

Selenium and Selenoproteins in Health
Abstract
:1. Introduction
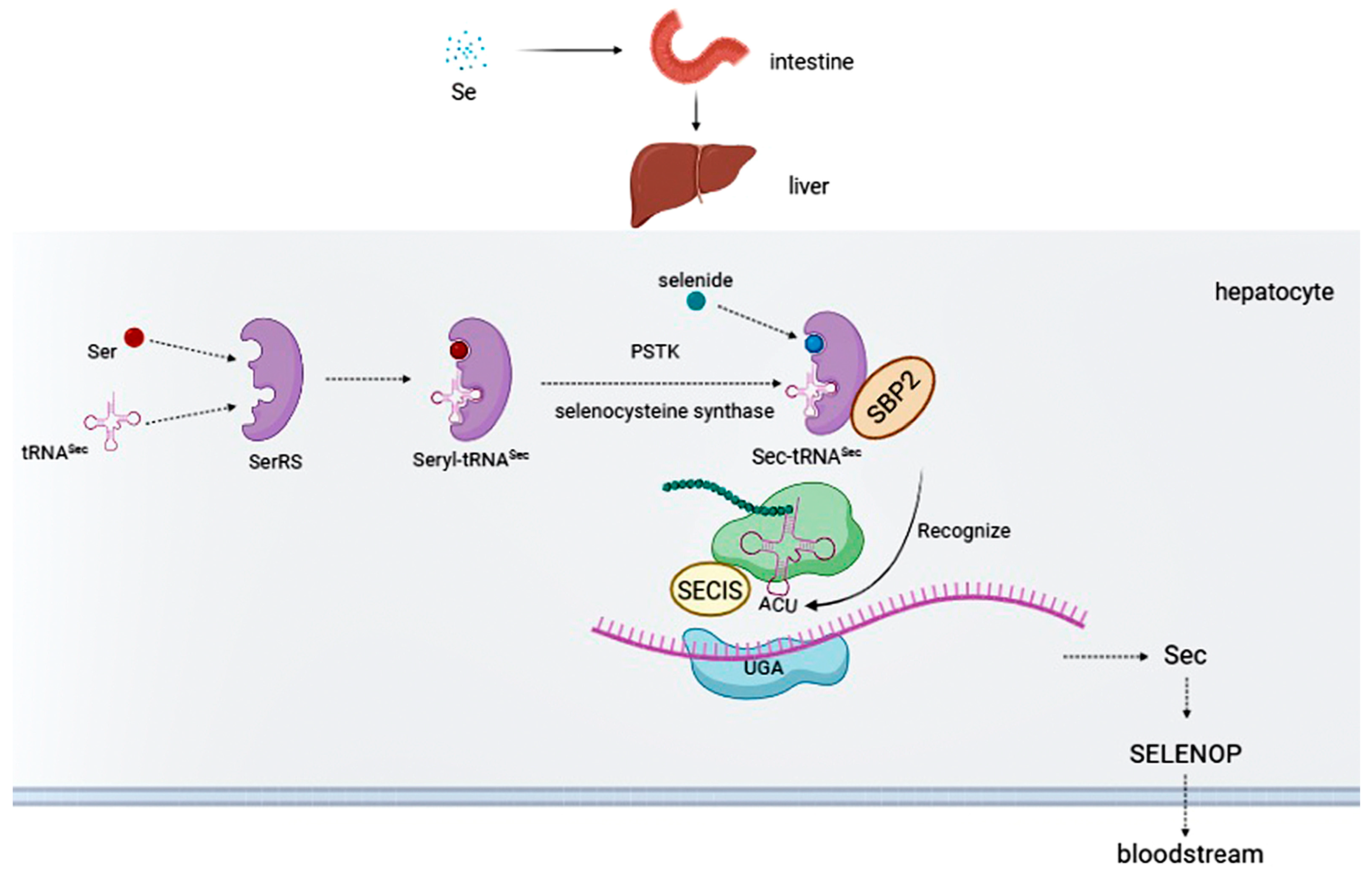
2. Selenium Intake
3. Selenoproteins
4. Health Effects of Selenium and Selenoproteins
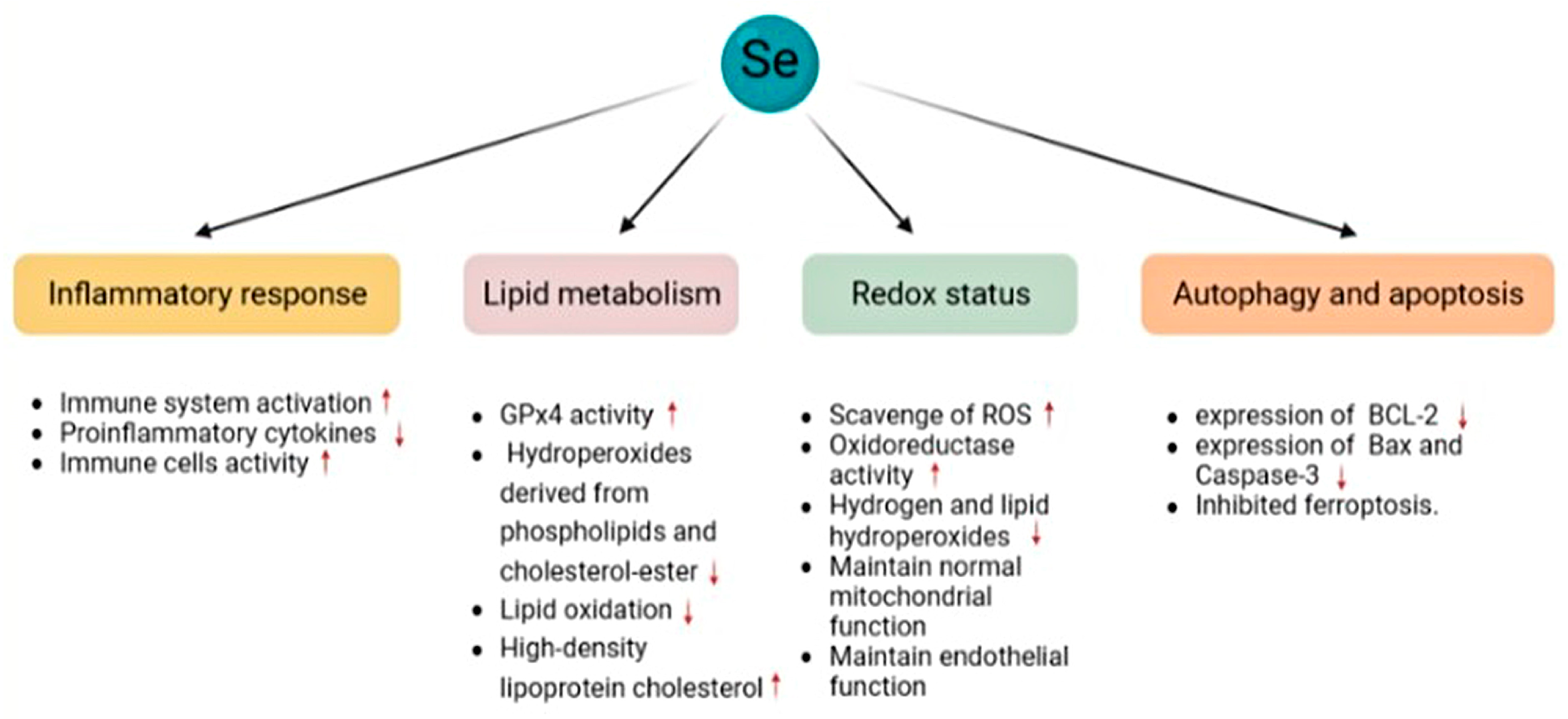
4.1. Oxidative Stress
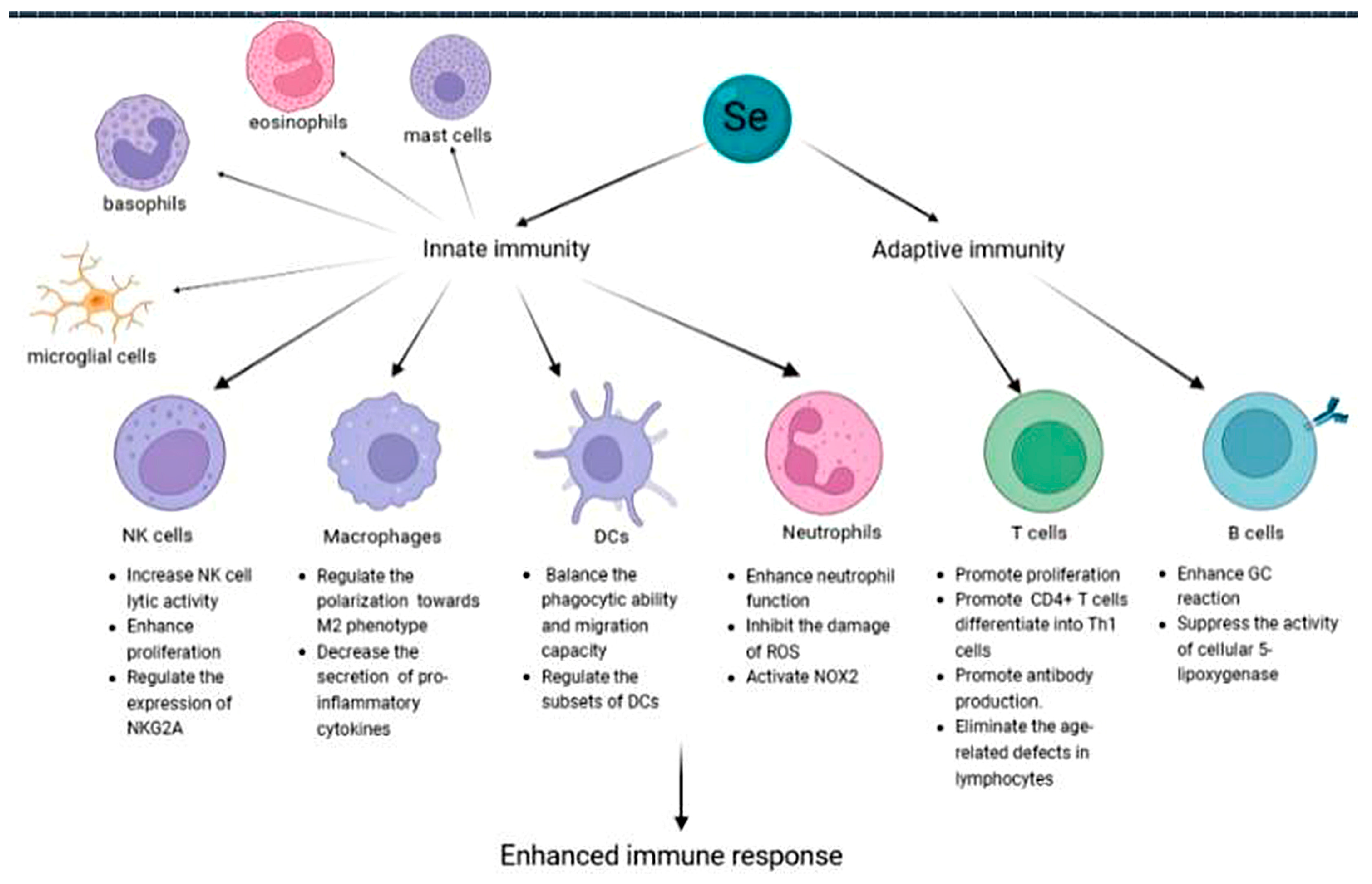
4.2. Immune System
4.2.1. Innate Immunity
4.2.2. Adaptive Immunity
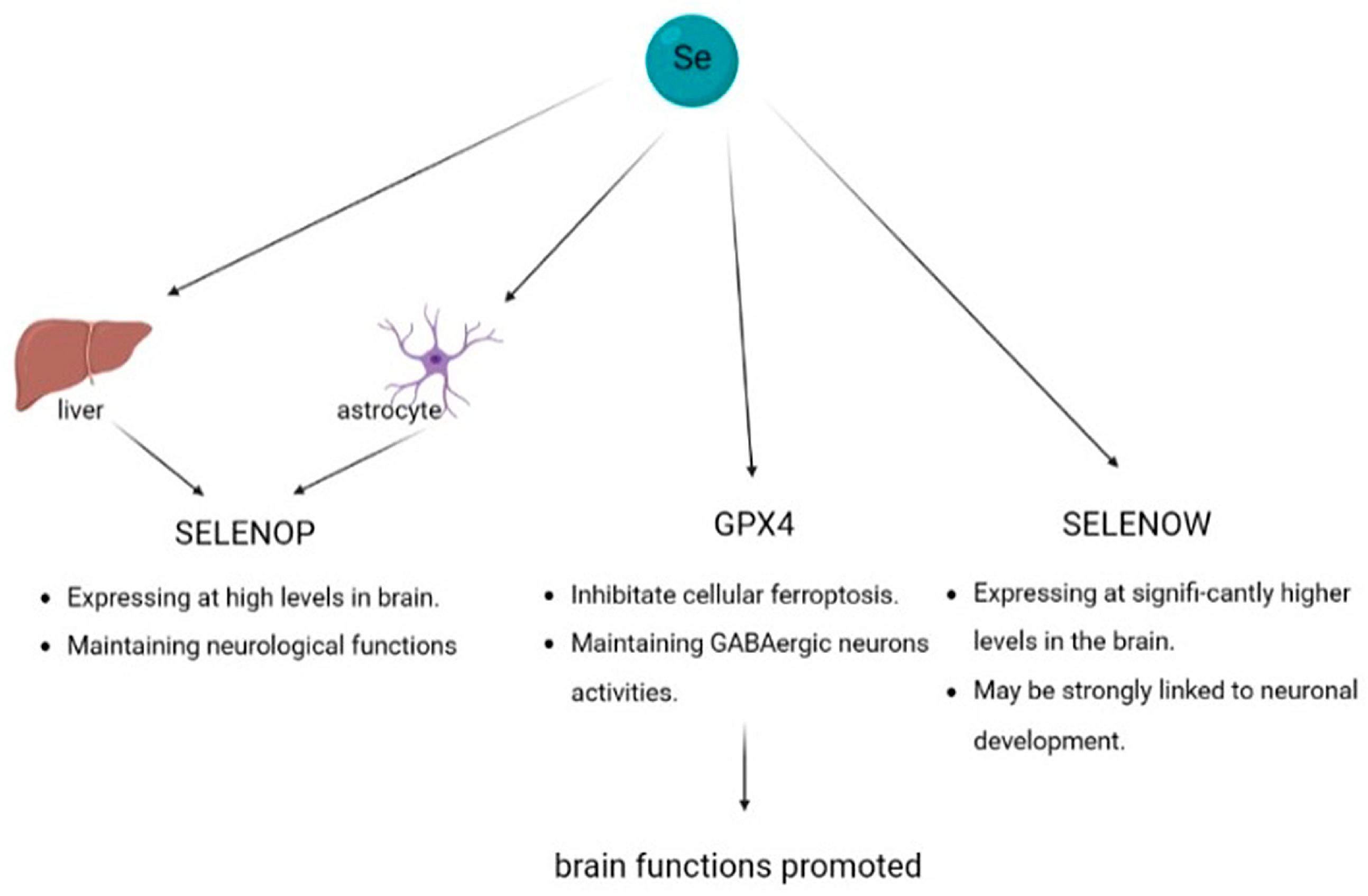
4.3. Brain Function
4.3.1. Selenoprotein P
4.3.2. Glutathione Peroxidase 4
4.3.3. Selenoprotein W
4.4. Cardiovascular System
4.5. Cancer
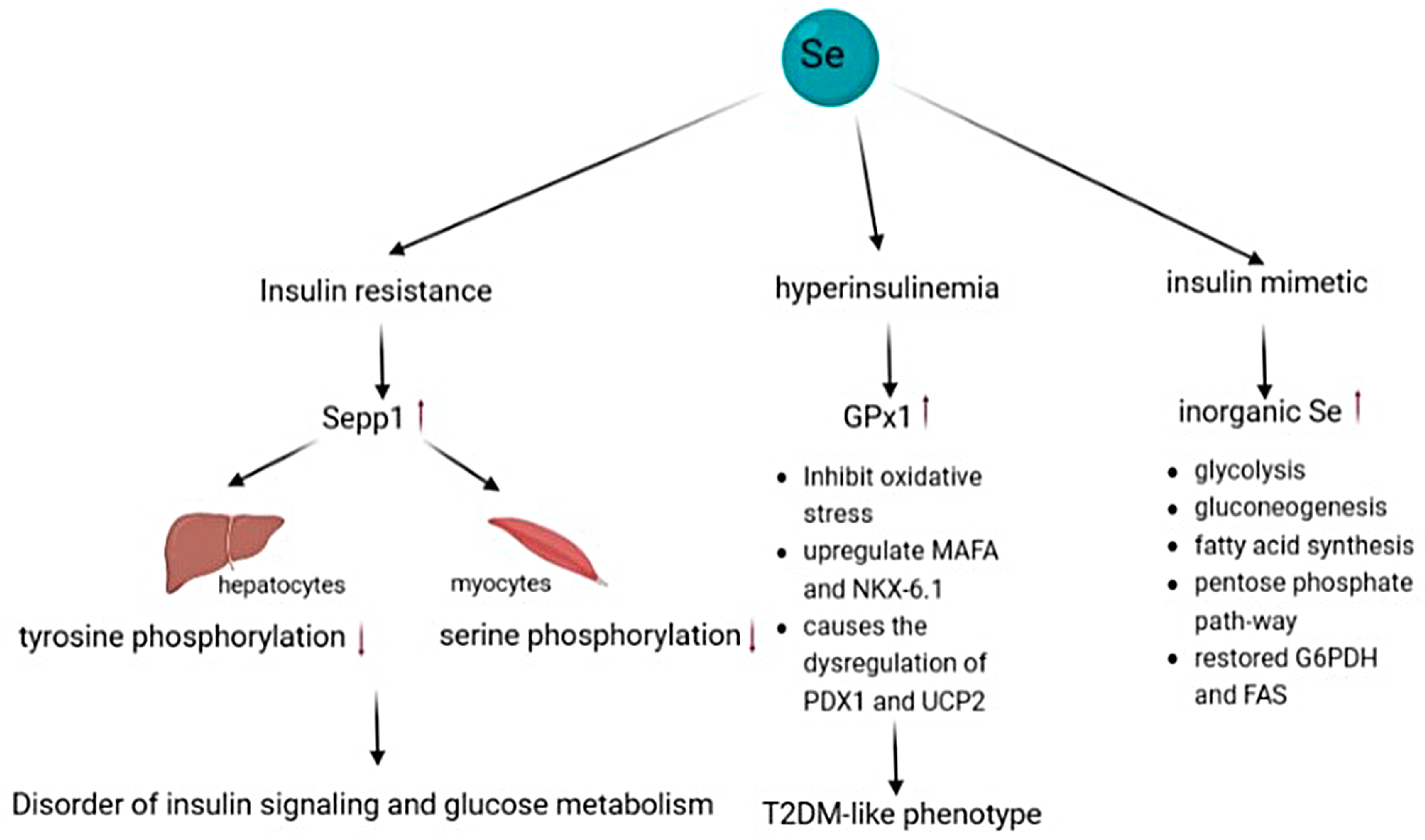
4.6. Type 2 Diabetes
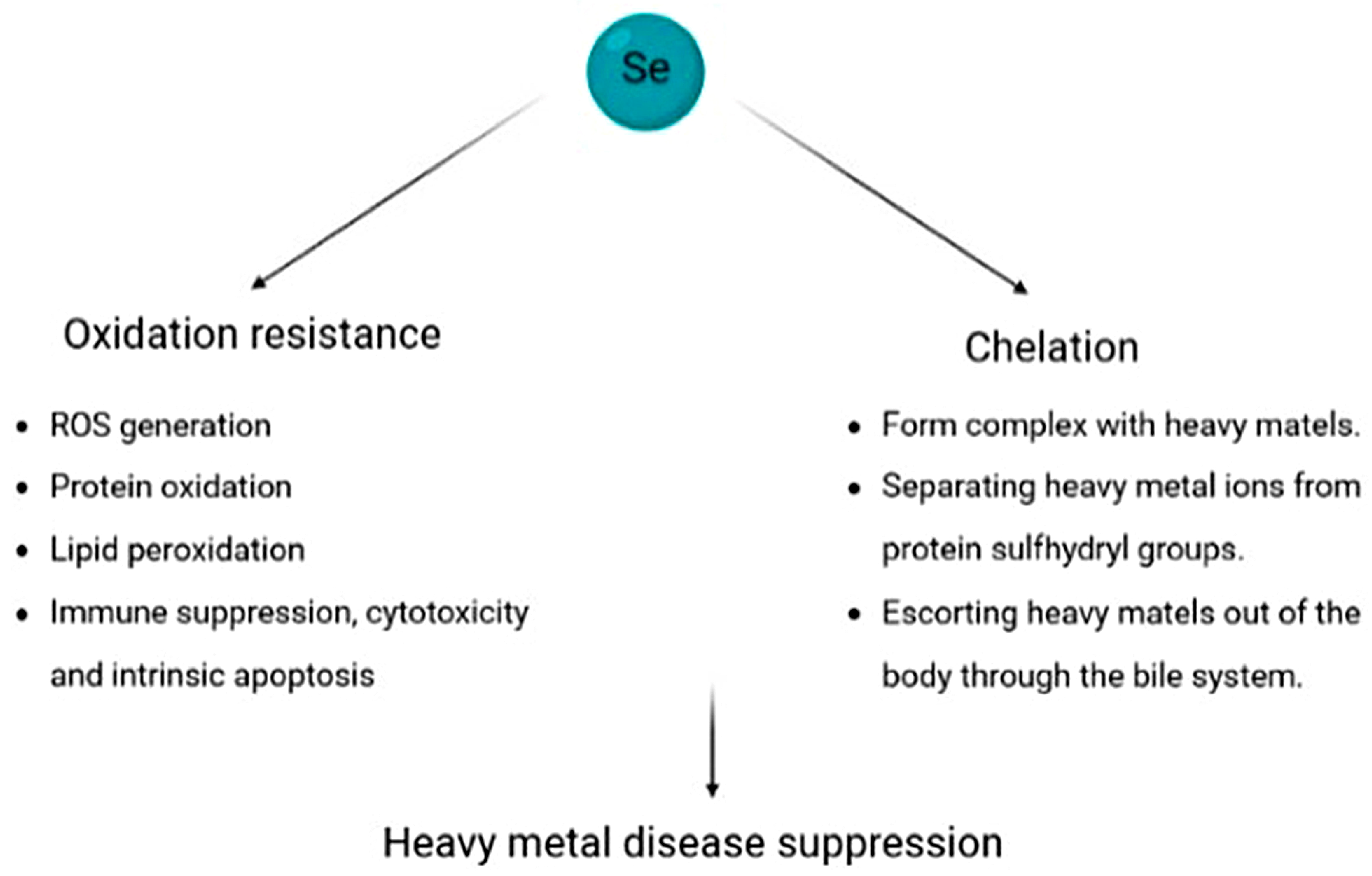
4.7. Heavy Metal-Based Illness
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mertz, W. The Essential Trace Elements. Science 1981, 213, 1332–1338. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; Foltz, C.M. Factor 3 activity of selenium compounds. J. Biol. Chem. 1958, 233, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Stadtman, T.C. Selenocysteine. Annu. Rev. Biochem. 1996, 65, 83–100. [Google Scholar] [CrossRef] [PubMed]
- Arnér, E.S.; Holmgren, A. The thioredoxin system in cancer. Semin. Cancer Biol. 2006, 16, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Lee, J.; Jung, J.; Carlson, B.A.; Chang, M.J.; Chang, C.B.; Kang, S.-B.; Lee, B.C.; Gladyshev, V.N.; Hatfield, D.L.; et al. Selenophosphate synthetase 1 deficiency exacerbates osteoarthritis by dysregulating redox homeostasis. Nat. Commun. 2022, 13, 779. [Google Scholar] [CrossRef]
- Xie, M.; Sun, X.; Li, P.; Shen, X.; Fang, Y. Selenium in cereals: Insight into species of the element from total amount. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2914–2940. [Google Scholar] [CrossRef]
- Alcântara, D.B.; Dionísio, A.P.; Artur, A.G.; Silveira, B.K.; Lopes, A.F.; Guedes, J.A.; Luz, L.R.; Nascimento, R.F.; Lopes, G.S.; Hermsdorff, H.H.; et al. Selenium in Brazil nuts: An overview of agronomical aspects, recent trends in analytical chemistry, and health outcomes. Food Chem. 2022, 372, 131207. [Google Scholar] [CrossRef]
- Yang, H.; Yang, X.; Ning, Z.; Kwon, S.Y.; Li, M.-L.; Tack, F.M.; Kwon, E.E.; Rinklebe, J.; Yin, R. The beneficial and hazardous effects of selenium on the health of the soil-plant-human system: An overview. J. Hazard. Mater. 2022, 422, 126876. [Google Scholar] [CrossRef]
- Pyrzynska, K.; Sentkowska, A. Selenium in plant foods: Speciation analysis, bioavailability, and factors affecting composition. Crit. Rev. Food Sci. Nutr. 2021, 61, 1340–1352. [Google Scholar] [CrossRef]
- Finley, J.W. Bioavailability of selenium from foods. Nutr. Rev. 2006, 64, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Ip, C.; Hayes, C.; Budnick, R.M.; Ganther, H.E. Chemical form of selenium, critical metabolites, and cancer prevention. Cancer Res. 1991, 51, 595–600. [Google Scholar]
- Klimaszewska, M.; Górska, S.; Dawidowski, M.; Podsadni, P.; Turło, J. Biosynthesis of Se-methyl-seleno-l-cysteine in Basidiomycetes fungus Lentinula edodes (Berk.) Pegler. Springerplus 2016, 5, 733. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E. Regulation of Selenium Metabolism and Transport. Annu. Rev. Nutr. 2015, 35, 109–134. [Google Scholar] [CrossRef] [PubMed]
- Adadi, P.; Barakova, N.V.; Muravyov, K.Y.; Krivoshapkina, E.F. Designing selenium functional foods and beverages: A review. Food Res. Int. 2019, 120, 708–725. [Google Scholar] [CrossRef] [PubMed]
- Mistry, H.D.; Pipkin, F.B.; Redman, C.W.; Poston, L. Selenium in reproductive health. Am. J. Obstet. Gynecol. 2012, 206, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Leiter, O.; Zhuo, Z.; Rust, R.; Wasielewska, J.M.; Grönnert, L.; Kowal, S.; Overall, R.W.; Adusumilli, V.S.; Blackmore, D.G.; Southon, A.; et al. Selenium mediates exercise-induced adult neurogenesis and reverses learning deficits induced by hippocampal injury and aging. Cell Metab. 2022, 34, 408–423. [Google Scholar] [CrossRef]
- Gharipour, M.; Ouguerram, K.; Nazih, E.; Salehi, M.; Behmanesh, M.; Roohafza, H.; Hosseini, S.M.; Nezafati, P.; Dianatkhah, M.; Gharipour, A.; et al. Effects of selenium supplementation on expression of SEPP1 in mRNA and protein levels in subjects with and without metabolic syndrome suffering from coronary artery disease: Selenegene study a double-blind randomized controlled trial. J. Cell Biochem. 2018, 119, 8282–8289. [Google Scholar] [CrossRef]
- Hill, K.E.; Wu, S.; Motley, A.K.; Stevenson, T.D.; Winfrey, V.P.; Capecchi, M.R.; Atkins, J.; Burk, R.F. Production of selenoprotein P (Sepp1) by hepatocytes is central to selenium homeostasis. J. Biol. Chem. 2012, 287, 40414–40424. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef]
- Zhang, Y.; Romero, H.; Salinas, G.; Gladyshev, V.N. Dynamic evolution of selenocysteine utilization in bacteria: A balance between selenoprotein loss and evolution of selenocysteine from redox active cysteine residues. Genome. Biol. 2006, 7, R94. [Google Scholar] [CrossRef] [PubMed]
- Castellano, S.; Andrés, A.M.; Bosch, E.; Bayes, M.; Guigó, R.; Clark, A.G. Low exchangeability of selenocysteine, the 21st amino acid, in vertebrate proteins. Mol. Biol. Evol. 2009, 26, 2031–2040. [Google Scholar] [CrossRef]
- Shu, N.; Cheng, Q.; Arnér, E.S.; Davies, M.J. Inhibition and crosslinking of the selenoprotein thioredoxin reductase-1 by p-benzoquinone. Redox Biol. 2020, 28, 101335. [Google Scholar] [CrossRef]
- Zeida, A.; Trujillo, M.; Ferrer-Sueta, G.; Denicola, A.; Estrin, D.A.; Radi, R. Catalysis of Peroxide Reduction by Fast Reacting Protein Thiols. Chem. Rev. 2019, 119, 10829–10855. [Google Scholar] [CrossRef] [PubMed]
- Mobli, M.; Morgenstern, D.; King, G.F.; Alewood, P.F.; Muttenthaler, M. Site-specific pK(a) determination of selenocysteine residues in selenovasopressin by using 77Se NMR spectroscopy. Angew. Chem. Int. Ed. Engl. 2011, 50, 11952–11955. [Google Scholar] [CrossRef] [PubMed]
- Mousa, R.; Dardashti, R.N.; Metanis, N. Selenium and Selenocysteine in Protein Chemistry. Angew. Chem. Int. Ed. Engl. 2017, 56, 15818–15827. [Google Scholar] [CrossRef]
- Sheppard, K.; Yuan, J.; Hohn, M.J.; Jester, B.; Devine, K.M.; Söll, D. From one amino acid to another: tRNA-dependent amino acid biosynthesis. Nucleic Acids. Res. 2008, 36, 1813–1825. [Google Scholar] [CrossRef]
- Arnér, E.S. Common modifications of selenocysteine in selenoproteins. Essays Biochem. 2020, 64, 45–53. [Google Scholar] [CrossRef]
- Commans, S.; Böck, A. Selenocysteine inserting tRNAs: An overview. FEMS Microbiol. Rev. 1999, 23, 335–351. [Google Scholar] [CrossRef]
- Wang, C.; Guo, Y.; Tian, Q.; Jia, Q.; Gao, Y.; Zhang, Q.; Zhou, C.; Xie, W. SerRS-tRNASec complex structures reveal mechanism of the first step in selenocysteine biosynthesis. Nucleic Acids. Res. 2015, 43, 10534–10545. [Google Scholar]
- Fischer, N.; Neumann, P.; Bock, L.V.; Maracci, C.; Wang, Z.; Paleskava, A.; Konevega, A.L.; Schröder, G.F.; Grubmüller, H.; Ficner, R.; et al. The pathway to GTPase activation of elongation factor SelB on the ribosome. Nature 2016, 540, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, E.; Agostini, M.; Mitchell, C.; Schoenmakers, N.; Papp, L.; Rajanayagam, O.; Padidela, R.; Ceron-Gutierrez, L.; Doffinger, R.; Prevosto, C.; et al. Mutations in the selenocysteine insertion sequence-binding protein 2 gene lead to a multisystem selenoprotein deficiency disorder in humans. J. Clin. Investig. 2010, 120, 4220–4235. [Google Scholar] [CrossRef] [PubMed]
- Dumitrescu, A.M.; Liao, X.-H.; Abdullah, M.S.Y.; Lado-Abeal, J.; Majed, F.A.; Moeller, L.; Boran, G.; Schomburg, L.; Weiss, R.E.; Refetoff, S. Mutations in SECISBP2 result in abnormal thyroid hormone metabolism. Nat. Genet. 2005, 37, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Chavatte, L.; Ii, B.A.B.; Driscoll, D.M. Ribosomal protein L30 is a component of the UGA-selenocysteine recoding machinery in eukaryotes. Nat. Struct. Mol. Biol. 2005, 12, 408–416. [Google Scholar] [CrossRef]
- Papp, L.V.; Wang, J.; Kennedy, D.; Boucher, D.; Zhang, Y.; Gladyshev, V.N.; Singh, R.N.; Khanna, K.K. Functional characterization of alternatively spliced human SECISBP2 transcript variants. Nucleic Acids. Res. 2008, 36, 7192–7206. [Google Scholar] [CrossRef]
- Peeler, J.C.; Weerapana, E. Chemical Biology Approaches to Interrogate the Selenoproteome. Acc. Chem. Res. 2019, 52, 2832–2840. [Google Scholar] [CrossRef]
- Stadtman, T.C. Selenium biochemistry. Annu. Rev. Biochem. 1990, 59, 111–127. [Google Scholar] [CrossRef]
