activated Protein Kinase (AMPK)
The AMP activated protein kinase (AMPK) plays an important role in both metabolic and neurodegenerative disorders as well as in other disease entities that involve inflammation and infection [8,26,86,177,264,323,324,325] (Figure 1). AMPK controls mTORC1 activity through the hamartin (tuberous sclerosis 1)/tuberin (tuberous sclerosis 2) (TSC1/TSC2) complex that inhibits mTORC1 function [325,326]. Modulation of the TSC1/TSC2 complex also can be overseen though phosphoinositide 3-kinase (PI 3-K), Akt, and its phosphorylation of TSC2. Extracellular signal-regulated kinases (ERKs), protein p90 ribosomal S6 kinase 1 (RSK1), and glycogen synthase kinase -3β (GSK-3β) can oversee the activity of the TSC1/TSC2 complex as well. TSC2 functions as a GTPase-activating protein (GAP) that changes G protein Rheb (Rheb-GTP) into the inactive GDP-bound form (Rheb-GDP). During Rheb-GTP activation, Rheb-GTP associates with Raptor to oversee the binding of 4EBP1 to mTORC1 and increase mTORC1 activity [327]. AMPK phosphorylates TSC2 to increase GAP activity to change Rheb-GTP into the inactive Rheb-GDP and to block mTORC1 activity [328].
As a pathway of mTOR, AMPK is a critical modulator of mTOR pathways and can oversee cellular metabolism and the pathways of programmed cell death. AMPK has been shown to affect insulin resistance and mitochondrial homeostasis [23,39,329,330]. During dietary restrictions that may increase lifespan [331], AMPK can alter cellular metabolism to shift to protective oxidative metabolism [332]. In relation to stem cell maintenance, AMPK can be necessary for resistance to senescence in mesenchymal stem cells [333] and can protect endothelial progenitor cells during periods of hyperglycemia [334]. Anti-senescence cell activity can be promoted through mTOR inhibition, AMPK activation, and the acceleration of autophagic flux [335]. In regard to neurodegeneration, AMPK activation can improve memory retention in models of AD and DM [336], assist with pathways for healthy aging [58], facilitate clearance of Aß [337] and tau [238] in the brain, diminish Aß neurotoxicity [338], and control chronic inflammation in neurodegenerative disorders [61,81,326].
AMPK employs autophagy activation to control cell survival and regulate metabolic homeostasis [6,23,41,47,113,322]. Modulation of AMPK activity may be required to increase basal autophagy activity [175,339] and maintain endothelial cell survival [322,340] during hyperglycemia. AMPK can control both apoptosis and autophagy during coronary artery disease [341] and oxidative stress cell injury [342,343]. In regard to growth factor protection in the nervous system, EPO can increase neuronal survival through increased AMPK activity and enhanced autophagy activity [344]. EPO oversees AMPK and mTOR activities to protect cells under conditions of oxidative stress [281] and inflammation [306,345,346]. The duration and concentration of EPO exposure can influence a specific level of AMPK and mTOR activity to alleviate the detrimental effects of oxidative stress [253,347]. This fine control over mTOR is important since high concentrations of EPO may result in cell death and reduce the activity of mTOR [348].
Interestingly, AMPK pathways are used by current agents to treat DM, such as metformin and biguanides, to limit neurodegenerative disease, such as peripheral neuropathy, demyelinating disease, and cognitive loss [27,224,329,349,350]. Metformin inhibits mTOR activity, promotes autophagy, and may function at times in an AMPK-independent manner [351]. Metformin can limit lipid peroxidation in the brain and spinal cord and decrease caspase activity to enhance cell survival [352]. Recent work also suggests that metformin through AMPK pathways can accelerate myelin recovery in animal models of multiple sclerosis [349]. In relation to infections of the nervous system that can lead to neurodegeneration, metformin reduced disability in obese patients or individuals with DM during coronavirus disease 2019 (COVID-19) [14,353].
9. Cellular Metabolism and the ε4 Allele of the Apolipoprotein E (APOE-ε4)
Since the sporadic form of AD represents most of the clinical cases, it can impact at least ten percent of the global population over the age of 65 [6,47,73,86,87,91,92,93]. Although multiple factors and cellular pathways may precipitate AD, individuals with the ε4 allele of the apolipoprotein E (APOE-ε4) gene have an increased risk of late-onset AD [215,237,354,355,356] (Figure 1). Apolipoprotein E (APOE) is produced in the liver and is important for cellular metabolism by regulating lipid homeostasis and the transport of cholesterol, triglycerides, and phospholipids in the body [96,147]. In the brain, APOE is generated in astrocytes and is used through APOE receptors to transfer cholesterol to neurons [99,215]. In some cases, APOE can assist with the destruction of Aβ in the brain (Table 1). However, the isoform APOE-ε4 is not effective in the destruction of Aβ, which may result in heightened risk of the development of AD [99,237,354,357,358,359]. Individuals with two ε4 alleles may have almost 20 times the risk of developing AD. Interestingly, PS membrane exposure [360,361,362,363], an initial phase of which is apoptotic cell death, may be related to Aβ aggregation and some isoforms of APOE may prevent the aggregation of Aβ through PS membrane exposure, but this is not the case for the isoform APOE-ε4 [358].
APOE-ε4 has been shown to affect mTOR signaling and increase mTOR activity [364,365]. The ability of APOE-ε4 to alter mTOR activity as well as autophagy flux has been suggested to increase the risk for the development of cerebrovascular disease and AD due to possible deficits in synaptic plasticity [366]. In addition, studies have suggested that at least twenty-two viral diseases have been identified to cause increased risk of neurodegenerative disorders, with the most prominent presenting disease being dementia [367]. In particular, APOE-ε4 may promote the susceptibility of viral infection and cerebrovascular disease during COVID-19 [368]. The β-coronavirus family virion, severe acute respiratory syndrome (SARS) -CoV-2 (SARS-CoV-2), has led to multiple and repeated infections throughout the global population [8,27,223,224,225,226,227,228,312,350,369,370,371,372]. Coronaviruses are ribonucleic acid (RNA) viruses and are members of the family of Coronaviridae and the subfamily of Orthocoronavirinae [27,241,373,374,375,376,377]. SARS-CoV-2 can attach to host cells, such as in the nasal epithelial region [378] and the brain [117], and lead to a heightened response of the immune system [10,375,379,380,381,382].
Cognitive disability can ensue after infection with SARS-CoV-2 [27,224,311,377,383,384]. The cognitive loss can be part of a long-COVID syndrome [227]. Long-COVID, also known as long-haul COVID, post-acute COVID-19, and chronic COVID, represents long-term effects that can occur following acute SARS-CoV-2 infection. Multiple mechanisms can account for this chronic syndrome, including metabolic pathways, apoptosis, autophagy, oxidative stress, mitochondrial dysfunction, and cytokine release [56,73,74,86,87,88,91,177,178,180,206,212,251,385,386,387,388,389,390,391,392,393]. With the knowledge of the role of APOE-ε4 in metabolism and autophagy signaling, APOE-ε4 has been linked to the effects of long COVID and dementia. Individuals with two ε4 alleles of APOE-ε4 have reduced expression of antiviral defense genes and have more severe neuroinflammation and microvascular injury in the brain [151,355,394]. APOE-ε4 during SARS-CoV-2 infection and long COVID may result in cognitive loss and cerebrovascular disease in the nervous system [11,30,61,66,85,167,177,199,395,396].
10. Future Perspectives
Metabolic disease, which includes DM, represents a significant healthcare concern for the global population (Table 1). The prevalence of DM continues to increase and by the year 2045 seven hundred million individuals are expected to suffer from DM according to the International Diabetes Federation [1]. Of further concern, it is estimated that approximately seven million individuals over the age of eighteen are undiagnosed with DM. Although the use of current pharmaceuticals and diet modification can assist in the treatment of DM and metabolic disorders, these treatments cannot reverse disease progression and have potential risks that can decrease organ mass, affect cellular organelles, and lead to neuronal loss through processes that involve autophagy. Of additional concern is the role of metabolic disorders in fostering neurodegeneration, especially since DM can be a critical factor for the development of cognitive loss and dementia. Coupled to these concerns for the onset of neurodegenerative disease as a result of underlying metabolic disorders is the limited treatment arsenal for disorders such as cognitive loss, now considered to be the seventh leading cause of death. Overcoming these challenges requires the implementation of novel and innovative strategies. The focus upon metabolic homeostasis, programmed cell death pathways, mTOR and its associated pathways of mTORC1, mTORC2, AMPK, growth factor signaling with EPO, and underlying risk factors such as the APOE-ε4 gene can offer new directions to identify potential therapeutic strategies for metabolic disorders and neurodegenerative disease (Figure 1).
The examination of these pathways, each intimately linked to one another, offers a number of valuable attributes. For example, mTOR activation can lead to retinal ganglion cell regeneration, a reduction in diabetic peripheral neuropathy, and limit Aß toxicity in the nervous system. Trophic factors, such as EPO, can offer cellular protection against neurotoxic insults by also fostering the activation of mTOR. Components of the mTOR pathway, such as mLST8, can oversee insulin signaling to help maintain glucose homeostasis. mTORC1 can promote lipogenesis and fat storage, improve glucose homeostasis, and increase pancreatic ß-cell mass, and mTORC2 signaling is necessary for the maintenance of pancreatic β-cell proliferation and mass. However, under some conditions, the activation of autophagy with the inhibition of mTOR may be necessary to prevent neurodegeneration. As an example, the activation of autophagy with blocked mTOR function may result in improved memory and more robust insulin signaling that can increase Aß clearance in the nervous system. The dysregulation of autophagy also may result in the progression of cognitive loss with AD and DM. AMPK activation during the inhibition of mTOR leads to memory retention, limits lipid accumulation and obesity, and results in neuroprotection. However, under alternate cellular conditions, AMPK inhibition with active mTOR signaling is necessary to offer protection to pancreatic islet cells, limit Aβ toxicity, and block nervous system inflammation.
11. Conclusions
A fine balance between glucose homeostasis, programmed cell death pathways, and mTOR signaling pathways is required. As a result, future studies are warranted to pursue such directions. For example, biological feedback pathways with mTOR, such as through AMPK inhibition, are believed to exist to prevent excessive mTOR activity. Periods of elevated mTOR activity can be detrimental and result in glucose intolerance by inhibiting the insulin receptor substrate 1 (IRS-1) [397]. In addition, the presence of growth factors can affect this balance, since the duration and concentration of EPO exposure can affect AMPK and mTOR activity to offer cellular protection with controlled EPO concentrations but limit the activity of mTOR and lead to cell death during elevated EPO concentrations.
Additional investigations must also seek to understand the underlying risk factors for neurodegenerative disease that can occur as a result of metabolic disorders. The role of APOE-ε4 as a significant risk factor for developing AD and the function of APOE-ε4 to regulate lipid homeostasis and the transport of cholesterol, triglycerides, and phospholipids in the body offers critical support for this conjecture. Aβ aggregation in the brain appears to be linked to the inability of APOE-ε4 to oversee apoptotic signaling with PS membrane exposure. Furthermore, APOE-ε4 has been shown to affect mTOR signaling, increase mTOR activity, and alter autophagy flux that can increase the risk for the development of cerebrovascular disease and AD. In addition, APOE-ε4 also may increase the susceptibility of viral infection and cerebrovascular disease during COVID-19 as well as promote long-term disability with dementia and long-COVID syndrome. Individuals with two ε4 alleles of APOE-ε4 have limited expression of antiviral defense genes and, as a result, have more severe neuroinflammation and microvascular injury in the brain. The future offers new challenges to unravel the complex and critical pathways that tie neurodegenerative disorders to metabolic disease and develop new therapeutic avenues for these diseases that impact the global population.
Funding
This research was supported by the following grants to Kenneth Maiese: American Diabetes Association, American Heart Association, NIH NIEHS, NIH NIA, NIH NINDS, NS053956, and NIH ARRA.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- International Diabetes Federation. Diabetes. In IDF Diabetes Atlas; International Diabetes Federation: Brussels, Belgium, 2019. [Google Scholar]
- Fadini, G.P.; Morieri, M.L.; Longato, E.; Avogaro, A. Prevalence and impact of diabetes among people infected with SARS-CoV-2. J. Endocrinol. Investig. 2020, 43, 867–869. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Prospects and Perspectives for WISP1 (CCN4) in Diabetes Mellitus. Curr. Neurovasc. Res. 2020, 17, 327–331. [Google Scholar] [CrossRef] [PubMed]
- Nie, X.; Wei, X.; Ma, H.; Fan, L.; Chen, W.D. The complex role of Wnt ligands in type 2 diabetes mellitus and related complications. J. Cell. Mol. Med. 2021, 25, 6475–6495. [Google Scholar] [CrossRef] [PubMed]
- Schell, M.; Wardelmann, K.; Kleinridders, A. Untangling the effect of insulin action on brain mitochondria and metabolism. J. Neuroendocr. 2021, 33, e12932. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Cognitive impairment with diabetes mellitus and metabolic disease: Innovative insights with the mechanistic target of rapamycin and circadian clock gene pathways. Expert Rev. Clin. Pharm. 2020, 13, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. SIRT1 and stem cells: In the forefront with cardiovascular disease, neurodegeneration and cancer. World J. Stem Cells 2015, 7, 235–242. [Google Scholar] [CrossRef]
- Alves, H.R.; Lomba, G.S.B.; Gonçalves-de-Albuquerque, C.F.; Burth, P. Irisin, Exercise, and COVID-19. Front. Endocrinol. 2022, 13, 879066. [Google Scholar] [CrossRef]
- Chong, M.C.; Silva, A.; James, P.F.; Wu, S.S.X.; Howitt, J. Exercise increases the release of NAMPT in extracellular vesicles and alters NAD(+) activity in recipient cells. Aging Cell 2022, 21, e13647. [Google Scholar] [CrossRef]
- Furtado, G.E.; Letieri, R.V.; Caldo-Silva, A.; Sardão, V.A.; Teixeira, A.M.; de Barros, M.P.; Vieira, R.P.; Bachi, A.L.L. Sustaining efficient immune functions with regular physical exercise in the COVID-19 era and beyond. Eur. J. Clin. Investig. 2021, 51, e13485. [Google Scholar] [CrossRef]
- Maiese, K. Picking a bone with WISP1 (CCN4): New strategies against degenerative joint disease. J. Transl. Sci. 2016, 1, 83–85. [Google Scholar] [CrossRef]
- Potthast, A.B.; Nebl, J.; Wasserfurth, P.; Haufe, S.; Eigendorf, J.; Hahn, A.; Das, A. Impact of Nutrition on Short-Term Exercise-Induced Sirtuin Regulation: Vegans Differ from Omnivores and Lacto-Ovo Vegetarians. Nutrients 2020, 12, 1004. [Google Scholar] [CrossRef]
- Zeng, Z.; Liang, J.; Wu, L.; Zhang, H.; Lv, J.; Chen, N. Exercise-Induced Autophagy Suppresses Sarcopenia Through Akt/mTOR and Akt/FoxO3a Signal Pathways and AMPK-Mediated Mitochondrial Quality Control. Front. Physiol. 2020, 11, 583478. [Google Scholar] [CrossRef]
- Bramante, C.; Ingraham, N.; Murray, T.; Marmor, S.; Hoversten, S.; Gronski, J.; McNeil, C.; Feng, R.; Guzman, G.; Abdelwahab, N.; et al. Observational Study of Metformin and Risk of Mortality in Patients Hospitalized with COVID-19. medRxiv 2020. [Google Scholar] [CrossRef]
- Hołowko-Ziółek, J.; Cięszczyk, P.; Biliński, J.; Basak, G.W.; Stachowska, E. What Model of Nutrition Can Be Recommended to People Ending Their Professional Sports Career? An Analysis of the Mediterranean Diet and the CRON Diet in the Context of Former Athletes. Nutrients 2020, 12, 3604. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hu, J.; Yang, L.; Wang, N.; Liu, Y.; Wei, X.; Gao, M.; Wang, Y.; Ma, Y.; Wen, D. Association of WISP1/CCN4 with Risk of Overweight and Gestational Diabetes Mellitus in Chinese Pregnant Women. Dis. Markers 2020, 2020, 4934206. [Google Scholar] [CrossRef]
- Lu, M.; Chen, C.; Lan, Y.; Xiao, J.; Li, R.; Huang, J.; Huang, Q.; Cao, Y.; Ho, C.T. Capsaicin-the major bioactive ingredient of chili peppers: Bio-efficacy and delivery systems. Food Funct. 2020, 11, 2848–2860. [Google Scholar] [CrossRef]
- Maiese, K. Paring down obesity and metabolic disease by targeting inflammation and oxidative stress. Curr. Neurovasc. Res. 2015, 12, 107–108. [Google Scholar] [CrossRef]
- Maiese, K. Erythropoietin and diabetes mellitus. World J. Diabetes 2015, 6, 1259–1273. [Google Scholar] [CrossRef] [PubMed]
- Quesada, I.; de Paola, M.; Torres-Palazzolo, C.; Camargo, A.; Ferder, L.; Manucha, W.; Castro, C. Effect of Garlic’s Active Constituents in Inflammation, Obesity and Cardiovascular Disease. Curr. Hypertens. Rep. 2020, 22, 6. [Google Scholar] [CrossRef]
- Raut, S.K.; Khullar, M. Oxidative stress in metabolic diseases: Current scenario and therapeutic relevance. Mol. Cell. Biochem. 2023, 478, 185–196. [Google Scholar] [CrossRef]
- Yamashima, T.; Ota, T.; Mizukoshi, E.; Nakamura, H.; Yamamoto, Y.; Kikuchi, M.; Yamashita, T.; Kaneko, S. Intake of ω-6 Polyunsaturated Fatty Acid-Rich Vegetable Oils and Risk of Lifestyle Diseases. Adv. Nutr. 2020, 11, 1489–1509. [Google Scholar] [CrossRef]
- Yang, J.; Suo, H.; Song, J. Protective role of mitoquinone against impaired mitochondrial homeostasis in metabolic syndrome. Crit. Rev. Food Sci. Nutr. 2020, 20, 1–19. [Google Scholar] [CrossRef]
- Zaiou, M. circRNAs Signature as Potential Diagnostic and Prognostic Biomarker for Diabetes Mellitus and Related Cardiovascular Complications. Cells 2020, 9, 659. [Google Scholar] [CrossRef] [PubMed]
- Beegum, F.; PV, A.; George, K.T.; KP, D.; Begum, F.; Krishnadas, N.; Shenoy, R.R. Sirtuins as therapeutic targets for improving delayed wound healing in diabetes. J. Drug Target. 2022, 30, 911–926. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. New Insights for nicotinamide: Metabolic disease, autophagy, and mTOR. Front. Biosci. (Landmark Ed.) 2020, 25, 1925–1973. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Nicotinamide: Oversight of Metabolic Dysfunction through SIRT1, mTOR, and Clock Genes. Curr. Neurovasc. Res. 2020, 17, 765–783. [Google Scholar] [CrossRef] [PubMed]
- Fischer, F.; Grigolon, G.; Benner, C.; Ristow, M. Evolutionarily conserved transcription factors as regulators of longevity and targets for geroprotection. Physiol. Rev. 2022, 102, 1449–1494. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Chong, Z.Z.; Wang, S.; Shang, Y.C. Oxidant Stress and Signal Transduction in the Nervous System with the PI 3-K, Akt, and mTOR Cascade. Int. J. Mol. Sci. 2013, 13, 13830–13866. [Google Scholar] [CrossRef]
- Maiese, K.; Li, F.; Chong, Z.Z.; Shang, Y.C. The Wnt signaling pathway: Aging gracefully as a protectionist? Pharmacol. Ther. 2008, 118, 58–81. [Google Scholar] [CrossRef]
- Rotllan, N.; Camacho, M.; Tondo, M.; Diarte-Añazco, E.M.G.; Canyelles, M.; Méndez-Lara, K.A.; Benitez, S.; Alonso, N.; Mauricio, D.; Escolà-Gil, J.C.; et al. Therapeutic Potential of Emerging NAD+-Increasing Strategies for Cardiovascular Diseases. Antioxidants 2021, 10, 1939. [Google Scholar] [CrossRef]
- Maiese, K. New Insights for Oxidative Stress and Diabetes Mellitus. Oxid. Med. Cell. Longev. 2015, 2015, 875961. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, B.T.; Monjure, T.A.; Al-Ghadban, S.; Ives, C.J.; L’Ecuyer, M.P.; Rhee, C.; Romero-Lopez, M.; Li, Z.; Goodman, S.B.; Lin, H.; et al. Aberrant Expression of COX-2 and FOXG1 in Infrapatellar Fat Pad-Derived ASCs from Pre-Diabetic Donors. Cells 2022, 11, 2367. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.Y.; Yu, T.Y.; Jiang, F.X.; Wang, W. Functional maturation of immature β cells: A roadblock for stem cell therapy for type 1 diabetes. World J. Stem Cells 2021, 13, 193–207. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Global Report on Diabetes; World Health Organization: Geneva, Switzerland, 2016; pp. 1–83.
