
Structural and Biochemical Characterization of Mycobacterium tuberculosis Zinc SufU-SufS Complex

Abstract
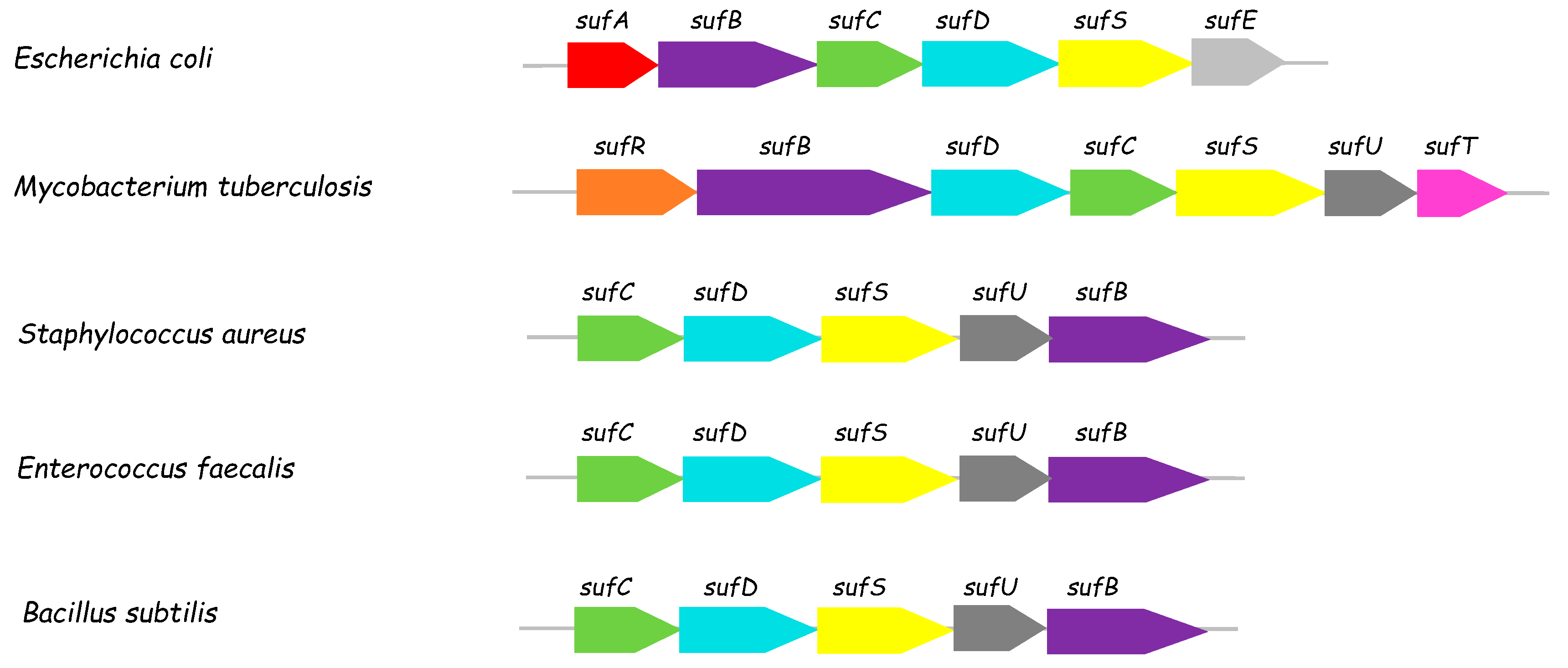
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Plasmid Construction
- ATGCACCACCACCACCACCACGAGAATCTTTATTTTCAGGGCATGACGGCCTCGGTGAACTCGCTCGATCTGGCGGCGATTCGCGCCGATTT
- CCCCATCCTCAAGCGCATCATGCGGGGTGGAAACCCGTTGGCGTATTTGGACTCCGGCGCCACCTCACAACGCCCGCTGCAGGTCCTCGACG
- CCGAGCGCGAGTTCCTGACCGCGTCCAACGGCGCGGTCCATCGTGGCGCGCACCAGCTGATGGAGGAGGCGACCGACGCCTACGAGCAGGGC
- CGCGCGGACATCGCGTTATTCGTCGGCGCCGACACGGACGAGCTGGTGTTCACCAAAAATGCCACCGAGGCGCTCAACCTGGTGTCATATGT
- GCTGGGGGACAGCCGTTTCGAGCGTGCCGTCGGCCCCGGCGACGTGATCGTCACCACCGAGCTGGAGCATCACGCCAACCTGATCCCGTGGC
- AGGAGCTGGCCCGGCGCACCGGGGCCACATTGCGCTGGTACGGGGTGACTGACGACGGGCGCATCGACCTGGACTCGCTGTATCTGGACGAC
- CGTGTCAAAGTCGTTGCGTTCACCCATCATTCCAATGTGACCGGGGTGCTGACACCGGTGAGCGAGCTGGTCTCCCGCGCCCACCAGTCGGG
- TGCGCTGACCGTGCTGGACGCCTGCCAGTCGGTGCCGCACCAGCCGGTTGACCTGCACGAACTCGGCGTCGACTTCGCCGCGTTTTCCGGAC
- ATAAAATGCTGGGCCCCAACGGAATCGGTGTGCTGTACGGCCGCCGTGAGCTGCTAGCGCAGATGCCCCCATTTCTCACCGGCGGTTCGATG
- ATCGAAACGGTGACCATGGAAGGCGCCACCTACGCGCCGGCGCCGCAACGGTTCGAGGCCGGTACCCCGATGACCTCCCAGGTGGTCGGGTT
- GGCCGCCGCGGCCCGCTATCTCGGCGCGATCGGCATGGCCGCGGTGGAGGCCCACGAGCGGGAGCTGGTAGCCGCGGCCATCGAAGGCCTGT
- CCGGCATCGACGGTGTGCGGATCCTTGGCCCGACGTCGATGCGGGACCGAGGGTCGCCGGTGGCGTTCGTCGTCGAGGGCGTGCACGCGCAC
- GACGTGGGTCAGGTACTCGACGACGGCGGCGTGGCGGTGCGGGTCGGGCACCACTGCGCGCTGCCGCTGCACCGCAGGTTCGGTCTGGCCGC
- CACCGCGCGGGCGTCGTTCGCGGTGTACAACACCGCAGACGAGGTGGACCGCTTGGTGGCCGGCGTGCGGCGATCCCGGCATTTCTTTGGAA
- GAGCGTGA
- it produces the following protein sequence (46.4 kDa):