- Spallholz, J.E.; Boylan, L.M.; Larsen, H.S. Advances in understanding selenium’s role in the immune system. Ann. N. Y. Acad. Sci. 1990, 587, 123–139. [Google Scholar] [CrossRef]
- Allan, C.B.; Lacourciere, G.M.; Stadtman, T.C. Responsiveness of selenoproteins to dietary selenium. Annu. Rev. Nutr. 1999, 19, 1–16. [Google Scholar] [CrossRef]
- Schomburg, L.; Köhrle, J. On the importance of selenium and iodine metabolism for thyroid hormone biosynthesis and human health. Mol. Nutr. Food Res. 2008, 52, 1235–1246. [Google Scholar] [CrossRef]
- Olson, G.E.; Winfrey, V.P.; Hill, K.E.; Burk, R.F. Megalin mediates selenoprotein P uptake by kidney proximal tubule epithelial cells. J. Biol. Chem. 2008, 283, 6854–6860. [Google Scholar] [CrossRef] [PubMed]
- Curran, J.E.; Jowett, J.B.M.; Elliott, K.S.; Gao, Y.; Gluschenko, K.; Wang, J.; Azim, D.M.A.; Cai, G.; Mahaney, M.C.; Comuzzie, A.G.; et al. Genetic variation in selenoprotein S influences inflammatory response. Nat. Genet. 2005, 37, 1234–1241. [Google Scholar] [CrossRef] [PubMed]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell Mol. Life. Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef] [PubMed]
- Beilstein, M.; Vendeland, S.; Barofsky, E.; Jensen, O.; Whanger, P. Selenoprotein W of rat muscle binds glutathione and an unknown small molecular weight moiety. J. Inorg. Biochem. 1996, 61, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Muntoni, F.; Brockington, M.; Godfrey, C.; Ackroyd, M.; Robb, S.; Manzur, A.; Kinali, M.; Mercuri, E.; Kaluarachchi, M.; Feng, L.; et al. Muscular dystrophies due to defective glycosylation of dystroglycan. Acta Myol. 2007, 26, 129–135. [Google Scholar] [PubMed]
- Zhao, Y.; Wang, H.; Zhou, J.; Shao, Q. Glutathione Peroxidase GPX1 and its Dichotomous Roles in Cancer. Cancers 2022, 14, 2560. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Guo, Y.; Fei, Y.; Zhang, R.; Han, Y.; Lu, S. GPx1 knockdown suppresses chondrogenic differentiation of ATDC5 cells through induction of reductive stress. Acta Biochim. Biophys. Sin. 2017, 49, 110–118. [Google Scholar] [CrossRef]
- Kafeel, S.; Hashim, Z.; Fawwad, A.; Nawab, S.N. Predisposition of SOD1, GPX1, CAT genetic variants and their haplotypes in cataractogenesis of type 2 diabetes mellitus in Pakistan. Acta Diabetol. 2022, 59, 623–632. [Google Scholar] [CrossRef]
- Johnson, L.A.; Phillips, J.A.; Mauer, C.; Edwards, M.; Balldin, V.H.; Hall, J.R.; Barber, R.; Conger, T.L.; Ho, E.J.; O’bryant, S.E. The impact of GPX1 on the association of groundwater selenium and depression: A Project FRONTIER study. BMC. Psychiatry 2013, 13, 7. [Google Scholar] [CrossRef]
- Pei, J.; Fu, W.; Yang, L.; Zhang, Z.; Liu, Y. Oxidative stress is involved in the pathogenesis of Keshan disease (an endemic dilated cardiomyopathy) in China. Oxid. Med. Cell Longev. 2013, 2013, 474203. [Google Scholar] [CrossRef]
- Orhan, H.; Marol, S.; Hepşen, I.F.; Şahin, G. Effects of some probable antioxidants on selenite-induced cataract formation and oxidative stress-related parameters in rats. Toxicology 1999, 139, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zheng, Y.; Wang, C.; Liu, Y. Glutathione depletion induces ferroptosis, autophagy, and premature cell senescence in retinal pigment epithelial cells. Cell Death Dis. 2018, 9, 753. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R.; Kipp, A.P. Physiological functions of GPx2 and its role in inflammation-triggered carcinogenesis. Ann. N. Y. Acad. Sci. 2012, 1259, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Esworthy, R.S.; Yang, L.; Frankel, P.H.; Chu, F.-F. Epithelium-specific glutathione peroxidase, Gpx2, is involved in the prevention of intestinal inflammation in selenium-deficient mice. J. Nutr. 2005, 135, 740–745. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Worley, B.L.; Phaëton, R.; Hempel, N. Extracellular Glutathione Peroxidase GPx3 and its Role in Cancer. Cancers 2020, 12, 2197. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Qin, Y.; Cheng, Z.; Cheng, X.; Wang, R.; Luo, X.; Zhao, Y.; Zhang, D.; Li, G. Gpx3 and Egr1 Are Involved in Regulating the Differentiation Fate of Cardiac Fibroblasts under Pressure Overload. Oxid. Med. Cell Longev. 2022, 2022, 3235250. [Google Scholar] [CrossRef]
- Covington, T.A.; Pilz, P.M.; Mulhern, R.M.; Ngoy, S.; Loscalzo, A.; Liu, J.; Fisch, S.; Grune, J. GPx3 deficiency exacerbates maladaptive right ventricular remodeling in experimental pulmonary artery banding. Am. J. Physiol. Lung. Cell Mol. Physiol. 2023, 324, L550–L556. [Google Scholar] [CrossRef]
- Hangauer, M.J.; Viswanathan, V.S.; Ryan, M.J.; Bole, D.; Eaton, J.K.; Matov, A.; Galeas, J.; Dhruv, H.D.; Berens, M.E.; Schreiber, S.L.; et al. Contribution of ferroptosis and GPX4’s dual functions to osteoarthritis progression. EBioMedicine 2022, 76, 103847. [Google Scholar]
- Hangauer, M.J.; Viswanathan, V.S.; Ryan, M.J.; Bole, D.; Eaton, J.K.; Matov, A.; Galeas, J.; Dhruv, H.D.; Berens, M.E.; Schreiber, S.L.; et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition. Nature 2017, 551, 247–250. [Google Scholar] [CrossRef]
- Wang, Y.; Yan, S.; Liu, X.; Deng, F.; Wang, P.; Yang, L.; Hu, L.; Huang, K.; He, J. PRMT4 promotes ferroptosis to aggravate doxorubicin-induced cardiomyopathy via inhibition of the Nrf2/GPX4 pathway. Cell Death Differ. 2022, 29, 1982–1995. [Google Scholar] [CrossRef]
- Zhang, J.; Bi, J.; Ren, Y.; Du, Z.; Li, T.; Wang, T.; Zhang, L.; Wang, M.; Wei, S.; Lv, Y.; et al. Involvement of GPX4 in irisin’s protection against ischemia reperfusion-induced acute kidney injury. J. Cell Physiol. 2021, 236, 931–945. [Google Scholar] [CrossRef]
- Reichert, C.O.; de Freitas, F.A.; Sampaio-Silva, J.; Rokita-Rosa, L.; Barros, P.d.L.; Levy, D.; Bydlowski, S.P. Ferroptosis Mechanisms Involved in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8765. [Google Scholar] [CrossRef] [PubMed]
- Shema, R.; Kulicke, R.; Cowley, G.S.; Stein, R.; Root, D.E.; Heiman, M. Synthetic lethal screening in the mammalian central nervous system identifies Gpx6 as a modulator of Huntington’s disease. Proc. Natl. Acad. Sci. USA 2015, 112, 268–272. [Google Scholar] [CrossRef]
- Lin, W.; Tang, Y.; Zhang, M.; Liang, B.; Wang, M.; Zha, L.; Yu, Z. Integrated Bioinformatic Analysis Reveals TXNRD1 as a Novel Biomarker and Potential Therapeutic Target in Idiopathic Pulmonary Arterial Hypertension. Front. Med. 2022, 9, 894584. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Meng, W.; Zeng, X.; Zhao, H.; Liu, W.; Zhang, T. TXNRD1 Is an Unfavorable Prognostic Factor for Patients with Hepatocellular Carcinoma. Biomed. Res. Int. 2017, 2017, 4698167. [Google Scholar] [CrossRef]
- Liang, J.; Wang, S.; Hu, J.; Hong, X.; Zhu, M.; Liu, X.; Alswadeh, M.; Mo, F.; Dai, M. Targeted inhibition of TXNRD1 prevents cartilage extracellular matrix degeneration by activating Nrf2 pathway in osteoarthritis. Biochem. Biophys. Res. Commun. 2022, 635, 267–276. [Google Scholar] [CrossRef]
- Kudin, A.P.; Baron, G.; Zsurka, G.; Hampel, K.G.; Elger, C.E.; Grote, A.; Weber, Y.; Lerche, H.; Thiele, H.; Nürnberg, P.; et al. Homozygous mutation in TXNRD1 is associated with genetic generalized epilepsy. Free Radic. Biol. Med. 2017, 106, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J.N.C.; ANZRAG Consortium; Loomis, S.J.; Kang, J.H.; Allingham, R.R.; Gharahkhani, P.; Khor, C.C.; Burdon, K.P.; Aschard, H.; Chasman, D.I.; et al. Genome-wide association analysis identifies TXNRD2, ATXN2 and FOXC1 as susceptibility loci for primary open-angle glaucoma. Nat. Genet. 2016, 48, 189–194. [Google Scholar] [CrossRef]
- Sibbing, D.; Pfeufer, A.; Perisic, T.; Mannes, A.M.; Fritz-Wolf, K.; Unwin, S.; Sinner, M.F.; Gieger, C.; Gloeckner, C.J.; Wichmann, H.-E.; et al. Mutations in the mitochondrial thioredoxin reductase gene TXNRD2 cause dilated cardiomyopathy. Eur. Heart J. 2011, 32, 1121–1133. [Google Scholar] [CrossRef]
- Kariž, S.; Mankoč, S.; Petrovič, D. Association of thioredoxin reductase 2 (TXNRD2) gene polymorphisms with myocardial infarction in Slovene patients with type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2015, 108, 323–328. [Google Scholar] [CrossRef]
- Gencheva, R.; Arnér, E.S.J. Thioredoxin Reductase Inhibition for Cancer Therapy. Annu. Rev. Pharmacol. Toxicol. 2022, 62, 177–196. [Google Scholar] [CrossRef] [PubMed]
- Aboobakar, I.F.; Kinzy, T.G.; Zhao, Y.; Fan, B.; Pasquale, L.R.; Qassim, A.; Kolovos, A.; Schmidt, J.M.; Craig, J.E.; Bailey, J.N.C.; et al. Mitochondrial TXNRD2 and ME3 genetic risk scores are associated with specific primary open-angle glaucoma phenotypes. Ophthalmology 2023, in press. [Google Scholar] [CrossRef] [PubMed]