- Ajiboye, B.O.; Shonibare, M.T.; Oyinloye, B.E. Antidiabetic activity of watermelon (Citrullus lanatus) juice in alloxan-induced diabetic rats. J. Diabetes Metab. Disord. 2020, 19, 343–352. [Google Scholar] [CrossRef]
- Burillo, J.; Marqués, P.; Jiménez, B.; González-Blanco, C.; Benito, M.; Guillén, C. Insulin Resistance and Diabetes Mellitus in Alzheimer’s Disease. Cells 2021, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Novel applications of trophic factors, Wnt and WISP for neuronal repair and regeneration in metabolic disease. Neural Regen. Res. 2015, 10, 518–528. [Google Scholar] [CrossRef]
- Maiese, K. Novel nervous and multi-system regenerative therapeutic strategies for diabetes mellitus with mTOR. Neural Regen. Res. 2016, 11, 372–385. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. National Diabetes Statistics Report, 2020; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2020; CS 314227-A; pp. 1–30.
- Maiese, K. Nicotinamide as a Foundation for Treating Neurodegenerative Disease and Metabolic Disorders. Curr. Neurovasc. Res. 2021, 18, 134–149. [Google Scholar] [CrossRef]
- Elnour, M.A.A.; Saleh, A.A.; Kalantan, M.M.; Mirghani, H.O. The relationship between coffee intake, obstructive sleep apnea risk, and type 2 diabetes glycemic control, in Tabuk City, The Kingdom of Saudi Arabia: A case-control study. BMC Res. Notes 2019, 12, 798. [Google Scholar] [CrossRef]
- Hill, J.H.; Solt, C.; Foster, M.T. Obesity associated disease risk: The role of inherent differences and location of adipose depots. Horm. Mol. Biol. Clin. Investig. 2018, 33, 20180012. [Google Scholar] [CrossRef]
- Liu, C.; Zhong, C.; Chen, R.; Zhou, X.; Wu, J.; Han, J.; Li, X.; Zhang, Y.; Gao, Q.; Xiao, M.; et al. Higher dietary vitamin C intake is associated with a lower risk of gestational diabetes mellitus: A longitudinal cohort study. Clin. Nutr. 2020, 39, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Orkaby, A.R.; Dushkes, R.; Ward, R.; Djousse, L.; Buring, J.E.; Lee, I.M.; Cook, N.R.; LeBoff, M.S.; Okereke, O.I.; Copeland, T.; et al. Effect of Vitamin D3 and Omega-3 Fatty Acid Supplementation on Risk of Frailty: An Ancillary Study of a Randomized Clinical Trial. JAMA Netw. Open 2022, 5, e2231206. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.; Jiang, W.; Zhou, L. Islet Autoantibodies in the Patients with Sjogren’s Syndrome and Thyroid Disease and Risk of Progression to Latent Autoimmune Diabetes in Adults: A Case Series. Diabetes Metab. Syndr. Obes. 2021, 14, 1025–1033. [Google Scholar] [CrossRef]
- Maiese, K. Dysregulation of metabolic flexibility: The impact of mTOR on autophagy in neurodegenerative disease. Int. Rev. Neurobiol. 2020, 155, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Heightened Attention for Wnt Signaling in Diabetes Mellitus. Curr. Neurovasc. Res. 2020, 17, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Chong, Z.Z.; Maiese, K. Mammalian Target of Rapamycin Signaling in Diabetic Cardiovascular Disease. Cardiovasc. Diabetol. 2012, 11, 45. [Google Scholar] [CrossRef]
- Centers for Medicare and Medicaid Services. National Health Expenditure Projections 2018–2027. 2019. Available online: www.cms.gov (accessed on 1 May 2023.).
- Maiese, K. Targeting the core of neurodegeneration: FoxO, mTOR, and SIRT1. Neural Regen. Res. 2021, 16, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Speer, H.; D’Cunha, N.M.; Alexopoulos, N.I.; McKune, A.J.; Naumovski, N. Anthocyanins and Human Health-A Focus on Oxidative Stress, Inflammation and Disease. Antioxidants 2020, 9, 366. [Google Scholar] [CrossRef]
- World Health Organization. Description of the Global Burden of NCDs, Their Risk Factors and Determinants. In Global Status Report on Noncommunicable Diseases 2010; World Health Organization: Geneva, Switzerland, 2011; pp. 1–176. [Google Scholar]
- World Health Organization. Global Action Plan on the Public Health Response to Dementia 2017–2025; World Health Organization: Geneva, Switzerland, 2017; pp. 1–44.
- Maiese, K. Sirtuins: Developing Innovative Treatments for Aged-Related Memory Loss and Alzheimer’s Disease. Curr. Neurovasc. Res. 2018, 15, 367–371. [Google Scholar] [CrossRef]
- Jalgaonkar, M.P.; Parmar, U.M.; Kulkarni, Y.A.; Oza, M.J. SIRT1-FOXOs activity regulates diabetic complications. Pharm. Res. 2022, 175, 106014. [Google Scholar] [CrossRef]
- Odnokoz, O.; Nakatsuka, K.; Wright, C.; Castellanos, J.; Klichko, V.I.; Kretzschmar, D.; Orr, W.C.; Radyuk, S.N. Mitochondrial Redox Signaling Is Critical to the Normal Functioning of the Neuronal System. Front. Cell Dev. Biol. 2021, 9, 613036. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, H.; Wang, B.; Zhang, Y.; Zheng, X.; Shao, B.; Zhuge, Q.; Jin, K. Key Signaling Pathways in Aging and Potential Interventions for Healthy Aging. Cells 2021, 10, 660. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Cutting through the Complexities of mTOR for the Treatment of Stroke. Curr. Neurovasc. Res. 2014, 11, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Minino, A.M. Death in the United States, 2011; NCHS Data Brief: Hyattsville, MD, USA, 2013; pp. 1–8. [Google Scholar]
- Maiese, K. Moving to the Rhythm with Clock (Circadian) Genes, Autophagy, mTOR, and SIRT1 in Degenerative Disease and Cancer. Curr. Neurovasc. Res. 2017, 14, 299–304. [Google Scholar] [CrossRef]
- Chen, Y.L.; Hsieh, C.C.; Chu, P.M.; Chen, J.Y.; Huang, Y.C.; Chen, C.Y. Roles of protein tyrosine phosphatases in hepatocellular carcinoma progression (Review). Oncol. Rep. 2023, 49, 48. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Ding, K.; Yue, R.; Lei, M. Therapeutic effects of icariin and icariside II on diabetes mellitus and its complications. Crit. Rev. Food Sci. Nutr. 2023, 63, 1–26. [Google Scholar] [CrossRef]
- Li, J.B.; Hu, X.Y.; Chen, M.W.; Xiong, C.H.; Zhao, N.; Ge, Y.H.; Wang, H.; Gao, X.L.; Xu, N.J.; Zhao, L.X.; et al. p85S6K sustains synaptic GluA1 to ameliorate cognitive deficits in Alzheimer’s disease. Transl. Neurodegener. 2023, 12, 1. [Google Scholar] [CrossRef]
- Kahmini, F.R.; Ghaleh, H.D.; Shahgaldi, S. Sirtuins: Subtle Regulators Involved in Convoluted Mechanisms of Pregnancy. Cell. Physiol. Biochem. 2022, 56, 644–662. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, M.; Tian, J.; Gao, M.; Liu, M.; Fu, X.; Jin, T.; Pan, J.; Chen, F.; An, F. WNT1-inducible signalling pathway protein 1 stabilizes atherosclerotic plaques in apolipoprotein-E-deficient mice via the focal adhesion kinase/mitogen-activated extracellular signal-regulated kinase/extracellular signal-regulated kinase pathway. J. Hypertens. 2022, 40, 1666–1681. [Google Scholar] [CrossRef]
- Maiese, K. Wnt Signaling and WISP1 (CCN4): Critical Components in Neurovascular Disease, Blood Brain Barrier Regulation, and Cerebral Hemorrhage. Curr. Neurovasc. Res. 2022, 19, 379–382. [Google Scholar] [CrossRef]
- Hacioglu, C.; Kar, F.; Kanbak, G. Reproductive Effects of Nicotinamide on Testicular Function and Structure in Old Male Rats: Oxidative, Apoptotic, Hormonal, and Morphological Analyses. Reprod. Sci. 2021, 28, 3352–3360. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Cognitive Impairment and Dementia: Gaining Insight through Circadian Clock Gene Pathways. Biomolecules 2021, 11, 1002. [Google Scholar] [CrossRef]
- Maiese, K. Neurodegeneration, memory loss, and dementia: The impact of biological clocks and circadian rhythm. Front. Biosci. (Landmark Ed.) 2021, 26, 614–627. [Google Scholar] [CrossRef]
- Patocka, J.; Kuca, K.; Oleksak, P.; Nepovimova, E.; Valis, M.; Novotny, M.; Klimova, B. Rapamycin: Drug Repurposing in SARS-CoV-2 Infection. Pharmaceuticals 2021, 14, 217. [Google Scholar] [CrossRef] [PubMed]
- Sorrells, S.F.; Paredes, M.F.; Zhang, Z.; Kang, G.; Pastor-Alonso, O.; Biagiotti, S.; Page, C.E.; Sandoval, K.; Knox, A.; Connolly, A.; et al. Positive Controls in Adults and Children Support That Very Few, If Any, New Neurons Are Born in the Adult Human Hippocampus. J. Neurosci. 2021, 41, 2554–2565. [Google Scholar] [CrossRef] [PubMed]
- Amanollahi, M.; Jameie, M.; Heidari, A.; Rezaei, N. The Dialogue Between Neuroinflammation and Adult Neurogenesis: Mechanisms Involved and Alterations in Neurological Diseases. Mol. Neurobiol. 2022, 60, 923–959. [Google Scholar] [CrossRef]
- Mishra, P.; Davies, D.A.; Albensi, B.C. The Interaction Between NF-κB and Estrogen in Alzheimer’s Disease. Mol. Neurobiol. 2022, 60, 1515–1526. [Google Scholar] [CrossRef]
- Salemi, M.; Mogavero, M.P.; Lanza, G.; Mongioì, L.M.; Calogero, A.E.; Ferri, R. Examples of Inverse Comorbidity between Cancer and Neurodegenerative Diseases: A Possible Role for Noncoding RNA. Cells 2022, 11, 1930. [Google Scholar] [CrossRef]
- Savu, D.I.; Moisoi, N. Mitochondria—Nucleus communication in neurodegenerative disease. Who talks first, who talks louder? Biochim. Biophys. Acta Bioenerg. 2022, 1863, 148588. [Google Scholar] [CrossRef]
- Yalçin, M.; Mundorf, A.; Thiel, F.; Amatriain-Fernández, S.; Kalthoff, I.S.; Beucke, J.C.; Budde, H.; Garthus-Niegel, S.; Peterburs, J.; Relógio, A. It’s About Time: The Circadian Network as Time-Keeper for Cognitive Functioning, Locomotor Activity and Mental Health. Front. Physiol. 2022, 13, 873237. [Google Scholar] [CrossRef]
- Maiese, K. Novel Treatment Strategies for Neurodegenerative Disease with Sirtuins. In Sirtuin Biology in Medicine: Targeting New Avenues of Care in Development, Aging, and Disease; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2021; ISBN 9780128224670. [Google Scholar]
- Maiese, K. Biomarkers for Parkinson’s Disease and Neurodegenerative Disorders: A Role for Non-coding RNAs. Curr. Neurovasc. Res. 2022, 19, 127–130. [Google Scholar] [CrossRef]
- Tang, B.; Zeng, W.; Song, L.L.; Wang, H.M.; Qu, L.Q.; Lo, H.H.; Yu, L.; Wu, A.G.; Wong, V.K.W.; Law, B.Y.K. Extracellular Vesicle Delivery of Neferine for the Attenuation of Neurodegenerative Disease Proteins and Motor Deficit in an Alzheimer’s Disease Mouse Model. Pharmaceuticals 2022, 15, 83. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Targeting molecules to medicine with mTOR, autophagy and neurodegenerative disorders. Br. J. Clin. Pharm. 2016, 82, 1245–1266. [Google Scholar] [CrossRef]
- World Health Organization. Dementia: A Public Health Priority; World Health Organization: Geneva, Switzerland, 2012; pp. 1–4.