- Dou, Q.; Turanov, A.A.; Mariotti, M.; Hwang, J.Y.; Lee, S.-G.; Paulo, J.A.; Yim, S.H.; Gygi, S.P.; Chung, J.-J.; Gladyshev, V.N. Selenoprotein TXNRD3 supports male fertility via the redox regulation of spermatogenesis. J. Biol. Chem. 2022, 298, 102183. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Du, P.; Zhu, Y.; Zhang, X.; Cai, J.; Zhang, Z. Thioredoxin reductase 3 suppression promotes colitis and carcinogenesis via activating pyroptosis and necrosis. Cell Mol. Life Sci. 2022, 79, 106. [Google Scholar] [CrossRef]
- França, M.M.; German, A.; Fernandes, G.W.; Liao, X.-H.; Bianco, A.C.; Refetoff, S.; Dumitrescu, A.M. Dumitrescu. Human Type 1 Iodothyronine Deiodinase (DIO1) Mutations Cause Abnormal Thyroid Hormone Metabolism. Thyroid 2021, 31, 202–207. [Google Scholar] [CrossRef]
- Bruinstroop, E.; Zhou, J.; Tripathi, M.; Yau, W.W.; Boelen, A.; Singh, B.K.; Yen, P.M. Early induction of hepatic deiodinase type 1 inhibits hepatosteatosis during NAFLD progression. Mol. Metab. 2021, 53, 101266. [Google Scholar] [CrossRef]
- Poplawski, P.; Rybicka, B.; Boguslawska, J.; Rodzik, K.; Visser, T.J.; Nauman, A.; Piekielko-Witkowska, A. Induction of type 1 iodothyronine deiodinase expression inhibits proliferation and migration of renal cancer cells. Mol. Cell Endocrinol. 2017, 442, 58–67. [Google Scholar] [CrossRef]
- Bomer, N.; Hollander, W.D.; Ramos, Y.F.M.; Bos, S.D.; van der Breggen, R.; Lakenberg, N.; Pepers, B.A.; van Eeden, A.E.; Darvishan, A.; Tobi, E.W.; et al. Underlying molecular mechanisms of DIO2 susceptibility in symptomatic osteoarthritis. Ann. Rheum. Dis. 2015, 74, 1571–1579. [Google Scholar] [CrossRef]
- Bradley, D.; Liu, J.; Blaszczak, A.; Wright, V.; Jalilvand, A.; Needleman, B.; Noria, S.; Renton, D.; Hsueh, W. Adipocyte DIO2 Expression Increases in Human Obesity but Is Not Related to Systemic Insulin Sensitivity. J. Diabetes Res. 2018, 2018, 2464652. [Google Scholar] [CrossRef]
- Guo, T.-W.; Zhang, F.-C.; Yang, M.-S.; Gao, X.-C.; Bian, L.; Duan, S.-W.; Zheng, Z.-J.; Gao, J.-J.; Wang, H.; Li, R.-L.; et al. Positive association of the DIO2 (deiodinase type 2) gene with mental retardation in the iodine-deficient areas of China. J. Med. Genet. 2004, 41, 585–590. [Google Scholar] [CrossRef]
- Meulenbelt, I.; Bos, S.D.; Chapman, K.; van der Breggen, R.; Houwing-Duistermaat, J.J.; Kremer, D.; Kloppenburg, M.; Carr, A.; Tsezou, A.; González, A.; et al. Meta-analyses of genes modulating intracellular T3 bio-availability reveal a possible role for the DIO3 gene in osteoarthritis susceptibility. Ann. Rheum. Dis. 2011, 70, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Brent, G.A. Thyroid hormone and the brain: Mechanisms of action in development and role in protection and promotion of recovery after brain injury. Pharmacol. Ther. 2018, 186, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Vidart, J.; Axelrud, L.; Braun, A.C.; Marschner, R.A.; Wajner, S.M. Relationship among Low T3 Levels, Type 3 Deiodinase, Oxidative Stress, and Mortality in Sepsis and Septic Shock: Defining Patient Outcomes. Int. J. Mol. Sci. 2023, 24, 3935. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Li, H.; Meng, F.; Sun, X.; Feng, X.; Chen, J.; Li, L.; Liu, J. Methionine Sulfoxide Reductase B1 Regulates Hepatocellular Carcinoma Cell Proliferation and Invasion via the Mitogen-Activated Protein Kinase Pathway and Epithelial-Mesenchymal Transition. Oxid. Med. Cell Longev. 2018, 2018, 5287971. [Google Scholar] [CrossRef]
- Lee, H.-J.; Park, J.S.; Yoo, H.J.; Lee, H.M.; Lee, B.C.; Kim, J.H. The Selenoprotein MsrB1 Instructs Dendritic Cells to Induce T-Helper 1 Immune Responses. Antioxidants 2020, 9, 1021. [Google Scholar] [CrossRef]
- Carlisle, A.E.; Lee, N.; Matthew-Onabanjo, A.N.; Spears, M.E.; Park, S.J.; Youkana, D.; Doshi, M.B.; Peppers, A.; Li, R.; Joseph, A.B.; et al. Selenium detoxification is required for cancer-cell survival. Nat. Metab. 2020, 2, 603–611. [Google Scholar] [CrossRef]
- Flowers, B.; Bochnacka, O.; Poles, A.; Diamond, A.M.; Kastrati, I. Distinct Roles of SELENOF in Different Human Cancers. Biomolecules 2023, 13, 486. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Y.; Zhou, J.; Liu, H. Selenoprotein F Knockout Caused Glucose Metabolism Disorder in Young Mice by Disrupting Redox Homeostasis. Antioxidants 2022, 11, 2105. [Google Scholar] [CrossRef]
- Bertz, M.; Kühn, K.; Koeberle, S.C.; Müller, M.F.; Hoelzer, D.; Thies, K.; Deubel, S.; Thierbach, R.; Kipp, A.P. Selenoprotein H controls cell cycle progression and proliferation of human colorectal cancer cells. Free Radic. Biol. Med. 2018, 127, 98–107. [Google Scholar] [CrossRef]
- Sarma, A.S.; Siddardha, B.; Pragna Lakshmi, T.; Ranganath, P.; Dalal, A. A novel homozygous synonymous splicing variant in SELENOI gene causes spastic paraplegia 81. J. Gene Med. 2023, 2023, e3501. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Song, G.L. Roles of Selenoproteins in Brain Function and the Potential Mechanism of Selenium in Alzheimer’s Disease. Front. Neurosci. 2021, 15, 646518. [Google Scholar] [CrossRef] [PubMed]
- Abdurahman, A.; Li, Y.; Jia, S.-Z.; Xu, X.-W.; Lin, S.-J.; Ouyang, P.; He, Z.J.; Zhang, Z.-H.; Liu, Q.; Xu, Y.; et al. Knockdown of the SELENOK gene induces ferroptosis in cervical cancer cells. Metallomics 2023, 15, mfad019. [Google Scholar] [CrossRef] [PubMed]
- Rogachev, V.V.; Goltyaev, M.V.; Varlamova, E.G.; Turovsky, E.A. Turovsky. Molecular Mechanisms of the Cytotoxic Effect of Recombinant Selenoprotein SELENOM on Human Glioblastoma Cells. Int. J. Mol. Sci. 2023, 24, 6469. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Huang, J.; Yang, J.; Chen, X.; Zhang, H.; Zhu, Y.; Liu, Q.; Zhang, Z. The protective effect of selenoprotein M on non-alcoholic fatty liver disease: The role of the AMPKα1-MFN2 pathway and Parkin mitophagy. Cell Mol. Life Sci. 2022, 79, 354. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chen, C.; Ouyang, P.; Cai, Z.; Liu, X.; Abdurahman, A.; Peng, J.; Li, Y.; Zhang, Z.; Song, G.-L. SELENOM Knockout Induces Synaptic Deficits and Cognitive Dysfunction by Influencing Brain Glucose Metabolism. J. Agric. Food Chem. 2023, 71, 1607–1619. [Google Scholar] [CrossRef]
- Pozzer, D.; Varone, E.; Chernorudskiy, A.; Schiarea, S.; Missiroli, S.; Giorgi, C.; Pinton, P.; Canato, M.; Germinario, E.; Nogara, L.; et al. A maladaptive ER stress response triggers dysfunction in highly active muscles of mice with SELENON loss. Redox Biol. 2019, 20, 354–366. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, P.; Lv, H.-J.; Wu, Y.; Liu, S.; Deng, X.; Shi, B.; Fu, J. Comprehensive Analysis of Expression and Prognostic Value of Selenoprotein Genes in Thyroid Cancer. Genet. Test. Mol. Biomarkers 2022, 26, 159–173. [Google Scholar] [CrossRef]
- Wang, C.; Yu, Q.; Song, T.; Wang, Z.; Song, L.; Yang, Y.; Shao, J.; Li, J.; Ni, Y.; Chao, N.; et al. The heterogeneous immune landscape between lung adenocarcinoma and squamous carcinoma revealed by single-cell RNA sequencing. Signal. Transduct. Target. Ther. 2022, 7, 289. [Google Scholar] [CrossRef]
- Short, S.P.; Pilat, J.M.; Barrett, C.W.; Reddy, V.K.; Haberman, Y.; Hendren, J.R.; Marsh, B.J.; Keating, C.E.; Motley, A.K.; Hill, K.E.; et al. Colonic Epithelial-Derived Selenoprotein P Is the Source for Antioxidant-Mediated Protection in Colitis-Associated Cancer. Gastroenterology 2021, 160, 1694–1708. [Google Scholar] [CrossRef]
- Ekoue, D.N.; Ansong, E.; Liu, L.; Macias, V.; Deaton, R.; Lacher, C.; Picklo, M.; Nonn, L.; Gann, P.H.; Kajdacsy-Balla, A.; et al. Correlations of SELENOF and SELENOP genotypes with serum selenium levels and prostate cancer. Prostate 2018, 78, 279–288. [Google Scholar] [CrossRef]
- Schweizer, U.; Wirth, E.K.; Klopstock, T.; Hölter, S.M.; Becker, L.; Moskovitz, J.; Grune, T.; Fuchs, H.; Gailus-Durner, V.; de Angelis, M.H.; et al. Seizures, ataxia and parvalbumin-expressing interneurons respond to selenium supply in Selenop-deficient mice. Redox Biol. 2022, 57, 102490. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Orho-Melander, M.; Struck, J.; Bergmann, A.; Melander, O. Selenoprotein-P Deficiency Predicts Cardiovascular Disease and Death. Nutrients 2019, 11, 1852. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.R.; Durães, C.; Ziros, P.G.; Pestana, A.; Esteves, C.; Neves, C.; Carvalho, D.; Bongiovanni, M.; Renaud, C.O.; Chartoumpekis, D.V.; et al. Interaction of Genetic Variations in NFE2L2 and SELENOS Modulates the Risk of Hashimoto’s Thyroiditis. Thyroid 2019, 29, 1302–1315. [Google Scholar] [CrossRef] [PubMed]