- Maiese, K. MicroRNAs for the Treatment of Dementia and Alzheimer’s Disease. Curr. Neurovasc. Res. 2019, 16, 1–2. [Google Scholar] [CrossRef]
- Maiese, K. Impacting dementia and cognitive loss with innovative strategies: Mechanistic target of rapamycin, clock genes, circular non-coding ribonucleic acids, and Rho/Rock. Neural Regen. Res. 2019, 14, 773–774. [Google Scholar] [CrossRef]
- Maiese, K. The mechanistic target of rapamycin (mTOR) and the silent mating-type information regulation 2 homolog 1 (SIRT1): Oversight for neurodegenerative disorders. Biochem. Soc. Trans. 2018, 46, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.R.; Qu, Y.J.; Hu, B.; An, H.M. Signal pathways in the treatment of Alzheimer’s disease with traditional Chinese medicine. Biomed. Pharm. 2022, 152, 113208. [Google Scholar] [CrossRef]
- Rapaka, D.; Bitra, V.R.; Challa, S.R.; Adiukwu, P.C. mTOR signaling as a molecular target for the alleviation of Alzheimer’s disease pathogenesis. Neurochem. Int. 2022, 155, 105311. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Apolipoprotein-ε4 allele (APOE-ε4) as a Mediator of Cognitive Loss and Dementia in Long COVID-19. Curr. Neurovasc. Res. 2022, 19, 435–439. [Google Scholar] [CrossRef]
- Ullah, H.; Hussain, A.; Asif, M.; Nawaz, F.; Rasool, M. Natural products as bioactive agents in the prevention of dementia. CNS Neurol. Disord. Drug Targets 2022, 22, 466–476. [Google Scholar] [CrossRef]
- Zhu, G.; Tong, Q.; Ye, X.; Li, J.; Zhou, L.; Sun, P.; Liang, F.; Zhong, S.; Cheng, R.; Zhang, J. Phototherapy for Cognitive Function in Patients with Dementia: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2022, 14, 936489. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, A.; Reynolds, R. Diverse pathways to neuronal necroptosis in Alzheimer’s disease. Eur. J. Neurosci. 2022, 56, 5428–5441. [Google Scholar] [CrossRef] [PubMed]
- Mavroidi, B.; Kaminari, A.; Matiadis, D.; Hadjipavlou-Litina, D.; Pelecanou, M.; Tzinia, A.; Sagnou, M. The Prophylactic and Multimodal Activity of Two Isatin Thiosemicarbazones against Alzheimer’s Disease In Vitro. Brain Sci. 2022, 12, 806. [Google Scholar] [CrossRef]
- Maiese, K. Addressing Alzheimer’s Disease and Cognitive Loss through Autophagy. Curr. Neurovasc. Res. 2020, 17, 339–341. [Google Scholar] [CrossRef]
- Engin, A.B.; Engin, A. Alzheimer’s Disease and Protein Kinases. Adv. Exp. Med. Biol. 2021, 1275, 285–321. [Google Scholar] [CrossRef]
- Hsieh, C.F.; Liu, C.K.; Lee, C.T.; Yu, L.E.; Wang, J.Y. Acute glucose fluctuation impacts microglial activity, leading to inflammatory activation or self-degradation. Sci. Rep. 2019, 9, 840. [Google Scholar] [CrossRef]
- Hu, Z.; Jiao, R.; Wang, P.; Zhu, Y.; Zhao, J.; De Jager, P.; Bennett, D.A.; Jin, L.; Xiong, M. Shared Causal Paths underlying Alzheimer’s dementia and Type 2 Diabetes. Sci. Rep. 2020, 10, 4107. [Google Scholar] [CrossRef] [PubMed]
- Min, A.Y.; Yoo, J.M.; Sok, D.E.; Kim, M.R. Mulberry Fruit Prevents Diabetes and Diabetic Dementia by Regulation of Blood Glucose through Upregulation of Antioxidative Activities and CREB/BDNF Pathway in Alloxan-Induced Diabetic Mice. Oxid. Med. Cell. Longev. 2020, 2020, 1298691. [Google Scholar] [CrossRef]
- Su, M.; Naderi, K.; Samson, N.; Youssef, I.; Fulop, L.; Bozso, Z.; Laroche, S.; Delatour, B.; Davis, S. Mechanisms Associated with Type 2 Diabetes as a Risk Factor for Alzheimer-Related Pathology. Mol. Neurobiol. 2019, 56, 5815–5834. [Google Scholar] [CrossRef]
- Caberlotto, L.; Nguyen, T.P.; Lauria, M.; Priami, C.; Rimondini, R.; Maioli, S.; Cedazo-Minguez, A.; Sita, G.; Morroni, F.; Corsi, M.; et al. Cross-disease analysis of Alzheimer’s disease and type-2 Diabetes highlights the role of autophagy in the pathophysiology of two highly comorbid diseases. Sci. Rep. 2019, 9, 3965. [Google Scholar] [CrossRef]
- Jiang, W.J.; Peng, Y.C.; Yang, K.M. Cellular signaling pathways regulating beta-cell proliferation as a promising therapeutic target in the treatment of diabetes. Exp. Ther. Med. 2018, 16, 3275–3285. [Google Scholar] [CrossRef]
- Khan, H.; Tundis, R.; Ullah, H.; Aschner, M.; Belwal, T.; Mirzaei, H.; Akkol, E.K. Flavonoids targeting NRF2 in neurodegenerative disorders. Food Chem. Toxicol. 2020, 146, 111817. [Google Scholar] [CrossRef]
- Lin, X.; Zhang, N. Berberine: Pathways to protect neurons. Phytother. Res. PTR 2018, 32, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C. Raves and risks for erythropoietin. Cytokine Growth Factor Rev. 2008, 19, 145–155. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, H.; Yu, P.; Qian, T.; Xu, X. Protective or Harmful: The Dual Roles of Autophagy in Diabetic Retinopathy. Front. Med. 2021, 8, 644121. [Google Scholar] [CrossRef]
- Ponnalagu, M.; Subramani, M.; Jayadev, C.; Shetty, R.; Das, D. Retinal pigment epithelium-secretome: A diabetic retinopathy perspective. Cytokine 2017, 95, 126–135. [Google Scholar] [CrossRef]
- Tomita, Y.; Lee, D.; Tsubota, K.; Kurihara, T. PPARα Agonist Oral Therapy in Diabetic Retinopathy. Biomedicines 2020, 8, 433. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chong, Z.Z.; Maiese, K. Cell Life Versus Cell Longevity: The Mysteries Surrounding the NAD(+) Precursor Nicotinamide. Curr. Med. Chem. 2006, 13, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Huo, F.; Liu, B.; Liu, J.; Chen, T.; Li, J.; Zhu, Z.; Lv, B. Crocin Inhibits Oxidative Stress and Pro-inflammatory Response of Microglial Cells Associated with Diabetic Retinopathy Through the Activation of PI3K/Akt Signaling Pathway. J. Mol. Neurosci. 2017, 61, 581–589. [Google Scholar] [CrossRef]
- Chong, Z.Z.; Li, F.; Maiese, K. The pro-survival pathways of mTOR and protein kinase B target glycogen synthase kinase-3beta and nuclear factor-kappaB to foster endogenous microglial cell protection. Int. J. Mol. Med. 2007, 19, 263–272. [Google Scholar]
- Li, F.; Chong, Z.Z.; Maiese, K. Microglial integrity is maintained by erythropoietin through integration of Akt and its substrates of glycogen synthase kinase-3beta, beta-catenin, and nuclear factor-kappaB. Curr. Neurovasc. Res. 2006, 3, 187–201. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.C.; Chong, Z.Z.; Hou, J.; Maiese, K. FoxO3a governs early microglial proliferation and employs mitochondrial depolarization with caspase 3, 8, and 9 cleavage during oxidant induced apoptosis. Curr. Neurovasc. Res. 2009, 6, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Albiero, M.; Poncina, N.; Tjwa, M.; Ciciliot, S.; Menegazzo, L.; Ceolotto, G.; Vigili de Kreutzenberg, S.; Moura, R.; Giorgio, M.; Pelicci, P.; et al. Diabetes causes bone marrow autonomic neuropathy and impairs stem cell mobilization via dysregulated p66Shc and Sirt1. Diabetes 2014, 63, 1353–1365. [Google Scholar] [CrossRef] [PubMed]
- Atef, M.M.; El-Sayed, N.M.; Ahmed, A.A.M.; Mostafa, Y.M. Donepezil improves neuropathy through activation of AMPK signalling pathway in streptozotocin-induced diabetic mice. Biochem. Pharm. 2019, 159, 1–10. [Google Scholar] [CrossRef]
- Gomes, M.B.; Negrato, C.A. Alpha-lipoic acid as a pleiotropic compound with potential therapeutic use in diabetes and other chronic diseases. Diabetol. Metab. Syndr. 2014, 6, 80. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. FoxO Transcription Factors and Regenerative Pathways in Diabetes Mellitus. Curr. Neurovasc. Res. 2015, 12, 404–413. [Google Scholar] [CrossRef]
- Prattichizzo, F.; De Nigris, V.; Spiga, R.; Mancuso, E.; La Sala, L.; Antonicelli, R.; Testa, R.; Procopio, A.D.; Olivieri, F.; Ceriello, A. Inflammageing and metaflammation: The yin and yang of type 2 diabetes. Ageing Res. Rev. 2018, 41, 1–17. [Google Scholar] [CrossRef]
- Swain, O.; Romano, S.K.; Miryala, R.; Tsai, J.; Parikh, V.; Umanah, G.K.E. SARS-CoV-2 Neuronal Invasion and Complications: Potential Mechanisms and Therapeutic Approaches. J. Neurosci. 2021, 41, 5338–5349. [Google Scholar] [CrossRef] [PubMed]
- Bayaraa, O.; Inman, C.K.; Thomas, S.A.; Al Jallaf, F.; Alshaikh, M.; Idaghdour, Y.; Ashall, L. Hyperglycemic conditions induce rapid cell dysfunction-promoting transcriptional alterations in human aortic endothelial cells. Sci. Rep. 2022, 12, 20912. [Google Scholar] [CrossRef]
- Maiese, K. mTOR: Driving apoptosis and autophagy for neurocardiac complications of diabetes mellitus. World J. Diabetes 2015, 6, 217–224. [Google Scholar] [CrossRef]
- Hajibabaie, F.; Abedpoor, N.; Safavi, K.; Taghian, F. Natural remedies medicine derived from flaxseed (secoisolariciresinol diglucoside, lignans, and α-linolenic acid) improve network targeting efficiency of diabetic heart conditions based on computational chemistry techniques and pharmacophore modeling. J. Food Biochem. 2022, 46, e14480. [Google Scholar] [CrossRef]
- Januszewski, A.S.; Watson, C.J.; O’Neill, V.; McDonald, K.; Ledwidge, M.; Robson, T.; Jenkins, A.J.; Keech, A.C.; McClements, L. FKBPL is associated with metabolic parameters and is a novel determinant of cardiovascular disease. Sci. Rep. 2020, 10, 21655. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Liu, J.; Wu, Y.; Xi, W.; Wei, Y.; Yuan, Z.; Zhuo, X. Zinc supplementation protects against diabetic endothelial dysfunction via GTP cyclohydrolase 1 restoration. Biochem. Biophys. Res. Commun. 2020, 521, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Disease onset and aging in the world of circular RNAs. J. Transl. Sci. 2016, 2, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Mocayar Marón, F.J.; Ferder, L.; Reiter, R.J.; Manucha, W. Daily and seasonal mitochondrial protection: Unraveling common possible mechanisms involving vitamin D and melatonin. J. Steroid Biochem. Mol. Biol. 2020, 199, 105595. [Google Scholar] [CrossRef]
- Pabel, S.; Hamdani, N.; Luedde, M.; Sossalla, S. SGLT2 Inhibitors and Their Mode of Action in Heart Failure-Has the Mystery Been Unravelled? Curr. Heart Fail. Rep. 2021, 18, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Zarneshan, S.N.; Fakhri, S.; Farzaei, M.H.; Khan, H.; Saso, L. Astaxanthin targets PI3K/Akt signaling pathway toward potential therapeutic applications. Food Chem. Toxicol. 2020, 145, 111714. [Google Scholar] [CrossRef]
- Zhou, Q.; Tang, S.; Zhang, X.; Chen, L. Targeting PRAS40: A novel therapeutic strategy for human diseases. J. Drug Target. 2021, 29, 1–44. [Google Scholar] [CrossRef]
- Chiareli, R.A.; Carvalho, G.A.; Marques, B.L.; Mota, L.S.; Oliveira-Lima, O.C.; Gomes, R.M.; Birbrair, A.; Gomez, R.S.; Simão, F.; Klempin, F.; et al. The Role of Astrocytes in the Neurorepair Process. Front. Cell Dev. Biol. 2021, 9, 665795. [Google Scholar] [CrossRef]
- Gu, H.F.; Li, N.; Tang, Y.L.; Yan, C.Q.; Shi, Z.; Yi, S.N.; Zhou, H.L.; Liao, D.F.; OuYang, X.P. Nicotinate-curcumin ameliorates cognitive impairment in diabetic rats by rescuing autophagic flux in CA1 hippocampus. CNS Neurosci. 2019, 25, 430–441. [Google Scholar] [CrossRef]
- Xu, T.; Liu, J.; Li, X.R.; Yu, Y.; Luo, X.; Zheng, X.; Cheng, Y.; Yu, P.Q.; Liu, Y. The mTOR/NF-κB Pathway Mediates Neuroinflammation and Synaptic Plasticity in Diabetic Encephalopathy. Mol. Neurobiol. 2021, 58, 3848–3862. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Huang, C.; Zhu, S.Y.; Zou, H.C.; Xu, C.Y.; Chen, Y.X. Overexpression of HOTAIR attenuates Pi-induced vascular calcification by inhibiting Wnt/β-catenin through regulating miR-126/Klotho/SIRT1 axis. Mol. Cell. Biochem. 2021, 476, 3551–3561. [Google Scholar] [CrossRef] [PubMed]
- Chong, Z.Z.; Maiese, K. The Src homology 2 domain tyrosine phosphatases SHP-1 and SHP-2: Diversified control of cell growth, inflammation, and injury. Histol. Histopathol. 2007, 22, 1251–1267. [Google Scholar]
- El-Beltagy, A.; Saleh, A.M.B.; Attaallah, A.; Gahnem, R.A. Therapeutic role of Azadirachta indica leaves ethanolic extract against diabetic nephropathy in rats neonatally induced by streptozotocin. Ultrastruct. Pathol. 2021, 45, 391–406. [Google Scholar] [CrossRef]
- Kita, A.; Saito, Y.; Miura, N.; Miyajima, M.; Yamamoto, S.; Sato, T.; Yotsuyanagi, T.; Fujimiya, M.; Chikenji, T.S. Altered regulation of mesenchymal cell senescence in adipose tissue promotes pathological changes associated with diabetic wound healing. Commun. Biol. 2022, 5, 310. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cao, Q.; Gao, W.; Li, B.Y.; Zeng, C.; Xia, Z.; Zhao, B. Melatonin ameliorates cerebral ischemia-reperfusion injury in diabetic mice by enhancing autophagy via the SIRT1-BMAL1 pathway. FASEB J. 2021, 35, e22040. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C. OutFOXOing disease and disability: The therapeutic potential of targeting FoxO proteins. Trends Mol. Med. 2008, 14, 219–227. [Google Scholar] [CrossRef]
- Tan, S.; Zang, G.; Wang, Y.; Sun, Z.; Li, Y.; Lu, C.; Wang, Z. Differences of Angiogenesis Factors in Tumor and Diabetes Mellitus. Diabetes Metab. Syndr. Obes. 2021, 14, 3375–3388. [Google Scholar] [CrossRef]
- Wasserfurth, P.; Nebl, J.; Rühling, M.R.; Shammas, H.; Bednarczyk, J.; Koehler, K.; Boßlau, T.K.; Krüger, K.; Hahn, A.; Das, A.M. Impact of Dietary Modifications on Plasma Sirtuins 1, 3 and 5 in Older Overweight Individuals Undergoing 12-Weeks of Circuit Training. Nutrients 2021, 13, 3824. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, Q.; Sun, Y. FoxO1 Regulates Neuropeptide Y and Pro-opiomelanocortin in the Hypothalamus of Rat Offspring Small for Gestational Age. Reprod. Sci. 2021, 29, 173–183. [Google Scholar] [CrossRef]
- Lee, J.H.; Lee, J.H.; Jin, M.; Han, S.D.; Chon, G.R.; Kim, I.H.; Kim, S.; Kim, S.Y.; Choi, S.B.; Noh, Y.H. Diet control to achieve euglycemia induces significant loss of heart and liver weight via increased autophagy compared with ad libitum diet in diabetic rats. Exp. Mol. Med. 2014, 46, e111. [Google Scholar] [CrossRef]
- Li, J.; Lin, F.H.; Zhu, X.M.; Lv, Z.M. Impact of diabetic hyperglycaemia and insulin therapy on autophagy and impairment in rat epididymis. Andrologia 2020, 52, e13889. [Google Scholar] [CrossRef]
- van Dyck, C.H.; Swanson, C.J.; Aisen, P.; Bateman, R.J.; Chen, C.; Gee, M.; Kanekiyo, M.; Li, D.; Reyderman, L.; Cohen, S.; et al. Lecanemab in Early Alzheimer’s Disease. N. Engl. J. Med. 2023, 388, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, U.K.; Khan, R.; Mikhael, M.; Balez, R.; David, M.A.; Mahns, D.; Hardy, J.; Tayebi, M. Therapeutic anti-amyloid β antibodies cause neuronal disturbances. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2022. [Google Scholar] [CrossRef]
- Mahmoudi, N.; Kiasalari, Z.; Rahmani, T.; Sanaierad, A.; Afshin-Majd, S.; Naderi, G.; Baluchnejadmojarad, T.; Roghani, M. Diosgenin Attenuates Cognitive Impairment in Streptozotocin-Induced Diabetic Rats: Underlying Mechanisms. Neuropsychobiology 2021, 80, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. FoxO Proteins in the Nervous System. Anal. Cell. Pathol. 2015, 2015, 569392. [Google Scholar] [CrossRef]
- Maiese, K. Forkhead Transcription Factors: Formulating a FOXO Target for Cognitive Loss. Curr. Neurovasc. Res. 2017, 14, 415–420. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Hou, J.; Shang, Y.C. New strategies for Alzheimer’s disease and cognitive impairment. Oxid. Med. Cell. Longev. 2009, 2, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Sedighi, M.; Baluchnejadmojarad, T.; Afshin-Majd, S.; Amiri, M.; Aminzade, M.; Roghani, M. Anti-aging Klotho Protects SH-SY5Y Cells Against Amyloid β1-42 Neurotoxicity: Involvement of Wnt1/pCREB/Nrf2/HO-1 Signaling. J. Mol. Neurosci. 2020, 71, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.K.; Singh, T.G.; Singh, S.; Garg, N.; Dhiman, S. Apoptotic Pathways and Alzheimer’s Disease: Probing Therapeutic Potential. Neurochem. Res. 2021, 46, 3103–3122. [Google Scholar] [CrossRef]
- Wang, H.; Li, Q.; Sun, S.; Chen, S. Neuroprotective Effects of Salidroside in a Mouse Model of Alzheimer’s Disease. Cell. Mol. Neurobiol. 2020, 40, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, J.; Hou, Y.; Leverenz, J.B.; Kallianpur, A.; Mehra, R.; Liu, Y.; Yu, H.; Pieper, A.A.; Jehi, L.; et al. Network medicine links SARS-CoV-2/COVID-19 infection to brain microvascular injury and neuroinflammation in dementia-like cognitive impairment. Alzheimers Res. 2021, 13, 110. [Google Scholar] [CrossRef]
- Wang, R.; Zhu, Y.; Qin, L.F.; Xu, Z.G.; Gao, X.R.; Liu, C.B.; Xu, G.T.; Chen, Y.Z. Comprehensive Bibliometric Analysis of Stem Cell Research in Alzheimer’s Disease from 2004 to 2022. Dement. Geriatr. Cogn. Disord. 2023, 51, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Fessel, J. Cure of Alzheimer’s Dementia Requires Addressing All of the Affected Brain Cell Types. J. Clin. Med. 2023, 12, 2049. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, F.; Zhang, S.; Xin, R.; Sun, Y. Genetic and environmental factors in Alzheimer’s and Parkinson’s diseases and promising therapeutic intervention via fecal microbiota transplantation. NPJ Park. Dis. 2021, 7, 70. [Google Scholar] [CrossRef]
- Wahl, D.; Solon-Biet, S.M.; Cogger, V.C.; Fontana, L.; Simpson, S.J.; Le Couteur, D.G.; Ribeiro, R.V. Aging, lifestyle and dementia. Neurobiol. Dis. 2019, 130, 104481. [Google Scholar] [CrossRef]
- Ghiasi, R.; Naderi, R.; Sheervalilou, R.; Alipour, M.R. Swimming training by affecting the pancreatic Sirtuin1 (SIRT1) and oxidative stress, improves insulin sensitivity in diabetic male rats. Horm. Mol. Biol. Clin. Investig. 2019, 40. [Google Scholar] [CrossRef]
- Maiese, K. Triple play: Promoting neurovascular longevity with nicotinamide, WNT, and erythropoietin in diabetes mellitus. Biomed. Pharm. 2008, 62, 218–232. [Google Scholar] [CrossRef]
- Osorio Alves, J.; Matta Pereira, L.; Cabral Coutinho do Rego Monteiro, I.; Pontes Dos Santos, L.H.; Soares Marreiros Ferraz, A.; Carneiro Loureiro, A.C.; Calado Lima, C.; Leal-Cardoso, J.H.; Pires Carvalho, D.; Soares Fortunato, R.; et al. Strenuous Acute Exercise Induces Slow and Fast Twitch-Dependent NADPH Oxidase Expression in Rat Skeletal Muscle. Antioxidants 2020, 9, 57. [Google Scholar] [CrossRef]
- Rezaee, Z.; Marandi, S.M.; Alaei, H.; Esfarjani, F. Exercise-Induced Neuroprotection in the 6-Hydroxydopamine Parkinson’s Disease Model. Neurotox. Res. 2020, 38, 850–858. [Google Scholar] [CrossRef]
- Zhang, H.; Liang, J.; Chen, N. Do not neglect the role of circadian rhythm in muscle atrophy. Ageing Res. Rev. 2020, 63, 101155. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. New Directions for Dementia. Curr. Neurovasc. Res. 2017, 14, 305. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z. Insights into oxidative stress and potential novel therapeutic targets for Alzheimer disease. Restor. Neurol. Neurosci. 2004, 22, 87–104. [Google Scholar]
- Querfurth, H.; Lee, H.K. Mammalian/mechanistic target of rapamycin (mTOR) complexes in neurodegeneration. Mol. Neurodegener. 2021, 16, 44. [Google Scholar] [CrossRef]
- Wang, F.; Cao, Y.; Ma, L.; Pei, H.; Rausch, W.D.; Li, H. Dysfunction of Cerebrovascular Endothelial Cells: Prelude to Vascular Dementia. Front. Aging Neurosci. 2018, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.M.; Chuang, Y.W.; Sun, K.T.; Yu, M.C.; Kung, S.C.; Lee, B.K.; Huang, S.T.; Chen, C.H.; Lin, C.L.; Kao, C.H. Polycystic kidney disease is significantly associated with dementia risk. Neurology 2017, 89, 1457–1463. [Google Scholar] [CrossRef]
- Chen, X.; Le, Y.; He, W.Y.; He, J.; Wang, Y.H.; Zhang, L.; Xiong, Q.M.; Zheng, X.Q.; Liu, K.X.; Wang, H.B. Abnormal Insulin-like Growth Factor 1 Signaling Regulates Neuropathic Pain by Mediating the Mechanistic Target of Rapamycin-Related Autophagy and Neuroinflammation in Mice. ACS Chem. Neurosci. 2021, 12, 2917–2928. [Google Scholar] [CrossRef]
- González-Fernández, C.; González, P.; González-Pérez, F.; Rodríguez, F. Characterization of Ex Vivo and In Vitro Wnt Transcriptome Induced by Spinal Cord Injury in Rat Microglial Cells. Brain Sci. 2022, 12, 708. [Google Scholar] [CrossRef]
- Maiese, K. Inflammatory glial cells of the nervous system: Assistants or assassins? Curr. Neurovasc. Res. 2005, 2, 187–188. [Google Scholar] [CrossRef]
- Maiese, K.; Li, F.; Chong, Z.Z. Erythropoietin in the brain: Can the promise to protect be fulfilled? Trends Pharm. Sci. 2004, 25, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Jarero-Basulto, J.; Rivera-Cervantes, M.; Gasca-Martínez, D.; García-Sierra, F.; Gasca-Martínez, Y.; Beas-Zárate, C. Current Evidence on the Protective Effects of Recombinant Human Erythropoietin and Its Molecular Variants against Pathological Hallmarks of Alzheimer’s Disease. Pharmaceuticals 2020, 13, 424. [Google Scholar] [CrossRef] [PubMed]
- Kaur, D.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Badavath, V.N.; Ul Hassan, S.S.; Hasan, M.M.; Bhatia, S.; Al-Harassi, A.; et al. Unravelling the potential neuroprotective facets of erythropoietin for the treatment of Alzheimer’s disease. Metab. Brain Dis. 2021, 37, 1–16. [Google Scholar] [CrossRef]
- Barchetta, I.; Cimini, F.A.; Ciccarelli, G.; Baroni, M.G.; Cavallo, M.G. Sick fat: The good and the bad of old and new circulating markers of adipose tissue inflammation. J. Endocrinol. Investig. 2019, 42, 1257–1272. [Google Scholar] [CrossRef]
- Li, L.; Sun, Y.; Zhang, Y.; Wang, W.; Ye, C. Mutant Huntingtin Impairs Pancreatic β-cells by Recruiting IRS-2 and Disturbing the PI3K/AKT/FoxO1 Signaling Pathway in Huntington’s Disease. J. Mol. Neurosci. 2021, 71, 2646–2658. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Wang, B.; Wu, C.; Li, D.; Wu, Y.; Ye, L.; Ye, L.; Chen, X.; Li, P.; Yuan, Y.; et al. Acidic fibroblast growth factor attenuates type 2 diabetes-induced demyelination via suppressing oxidative stress damage. Cell Death Dis. 2021, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition). Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C.; Wang, S. Targeting disease through novel pathways of apoptosis and autophagy. Expert Opin. Ther. Targets 2012, 16, 1203–1214. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, H.; Rong, Z.; Chen, L. Wnt family member 1 (Wnt1) overexpression-induced M2 polarization of microglia alleviates inflammation-sensitized neonatal brain injuries. Bioengineered 2022, 13, 12409–12420. [Google Scholar] [CrossRef]
- He, C.; Xu, Y.; Sun, J.; Li, L.; Zhang, J.H.; Wang, Y. Autophagy and Apoptosis in Acute CNS injuries: From Mechanism to Treatment. Antioxid. Redox Signal. 2022, 38, 234–257. [Google Scholar] [CrossRef]
- Qin, C.; Lu, Y.; Bai, L.; Wang, K. The molecular regulation of autophagy in antimicrobial immunity. J. Mol. Cell Biol. 2022, 14, mjac015. [Google Scholar] [CrossRef]
- Senousy, M.A.; Hanafy, M.E.; Shehata, N.; Rizk, S.M. Erythropoietin and Bacillus Calmette-Guérin Vaccination Mitigate 3-Nitropropionic Acid-Induced Huntington-like Disease in Rats by Modulating the PI3K/Akt/mTOR/P70S6K Pathway and Enhancing the Autophagy. ACS Chem. Neurosci. 2022, 13, 721–732. [Google Scholar] [CrossRef]
- Hou, J.; Chong, Z.Z.; Shang, Y.C.; Maiese, K. Early apoptotic vascular signaling is determined by Sirt1 through nuclear shuttling, forkhead trafficking, bad, and mitochondrial caspase activation. Curr. Neurovasc. Res. 2010, 7, 95–112. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.C.; Chong, Z.Z.; Hou, J.; Maiese, K. Wnt1, FoxO3a, and NF-kappaB oversee microglial integrity and activation during oxidant stress. Cell Signal. 2010, 22, 1317–1329. [Google Scholar] [CrossRef]
- Taveira, G.B.; Mello, E.O.; Souza, S.B.; Monteiro, R.M.; Ramos, A.C.; Carvalho, A.O.; Rodrigues, R.; Okorokov, L.A.; Gomes, V.M. Programmed cell death in yeast by thionin-like peptide from Capsicum annuum fruits involving activation of capases and extracelullar H(+) flux. Biosci. Rep. 2018, 38, BSR20180119. [Google Scholar] [CrossRef]
- Almasieh, M.; Catrinescu, M.M.; Binan, L.; Costantino, S.; Levin, L.A. Axonal Degeneration in Retinal Ganglion Cells is Associated with a Membrane Polarity-Sensitive Redox Process. J. Neurosci. 2017, 37, 3824–3839. [Google Scholar] [CrossRef]
- Viola, G.; Bortolozzi, R.; Hamel, E.; Moro, S.; Brun, P.; Castagliuolo, I.; Ferlin, M.G.; Basso, G. MG-2477, a new tubulin inhibitor, induces autophagy through inhibition of the Akt/mTOR pathway and delayed apoptosis in A549 cells. Biochem. Pharm. 2012, 83, 16–26. [Google Scholar] [CrossRef]
- Chong, Z.Z.; Kang, J.Q.; Maiese, K. Erythropoietin is a novel vascular protectant through activation of Akt1 and mitochondrial modulation of cysteine proteases. Circulation 2002, 106, 2973–2979. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. The bright side of reactive oxygen species: Lifespan extension without cellular demise. J. Transl. Sci. 2016, 2, 185–187. [Google Scholar] [CrossRef]
- Watroba, M.; Szukiewicz, D. Sirtuins at the Service of Healthy Longevity. Front. Physiol. 2021, 12, 724506. [Google Scholar] [CrossRef]
- Yousafzai, N.A.; Jin, H.; Ullah, M.; Wang, X. Recent advances of SIRT1 and implications in chemotherapeutics resistance in cancer. Am. J. Cancer Res. 2021, 11, 5233–5248. [Google Scholar] [PubMed]
- Maiese, K. A Common Link in Neurovascular Regenerative Pathways: Protein Kinase B (Akt). Curr. Neurovasc. Res. 2022, 19, 1–4. [Google Scholar] [CrossRef]
- Maiese, K. Pyroptosis, Apoptosis, and Autophagy: Critical Players of Inflammation and Cell Demise in the Nervous System. Curr. Neurovasc. Res. 2022, 19, 241–244. [Google Scholar] [CrossRef]
- Pang, Y.; Qin, M.; Hu, P.; Ji, K.; Xiao, R.; Sun, N.; Pan, X.; Zhang, X. Resveratrol protects retinal ganglion cells against ischemia induced damage by increasing Opa1 expression. Int. J. Mol. Med. 2020, 46, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.J.; Fossum, S.L.; Fimbel, S.M.; Montgomery, J.E.; Hyde, D.R. The inhibitor of phagocytosis, O-phospho-L-serine, suppresses Muller glia proliferation and cone cell regeneration in the light-damaged zebrafish retina. Exp. Eye Res. 2010, 91, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Sun, C.; Lei, M.; Li, G.; Yi, L.; Luo, F.; Li, Y.; Ding, L.; Liu, Z.; Li, S.; et al. Activation of Wnt/beta-catenin Pathway by Exogenous Wnt1 Protects SH-SY5Y Cells Against 6-Hydroxydopamine Toxicity. J. Mol. Neurosci. 2013, 49, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kang, I.H.; Nam, J.B.; Cho, Y.; Chung, D.Y.; Kim, S.H.; Kim, J.S.; Cho, Y.D.; Hong, E.K.; Sohn, N.W.; et al. Ameliorating the Effect of Astragaloside IV on Learning and Memory Deficit after Chronic Cerebral Hypoperfusion in Rats. Molecules 2015, 20, 1904–1921. [Google Scholar] [CrossRef]