- Strauss, E.; Tomczak, J.; Staniszewski, R.; Oszkinis, G. Associations and interactions between variants in selenoprotein genes, selenoprotein levels and the development of abdominal aortic aneurysm, peripheral arterial disease, and heart failure. PLoS ONE 2018, 13, e0203350. [Google Scholar] [CrossRef]
- Varlamova, E.G.; Goltyaev, M.V.; Turovsky, E.A. The Role of Selenoproteins SELENOM and SELENOT in the Regulation of Apoptosis, ER Stress, and Calcium Homeostasis in the A-172 Human Glioblastoma Cell Line. Biology 2022, 11, 811. [Google Scholar] [CrossRef]
- Alsharif, I.; Boukhzar, L.; Lefranc, B.; Godefroy, D.; Aury-Landas, J.; Rego, J.D.; Rego, J.D.; Naudet, F.; Arabo, A.; Chagraoui, A.; et al. Cell-penetrating, antioxidant SELENOT mimetic protects dopaminergic neurons and ameliorates motor dysfunction in Parkinson’s disease animal models. Redox Biol. 2021, 40, 101839. [Google Scholar] [CrossRef]
- Rocca, C.; Pasqua, T.; Boukhzar, L.; Anouar, Y.; Angelone, T. Progress in the emerging role of selenoproteins in cardiovascular disease: Focus on endoplasmic reticulum-resident selenoproteins. Cell Mol. Life. Sci. 2019, 76, 3969–3985. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.; Kim, J.M.; Kim, M.Y.; Kim, J.-R.; Lee, H.-W.; Chung, Y.W.; Shin, H.-I.; Kim, T.; Park, E.-S.; et al. Selenoprotein W ensures physiological bone remodeling by preventing hyperactivity of osteoclasts. Nat. Commun. 2021, 12, 2258. [Google Scholar] [CrossRef]
- Liao, C.; Hardison, R.C.; Kennett, M.J.; Carlson, B.A.; Paulson, R.F.; Prabhu, K.S. Selenoproteins regulate stress erythroid progenitors and spleen microenvironment during stress erythropoiesis. Blood 2018, 131, 2568–2580. [Google Scholar] [CrossRef]
- Shengyu, C.; Yinhua, L.; Yuanhong, L.; Jinbo, Z.; Can, F.; Hao, X.; Changjiang, Z. Selenium alleviates heart remodeling through Sirt1/AKT/GSK-3β pathway. Int. Immunopharmacol. 2022, 111, 109158. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, B.; Wu, P.; Chu, Y.; Gui, S.; Zheng, Y.; Chen, X. Dietary Selenium Alleviated Mouse Liver Oxidative Stress and NAFLD Induced by Obesity by Regulating the KEAP1/NRF2 Pathway. Antioxidants 2022, 11, 349. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, Q.; Zhang, J.; Song, M.; Shao, B.; Han, Y.; Yang, X.; Li, Y. The Protective Effect of Selenium on T-2-Induced Nephrotoxicity Is Related to the Inhibition of ROS-Mediated Apoptosis in Mice Kidney. Biol. Trace Elem. Res. 2022, 200, 206–216. [Google Scholar] [CrossRef] [PubMed]
- Marcocci, C.; Leo, M.; Altea, M.A. Oxidative stress in graves’ disease. Eur. Thyroid. J. 2012, 1, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Maraldi, T.; Riccio, M.; Zambonin, L.; Vinceti, M.; De Pol, A.; Hakim, G. Low levels of selenium compounds are selectively toxic for a human neuron cell line through ROS/RNS increase and apoptotic process activation. Neurotoxicology 2011, 32, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Tosatto, S.C.; Bosello, V.; Fogolari, F.; Mauri, P.; Roveri, A.; Toppo, S.; Flohé, L.; Ursini, F.; Maiorino, M. The catalytic site of glutathione peroxidases. Antioxid. Redox Signal. 2008, 10, 1515–1526. [Google Scholar] [CrossRef] [PubMed]
- Masuda, R.; Kimura, R.; Karasaki, T.; Sase, S.; Goto, K. Modeling the Catalytic Cycle of Glutathione Peroxidase by Nuclear Magnetic Resonance Spectroscopic Analysis of Selenocysteine Selenenic Acids. J. Am. Chem. Soc. 2021, 143, 6345–6350. [Google Scholar] [CrossRef]
- Chu, F.; Doroshow, J.; Esworthy, R. Expression, characterization, and tissue distribution of a new cellular selenium-dependent glutathione peroxidase, GSHPx-GI. J. Biol. Chem. 1993, 268, 2571–2576. [Google Scholar] [CrossRef]
- Wingler, K.; Böcher, M.; Flohé, L.; Kollmus, H.; Brigelius-Flohé, R. mRNA stability and selenocysteine insertion sequence efficiency rank gastrointestinal glutathione peroxidase high in the hierarchy of selenoproteins. Eur. J. Biochem. 1999, 259, 149–157. [Google Scholar] [CrossRef]
- Björnstedt, M.; Xue, J.; Huang, W.; Akesson, B.; Holmgren, A. The thioredoxin and glutaredoxin systems are efficient electron donors to human plasma glutathione peroxidase. J. Biol. Chem. 1994, 269, 29382–29384. [Google Scholar] [CrossRef]
- Schwarz, M.; Löser, A.; Cheng, Q.; Wichmann-Costaganna, M.; Schädel, P.; Werz, O.; Arnér, E.S.; Kipp, A.P. Side-by-side comparison of recombinant human glutathione peroxidases identifies overlapping substrate specificities for soluble hydroperoxides. Redox Biol. 2023, 59, 102593. [Google Scholar] [CrossRef]
- Chi, Q.; Zhang, Q.; Lu, Y.; Zhang, Y.; Xu, S.; Li, S. Roles of selenoprotein S in reactive oxygen species-dependent neutrophil extracellular trap formation induced by selenium-deficient arteritis. Redox Biol. 2021, 44, 102003. [Google Scholar] [CrossRef]
- Serfass, R.E.; Ganther, H.E. Defective microbicidal activity in glutathione peroxidase-deficient neutrophils of selenium-deficient rats. Nature 1975, 255, 640–641. [Google Scholar] [CrossRef] [PubMed]
- Pagmantidis, V.; Méplan, C.; van Schothorst, E.M.; Keijer, J.; Hesketh, J.E. Supplementation of healthy volunteers with nutritionally relevant amounts of selenium increases the expression of lymphocyte protein biosynthesis genes. Am. J. Clin. Nutr. 2008, 87, 181–189. [Google Scholar] [CrossRef]
- Pan, S.; Li, T.; Tan, Y.; Xu, H. Selenium-containing nanoparticles synergistically enhance Pemetrexed&NK cell-based chemoimmunotherapy. Biomaterials 2022, 280, 121321. [Google Scholar] [PubMed]
- Gong, D.; Sun, K.; Yin, K.; Wang, X. Selenium mitigates the inhibitory effect of TBBPA on NETs release by regulating ROS/MAPK pathways-induced carp neutrophil apoptosis and necroptosis. Fish Shellfish Immunol. 2023, 132, 108501. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-J.; Mao, X.-T.; Li, Y.-Y.; Liu, D.-D.; Fan, K.-Q.; Liu, R.-B.; Wu, T.-T.; Wang, H.-L.; Zhang, Y.; Yang, B.; et al. Multiomics analyses reveal a critical role of selenium in controlling T cell differentiation in Crohn’s disease. Immunity 2021, 54, 1728–1744.e7. [Google Scholar] [CrossRef]
- Kiremidjian-Schumacher, L.; Roy, M.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Supplementation with selenium and human immune cell functions. II. Effect on cytotoxic lymphocytes and natural killer cells. Biol. Trace Elem. Res. 1994, 41, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Enqvist, M.; Nilsonne, G.; Hammarfjord, O.; Wallin, R.P.; Björkström, N.K.; Björnstedt, M.; Hjerpe, A.; Ljunggren, H.G.; Dobra, K.; Malmberg, K.J.; et al. Selenite induces posttranscriptional blockade of HLA-E expression and sensitizes tumor cells to CD94/NKG2A-positive NK cells. J. Immunol. 2011, 187, 3546–3554. [Google Scholar] [CrossRef]
- Kiremidjian-Schumacher, L.; Roy, M.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Supplementation with selenium and human immune cell functions. I. Effect on lymphocyte proliferation and interleukin 2 receptor expression. Biol. Trace Elem. Res. 1994, 41, 103–114. [Google Scholar] [CrossRef]
- Nelson, S.M.; Lei, X.; Prabhu, K.S. Selenium levels affect the IL-4-induced expression of alternative activation markers in murine macrophages. J. Nutr. 2011, 141, 1754–1761. [Google Scholar] [CrossRef]
- de Toledo, J.; Fraga-Silva, T.F.; Borim, P.A.; de Oliveira, L.R.; Oliveira, E.D.; Périco, L.L.; Hiruma-Lima, C.A.; de Souza, A.A.L.; de Oliveira, C.A.F.; Padilha, P.M.; et al. Organic Selenium Reaches the Central Nervous System and Downmodulates Local Inflammation: A Complementary Therapy for Multiple Sclerosis? Front. Immunol. 2020, 11, 571844. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive Oxygen Species and Neutrophil Function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef] [PubMed]
- Boyne, R.; Arthur, J.R. The response of selenium-deficient mice to Candida albicans infection. J. Nutr. 1986, 116, 816–822. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.L. The etiologies, pathophysiology, and alternative/complementary treatment of asthma. Altern. Med. Rev. 2001, 6, 20–47. [Google Scholar]
- Gholami, M.; Zendedel, A.; Khayat, Z.K.; Ghanad, K.; Nazari, A.; Pirhadi, A. Selenium effect on ischemia-reperfusion injury of gastrocnemius muscle in adult rats. Biol. Trace Elem. Res. 2015, 164, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Zendehdel, A.; Gharibi, Z.; Anbari, K.; Abbaszadeh, A.; Khayat, Z.K.; Khorramabadi, R.M.; Soleymaninejad, M.; Gholami, M. Ameliorate Peripheral Nerve Ischemic-Reperfusion Injury via Decreased TNF-α. Biol. Trace Elem. Res. 2017, 176, 328–337. [Google Scholar] [CrossRef]