- Xin, Y.J.; Yuan, B.; Yu, B.; Wang, Y.Q.; Wu, J.J.; Zhou, W.H.; Qiu, Z. Tet1-mediated DNA demethylation regulates neuronal cell death induced by oxidative stress. Sci. Rep. 2015, 5, 7645. [Google Scholar] [CrossRef]
- Yu, T.; Li, L.; Chen, T.; Liu, Z.; Liu, H.; Li, Z. Erythropoietin attenuates advanced glycation endproducts-induced toxicity of schwann cells in vitro. Neurochem. Res. 2015, 40, 698–712. [Google Scholar] [CrossRef]
- Lan, T.; Xu, Y.; Li, S.; Li, N.; Zhang, S.; Zhu, H. Cornin protects against cerebral ischemia/reperfusion injury by preventing autophagy via the PI3K/Akt/mTOR pathway. BMC Pharm. Toxicol. 2022, 23, 82. [Google Scholar] [CrossRef]
- Liu, L.; Xu, S.; Li, P.; Li, L. A novel adipokine WISP1 attenuates lipopolysaccharide-induced cell injury in 3T3-L1 adipocytes by regulating the PI3K/Akt pathway. Obes. Res. Clin. Pract. 2022, 16, 122–129. [Google Scholar] [CrossRef]
- Mansour, R.M.; El Sayed, N.S.; Ahmed, M.A.E.; El-Sahar, A.E. Addressing Peroxisome Proliferator-Activated Receptor-gamma in 3-Nitropropionic Acid-Induced Striatal Neurotoxicity in Rats. Mol. Neurobiol. 2022, 59, 4368–4383. [Google Scholar] [CrossRef]
- Sabzali, M.; Eidi, A.; Khaksari, M.; Khastar, H. Anti-inflammatory, Antioxidant, and Antiapoptotic Action of Metformin Attenuates Ethanol Neurotoxicity in the Animal Model of Fetal Alcohol Spectrum Disorders. Neurotox. Res. 2022, 40, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Chong, Z.Z.; Li, F.; Maiese, K. Oxidative stress in the brain: Novel cellular targets that govern survival during neurodegenerative disease. Prog. Neurobiol. 2005, 75, 207–246. [Google Scholar] [CrossRef]
- Maiese, K. WISP1: Clinical Insights for a Proliferative and Restorative Member of the CCN Family. Curr. Neurovasc. Res. 2014, 11, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Xue, M.; Deng, H.; Cheng, S.; Hu, Y.; Zhou, C. Ginsenoside and Its Therapeutic Potential for Cognitive Impairment. Biomolecules 2022, 12, 1310. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wu, Z.; Peng, Y.; Gao, J.; Zheng, F.; Tan, J.; Xu, J.; Wang, T. Neuroprotection of Triptolide against Amyloid-Beta1-42-induced toxicity via the Akt/mTOR/p70S6K-mediated Autophagy Pathway. Acad. Bras. Cienc. 2022, 94, e20210938. [Google Scholar] [CrossRef]
- Zhuang, X.; Ma, J.; Xu, G.; Sun, Z. SHP-1 knockdown suppresses mitochondrial biogenesis and aggravates mitochondria-dependent apoptosis induced by all trans retinal through the STING/AMPK pathways. Mol. Med. 2022, 28, 125. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, D.; Kumari, R.; Ilyas, A.; Tyagi, S.; Kumar, R.; Poddar, N.K. Crosstalk between epigenetics and mTOR as a gateway to new insights in pathophysiology and treatment of Alzheimer’s disease. Int. J. Biol. Macromol. 2021, 192, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Ali, E.S.; Mitra, K.; Akter, S.; Ramproshad, S.; Mondal, B.; Khan, I.N.; Islam, M.T.; Sharifi-Rad, J.; Calina, D.; Cho, W.C. Recent advances and limitations of mTOR inhibitors in the treatment of cancer. Cancer Cell Int. 2022, 22, 284. [Google Scholar] [CrossRef]
- Arias, C.; Salazar, L.A. Autophagy and Polyphenols in Osteoarthritis: A Focus on Epigenetic Regulation. Int. J. Mol. Sci. 2021, 23, 421. [Google Scholar] [CrossRef]
- Barthels, D.; Prateeksha, P.; Nozohouri, S.; Villalba, H.; Zhang, Y.; Sharma, S.; Anderson, S.; Howlader, M.S.I.; Nambiar, A.; Abbruscato, T.J.; et al. Dental Pulp-Derived Stem Cells Preserve Astrocyte Health During Induced Gliosis by Modulating Mitochondrial Activity and Functions. Cell. Mol. Neurobiol. 2022. [Google Scholar] [CrossRef]
- Bonam, S.R.; Tranchant, C.; Muller, S. Autophagy-Lysosomal Pathway as Potential Therapeutic Target in Parkinson’s Disease. Cells 2021, 10, 3547. [Google Scholar] [CrossRef]
- Casciano, F.; Zauli, E.; Rimondi, E.; Mura, M.; Previati, M.; Busin, M.; Zauli, G. The role of the mTOR pathway in diabetic retinopathy. Front. Med. 2022, 9, 973856. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zeng, L.; Yan, F.; Liu, J.; Qin, M.; Wang, F.; Zhang, X. Long-term oral administration of naringenin counteracts aging-related retinal degeneration via regulation of mitochondrial dynamics and autophagy. Front. Pharmacol. 2022, 13, 919905. [Google Scholar] [CrossRef]
- Jobst, M.; Kiss, E.; Gerner, C.; Marko, D.; Del Favero, G. Activation of autophagy triggers mitochondrial loss and changes acetylation profile relevant for mechanotransduction in bladder cancer cells. Arch. Toxicol. 2022, 97, 217–233. [Google Scholar] [CrossRef]
- Maiese, K. Taking aim at Alzheimer’s disease through the mammalian target of rapamycin. Ann. Med. 2014, 46, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Sakai, M.; Yu, Z.; Hirayama, R.; Nakasato, M.; Kikuchi, Y.; Ono, C.; Komatsu, H.; Nakanishi, M.; Yoshii, H.; Stellwagen, D.; et al. Deficient Autophagy in Microglia Aggravates Repeated Social Defeat Stress-Induced Social Avoidance. Neural Plast. 2022, 2022, 7503553. [Google Scholar] [CrossRef]
- Corti, O.; Blomgren, K.; Poletti, A.; Beart, P.M. Autophagy in neurodegeneration: New insights underpinning therapy for neurological diseases. J. Neurochem. 2020, 154, e15002. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Warming Up to New Possibilities with the Capsaicin Receptor TRPV1: mTOR, AMPK, and Erythropoietin. Curr. Neurovasc. Res. 2017, 14, 184–189. [Google Scholar] [CrossRef]
- Maiese, K. Driving neural regeneration through the mammalian target of rapamycin. Neural Regen. Res. 2014, 9, 1413–1417. [Google Scholar] [CrossRef]
- Moors, T.E.; Hoozemans, J.J.; Ingrassia, A.; Beccari, T.; Parnetti, L.; Chartier-Harlin, M.C.; van de Berg, W.D. Therapeutic potential of autophagy-enhancing agents in Parkinson’s disease. Mol. Neurodegener. 2017, 12, 11. [Google Scholar] [CrossRef]
- Zhou, Z.D.; Selvaratnam, T.; Lee, J.C.T.; Chao, Y.X.; Tan, E.K. Molecular targets for modulating the protein translation vital to proteostasis and neuron degeneration in Parkinson’s disease. Transl. Neurodegener. 2019, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. NoNovel Stem Cell Strategies with mTOR. In Molecules to Medicine with mTOR: Translating Critical Pathways into Novel Therapeutic Strategies; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2016; pp. 3–22. [Google Scholar] [CrossRef]
- He, W.; Gao, Y.; Zhou, J.; Shi, Y.; Xia, D.; Shen, H.M. Friend or Foe? Implication of the autophagy-lysosome pathway in SARS-CoV-2 infection and COVID-19. Int. J. Biol. Sci. 2022, 18, 4690–4703. [Google Scholar] [CrossRef]
- Maiese, K. The Mechanistic Target of Rapamycin (mTOR): Novel Considerations as an Antiviral Treatment. Curr. Neurovasc Res. 2020, 17, 332–337. [Google Scholar] [CrossRef]
- Maiese, K. Circadian Clock Genes: Targeting Innate Immunity for Antiviral Strategies Against COVID-19. Curr. Neurovasc. Res. 2020, 17, 531–533. [Google Scholar] [CrossRef] [PubMed]
- Pinchera, B.; Scotto, R.; Buonomo, A.R.; Zappulo, E.; Stagnaro, F.; Gallicchio, A.; Viceconte, G.; Sardanelli, A.; Mercinelli, S.; Villari, R.; et al. Diabetes and COVID-19: The potential role of mTOR. Diabetes Res. Clin. Pract. 2022, 186, 109813. [Google Scholar] [CrossRef]
- Theoharides, T.C. Could SARS-CoV-2 Spike Protein Be Responsible for Long-COVID Syndrome? Mol. Neurobiol. 2022, 59, 1850–1861. [Google Scholar] [CrossRef] [PubMed]
- You, H.; Zhao, Q.; Dong, M. The Key Genes Underlying Pathophysiology Correlation Between the Acute Myocardial Infarction and COVID-19. Int. J. Gen. Med. 2022, 15, 2479–2490. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, T.; Wang, Y.; Yang, S.; Luo, J.; Fang, F.; Liao, J.; Wen, W.; Cui, H.; Shang, H. Qing-Wen-Jie-Re Mixture Ameliorates Poly (I:C)-Induced Viral Pneumonia Through Regulating the Inflammatory Response and Serum Metabolism. Front. Pharmacol. 2022, 13, 891851. [Google Scholar] [CrossRef]
- Bello-Perez, M.; Sola, I.; Novoa, B.; Klionsky, D.J.; Falco, A. Canonical and Noncanonical Autophagy as Potential Targets for COVID-19. Cells 2020, 9, 1619. [Google Scholar] [CrossRef]
- Cavalli, E.; Bramanti, A.; Ciurleo, R.; Tchorbanov, A.I.; Giordano, A.; Fagone, P.; Belizna, C.; Bramanti, P.; Shoenfeld, Y.; Nicoletti, F. Entangling COVID-19 associated thrombosis into a secondary antiphospholipid antibody syndrome: Diagnostic and therapeutic perspectives (Review). Int. J. Mol. Med. 2020, 46, 903–912. [Google Scholar] [CrossRef]
- Evans, T.; Kok, W.L.; Cowan, K.; Hefford, M.; Anichtchik, O. Accumulation of beta-synuclein in cortical neurons is associated with autophagy attenuation in the brains of dementia with Lewy body patients. Brain Res. 2017, 1681, 1–13. [Google Scholar] [CrossRef]
- Maiese, K. Forkhead transcription factors: New considerations for alzheimer’s disease and dementia. J. Transl. Sci. 2016, 2, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Movahedpour, A.; Vakili, O.; Khalifeh, M.; Mousavi, P.; Mahmoodzadeh, A.; Taheri-Anganeh, M.; Razmeh, S.; Shabaninejad, Z.; Yousefi, F.; Behrouj, H.; et al. Mammalian target of rapamycin (mTOR) signaling pathway and traumatic brain injury: A novel insight into targeted therapy. Cell Biochem. Funct. 2022, 40, 232–247. [Google Scholar] [CrossRef] [PubMed]
- Perluigi, M.; Di Domenico, F.; Barone, E.; Butterfield, D.A. mTOR in Alzheimer disease and its earlier stages: Links to oxidative damage in the progression of this dementing disorder. Free. Radic. Biol. Med. 2021, 169, 382–396. [Google Scholar] [CrossRef]
- Cheng, J.; North, B.J.; Zhang, T.; Dai, X.; Tao, K.; Guo, J.; Wei, W. The emerging roles of protein homeostasis-governing pathways in Alzheimer’s disease. Aging Cell 2018, 17, e12801. [Google Scholar] [CrossRef]
- Morris, G.; Berk, M.; Maes, M.; Puri, B.K. Could Alzheimer’s Disease Originate in the Periphery and If So How So? Mol. Neurobiol. 2019, 56, 406–434. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Wu, Q.Y.; Zheng, R.; Chen, C.; Chen, Y.; Liu, Q.; Hoffmann, P.R.; Ni, J.Z.; Song, G.L. Selenomethionine mitigates cognitive decline by targeting both tau hyperphosphorylation and autophagic clearance in an Alzheimer’s disease mouse model. J. Neurosci. 2017, 37, 2449–2462. [Google Scholar] [CrossRef]
- Saleem, S.; Biswas, S.C. Tribbles Pseudokinase 3 Induces Both Apoptosis and Autophagy in Amyloid-beta-induced Neuronal Death. J. Biol. Chem. 2017, 292, 2571–2585. [Google Scholar] [CrossRef]
- Maiese, K. Novel Insights for Multiple Sclerosis and Demyelinating Disorders with Apoptosis, Autophagy, FoxO, and mTOR. Curr. Neurovasc. Res. 2021, 18, 169–171. [Google Scholar] [CrossRef]
- Farahani, M.; Niknam, Z.; Mohammadi Amirabad, L.; Amiri-Dashatan, N.; Koushki, M.; Nemati, M.; Danesh Pouya, F.; Rezaei-Tavirani, M.; Rasmi, Y.; Tayebi, L. Molecular pathways involved in COVID-19 and potential pathway-based therapeutic targets. Biomed. Pharm. 2021, 145, 112420. [Google Scholar] [CrossRef]
- Geng, K.; Ma, X.; Jiang, Z.; Huang, W.; Gao, C.; Pu, Y.; Luo, L.; Xu, Y.; Xu, Y. Innate Immunity in Diabetic Wound Healing: Focus on the Mastermind Hidden in Chronic Inflammatory. Front. Pharmacol. 2021, 12, 653940. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhao, Y.; Wang, Y.; Xie, R.; Tong, Y.; Sauer, J.D.; Gong, S. NAD(H)-loaded nanoparticles for efficient sepsis therapy via modulating immune and vascular homeostasis. Nat. Nanotechnol. 2022, 17, 880–890. [Google Scholar] [CrossRef]
- Wu, L.; Xiong, X.; Wu, X.; Ye, Y.; Jian, Z.; Zhi, Z.; Gu, L. Targeting Oxidative Stress and Inflammation to Prevent Ischemia-Reperfusion Injury. Front. Mol. Neurosci. 2020, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Chong, Z.Z.; Li, F.; Maiese, K. Stress in the brain: Novel cellular mechanisms of injury linked to Alzheimer’s disease. Brain Res. Brain Res. Rev. 2005, 49, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Li, Z.; Chen, C.; Liu, J.; Zhu, W.; She, L.; Huang, H.; Qin, Y.; Liu, G.; Wang, J.; et al. The Molecular Landscape and Biological Alterations Induced by PRAS40-Knockout in Head and Neck Squamous Cell Carcinoma. Front. Oncol. 2020, 10, 565669. [Google Scholar] [CrossRef]
- Damstra-Oddy, J.L.; Warren, E.C.; Perry, C.J.; Desfougères, Y.; Fitzpatrick, J.K.; Schaf, J.; Costelloe, L.; Hind, W.; Downer, E.J.; Saiardi, A.; et al. Phytocannabinoid-dependent mTORC1 regulation is dependent upon inositol polyphosphate multikinase activity. Br. J. Pharm. 2021, 178, 1149–1163. [Google Scholar] [CrossRef]
- Gao, J.; Yao, M.; Chang, D.; Liu, J. mTOR (Mammalian Target of Rapamycin): Hitting the Bull’s Eye for Enhancing Neurogenesis After Cerebral Ischemia? Stroke 2022, 54, 279–285. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C.; Wang, S. mTOR: On target for novel therapeutic strategies in the nervous system. Trends Mol. Med. 2013, 19, 51–60. [Google Scholar] [CrossRef]
- Xue, Q.; Nagy, J.A.; Manseau, E.J.; Phung, T.L.; Dvorak, H.F.; Benjamin, L.E. Rapamycin inhibition of the Akt/mTOR pathway blocks select stages of VEGF-A164-driven angiogenesis, in part by blocking S6Kinase. Arter. Thromb. Vasc. Biol. 2009, 29, 1172–1178. [Google Scholar] [CrossRef]
- Sergio, C.M.; Rolando, C.A. Erythropoietin regulates signaling pathways associated with neuroprotective events. Exp. Brain Res. 2022, 240, 1303–1315. [Google Scholar] [CrossRef]
- Maiese, K. Regeneration in the nervous system with erythropoietin. Front. Biosci. (Landmark Ed.) 2016, 21, 561–596. [Google Scholar] [CrossRef]
- Chong, Z.Z.; Shang, Y.C.; Wang, S.; Maiese, K. PRAS40 Is an Integral Regulatory Component of Erythropoietin mTOR Signaling and Cytoprotection. PLoS ONE 2012, 7, e45456. [Google Scholar] [CrossRef]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. WNT1 Inducible Signaling Pathway Protein 1 (WISP1) Targets PRAS40 to Govern beta-Amyloid Apoptotic Injury of Microglia. Curr. Neurovasc. Res. 2012, 9, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, Q.; Wen, Q.; Zheng, Y.; Philip, L.; Jiang, H.; Lin, J.; Zheng, W. Proline-rich Akt substrate of 40kDa (PRAS40): A novel downstream target of PI3k/Akt signaling pathway. Cell Signal. 2012, 24, 17–24. [Google Scholar] [CrossRef]
- Kim, D.H.; Sarbassov, D.D.; Ali, S.M.; Latek, R.R.; Guntur, K.V.; Erdjument-Bromage, H.; Tempst, P.; Sabatini, D.M. GbetaL, a positive regulator of the rapamycin-sensitive pathway required for the nutrient-sensitive interaction between raptor and mTOR. Mol. Cell 2003, 11, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Guertin, D.A.; Stevens, D.M.; Thoreen, C.C.; Burds, A.A.; Kalaany, N.Y.; Moffat, J.; Brown, M.; Fitzgerald, K.J.; Sabatini, D.M. Ablation in mice of the mTORC components raptor, rictor, or mLST8 reveals that mTORC2 is required for signaling to Akt-FOXO and PKCalpha, but not S6K1. Dev. Cell 2006, 11, 859–871. [Google Scholar] [CrossRef]
- Saenwongsa, W.; Nithichanon, A.; Chittaganpitch, M.; Buayai, K.; Kewcharoenwong, C.; Thumrongwilainet, B.; Butta, P.; Palaga, T.; Takahashi, Y.; Ato, M.; et al. Metformin-induced suppression of IFN-alpha via mTORC1 signalling following seasonal vaccination is associated with impaired antibody responses in type 2 diabetes. Sci. Rep. 2020, 10, 3229. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Y.; Li, D.; Zhang, N.; Liu, R.; Han, B.; Wei, C.; Liu, H.; Xu, X.; Hao, J. Everolimus (RAD001) ameliorates vascular cognitive impairment by regulating microglial function via the mTORC1 signaling pathway. J. Neuroimmunol. 2016, 299, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, P.; English, T.; Shi, J.; Smas, C.M.; Kandror, K.V. Mammalian target of rapamycin complex 1 suppresses lipolysis, stimulates lipogenesis, and promotes fat storage. Diabetes 2010, 59, 775–781. [Google Scholar] [CrossRef]
- Malla, R.; Wang, Y.; Chan, W.K.; Tiwari, A.K.; Faridi, J.S. Genetic ablation of PRAS40 improves glucose homeostasis via linking the AKT and mTOR pathways. Biochem. Pharm. 2015, 96, 65–75. [Google Scholar] [CrossRef]
- Hamada, S.; Hara, K.; Hamada, T.; Yasuda, H.; Moriyama, H.; Nakayama, R.; Nagata, M.; Yokono, K. Upregulation of the mammalian target of rapamycin complex 1 pathway by Ras homolog enriched in brain in pancreatic beta-cells leads to increased beta-cell mass and prevention of hyperglycemia. Diabetes 2009, 58, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Katsianou, M.A.; Papavassiliou, K.A.; Gargalionis, A.N.; Agrogiannis, G.; Korkolopoulou, P.; Panagopoulos, D.; Themistocleous, M.S.; Piperi, C.; Basdra, E.K.; Papavassiliou, A.G. Polycystin-1 regulates cell proliferation and migration through AKT/mTORC2 pathway in a human craniosynostosis cell model. J. Cell. Mol. Med. 2022, 26, 2428–2437. [Google Scholar] [CrossRef] [PubMed]
- Sadria, M.; Seo, D.; Layton, A.T. The mixed blessing of AMPK signaling in Cancer treatments. BMC Cancer 2022, 22, 105. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Martinez, J.M.; Alessi, D.R. mTOR complex 2 (mTORC2) controls hydrophobic motif phosphorylation and activation of serum- and glucocorticoid-induced protein kinase 1 (SGK1). Biochem. J. 2008, 416, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.R.; Sommer, E.M.; Sakamoto, K.; Wullschleger, S.; Alessi, D.R. Protor-1 is required for efficient mTORC2-mediated activation of SGK1 in the kidney. Biochem. J. 2011, 436, 169–179. [Google Scholar] [CrossRef]
- Frias, M.A.; Thoreen, C.C.; Jaffe, J.D.; Schroder, W.; Sculley, T.; Carr, S.A.; Sabatini, D.M. mSin1 is necessary for Akt/PKB phosphorylation, and its isoforms define three distinct mTORC2s. Curr. Biol. 2006, 16, 1865–1870. [Google Scholar] [CrossRef]
- Gu, Y.; Lindner, J.; Kumar, A.; Yuan, W.; Magnuson, M.A. Rictor/mTORC2 is essential for maintaining a balance between beta-cell proliferation and cell size. Diabetes 2011, 60, 827–837. [Google Scholar] [CrossRef]
- Wang, R.H.; Kim, H.S.; Xiao, C.; Xu, X.; Gavrilova, O.; Deng, C.X. Hepatic Sirt1 deficiency in mice impairs mTorc2/Akt signaling and results in hyperglycemia, oxidative damage, and insulin resistance. J. Clin. Investig. 2011, 121, 4477–4490. [Google Scholar] [CrossRef]
- Treins, C.; Alliouachene, S.; Hassouna, R.; Xie, Y.; Birnbaum, M.J.; Pende, M. The combined deletion of S6K1 and Akt2 deteriorates glycaemic control in high fat diet. Mol. Cell. Biol. 2012, 32, 4001–4011. [Google Scholar] [CrossRef]
- Chong, Z.Z.; Shang, Y.C.; Wang, S.; Maiese, K. Shedding new light on neurodegenerative diseases through the mammalian target of rapamycin. Prog. Neurobiol. 2012, 99, 128–148. [Google Scholar] [CrossRef]
- Maiese, K. Novel Treatment Strategies for the Nervous System: Circadian Clock Genes, Non-coding RNAs, and Forkhead Transcription Factors. Curr. Neurovasc. Res. 2018, 15, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Zhang, J.; Tang, P.; Tu, N.; Wang, K.; Wu, G. Overexpression of miR185 inhibits autophagy and apoptosis of dopaminergic neurons by regulating the AMPK/mTOR signaling pathway in Parkinson’s disease. Mol. Med. Rep. 2018, 17, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Chen, B.; Wang, X.; Wu, L.; Yang, Y.; Cheng, X.; Hu, Z.; Cai, X.; Yang, J.; Sun, X.; et al. Sulforaphane protects against rotenone-induced neurotoxicity in vivo: Involvement of the mTOR, Nrf2, and autophagy pathways. Sci. Rep. 2016, 6, 32206. [Google Scholar] [CrossRef]
- Teotia, P.; Van Hook, M.J.; Fischer, D.; Ahmad, I. Human retinal ganglion cell axon regeneration by recapitulating developmental mechanisms: Effects of recruitment of the mTOR pathway. Development 2019, 146, 178012. [Google Scholar] [CrossRef]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. Erythropoietin and Wnt1 Govern Pathways of mTOR, Apaf-1, and XIAP in Inflammatory Microglia. Curr. Neurovasc. Res. 2011, 8, 270–285. [Google Scholar] [CrossRef]
- Dong, J.; Li, H.; Bai, Y.; Wu, C. Muscone ameliorates diabetic peripheral neuropathy through activating AKT/mTOR signalling pathway. J. Pharm. Pharm. 2019, 71, 1706–1713. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Peripheral Neuropathy: An Early Indication for Systemic Disease that Involves the Mechanistic Target of Rapamycin (mTOR). Curr. Neurovasc. Res. 2023. ahead of print. [Google Scholar] [CrossRef]
- Bellozi, P.M.; Lima, I.V.; Doria, J.G.; Vieira, E.L.; Campos, A.C.; Candelario-Jalil, E.; Reis, H.J.; Teixeira, A.L.; Ribeiro, F.M.; de Oliveira, A.C. Neuroprotective effects of the anticancer drug NVP-BEZ235 (dactolisib) on amyloid-beta 1-42 induced neurotoxicity and memory impairment. Sci. Rep. 2016, 6, 25226. [Google Scholar] [CrossRef]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. Prevention of beta-amyloid degeneration of microglia by erythropoietin depends on Wnt1, the PI 3-K/mTOR pathway, Bad, and Bcl-xL. Aging 2012, 4, 187–201. [Google Scholar] [CrossRef]
- Shang, Y.C.; Chong, Z.Z.; Wang, S.; Maiese, K. Tuberous sclerosis protein 2 (TSC2) modulates CCN4 cytoprotection during apoptotic amyloid toxicity in microglia. Curr. Neurovasc. Res. 2013, 10, 29–38. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.X.; Liu, T.; Law, P.Y.; Loh, H.H.; Qiu, Y.; Chen, H.Z. mu-Opioid receptor attenuates Abeta oligomers-induced neurotoxicity through mTOR signaling. CNS Neurosci. 2015, 21, 8–14. [Google Scholar] [CrossRef]
- Park, J.A.; Lee, C.H. Temporal changes in mammalian target of rapamycin (mTOR) and phosphorylated-mTOR expressions in the hippocampal CA1 region of rat with vascular dementia. J. Vet. Sci. 2017, 18, 11–16. [Google Scholar] [CrossRef]
- Farmer, K.; Abd-Elrahman, K.S.; Derksen, A.; Rowe, E.M.; Thompson, A.M.; Rudyk, C.A.; Prowse, N.A.; Dwyer, Z.; Bureau, S.C.; Fortin, T.; et al. mGluR5 Allosteric Modulation Promotes Neurorecovery in a 6-OHDA-Toxicant Model of Parkinson’s Disease. Mol. Neurobiol. 2020, 57, 1418–1431. [Google Scholar] [CrossRef]
- Dai, C.; Tang, S.; Biao, X.; Xiao, X.; Chen, C.; Li, J. Colistin induced peripheral neurotoxicity involves mitochondrial dysfunction and oxidative stress in mice. Mol. Biol. Rep. 2019, 46, 1963–1972. [Google Scholar] [CrossRef]
- Huang, D.; Shen, S.; Cai, M.; Jin, L.; Lu, J.; Xu, K.; Zhang, J.; Feng, G.; Hu, Y.; Zheng, K.; et al. Role of mTOR complex in IGF-1 induced neural differentiation of DPSCs. J. Mol. Histol. 2019, 50, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.S.; Wang, Y.F.; Long, X.X.; Ma, Y. Mangiferin Potentiates Neuroprotection by Isoflurane in Neonatal Hypoxic Brain Injury by Reducing Oxidative Stress and Activation of Phosphatidylinositol-3-Kinase/Akt/Mammalian Target of Rapamycin (PI3K/Akt/mTOR) Signaling. Med. Sci. Monit. 2018, 24, 7459–7468. [Google Scholar] [CrossRef]
- Dong, W.; Wang, R.; Ma, L.N.; Xu, B.L.; Zhang, J.S.; Zhao, Z.W.; Wang, Y.L.; Zhang, X. Influence of age-related learning and memory capacity of mice: Different effects of a high and low caloric diet. Aging Clin. Exp. Res. 2016, 28, 303–311. [Google Scholar] [CrossRef]
- Han, K.; Jia, N.; Zhong, Y.; Shang, X. S14G-humanin alleviates insulin resistance and increases autophagy in neurons of APP/PS1 transgenic mouse. J. Cell. Biochem. 2017, 119, 3111–3119. [Google Scholar] [CrossRef]
- Dai, C.; Ciccotosto, G.D.; Cappai, R.; Wang, Y.; Tang, S.; Hoyer, D.; Schneider, E.K.; Velkov, T.; Xiao, X. Rapamycin confers neuroprotection against colistin-induced oxidative stress, mitochondria dysfunction and apoptosis through the activation of autophagy and mTOR/Akt/CREB signaling pathways. ACS Chem. Neurosci. 2017, 9, 824–837. [Google Scholar] [CrossRef] [PubMed]
- Park, A.; Koh, H.C. NF-kappaB/mTOR-mediated autophagy can regulate diquat-induced apoptosis. Arch. Toxicol. 2019, 93, 1239–1253. [Google Scholar] [CrossRef]
- Javdan, N.; Ayatollahi, S.A.; Choudhary, M.I.; Al-Hasani, S.; Kobarfard, F.; Athar, A.; Pazoki-Toroudi, H. Capsaicin protects against testicular torsion injury through mTOR-dependent mechanism. Theriogenology 2018, 113, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, Q.; Wang, Y.; Li, J.; Lu, G.; Liu, Z. Glutamine protects against oxidative stress injury through inhibiting the activation of PI3K/Akt signaling pathway in parkinsonian cell model. Environ. Health Prev. Med. 2019, 24, 4. [Google Scholar] [CrossRef]
- Liu, P.; Yang, X.; Hei, C.; Meli, Y.; Niu, J.; Sun, T.; Li, P.A. Rapamycin Reduced Ischemic Brain Damage in Diabetic Animals Is Associated with Suppressions of mTOR and ERK1/2 Signaling. Int. J. Biol. Sci. 2016, 12, 1032–1040. [Google Scholar] [CrossRef]
- Lim, Y.M.; Lim, H.; Hur, K.Y.; Quan, W.; Lee, H.Y.; Cheon, H.; Ryu, D.; Koo, S.H.; Kim, H.L.; Kim, J.; et al. Systemic autophagy insufficiency compromises adaptation to metabolic stress and facilitates progression from obesity to diabetes. Nat. Commun. 2014, 5, 4934. [Google Scholar] [CrossRef]
- Ma, L.; Fu, R.; Duan, Z.; Lu, J.; Gao, J.; Tian, L.; Lv, Z.; Chen, Z.; Han, J.; Jia, L.; et al. Sirt1 is essential for resveratrol enhancement of hypoxia-induced autophagy in the type 2 diabetic nephropathy rat. Pathol. Res. Pract. 2016, 212, 310–318. [Google Scholar] [CrossRef]
- Liu, Z.; Stanojevic, V.; Brindamour, L.J.; Habener, J.F. GLP1-derived nonapeptide GLP1(28-36)amide protects pancreatic beta-cells from glucolipotoxicity. J. Endocrinol. 2012, 213, 143–154. [Google Scholar] [CrossRef]
- He, C.; Bassik, M.C.; Moresi, V.; Sun, K.; Wei, Y.; Zou, Z.; An, Z.; Loh, J.; Fisher, J.; Sun, Q.; et al. Exercise-induced BCL2-regulated autophagy is required for muscle glucose homeostasis. Nature 2012, 481, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Palanivel, R.; Rai, E.; Park, M.; Gabor, T.V.; Scheid, M.P.; Xu, A.; Sweeney, G. Adiponectin stimulates autophagy and reduces oxidative stress to enhance insulin sensitivity during high fat diet feeding in mice. Diabetes 2014, 64, 36–48. [Google Scholar] [CrossRef]
- Radulovic, J.; Gabbay, V. PFC mTOR signaling as a biological signature for cognitive deficits in bipolar disorder without psychosis. Cell. Rep. Med. 2021, 2, 100282. [Google Scholar] [CrossRef]