- Safaralizadeh, R.; Nourizadeh, M.; Zare, A.; Kardar, G.A.; Pourpak, Z. Influence of selenium on mast cell mediator release. Biol. Trace Elem. Res. 2013, 154, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.; Chow, C. Induction of eosinophilic enteritis and eosinophilia in rats by vitamin E and selenium deficiency. Exp. Mol. Pathol. 1988, 48, 182–192. [Google Scholar] [CrossRef]
- Weitzel, F.; Wendel, A. Selenoenzymes regulate the activity of leukocyte 5-lipoxygenase via the peroxide tone. J. Biol. Chem. 1993, 268, 6288–6292. [Google Scholar] [CrossRef]
- Dalla Puppa, L.; Savaskan, N.E.; Bräuer, A.U.; Behne, D.; Kyriakopoulos, A. The role of selenite on microglial migration. Ann. N. Y. Acad. Sci. 2007, 1096, 179–183. [Google Scholar] [CrossRef]
- Branco, V.; Coppo, L.; Aschner, M.; Carvalho, C. N-Acetylcysteine or Sodium Selenite Prevent the p38-Mediated Production of Proinflammatory Cytokines by Microglia during Exposure to Mercury (II). Toxics 2022, 10, 433. [Google Scholar] [CrossRef] [PubMed]
- Shrimali, R.K.; Irons, R.D.; Carlson, B.A.; Sano, Y.; Gladyshev, V.N.; Park, J.M.; Hatfield, D.L. Hatfield. Selenoproteins mediate T cell immunity through an antioxidant mechanism. J. Biol. Chem. 2008, 283, 20181–20185. [Google Scholar] [CrossRef] [PubMed]
- Smith-Garvin, J.E.; Koretzky, G.A.; Jordan, M.S. T cell activation. Annu. Rev. Immunol. 2009, 27, 591–619. [Google Scholar] [CrossRef] [PubMed]
- Kuhns, M.S.; Davis, M.M.; Garcia, K.C. Deconstructing the form and function of the TCR/CD3 complex. Immunity 2006, 24, 133–139. [Google Scholar] [CrossRef]
- Yue, T.; Zhan, X.; Zhang, D.; Jain, R.; Wang, K.-W.; Choi, J.H.; Misawa, T.; Su, L.; Quan, J.; Hildebrand, S.; et al. SLFN2 protection of tRNAs from stress-induced cleavage is essential for T cell-mediated immunity. Science 2021, 372, eaba4220. [Google Scholar] [CrossRef]
- Sena, L.A.; Li, S.; Jairaman, A.; Prakriya, M.; Ezponda, T.; Hildeman, D.A.; Wang, C.-R.; Schumacker, P.T.; Licht, J.D.; Perlman, H.; et al. Mitochondria are required for antigen-specific T cell activation through reactive oxygen species signaling. Immunity 2013, 38, 225–236. [Google Scholar] [CrossRef]
- Roy, M.; Kiremidjian-Schumacher, L.; Wishe, H.I.; Cohen, M.W.; Stotzky, G. Selenium supplementation enhances the expression of interleukin 2 receptor subunits and internalization of interleukin 2. Proc. Soc. Exp. Biol. Med. 1993, 202, 295–301. [Google Scholar] [CrossRef]
- Thikkurissy, S.; Pavone, A.; Rega, A.; Bae, R.; Roy, M.; Wishe, H.I.; Kiremidjian-Schumacher, L. Effect of interleukin-2 and selenium on the growth of squamous cell carcinoma cells. Otolaryngol. Head. Neck. Surg. 2001, 124, 142–149. [Google Scholar] [CrossRef]
- Verma, S.; Hoffmann, F.W.; Kumar, M.; Huang, Z.; Roe, K.; Nguyen-Wu, E.; Hashimoto, A.S.; Hoffmann, P.R. Selenoprotein K knockout mice exhibit deficient calcium flux in immune cells and impaired immune responses. J. Immunol. 2011, 186, 2127–2137. [Google Scholar] [CrossRef]
- Ren, X.; Wang, S.; Zhang, C.; Hu, X.; Zhou, L.; Li, Y.; Xu, L. Selenium ameliorates cadmium-induced mouse leydig TM3 cell apoptosis via inhibiting the ROS/JNK/c-jun signaling pathway. Ecotoxicol. Environ. Saf. 2020, 192, 110266. [Google Scholar] [CrossRef]
- Dehghani, M.; Shokrgozar, N.; Ramzi, M.; Kalani, M.; Golmoghaddam, H.; Arandi, N. The impact of selenium on regulatory T cell frequency and immune checkpoint receptor expression in patients with diffuse large B cell lymphoma (DLBCL). Cancer Immunol. Immunother. 2021, 70, 2961–2969. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M. Promotion of interleukin-2 activity as a strategy for ‘rejuvenating’ geriatric immune function. Med. Hypotheses 1997, 48, 47–54. [Google Scholar] [CrossRef]
- Peretz, A.A.; Nève, J.; Desmedt, J.; Duchateau, J.; Dramaix, M.; Famaey, J.P. Lymphocyte response is enhanced by supplementation of elderly subjects with selenium-enriched yeast. Am. J. Clin. Nutr. 1991, 53, 1323–1328. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, F.W.; Hashimoto, A.C.; Shafer, L.A.; Dow, S.; Berry, M.J.; Hoffmann, P.R. Dietary selenium modulates activation and differentiation of CD4+ T cells in mice through a mechanism involving cellular free thiols. J. Nutr. 2010, 140, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Yang, T.; Wang, W.; Xu, S. Effect of selenium antagonist lead-induced damage on Th1/Th2 imbalance in the peripheral blood lymphocytes of chickens. Ecotoxicol. Environ. Saf. 2019, 175, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Romagnani, S. Th1 and Th2 in human diseases. Clin. Immunol. Immunopathol. 1996, 80, 225–235. [Google Scholar] [CrossRef]
- Hawkes, W.C.; Hwang, A.; Alkan, Z. The effect of selenium supplementation on DTH skin responses in healthy North American men. J. Trace Elem. Med. Biol. 2009, 23, 272–280. [Google Scholar] [CrossRef]
- Hasselmark, L.; Malmgren, R.; Zetterström, O.; Unge, G. Selenium supplementation in intrinsic asthma. Allergy 1993, 48, 30–36. [Google Scholar]
- Yao, Y.; Chen, Z.; Zhang, H.; Chen, C.; Zeng, M.; Yunis, J.; Wei, Y.; Wan, Y.; Wang, N.; Zhou, M.; et al. Selenium-GPX4 axis protects follicular helper T cells from ferroptosis. Nat. Immunol. 2021, 22, 1127–1139. [Google Scholar] [CrossRef]
- Werz, O.; Steinhilber, D. Selenium-dependent peroxidases suppress 5-lipoxygenase activity in B-lymphocytes and immature myeloid cells. The presence of peroxidase-insensitive 5-lipoxygenase activity in differentiated myeloid cells. Eur. J. Biochem. 1996, 242, 90–97. [Google Scholar] [CrossRef]
- Vega, L.; Rodríguez-Sosa, M.; García-Montalvo, E.A.; Del Razo, L.M.; Elizondo, G. Non-optimal levels of dietary selenomethionine alter splenocyte response and modify oxidative stress markers in female mice. Food Chem. Toxicol. 2007, 45, 1147–1153. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.-H.; Holmström, A.; Li, X.; Wu, R.T.Y.; Zeng, H.; Xiao, Z. Effect of dietary selenium and cancer cell xenograft on peripheral T and B lymphocytes in adult nude mice. Biol. Trace Elem. Res. 2012, 146, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Jin, Y.; Hall, K.S.; Liang, C.; Unverzagt, F.W.; Ji, R.; Murrell, J.R.; Cao, J.; Shen, J.; Ma, F.; et al. Selenium level and cognitive function in rural elderly Chinese. Am. J. Epidemiol. 2007, 165, 955–965. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, B.R.; Roberts, B.R.; Malpas, C.B.; Vivash, L.; Genc, S.; Saling, M.M.; Desmond, P.; Steward, C.; Hicks, R.J.; Callahan, J.; et al. Supranutritional Sodium Selenate Supplementation Delivers Selenium to the Central Nervous System: Results from a Randomized Controlled Pilot Trial in Alzheimer’s Disease. Neurotherapeutics 2019, 16, 192–202. [Google Scholar] [CrossRef]
- Corcoran, N.M.; Martin, D.; Hutter-Paier, B.; Windisch, M.; Nguyen, T.; Nheu, L.; Sundstrom, L.E.; Costello, A.J.; Hovens, C.M. Sodium selenate specifically activates PP2A phosphatase, dephosphorylates tau and reverses memory deficits in an Alzheimer’s disease model. J. Clin. Neurosci. 2010, 17, 1025–1033. [Google Scholar] [CrossRef]
- van Eersel, J.; Ke, Y.D.; Liu, X.; Delerue, F.; Kril, J.J.; Götz, J.; Ittner, L.M. Sodium selenate mitigates tau pathology, neurodegeneration, and functional deficits in Alzheimer’s disease models. Proc. Natl. Acad. Sci. USA 2010, 107, 13888–13893. [Google Scholar] [CrossRef]
- Sun, C.; Du, Z.; Liu, X.; Yang, Y.; Zhou, S.; Li, C.; Cao, X.; Zhao, Q.; Wong, K.; Chen, W.; et al. Selenium Forms and Dosages Determined Their Biological Actions in Mouse Models of Parkinson’s Disease. Nutrients 2022, 15, 11. [Google Scholar] [CrossRef]
- Sophiabadi, M.; Rastgoo, N.; Haghdoost-Yazdi, H. Dopaminergic Neuronal Death in Substantia Nigra Associates with Serum Levels of Total Bilirubin, Selenium, and Zinc: Evidences from 6-Hydroxydopamine Animal Model of Parkinson’s Disease. Biol. Trace Elem. Res. 2022, 200, 4058–4067. [Google Scholar] [CrossRef]
- Ellwanger, J.H.; Molz, P.; Dallemole, D.R.; dos Santos, A.P.; Müller, T.E.; Cappelletti, L.; da Silva, M.G.; Franke, S.I.R.; Prá, D.; Henriques, J.A.P. Selenium reduces bradykinesia and DNA damage in a rat model of Parkinson’s disease. Nutrition 2015, 31, 359–365. [Google Scholar] [CrossRef]
- Tawfik, K.M.; Moustafa, Y.M.; El-Azab, M.F. Neuroprotective mechanisms of sildenafil and selenium in PTZ-kindling model: Implications in epilepsy. Eur. J. Pharmacol. 2018, 833, 131–144. [Google Scholar] [CrossRef]
- Nazıroglu, M. Role of selenium on calcium signaling and oxidative stress-induced molecular pathways in epilepsy. Neurochem. Res. 2009, 34, 2181–2191. [Google Scholar] [CrossRef]
- Schibler, U. Selenium cysteine and epileptic seizures. Nat. Rev. Mol. Cell Biol. 2018, 19, 753. [Google Scholar] [CrossRef]
- Burk, R.F.; Hill, K.E.; Motley, A.K.; Winfrey, V.P.; Kurokawa, S.; Mitchell, S.L.; Zhang, W. Selenoprotein P and apolipoprotein E receptor-2 interact at the blood-brain barrier and also within the brain to maintain an essential selenium pool that protects against neurodegeneration. FASEB J. 2014, 28, 3579–3588. [Google Scholar] [CrossRef] [PubMed]
- Burk, R.F.; Hill, K.E.; Olson, G.E.; Weeber, E.J.; Motley, A.K.; Winfrey, V.P.; Austin, L.M. Deletion of apolipoprotein E receptor-2 in mice lowers brain selenium and causes severe neurological dysfunction and death when a low-selenium diet is fed. J. Neurosci. 2007, 27, 6207–6211. [Google Scholar] [CrossRef]
- Schweizer, U.; Streckfuss, F.; Pelt, P.; Carlson, B.A.; Hatfield, D.L.; Köhrle, J.; Schomburg, L. Hepatically derived selenoprotein P is a key factor for kidney but not for brain selenium supply. Biochem. J. 2005, 386, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L.; Schweizer, U.; Holtmann, B.; Flohé, L.; Sendtner, M.; Köhrle, J. Gene disruption discloses role of selenoprotein P in selenium delivery to target tissues. Biochem. J. 2003, 370, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Sasuclark, A.R.; Khadka, V.S.; Pitts, M.W. Cell-Type Specific Analysis of Selenium-Related Genes in Brain. Antioxidants 2019, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Scharpf, M.; Schweizer, U.; Arzberger, T.; Roggendorf, W.; Schomburg, L.; Köhrle, J. Neuronal and ependymal expression of selenoprotein P in the human brain. J. Neural. Transm. 2007, 114, 877–884. [Google Scholar] [CrossRef]
- Caito, S.W.; Milatovic, D.; Hill, K.E.; Aschner, M.; Burk, R.F.; Valentine, W.M. Progression of neurodegeneration and morphologic changes in the brains of juvenile mice with selenoprotein P deleted. Brain Res. 2011, 1398, 1–12. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B.; et al. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef]
- Ingold, I.; Berndt, C.; Schmitt, S.; Doll, S.; Poschmann, G.; Buday, K.; Roveri, A.; Peng, X.; Porto Freitas, F.P.; Seibt, T.; et al. Selenium Utilization by GPX4 Is Required to Prevent Hydroperoxide-Induced Ferroptosis. Cell 2018, 172, 409–422.e21. [Google Scholar] [CrossRef] [PubMed]
- Yant, L.J.; Ran, Q.; Rao, L.; Van Remmen, H.; Shibatani, T.; Belter, J.G.; Motta, L.; Richardson, A.; Prolla, T.A. The selenoprotein GPX4 is essential for mouse development and protects from radiation and oxidative damage insults. Free Radic. Biol. Med. 2003, 34, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.-E.; Chen, L.; Na, R.; Liu, Y.; Rios, C.; Van Remmen, H.; Richardson, A.; Ran, Q. Gpx4 ablation in adult mice results in a lethal phenotype accompanied by neuronal loss in brain. Free Radic. Biol. Med. 2012, 52, 1820–1827. [Google Scholar] [CrossRef]
- Wirth, E.K.; Conrad, M.; Winterer, J.; Wozny, C.; Carlson, B.A.; Roth, S.; Schmitz, D.; Bornkamm, G.W.; Coppola, V.; Tessarollo, L.; et al. Neuronal selenoprotein expression is required for interneuron development and prevents seizures and neurodegeneration. FASEB J. 2010, 24, 844–852. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Chen, Y.; Zhang, Y.; Kuang, X.; Liu, Y.; Guo, M.; Ma, L.; Zhang, D.; Li, Q. Iron Metabolism and Ferroptosis in Epilepsy. Front. Neurosci. 2020, 14, 601193. [Google Scholar] [CrossRef] [PubMed]
- Kahn-Kirby, A.H.; Amagata, A.; Maeder, C.I.; Mei, J.J.; Sideris, S.; Kosaka, Y.; Hinman, A.; Malone, S.A.; Bruegger, J.J.; Wang, L.; et al. Targeting ferroptosis: A novel therapeutic strategy for the treatment of mitochondrial disease-related epilepsy. PLoS ONE 2019, 14, e0214250. [Google Scholar] [CrossRef]
- Yoo, M.H.; Gu, X.; Xu, X.M.; Kim, J.Y.; Carlson, B.A.; Patterson, A.D.; Cai, H.; Gladyshev, V.N.; Hatfield, D.L. Delineating the role of glutathione peroxidase 4 in protecting cells against lipid hydroperoxide damage and in Alzheimer’s disease. Antioxid. Redox Signal 2010, 12, 819–827. [Google Scholar] [CrossRef]
- Bellinger, F.P.; Bellinger, M.T.; Seale, L.A.; Takemoto, A.S.; Raman, A.V.; Miki, T.; Manning-Boğ, A.B.; Berry, M.J.; White, L.R.; Ross, G.W. Glutathione Peroxidase 4 is associated with Neuromelanin in Substantia Nigra and Dystrophic Axons in Putamen of Parkinson’s brain. Mol. Neurodegener. 2011, 6, 8. [Google Scholar] [CrossRef]
- Hambright, W.S.; Fonseca, R.S.; Chen, L.; Na, R.; Ran, Q. Ablation of ferroptosis regulator glutathione peroxidase 4 in forebrain neurons promotes cognitive impairment and neurodegeneration. Redox Biol. 2017, 12, 8–17. [Google Scholar] [CrossRef]
- Chung, Y.W.; Jeong, D.; Noh, O.J.; Park, Y.H.; Kang, S.I.; Lee, M.G.; Lee, T.-H.; Bin Yim, M.; Kim, I.Y. Antioxidative role of selenoprotein W in oxidant-induced mouse embryonic neuronal cell death. Mol. Cells 2009, 27, 609–613. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Schweizer, U.; Savaskan, N.E.; Hua, D.; Kipnis, J.; Hatfield, D.L.; Gladyshev, V.N. Comparative analysis of selenocysteine machinery and selenoproteome gene expression in mouse brain identifies neurons as key functional sites of selenium in mammals. J. Biol. Chem. 2008, 283, 2427–2438. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, J. Keshan disease, selenium deficiency, and the selenoproteome. N. Engl. J. Med. 2014, 370, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, Y.; Han, S.; Zhang, Y.; Zou, Y.; Su, S.; Zhou, H.; Zhang, X.; Liang, H.; Hou, J.; et al. A Spatial Ecological Study on Serum Selenium and Keshan Disease in Heilongjiang Province, China. Biol. Trace Elem. Res. 2021, 199, 3253–3261. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.A.; Noiva, R.; Wells, I.C. Selenium concentrations in plasma of patients with arteriographically defined coronary atherosclerosis. Clin. Chem. 1984, 30, 1171–1173. [Google Scholar] [CrossRef]
- Krohn, R.M.; Lemaire, M.; Silva, L.F.N.; Lemarié, C.; Bolt, A.; Mann, K.K.; Smits, J. High-selenium lentil diet protects against arsenic-induced atherosclerosis in a mouse model. J. Nutr. Biochem. 2016, 27, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Schwenke, D.C.; Behr, S.R. Vitamin E combined with selenium inhibits atherosclerosis in hypercholesterolemic rabbits independently of effects on plasma cholesterol concentrations. Circ. Res. 1998, 83, 366–377. [Google Scholar] [CrossRef]
- Zhang, Y.; Cartland, S.P.; Henriquez, R.; Patel, S.; Gammelgaard, B.; Flouda, K.; Hawkins, C.L.; Rayner, B.S. Selenomethionine supplementation reduces lesion burden, improves vessel function and modulates the inflammatory response within the setting of atherosclerosis. Redox Biol. 2020, 29, 101409. [Google Scholar] [CrossRef]
- Luoma, P.V.; Sotaniemi, E.A.; Korpela, H.; Kumpulainen, J. Serum selenium, glutathione peroxidase activity and high-density lipoprotein cholesterol—Effect of selenium supplementation. Res. Commun. Chem. Pathol. Pharmacol. 1984, 46, 469–472. [Google Scholar]
- Maiorino, M.; Conrad, M.; Ursini, F. GPx4, Lipid Peroxidation, and Cell Death: Discoveries, Rediscoveries, and Open Issues. Antioxid. Redox Signal 2018, 29, 61–74. [Google Scholar] [CrossRef]
- Leitinger, N. Cholesteryl ester oxidation products in atherosclerosis. Mol. Aspects. Med. 2003, 24, 239–250. [Google Scholar] [CrossRef]
- Hussein, O.; Rosenblat, M.; Refael, G.; Aviram, M. Dietary selenium increases cellular glutathione peroxidase activity and reduces the enhanced susceptibility to lipid peroxidation of plasma and low-density lipoprotein in kidney transplant recipients. Transplantation 1997, 63, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Toivanen, J.L. Effects of selenium, vitamin E and vitamin C on human prostacyclin and thromboxane synthesis in vitro. Prostaglandins. Leukot. Med. 1987, 26, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Luoma, P.V.; Stengård, J.; Korpela, H.; Rautio, A.; Sotaniemi, E.A.; Suvanto, E.; Marniemi, J. Lipid peroxides, glutathione peroxidase, high density lipoprotein subfractions and apolipoproteins in young adults. J. Intern. Med. 1990, 227, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Choi, A.M.; Ryter, S.W.; Levine, B. Autophagy in human health and disease. N. Engl. J. Med. 2013, 368, 651–662. [Google Scholar] [CrossRef]