- Thomas, S.D.; Jha, N.K.; Ojha, S.; Sadek, B. mTOR Signaling Disruption and Its Association with the Development of Autism Spectrum Disorder. Molecules 2023, 28, 1889. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.A.; Shin, Y.J.; Akram, M.; Kim, E.S.; Choi, K.W.; Suh, H.; Lee, C.H.; Bae, O.N. High glucose condition induces autophagy in endothelial progenitor cells contributing to angiogenic impairment. Biol. Pharm. Bull. 2014, 37, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Lai, D.; Lu, P.; Gao, J.; He, H. ERK and Akt signaling pathways are involved in advanced glycation end product-induced autophagy in rat vascular smooth muscle cells. Int. J. Mol. Med. 2012, 29, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hong, Y.; Lee, S.R.; Chang, K.T. Autophagy contributes to retardation of cardiac growth in diabetic rats. Lab. Anim. Res. 2012, 28, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Martino, L.; Masini, M.; Novelli, M.; Beffy, P.; Bugliani, M.; Marselli, L.; Masiello, P.; Marchetti, P.; De Tata, V. Palmitate activates autophagy in INS-1E beta-cells and in isolated rat and human pancreatic islets. PLoS ONE 2012, 7, e36188. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Li, F.; Chong, Z.Z. New avenues of exploration for erythropoietin. JAMA 2005, 293, 90–95. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Koh, S.H.; Song, K.M.; Seol, I.J.; Park, H.K. The Akt/mTOR/p70S6K Pathway Is Involved in the Neuroprotective Effect of Erythropoietin on Hypoxic/Ischemic Brain Injury in a Neonatal Rat Model. Neonatology 2016, 110, 93–100. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Shang, Y.C.; Wang, S. Erythropoietin: New directions for the nervous system. Int. J. Mol. Sci. 2012, 13, 11102–11129. [Google Scholar] [CrossRef]
- Wang, G.B.; Ni, Y.L.; Zhou, X.P.; Zhang, W.F. The AKT/mTOR pathway mediates neuronal protective effects of erythropoietin in sepsis. Mol. Cell. Biochem. 2014, 385, 125–132. [Google Scholar] [CrossRef]
- Ka, M.; Smith, A.L.; Kim, W.Y. MTOR controls genesis and autophagy of GABAergic interneurons during brain development. Autophagy 2017, 13, 1348–1363. [Google Scholar] [CrossRef]
- Blagosklonny, M.V. From causes of aging to death from COVID-19. Aging 2020, 12, 10004–10021. [Google Scholar] [CrossRef]
- Lally, M.A.; Tsoukas, P.; Halladay, C.W.; O’Neill, E.; Gravenstein, S.; Rudolph, J.L. Metformin is Associated with Decreased 30-Day Mortality Among Nursing Home Residents Infected with SARS-CoV2. J. Am. Med. Dir. Assoc. 2021, 22, 193–198. [Google Scholar] [CrossRef]
- Pende, M.; Kozma, S.C.; Jaquet, M.; Oorschot, V.; Burcelin, R.; Le Marchand-Brustel, Y.; Klumperman, J.; Thorens, B.; Thomas, G. Hypoinsulinaemia, glucose intolerance and diminished beta-cell size in S6K1-deficient mice. Nature 2000, 408, 994–997. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Flati, V.; Paiardi, S.; Rizzoni, D.; Porteri, E.; Aquilani, R.; Assanelli, D.; Corsetti, G.; Speca, S.; Rezzani, R.; et al. Intracellular molecular effects of insulin resistance in patients with metabolic syndrome. Cardiovasc. Diabetol. 2010, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wu, J.; Zheng, F.; Jin, M.; Li, H. Glucagon-like peptide-1 analog-mediated protection against cholesterol-induced apoptosis via mammalian target of rapamycin activation in pancreatic betaTC-6 cells -1mTORbetaTC-6. J. Diabetes 2015, 7, 231–239. [Google Scholar] [CrossRef]
- Liu, Y.W.; Zhang, L.; Li, Y.; Cheng, Y.Q.; Zhu, X.; Zhang, F.; Yin, X.X. Activation of mTOR signaling mediates the increased expression of AChE in high glucose condition: In vitro and in vivo evidences. Mol. Neurobiol. 2015, 53, 4972–4980. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.Y.; Gu, Z.Y.; Liu, P.; Hu, Y.; Li, L.; Gong, Y.P.; Shu, H.; Liu, Y.; Li, C.L. The human glucagon-like peptide-1 analogue liraglutide regulates pancreatic beta-cell proliferation and apoptosis via an AMPK/mTOR/P70S6K signaling pathway. Peptides 2013, 39, 71–79. [Google Scholar] [CrossRef]
- Fraenkel, M.; Ketzinel-Gilad, M.; Ariav, Y.; Pappo, O.; Karaca, M.; Castel, J.; Berthault, M.F.; Magnan, C.; Cerasi, E.; Kaiser, N.; et al. mTOR inhibition by rapamycin prevents beta-cell adaptation to hyperglycemia and exacerbates the metabolic state in type 2 diabetes. Diabetes 2008, 57, 945–957. [Google Scholar] [CrossRef]
- Sataranatarajan, K.; Ikeno, Y.; Bokov, A.; Feliers, D.; Yalamanchili, H.; Lee, H.J.; Mariappan, M.M.; Tabatabai-Mir, H.; Diaz, V.; Prasad, S.; et al. Rapamycin Increases Mortality in db/db Mice, a Mouse Model of Type 2 Diabetes. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 850–857. [Google Scholar] [CrossRef]
- Deblon, N.; Bourgoin, L.; Veyrat-Durebex, C.; Peyrou, M.; Vinciguerra, M.; Caillon, A.; Maeder, C.; Fournier, M.; Montet, X.; Rohner-Jeanrenaud, F.; et al. Chronic mTOR inhibition by rapamycin induces muscle insulin resistance despite weight loss in rats. Br. J. Pharm. 2012, 165, 2325–2340. [Google Scholar] [CrossRef]
- Crespo, M.C.; Tome-Carneiro, J.; Pintado, C.; Davalos, A.; Visioli, F.; Burgos-Ramos, E. Hydroxytyrosol restores proper insulin signaling in an astrocytic model of Alzheimer’s disease. BioFactors 2017, 43, 540–548. [Google Scholar] [CrossRef]
- Pal, P.B.; Sonowal, H.; Shukla, K.; Srivastava, S.K.; Ramana, K.V. Aldose reductase regulates hyperglycemia-induced HUVEC death via SIRT1/AMPK-alpha1/mTOR pathway. J. Mol. Endocrinol. 2019, 63, 11–25. [Google Scholar] [CrossRef]
- Hua, K.; Li, T.; He, Y.; Guan, A.; Chen, L.; Gao, Y.; Xu, Q.; Wang, H.; Luo, R.; Zhao, L.; et al. Resistin secreted by porcine alveolar macrophages leads to endothelial cell dysfunction during Haemophilus parasuis infection. Virulence 2023, 14, 2171636. [Google Scholar] [CrossRef] [PubMed]
- Guimera, A.M.; Clark, P.; Wordsworth, J.; Anugula, S.; Rasmussen, L.J.; Shanley, D.P. Systems modelling predicts chronic inflammation and genomic instability prevent effective mitochondrial regulation during biological ageing. Exp. Gerontol. 2022, 166, 111889. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Harnessing the Power of SIRT1 and Non-coding RNAs in Vascular Disease. Curr. Neurovasc. Res. 2017, 14, 82–88. [Google Scholar] [CrossRef]
- Peixoto, C.A.; de Oliveira, W.H.; da Rocha Araujo, S.M.; Nunes, A.K.S. AMPK activation: Role in the signaling pathways of neuroinflammation and neurodegeneration. Exp. Neurol. 2017, 298 Pt A, 31–41. [Google Scholar] [CrossRef]
- Sato, T.; Nakashima, A.; Guo, L.; Tamanoi, F. Specific activation of mTORC1 by Rheb G-protein in vitro involves enhanced recruitment of its substrate protein. J. Biol. Chem. 2009, 284, 12783–12791. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Zhu, T.; Guan, K.L. TSC2 mediates cellular energy response to control cell growth and survival. Cell 2003, 115, 577–590. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Samadi, N.; Shahnazi, V.; Mihanfar, A.; Fattahi, A.; Latifi, Z.; Bahrami-Asl, Z.; Roshangar, L.; Nouri, M. Nicotinamide and its metabolite N1-Methylnicotinamide alleviate endocrine and metabolic abnormalities in adipose and ovarian tissues in rat model of Polycystic Ovary Syndrome. Chem. Biol. Interact. 2020, 324, 109093. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.K.; Xu, X.J.; Lawson, E.; Deoliveira, R.; Brandon, A.E.; Kraegen, E.W.; Ruderman, N.B. Downregulation of AMPK accompanies leucine- and glucose-induced increases in protein synthesis and insulin resistance in rat skeletal muscle. Diabetes 2010, 59, 2426–2434. [Google Scholar] [CrossRef]
- Balan, V.; Miller, G.S.; Kaplun, L.; Balan, K.; Chong, Z.Z.; Li, F.; Kaplun, A.; VanBerkum, M.F.; Arking, R.; Freeman, D.C.; et al. Life span extension and neuronal cell protection by Drosophila nicotinamidase. J. Biol. Chem. 2008, 283, 27810–27819. [Google Scholar] [CrossRef]
- Moroz, N.; Carmona, J.J.; Anderson, E.; Hart, A.C.; Sinclair, D.A.; Blackwell, T.K. Dietary restriction involves NAD -dependent mechanisms and a shift toward oxidative metabolism. Aging Cell 2014, 13, 1075–1085. [Google Scholar] [CrossRef]
- Xia, W.; Zhang, F.; Xie, C.; Jiang, M.; Hou, M. Macrophage migration inhibitory factor confers resistance to senescence through CD74-dependent AMPK-FOXO3a signaling in mesenchymal stem cells. Stem Cell Res. Ther. 2015, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Chiu, S.C.; Chao, C.Y.; Chiang, E.I.; Syu, J.N.; Rodriguez, R.L.; Tang, F.Y. N-3 polyunsaturated fatty acids alleviate high glucose-mediated dysfunction of endothelial progenitor cells and prevent ischemic injuries both in vitro and in vivo. J. Nutr. Biochem. 2017, 42, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yang, X.; Pang, X.; Zhao, Z.; Yu, H.; Zhou, H. Genistein protects against ox-LDL-induced senescence through enhancing SIRT1/LKB1/AMPK-mediated autophagy flux in HUVECs. Mol. Cell. Biochem. 2019, 455, 127–134. [Google Scholar] [CrossRef]
- Du, L.L.; Chai, D.M.; Zhao, L.N.; Li, X.H.; Zhang, F.C.; Zhang, H.B.; Liu, L.B.; Wu, K.; Liu, R.; Wang, J.Z.; et al. AMPK Activation Ameliorates Alzheimer’s Disease-Like Pathology and Spatial Memory Impairment in a Streptozotocin-Induced Alzheimer’s Disease Model in Rats. J. Alzheimers Dis. 2015, 43, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, Z.C.; Wang, K.F.; Chen, X.Y. Abeta peptide secretion is reduced by Radix Polygalaeinduced autophagy via activation of the AMPK/mTOR pathway. Mol. Med. Rep. 2015, 12, 2771–2776. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.L.; Huang, W.N.; Li, H.H.; Huang, C.N.; Hsieh, S.; Lai, C.; Lu, F.J. Hydrogen-rich water attenuates amyloid beta-induced cytotoxicity through upregulation of Sirt1-FoxO3a by stimulation of AMP-activated protein kinase in SK-N-MC cells. Chem. Biol. Interact. 2015, 240, 12–21. [Google Scholar] [CrossRef]
- Maiese, K. The Many Facets of Cell Injury: Angiogenesis to Autophagy. Curr. Neurovasc. Res. 2012, 9, 1–2. [Google Scholar] [CrossRef]
- Weikel, K.A.; Cacicedo, J.M.; Ruderman, N.B.; Ido, Y. Knockdown of GSK3beta Increases Basal Autophagy and AMPK Signaling in Nutrient-laden Human Aortic Endothelial Cells. Biosci. Rep. 2016, 36, e00382. [Google Scholar] [CrossRef]
- Dong, Y.; Chen, H.; Gao, J.; Liu, Y.; Li, J.; Wang, J. Molecular machinery and interplay of apoptosis and autophagy in coronary heart disease. J. Mol. Cell. Cardiol. 2019, 136, 27–41. [Google Scholar] [CrossRef]
- Shokri Afra, H.; Zangooei, M.; Meshkani, R.; Ghahremani, M.H.; Ilbeigi, D.; Khedri, A.; Shahmohamadnejad, S.; Khaghani, S.; Nourbakhsh, M. Hesperetin is a potent bioactivator that activates SIRT1-AMPK signaling pathway in HepG2 cells. J. Physiol. Biochem. 2019, 75, 125–133. [Google Scholar] [CrossRef]
- Zhao, D.; Sun, X.; Lv, S.; Sun, M.; Guo, H.; Zhai, Y.; Wang, Z.; Dai, P.; Zheng, L.; Ye, M.; et al. Salidroside attenuates oxidized lowdensity lipoproteininduced endothelial cell injury via promotion of the AMPK/SIRT1 pathway. Int. J. Mol. Med. 2019, 43, 2279–2290. [Google Scholar] [CrossRef] [PubMed]
- Jang, W.; Kim, H.J.; Li, H.; Jo, K.D.; Lee, M.K.; Yang, H.O. The Neuroprotective Effect of Erythropoietin on Rotenone-Induced Neurotoxicity in SH-SY5Y Cells Through the Induction of Autophagy. Mol. Neurobiol. 2015, 53, 3812–3821. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.F.; Kuo, Y.H.; Yeh, W.L.; Wu, C.Y.; Lin, H.Y.; Lai, S.W.; Liu, Y.S.; Wu, L.H.; Lu, J.K.; Lu, D.Y. Regulatory Effects of Caffeic Acid Phenethyl Ester on Neuroinflammation in Microglial Cells. Int. J. Mol. Sci. 2015, 16, 5572–5589. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Erythropoietin and mTOR: A “One-Two Punch” for Aging-Related Disorders Accompanied by Enhanced Life Expectancy. Curr. Neurovasc. Res. 2016, 13, 329–340. [Google Scholar] [CrossRef]
- Wang, L.; Di, L.; Noguchi, C.T. AMPK is involved in mediation of erythropoietin influence on metabolic activity and reactive oxygen species production in white adipocytes. Int. J. Biochem. Cell. Biol. 2014, 54, 1–9. [Google Scholar] [CrossRef]
- Andreucci, M.; Fuiano, G.; Presta, P.; Lucisano, G.; Leone, F.; Fuiano, L.; Bisesti, V.; Esposito, P.; Russo, D.; Memoli, B.; et al. Downregulation of cell survival signalling pathways and increased cell damage in hydrogen peroxide-treated human renal proximal tubular cells by alpha-erythropoietin. Cell Prolif. 2009, 42, 554–561. [Google Scholar] [CrossRef]
- Sanadgol, N.; Barati, M.; Houshmand, F.; Hassani, S.; Clarner, T.; Shahlaei, M.; Golab, F. Metformin accelerates myelin recovery and ameliorates behavioral deficits in the animal model of multiple sclerosis via adjustment of AMPK/Nrf2/mTOR signaling and maintenance of endogenous oligodendrogenesis during brain self-repairing period. Pharm. Rep. 2020, 72, 641–658. [Google Scholar] [CrossRef]
- Maiese, K. The Oversight of Circadian Clock Genes for the Detection, Prevention, and Treatment of COVID-19 Infection. Curr. Neurovasc. Res. 2021, 18, 471–473. [Google Scholar] [CrossRef]
- Kalender, A.; Selvaraj, A.; Kim, S.Y.; Gulati, P.; Brule, S.; Viollet, B.; Kemp, B.E.; Bardeesy, N.; Dennis, P.; Schlager, J.J.; et al. Metformin, independent of AMPK, inhibits mTORC1 in a rag GTPase-dependent manner. Cell Metab. 2010, 11, 390–401. [Google Scholar] [CrossRef]
- Oda, S.S. Metformin Protects against Experimental Acrylamide Neuropathy in Rats. Drug Dev. Res. 2017, 78, 349–359. [Google Scholar] [CrossRef]