- Blankenberg, S.; Rupprecht, H.J.; Bickel, C.; Torzewski, M.; Hafner, G.; Tiret, L.; Smieja, M.; Cambien, F.; Meyer, J.; Lackner, K.J. Glutathione peroxidase 1 activity and cardiovascular events in patients with coronary artery disease. N. Engl. J. Med. 2003, 349, 1605–1613. [Google Scholar] [CrossRef]
- Sordillo, L.M.; Streicher, K.L.; Mullarky, I.K.; Gandy, J.C.; Trigona, W.; Corl, C.M. Selenium inhibits 15-hydroperoxyoctadecadienoic acid-induced intracellular adhesion molecule expression in aortic endothelial cells. Free Radic. Biol. Med. 2008, 44, 34–43. [Google Scholar] [CrossRef]
- Gao, J.; Tian, X.; Yan, X.; Wang, Y.; Wei, J.; Wang, X.; Yan, X.; Song, G. Selenium Exerts Protective Effects Against Fluoride-Induced Apoptosis and Oxidative Stress and Altered the Expression of Bcl-2/Caspase Family. Biol. Trace Elem. Res. 2021, 199, 682–692. [Google Scholar] [CrossRef]
- Han, Q.; Wang, A.; Fu, Q.; Zhou, S.; Bao, J.; Xing, H. Protective role of selenium on ammonia-mediated nephrotoxicity via PI3K/AKT/mTOR pathway: Crosstalk between autophagy and cytokine release. Ecotoxicol. Environ. Saf. 2022, 242, 113918. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Selenium and outcome in heart failure. Eur. J. Heart Fail 2020, 22, 1415–1423. [Google Scholar]
- Fleming, C.R.; Lie, J.; McCall, J.; O’Brien, J.; Baillie, E.E.; Thistle, J.L. Selenium deficiency and fatal cardiomyopathy in a patient on home parenteral nutrition. Gastroenterology 1982, 83, 689–693. [Google Scholar] [CrossRef]
- Jenkins, D.J.A.; Spence, J.D.; Giovannucci, E.L.; Kim, Y.-I.; Josse, R.; Vieth, R.; Blanco Mejia, S.; Viguiliouk, E.; Nishi, S.; Sahye-Pudaruth, S.; et al. Supplemental Vitamins and Minerals for CVD Prevention and Treatment. J. Am. Coll. Cardiol. 2018, 71, 2570–2584. [Google Scholar] [CrossRef] [PubMed]
- Virtamo, J.; Valkeila, E.; Alfthan, G.; Punsar, S.; Huttunen, J.K.; Karvonen, M.J. Serum selenium and the risk of coronary heart disease and stroke. Am. J. Epidemiol. 1985, 122, 276–282. [Google Scholar] [CrossRef]
- Salvini, S.; Hennekens, C.H.; Morris, J.; Willett, W.C.; Stampfer, M.J. Plasma levels of the antioxidant selenium and risk of myocardial infarction among U.S. physicians. Am. J. Cardiol. 1995, 76, 1218–1221. [Google Scholar] [CrossRef] [PubMed]
- Eagle, K.; Jiang, Y.; Shi, X.; Li, M.; Obholzer, N.P.; Hu, T.; Perez, M.W.; Koren, J.V.; Kitano, A.; Yi, J.S.; et al. An oncogenic enhancer encodes selective selenium dependency in AML. Cell Stem. Cell 2022, 29, 386–399.e7. [Google Scholar] [CrossRef]
- Sundström, H.; Korpela, H.; Viinikka, L.; Kauppila, A. Serum selenium and glutathione peroxidase, and plasma lipid peroxides in uterine, ovarian or vulvar cancer, and their responses to antioxidants in patients with ovarian cancer. Cancer Lett. 1984, 24, 1–10. [Google Scholar] [CrossRef]
- Schwartz, M.K. Role of trace elements in cancer. Cancer Res. 1975, 35, 3481–3487. [Google Scholar]
- Willett, W.; Morris, J.S.; Pressel, S.; Taylor, J.; Polk, B.F.; Stampfer, M.; Rosner, B.; Schneider, K.; Hames, C. Prediagnostic serum selenium and risk of cancer. Lancet 1983, 2, 130–134. [Google Scholar] [CrossRef]
- Lotan, Y.; Goodman, P.J.; Youssef, R.F.; Svatek, R.S.; Shariat, S.F.; Tangen, C.M.; Thompson, I.M.; Klein, E.A. Evaluation of vitamin E and selenium supplementation for the prevention of bladder cancer in SWOG coordinated SELECT. J. Urol. 2012, 187, 2005–2010. [Google Scholar] [CrossRef]
- Reid, M.E.; Duffield-Lillico, A.J.; Garland, L.; Turnbull, B.W.; Clark, L.C.; Marshall, J.R. Selenium supplementation and lung cancer incidence: An update of the nutritional prevention of cancer trial. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 1285–1291. [Google Scholar] [PubMed]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: The Selenium and Vitamin E Cancer Prevention Trial (SELECT). JAMA 2009, 301, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Y.; Zhu, Y.J.; Li, W.G. Protective role of selenium against hepatitis B virus and primary liver cancer in Qidong. Biol. Trace Elem. Res. 1997, 56, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Flowers, B.; Poles, A.; Kastrati, I. Selenium and breast cancer—An update of clinical and epidemiological data. Arch. Biochem. Biophys. 2022, 732, 109465. [Google Scholar] [CrossRef]
- Clark, L.C.; Dalkin, B.; Krongrad, A.; Combs, G.F., Jr.; Turnbull, B.W.; Slate, E.H.; Witherington, R.; Herlong, J.H.; Janosko, E.; Carpenter, D.; et al. Decreased incidence of prostate cancer with selenium supplementation: Results of a double-blind cancer prevention trial. Br. J. Urol. 1998, 81, 730–734. [Google Scholar] [CrossRef]
- Mukhtar, M.; Ashfield, N.; Vodickova, L.; Vymetalkova, V.; Levy, M.; Liska, V.; Bruha, J.; Bendova, P.; O’sullivan, J.; Doherty, G.; et al. The Associations of Selenoprotein Genetic Variants with the Risks of Colorectal Adenoma and Colorectal Cancer: Case-Control Studies in Irish and Czech Populations. Nutrients. 2022, 14, 2718. [Google Scholar] [CrossRef]
- Efendić, S.; Luft, R.; Wajngot, A. Aspects of the pathogenesis of type 2 diabetes. Endocr. Rev. 1984, 5, 395–410. [Google Scholar] [CrossRef]
- Stranges, S.; Marshall, J.R.; Natarajan, R.; Donahue, R.P.; Trevisan, M.; Combs, G.F.; Cappuccio, F.P.; Ceriello, A.; Reid, M.E. Effects of long-term selenium supplementation on the incidence of type 2 diabetes: A randomized trial. Ann. Intern. Med. 2007, 147, 217–223. [Google Scholar] [CrossRef]
- Vinceti, M.; Filippini, T.; Rothman, K.J. Selenium exposure and the risk of type 2 diabetes: A systematic review and meta-analysis. Eur. J. Epidemiol. 2018, 33, 789–810. [Google Scholar] [CrossRef]
- Misu, H.; Takamura, T.; Takayama, H.; Hayashi, H.; Matsuzawa-Nagata, N.; Kurita, S.; Ishikura, K.; Ando, H.; Takeshita, Y.; Ota, T.; et al. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab. 2010, 12, 483–495. [Google Scholar] [CrossRef]
- Misu, H.; Takayama, H.; Saito, Y.; Mita, Y.; Kikuchi, A.; Ishii, K.-A.; Chikamoto, K.; Kanamori, T.; Tajima, N.; Lan, F.; et al. Deficiency of the hepatokine selenoprotein P increases responsiveness to exercise in mice through upregulation of reactive oxygen species and AMP-activated protein kinase in muscle. Nat. Med. 2017, 23, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Harmon, J.S.; Bogdani, M.; Parazzoli, S.D.; Mak, S.S.; Oseid, E.A.; Berghmans, M.; Leboeuf, R.C.; Robertson, R.P. beta-Cell-specific overexpression of glutathione peroxidase preserves intranuclear MafA and reverses diabetes in db/db mice. Endocrinology 2009, 150, 4855–4862. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Dai, C.; Guo, M.; Taylor, B.; Harmon, J.S.; Sander, M.; Robertson, R.P.; Powers, A.C.; Stein, R. Inactivation of specific β cell transcription factors in type 2 diabetes. J. Clin. Invest. 2013, 123, 3305–3316. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.D.; Vatamaniuk, M.Z.; Wang, S.K.; Roneker, C.A.; Simmons, R.A.; Lei, X.G. Molecular mechanisms for hyperinsulinaemia induced by overproduction of selenium-dependent glutathione peroxidase-1 in mice. Diabetologia 2008, 51, 1515–1524. [Google Scholar] [CrossRef]
- Erbayraktar, Z.; Yılmaz, O.; Artmann, A.T.; Cehreli, R.; Coker, C. Effects of selenium supplementation on antioxidant defense and glucose homeostasis in experimental diabetes mellitus. Biol. Trace Elem. Res. 2007, 118, 217–226. [Google Scholar] [CrossRef]
- Ayaz, M.; Can, B.; Ozdemir, S.; Turan, B. Protective effect of selenium treatment on diabetes-induced myocardial structural alterations. Biol. Trace Elem. Res. 2002, 89, 215–226. [Google Scholar] [CrossRef]
- Ezaki, O. The insulin-like effects of selenate in rat adipocytes. J. Biol. Chem. 1990, 265, 1124–1128. [Google Scholar] [CrossRef]
- McNeill, J.H.; Delgatty, H.L.; Battell, M.L. Insulinlike effects of sodium selenate in streptozocin-induced diabetic rats. Diabetes 1991, 40, 1675–1678. [Google Scholar] [CrossRef]
- Ghosh, R.; Mukherjee, B.; Chatterjee, M. A novel effect of selenium on streptozotocin-induced diabetic mice. Diabetes Res. 1994, 25, 165–171. [Google Scholar]
- Magnuson, M.A.; Andreone, T.L.; Printz, R.L.; Koch, S.; Granner, D.K. Rat glucokinase gene: Structure and regulation by insulin. Proc. Natl. Acad. Sci. USA 1989, 86, 4838–4842. [Google Scholar] [CrossRef]
- Lv, Y.; Xie, L.; Dong, C.; Yang, R.; Long, T.; Yang, H.; Chen, L.; Zhang, L.; Chen, X.; Luo, X.; et al. Co-exposure of serum calcium, selenium and vanadium is nonlinearly associated with increased risk of type 2 diabetes mellitus in a Chinese population. Chemosphere 2021, 263, 128021. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.; Yang, N.; Li, X. Advances in Understanding How Heavy Metal Pollution Triggers Gastric Cancer. Biomed. Res. Int. 2016, 2016, 7825432. [Google Scholar] [CrossRef] [PubMed]
- Aaseth, J.; Alexander, J.; Alehagen, U.; Tinkov, A.; Skalny, A.; Larsson, A.; Crisponi, G.; Nurchi, V.M. The Aging Kidney-As Influenced by Heavy Metal Exposure and Selenium Supplementation. Biomolecules 2021, 11, 1078. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Mahmood, R. Mercury chloride toxicity in human erythrocytes: Enhanced generation of ROS and RNS, hemoglobin oxidation, impaired antioxidant power, and inhibition of plasma membrane redox system. Environ. Sci. Pollut. Res. Int. 2019, 26, 5645–5657. [Google Scholar] [CrossRef]
- Kim, S.H.; Sharma, R.P. Mercury-induced apoptosis and necrosis in murine macrophages: Role of calcium-induced reactive oxygen species and p38 mitogen-activated protein kinase signaling. Toxicol. Appl. Pharmacol. 2004, 196, 47–57. [Google Scholar] [CrossRef]
- Casalino, E.; Sblano, C.; Landriscina, C. Enzyme activity alteration by cadmium administration to rats: The possibility of iron involvement in lipid peroxidation. Arch. Biochem. Biophys. 1997, 346, 171–179. [Google Scholar] [CrossRef]
- Cao, X.; Fu, M.; Bi, R.; Zheng, X.; Fu, B.; Tian, S.; Liu, C.; Li, Q.; Liu, J. Cadmium induced BEAS-2B cells apoptosis and mitochondria damage via MAPK signaling pathway. Chemosphere 2021, 263, 128346. [Google Scholar] [CrossRef]
- Chayapong, J.; Madhyastha, H.; Madhyastha, R.; Nurrahmah, Q.I.; Nakajima, Y.; Choijookhuu, N.; Hishikawa, Y.; Maruyama, M. Arsenic trioxide induces ROS activity and DNA damage, leading to G0/G1 extension in skin fibroblasts through the ATM-ATR-associated Chk pathway. Environ. Sci. Pollut. Res. Int. 2017, 24, 5316–5325. [Google Scholar] [CrossRef]
- Bramanti, E.; Onor, M.; Colombaioni, L. Neurotoxicity Induced by Low Thallium Doses in Living Hippocampal Neurons: Evidence of Early Onset Mitochondrial Dysfunction and Correlation with Ethanol Production. ACS. Chem. Neurosci. 2019, 10, 451–459. [Google Scholar] [CrossRef]
- Fan, Y.; Zhao, X.; Yu, J.; Xie, J.; Li, C.; Liu, D.; Tang, C.; Wang, C. Lead-induced oxidative damage in rats/mice: A meta-analysis. J. Trace Elem. Med. Biol. 2020, 58, 126443. [Google Scholar] [CrossRef]
- Park, H.-J.; Kim, J.Y.; Kim, J.; Lee, J.-H.; Hahn, J.-S.; Gu, M.B.; Yoon, J. Silver-ion-mediated reactive oxygen species generation affecting bactericidal activity. Water Res. 2009, 43, 1027–1032. [Google Scholar] [CrossRef]
- Chitta, K.R.; Figueroa, J.A.L.; Caruso, J.A.; Merino, E.J. Selenium mediated arsenic toxicity modifies cytotoxicity, reactive oxygen species and phosphorylated proteins. Metallomics 2013, 5, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Saikiran, G.; Mitra, P.; Sharma, S.; Kumar, P.K.; Sharma, P. Selenium, oxidative stress and inflammatory markers in handicraft workers occupationally exposed to lead. Arch. Environ. Occup. Health 2022, 77, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Ganther, H.E. Interactions of vitamin E and selenium with mercury and silver. Ann. N. Y. Acad. Sci. 1980, 355, 212–226. [Google Scholar] [CrossRef]
- Sun, H.-J.; Rathinasabapathi, B.; Wu, B.; Luo, J.; Pu, L.-P.; Ma, L.Q. Arsenic and selenium toxicity and their interactive effects in humans. Environ. Int. 2014, 69, 148–158. [Google Scholar] [CrossRef]
- Schöpfer, J.; Drasch, G.; Schrauzer, G.N. Selenium and cadmium levels and ratios in prostates, livers, and kidneys of nonsmokers and smokers. Biol. Trace Elem. Res. 2010, 134, 180–187. [Google Scholar] [CrossRef]
- Kazi, T.G.; Kolachi, N.F.; Afridi, H.I.; Kazi, N.G.; Sirajuddin; Naeemullah; Arain, S.S. Effects of mineral supplementation on liver cirrhotic/cancer male patients. Biol. Trace Elem. Res. 2012, 150, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Hsueh, Y.-M.; Lin, Y.-C.; Huang, Y.-L.; Shiue, H.-S.; Pu, Y.-S.; Huang, C.-Y.; Chung, C.-J. Effect of plasma selenium, red blood cell cadmium, total urinary arsenic levels, and eGFR on renal cell carcinoma. Sci. Total Environ. 2021, 750, 141547. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Health Effects | |
|---|---|---|
| GPX1 | Antioxidant activity; reduce cellular H2O2. | Cancers [46]; chondrogenic differentiation [47]; T2DM [48]; depression [49]; Keshan disease [50]; cataracts [51]; macular degeneration [52]. |
| GPX2 | Antioxidant activity, protect the mucosa of the gastrointestinal tract and various endothelial cells from oxidative damage. | Cancers [53]; intestinal inflammation [54]. |
| GPX3 | Reduce lipid hydro peroxides and H2O2. | Cancers [55]; myocardial fibrosis [56]; ventricular remodeling [57]. |
| GPX4 | Antioxidant activity; decrease phosphatidylcholine hydroperoxide; suppress cellular ferroptosis. | Osteoarthritis [58]; cancers [59]; cardiomyopathy [60]; ischemia-reperfusion injury [61]; brain function [62]. |
| GPX6 | Not known. | Huntington’s disease [63]. |
| TXNRD1 | Antioxidant activity; regenerate thioredoxin; suppress cell ferroptosis. | Idiopathic pulmonary arterial hypertension [64] hepatocellular carcinoma [65]; osteoarthritis [66]; genetic generalized epilepsy [67]; Keshan disease [50]. |
| TXNRD2 | Regenerate thioredoxin; regulate cell proliferation and apoptosis. | Primary open-angle glaucoma [68]; CVDs [69,70]; cancers [71]; glaucoma [72]. |
| TXNRD3 | Antioxidant activity; suppress pyroptosis. | Male reproduction [73]; colitis and carcinogenesis [74]. |
| DIO1 | Activate T3. | Thyroid hormone metabolism [75]; inhibit hepatosteatosis [76]; renal cancer [77]. |
| DIO2 | Activate T3. | Osteoarthritis [78]; obesity [79]; mental retardation [80]. |
| DIO3 | Inactivate T3. | Osteoarthritis [81]; brain development [82]; sepsis and septic shock [83]. |
| MSRB1 | Antioxidant activity; anti-inflammatory effect; regulate immune responses. | Hepatocellular carcinoma [84]; inflammatory response [85]. |
| SEPHS2 | Sec synthesis. | Cancers [86]. |
| SELENOF | Immunomodulation; regulate glycogenolysis and lipogenesis; participate in vitamin A metabolism. | Cancers [87]; glucose metabolism disorder [88]. |
| SELENOH | Regulate cell cycle progression and proliferation. | Colorectal cancer [89]. |
| SELENOI | Critical enzyme in the central nervous system; T cell activation; neural development; plasmalogen maintenance. | Hereditary spastic paraplegia 81 [90]. |
| SELENOK | Oxidation resistance; Ca2+ flux regulation; immune regulation; apoptosis regulation; suppress cellular ferroptosis. | AD [91]; cervical cancer [92]. |
| SELENOM | Glucose metabolism; Ca2+ flux regulation; apoptosis regulation. | Glioblastoma [93]; non-alcoholic fatty liver disease [94]; synaptic deficits and cognitive dysfunction [95]. |
| SELENON | Muscle development; calcium haemostasis. | Myopathies [96]. |
| SELENOO | Not known. | Thyroid cancer [97]. |
| SELENOP | Antioxidant activity; maintain neuronal activity; transport selenium to tissues; regulate pancreatic β cell function. | Cancers [98,99,100]; seizures and ataxia [101]; CVDs [102]. |
| SELENOS | Regulate inflammation; induce ER stress apoptosis; immune regulation. | Hashimoto’s thyroiditis [103]; CVDs [104]. |
| SELENOT | Promote nerve regeneration; Ca2+ flux regulation; apoptosis regulation; maintain endoplasmic reticulum homeostasis; regulate glucose and lipid metabolism. | Glioblastoma [105]; AD [106]; CVDs [107]. |
| SELENOV | Regulate glucose and lipid metabolism; prevent endoplasmic reticulum stress and oxidative injury; maintain male reproduction. | Not known. |
| SELENOW | Oxidation resistance; regulate bone metabolism; support erythroblast development; muscle development. | Osteoporosis [108]; anemia [109]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Li, X.; Wei, Y. Selenium and Selenoproteins in Health. Biomolecules 2023, 13, 799. https://doi.org/10.3390/biom13050799
Zhang F, Li X, Wei Y. Selenium and Selenoproteins in Health. Biomolecules. 2023; 13(5):799. https://doi.org/10.3390/biom13050799
Chicago/Turabian StyleZhang, Fan, Xuelian Li, and Yumiao Wei. 2023. "Selenium and Selenoproteins in Health" Biomolecules 13, no. 5: 799. https://doi.org/10.3390/biom13050799