- Ong, A.N.; Tan, C.C.; Cañete, M.T.; Lim, B.A.; Robles, J. Association Between Metformin Use and Mortality among Patients with Type 2 Diabetes Mellitus Hospitalized for COVID-19 Infection. J. ASEAN Fed. Endocr. Soc. 2021, 36, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Margrett, J.A.; Schofield, T.; Martin, P.; Poon, L.W.; Masaki, K.; Donlon, T.A.; Kallianpur, K.J.; Willcox, B.J. Novel Functional, Health, and Genetic Determinants of Cognitive Terminal Decline: Kuakini Honolulu Heart Program/Honolulu-Asia Aging Study. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 77, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Safdari Lord, J.; Soltani Rezaiezadeh, J.; Yekaninejad, M.S.; Izadi, P. The association of APOE genotype with COVID-19 disease severity. Sci. Rep. 2022, 12, 13483. [Google Scholar] [CrossRef]
- Maiese, K. Late Onset Alzheimer’s Disease: Novel Clinical Prospects for the Future. Curr. Neurovasc. Res. 2017, 14, 89. [Google Scholar] [CrossRef]
- Cacabelos, R.; Carril, J.C.; Cacabelos, N.; Kazantsev, A.G.; Vostrov, A.V.; Corzo, L.; Cacabelos, P.; Goldgaber, D. Sirtuins in Alzheimer’s Disease: SIRT2-Related GenoPhenotypes and Implications for PharmacoEpiGenetics. Int. J. Mol. Sci. 2019, 20, 1249. [Google Scholar] [CrossRef]
- Lee, G.; Pollard, H.B.; Arispe, N. Annexin 5 and apolipoprotein E2 protect against Alzheimer’s amyloid-beta-peptide cytotoxicity by competitive inhibition at a common phosphatidylserine interaction site. Peptides 2002, 23, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Jia, L.; Liu, C.C.; Li Zhong, Z.R.; Yang, L.; Chen, X.F.; Fryer, J.D.; Wang, X.; Zhang, Y.W.; Xu, H.; et al. TREM2 promotes microglial survival by activating Wnt/beta-catenin pathway. J. Neurosci. 2017, 37, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Chong, Z.Z.; Shang, Y.C.; Hou, J.; Maiese, K. Wnt1 neuroprotection translates into improved neurological function during oxidant stress and cerebral ischemia through AKT1 and mitochondrial apoptotic pathways. Oxid. Med. Cell. Longev. 2010, 3, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Chong, Z.Z. Nicotinamide: Necessary nutrient emerges as a novel cytoprotectant for the brain. Trends Pharm. Sci. 2003, 24, 228–232. [Google Scholar] [CrossRef]
- Maiese, K.; Chong, Z.Z.; Hou, J.; Shang, Y.C. The vitamin nicotinamide: Translating nutrition into clinical care. Molecules 2009, 14, 3446–3485. [Google Scholar] [CrossRef]
- Maiese, K.; Vincent, A.M. Membrane asymmetry and DNA degradation: Functionally distinct determinants of neuronal programmed cell death. J. Neurosci. Res. 2000, 59, 568–580. [Google Scholar] [CrossRef]
- Li, W.; Su, D.; Zhai, Q.; Chi, H.; She, X.; Gao, X.; Wang, K.; Yang, H.; Wang, R.; Cui, B. Proteomes analysis reveals the involvement of autophagy in AD-like neuropathology induced by noise exposure and ApoE4. Environ. Res. 2019, 176, 108537. [Google Scholar] [CrossRef] [PubMed]
- Ojo, J.O.; Reed, J.M.; Crynen, G.; Vallabhaneni, P.; Evans, J.; Shackleton, B.; Eisenbaum, M.; Ringland, C.; Edsell, A.; Mullan, M.; et al. APOE genotype dependent molecular abnormalities in the cerebrovasculature of Alzheimer’s disease and age-matched non-demented brains. Mol. Brain 2021, 14, 110. [Google Scholar] [CrossRef]
- Pontifex, M.G.; Martinsen, A.; Saleh, R.N.M.; Harden, G.; Tejera, N.; Müller, M.; Fox, C.; Vauzour, D.; Minihane, A.M. APOE4 genotype exacerbates the impact of menopause on cognition and synaptic plasticity in APOE-TR mice. FASEB J. 2021, 35, e21583. [Google Scholar] [CrossRef] [PubMed]
- Levine, K.S.; Leonard, H.L.; Blauwendraat, C.; Iwaki, H.; Johnson, N.; Bandres-Ciga, S.; Ferrucci, L.; Faghri, F.; Singleton, A.B.; Nalls, M.A. Virus exposure and neurodegenerative disease risk across national biobanks. Neuron 2023, 111, 1086–1093. [Google Scholar] [CrossRef]
- Kurki, S.N.; Kantonen, J.; Kaivola, K.; Hokkanen, L.; Mäyränpää, M.I.; Puttonen, H.; Martola, J.; Pöyhönen, M.; Kero, M.; Tuimala, J.; et al. APOE ε4 associates with increased risk of severe COVID-19, cerebral microhaemorrhages and post-COVID mental fatigue: A Finnish biobank, autopsy and clinical study. Acta Neuropathol. Commun. 2021, 9, 199. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Buhadily, A.K.; Al-Gareeb, A.I.; Alorabi, M.; Hadi Al-Harcan, N.A.; El-Bouseary, M.M.; Batiha, G.E. Citicoline and COVID-19: Vis-à-vis conjectured. Naunyn Schmiedebergs Arch. Pharm. 2022, 395, 1463–1475. [Google Scholar] [CrossRef]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-Maiahy, T.J.; Alexiou, A.; Mukerjee, N.; Batiha, G.E. Prostaglandins and non-steroidal anti-inflammatory drugs in COVID-19. Biotechnol. Genet. Eng. Rev. 2022, 38, 1–21. [Google Scholar] [CrossRef]
- Mahmud, N.; Anik, M.I.; Hossain, M.K.; Khan, M.I.; Uddin, S.; Ashrafuzzaman, M.; Rahaman, M.M. Advances in Nanomaterial-Based Platforms to Combat COVID-19: Diagnostics, Preventions, Therapeutics, and Vaccine Developments. ACS Appl. Bio Mater. 2022, 5, 2431–2460. [Google Scholar] [CrossRef]
- Shirzad, M.; Nourigorji, M.; Sajedi, A.; Ranjbar, M.; Rasti, F.; Sourani, Z.; Moradi, M.; Mostafa Mir, S.; Memar, M.Y. Targeted therapy in Coronavirus disease 2019 (COVID-19): Implication from cell and gene therapy to immunotherapy and vaccine. Int. Immunopharmacol. 2022, 111, 109161. [Google Scholar] [CrossRef]
- Benotmane, I.; Perrin, P.; Vargas, G.G.; Bassand, X.; Keller, N.; Lavaux, T.; Ohana, M.; Bedo, D.; Baldacini, C.; Sagnard, M.; et al. Biomarkers of Cytokine Release Syndrome Predict Disease Severity and Mortality From COVID-19 in Kidney Transplant Recipients. Transplantation 2021, 105, 158–169. [Google Scholar] [CrossRef]
- Cheema, P.S.; Nandi, D.; Nag, A. Exploring the therapeutic potential of forkhead box O for outfoxing COVID-19. Open Biol. 2021, 11, 210069. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ruiz, R.; Paredes, J.L.; Niewold, T.B. COVID-19 in patients with systemic lupus erythematosus: Lessons learned from the inflammatory disease. Transl. Res. 2021, 232, 13–36. [Google Scholar] [CrossRef]
- Maity, S.; Saha, A. Therapeutic Potential of Exploiting Autophagy Cascade Against Coronavirus Infection. Front. Microbiol. 2021, 12, 675419. [Google Scholar] [CrossRef] [PubMed]
- Jansen van Vuren, E.; Steyn, S.F.; Brink, C.B.; Möller, M.; Viljoen, F.P.; Harvey, B.H. The neuropsychiatric manifestations of COVID-19: Interactions with psychiatric illness and pharmacological treatment. Biomed. Pharm. 2021, 135, 111200. [Google Scholar] [CrossRef]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020, 26, 681–687. [Google Scholar] [CrossRef]
- Abu-Eid, R.; Ward, F.J. Targeting the PI3K/Akt/mTOR pathway: A therapeutic strategy in COVID-19 patients. Immunol. Lett. 2021, 240, 1–8. [Google Scholar] [CrossRef]
- Diallo, A.B.; Gay, L.; Coiffard, B.; Leone, M.; Mezouar, S.; Mege, J.L. Daytime variation in SARS-CoV-2 infection and cytokine production. Microb. Pathog. 2021, 158, 105067. [Google Scholar] [CrossRef] [PubMed]
- Ghasemnejad-Berenji, M. mTOR inhibition: A double-edged sword in patients with COVID-19? Hum. Cell 2021, 34, 698–699. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Chiramel, A.I.; Majdoul, S.; Lai, K.K.; Das, S.; Beare, P.A.; Andresson, T.; Best, S.M.; Compton, A.A. Rapalogs downmodulate intrinsic immunity and promote cell entry of SARS-CoV-2. bioRxiv 2021. [Google Scholar] [CrossRef] [PubMed]
- Borges do Nascimento, I.J.; Cacic, N.; Abdulazeem, H.M.; von Groote, T.C.; Jayarajah, U.; Weerasekara, I.; Esfahani, M.A.; Civile, V.T.; Marusic, A.; Jeroncic, A.; et al. Novel Coronavirus Infection (COVID-19) in Humans: A Scoping Review and Meta-Analysis. J. Clin. Med. 2020, 9, 941. [Google Scholar] [CrossRef] [PubMed]
- Gusev, E.; Sarapultsev, A.; Hu, D.; Chereshnev, V. Problems of Pathogenesis and Pathogenetic Therapy of COVID-19 from the Perspective of the General Theory of Pathological Systems (General Pathological Processes). Int. J. Mol. Sci. 2021, 22, 7582. [Google Scholar] [CrossRef] [PubMed]
- Hardeland, R. Redox Biology of Melatonin: Discriminating between Circadian and Non-circadian Functions. Antioxid. Redox Signal. 2022, 37, 704–725. [Google Scholar] [CrossRef] [PubMed]
- Pouresmaeil, V.; Al Abudi, A.H.; Mahimid, A.H.; Sarafraz Yazdi, M.; Es-Haghi, A. Evaluation of Serum Selenium and Copper Levels with Inflammatory Cytokines and Indices of Oxidative Stress in Type 2 Diabetes. Biol. Trace Elem. Res. 2022, 201, 617–626. [Google Scholar] [CrossRef]
- Kirchenwitz, M.; Stahnke, S.; Grunau, K.; Melcher, L.; van Ham, M.; Rottner, K.; Steffen, A.; Stradal, T.E.B. The autophagy inducer SMER28 attenuates microtubule dynamics mediating neuroprotection. Sci. Rep. 2022, 12, 17805. [Google Scholar] [CrossRef]
- Puri, D.; Kelkar, A.; Gaurishankar, B.; Subramanyam, D. Balance between autophagy and cell death is maintained by Polycomb-mediated regulation during stem cell differentiation. FEBS J. 2022, 290, 1625–1644. [Google Scholar] [CrossRef]
- Hu, G.; Wang, T.; Ma, C. EPO activates PI3K-IKKα-CDK1 signaling pathway to promote the proliferation of Glial Cells under hypoxia environment. Genet. Mol. Biol. 2022, 45, e20210249. [Google Scholar] [CrossRef]
- Temiz-Resitoglu, M.; Guden, D.S.; Senol, S.P.; Vezir, O.; Sucu, N.; Kibar, D.; Yılmaz, S.N.; Tunctan, B.; Malik, K.U.; Sahan-Firat, S. Pharmacological Inhibition of Mammalian Target of Rapamycin Attenuates Deoxycorticosterone Acetate Salt-Induced Hypertension and Related Pathophysiology: Regulation of Oxidative Stress, Inflammation, and Cardiovascular Hypertrophy in Male Rats. J. Cardiovasc. Pharm. 2022, 79, 355–367. [Google Scholar] [CrossRef]
- Xiong, J.; Bonney, S.; Gonçalves, R.V.; Esposito, D. Brassinosteroids control the inflammation, oxidative stress and cell migration through the control of mitochondrial function on skin regeneration. Life Sci. 2022, 307, 120887. [Google Scholar] [CrossRef]
- Zuo, J.; Zhang, Z.; Luo, M.; Zhou, L.; Nice, E.C.; Zhang, W.; Wang, C.; Huang, C. Redox signaling at the crossroads of human health and disease. MedComm 2022, 3, e127. [Google Scholar] [CrossRef]
- Toniolo, S.; Scarioni, M.; Di Lorenzo, F.; Hort, J.; Georges, J.; Tomic, S.; Nobili, F.; Frederiksen, K.S. Dementia and COVID-19, a Bidirectional Liaison: Risk Factors, Biomarkers, and Optimal Health Care. J. Alzheimers Dis. 2021, 82, 883–898. [Google Scholar] [CrossRef] [PubMed]
- Assogna, M.; Di Lorenzo, F.; Martorana, A.; Koch, G. Synaptic Effects of Palmitoylethanolamide in Neurodegenerative Disorders. Biomolecules 2022, 12, 1161. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.L.; Zhou, J.Y.; Liang, S.J.; Wang, X.Q. Impaired intestinal stem cell activity in ETEC infection: Enterotoxins, cyclic nucleotides, and Wnt signaling. Arch. Toxicol. 2022, 96, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Chen, Y.; Liu, R.; Li, W.; Hua, B.; Bao, Y. Wnt Signaling Pathways: A Role in Pain Processing. Neuromol. Med. 2022, 24, 233–249. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Chemaly, E.R.; Hajjar, R.J.; Lebeche, D. Resistin promotes cardiac hypertrophy via the AMP-activated protein kinase/mammalian target of rapamycin (AMPK/mTOR) and c-Jun N-terminal kinase/insulin receptor substrate 1 (JNK/IRS1) pathways. J. Biol. Chem. 2011, 286, 18465–18473. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Circadian Rhythm Pathways during Cognitive Impairment. Therapeutic strategies that can target cellular metabolism, apoptosis, autophagy, and pyroptosis, the mechanistic target of rapamycin (mTOR), AMP activated protein kinase (AMPK), growth factor signaling with erythropoietin (EPO), and risk factors such as the apolipoprotein E (APOE-ε4) gene and coronavirus disease 2019 (COVID-19) can offer exciting potential for the treatment of metabolic disorders linked to neurodegenerative disease. Conditions that involve long-COVID syndrome can be multi-factorial and involve metabolic pathways, programmed cell death mechanisms, mTOR pathways, growth factors, and APOE-ε4.
Figure 1.
Circadian Rhythm Pathways during Cognitive Impairment. Therapeutic strategies that can target cellular metabolism, apoptosis, autophagy, and pyroptosis, the mechanistic target of rapamycin (mTOR), AMP activated protein kinase (AMPK), growth factor signaling with erythropoietin (EPO), and risk factors such as the apolipoprotein E (APOE-ε4) gene and coronavirus disease 2019 (COVID-19) can offer exciting potential for the treatment of metabolic disorders linked to neurodegenerative disease. Conditions that involve long-COVID syndrome can be multi-factorial and involve metabolic pathways, programmed cell death mechanisms, mTOR pathways, growth factors, and APOE-ε4.


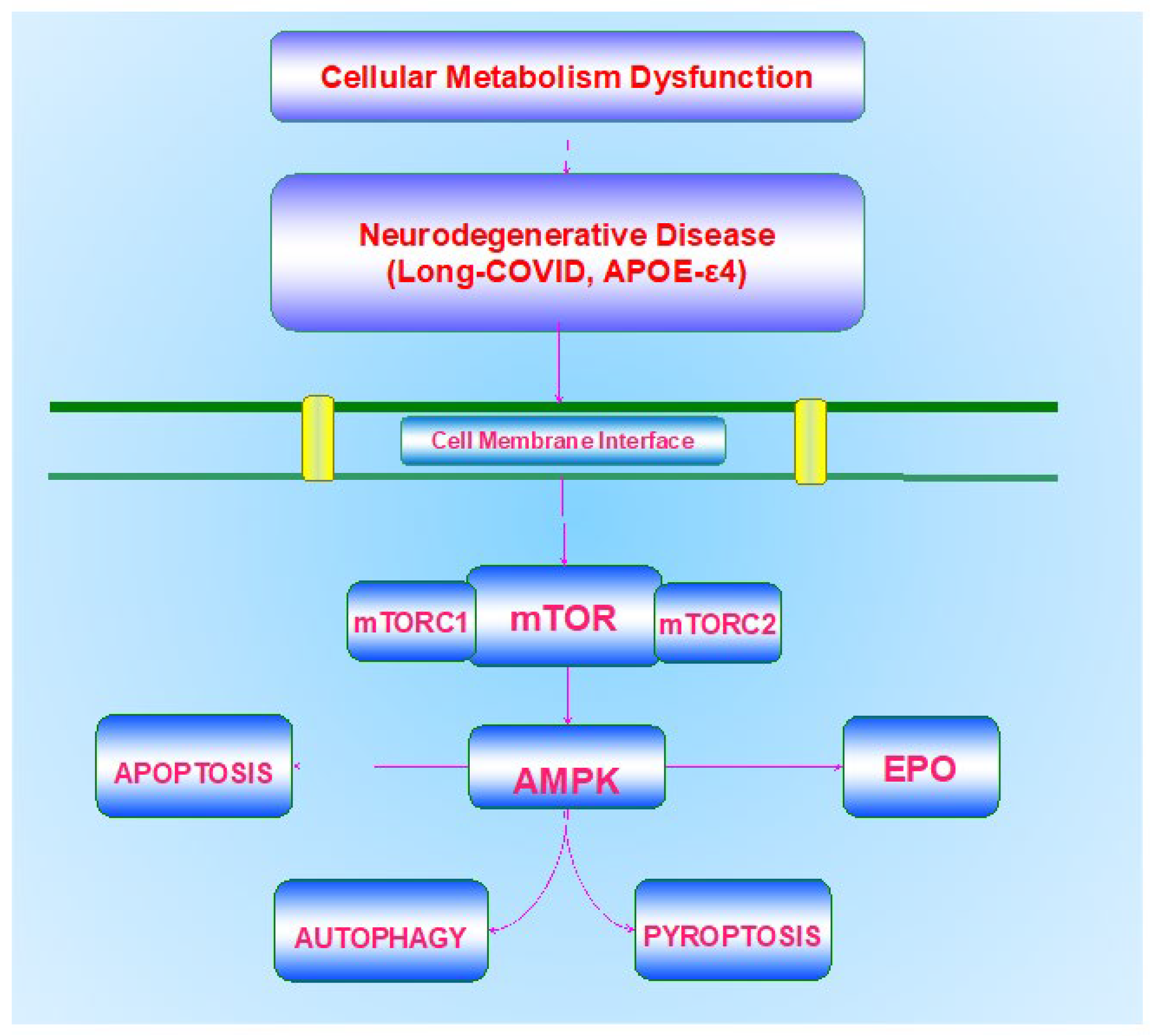{kind=link}
Table 1.
Highlighting Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System.
Table 1.
Highlighting Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System.
|
|
|
|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maiese, K. Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System. Biomolecules 2023, 13, 816. https://doi.org/10.3390/biom13050816
AMA Style
Maiese K. Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System. Biomolecules. 2023; 13(5):816. https://doi.org/10.3390/biom13050816
Chicago/Turabian StyleMaiese, Kenneth. 2023. "Cellular Metabolism: A Fundamental Component of Degeneration in the Nervous System" Biomolecules 13, no. 5: 816. https://doi.org/10.3390/biom13050816
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.