- MHHHHHHENLYFQGMTASVNSLDLAAIRADFPILKRIMRGGNPLAYLDSGATSQRPLQVLDAEREFLTASNGAVHRGAHQLMEEATDAYEQG
- RADIALFVGADTDELVFTKNATEALNLVSYVLGDSRFERAVGPGDVIVTTELEHHANLIPWQELARRTGATLRWYGVTDDGRIDLDSLYLDD
- RVKVVAFTHHSNVTGVLTPVSELVSRAHQSGALTVLDACQSVPHQPVDLHELGVDFAAFSGHKMLGPNGIGVLYGRRELLAQMPPFLTGGSM
- IETVTMEGATYAPAPQRFEAGTPMTSQVVGLAAAARYLGAIGMAAVEAHERELVAAAIEGLSGIDGVRILGPTSMRDRGSPVAFVVEGVHAH
- DVGQVLDDGGVAVRVGHHCALPLHRRFGLAATARASFAVYNTADEVDRLVAGVRRSRHFFGRA
- GMTASVNSLDLAAIRADFPILKRIMRGGNPLAYLDSGATSQRPLQVLDAEREFLTASNGAVHRGAHQLMEEATDAYEQGRADIALFVGADTDELVFTKNATEALNLVSYVLGDSRFERAVGPGDVIVTTELEHHANLIPWQELARRTGATLRWYGVTDDGRIDLDSLYLDDRVKVVAFTHHSNVTGVLTPVSELVSRAHQSGALTVLDACQSVPHQPVDLHELGVDFAAFSGHKMLGPNGIGVLYGRRELLAQMPPFLTGGSMIETVTMEGATYAPAPQRFEAGTPMTSQVVGLAAAARYLGAIGMAAVEAHERELVAAAIEGLSGIDGVRILGPTSMRDRGSPVAFVVEGVHAHDVGQVLDDGGVAVRVGHHCALPLHRRFGLAATARASFAVYNTADEVDRLVAGVRRSRHFFGRA
- CATATGGTTACGCTGCGTCTGGAGCAAATCTATCAGGACGTAATCCTGGACCATTACAAACATCCGCAGCACCGTGGTCTGCGTGAACCGTT
- CGGCGCACAGGTTTACCACGTTAACCCGATCTGCGGTGATGAAGTTACTCTGCGTGTTGCCCTGTCTGAAGACGGTACCCGTGTTACTGACG
- TTTCTTACGACGGTCAGGGTTGCAGCATTTCTCAGGCTGCTACTAGCGTTCTGACCGAGCAGGTTATTGGCCAGCGTGTGCCGCGTGCTCTG
- AACATCGTCGACGCCTTCACTGAAATGGTGAGCTCTCGTGGTACCGTGCCGGGCGATGAGGATGTTCTGGGTGATGGCGTGGCGTTCGCAGG
- TGTTGCAAAGTACCCAGCGCGTGTTAAGTGCGCACTGCTGGGTTGGATGGCTTTCAAAGATGCCCTGGCACAAGCGAGCGAAGCCTTCGAAG
- AAGTCACGGACGAACGTAACCAACGCACCGGTTGACTCGAG
- ATGGGCAGCAGCCATCATCATCATCATCACAGCAGCGGCCTGGTGCCGCGCGGCAGCCATATGGTTACGCTGCGTCTGGAGCAAATCTATCA
- GGACGTAATCCTGGACCATTACAAACATCCGCAGCACCGTGGTCTGCGTGAACCGTTCGGCGCACAGGTTTACCACGTTAACCCGATCTGCG
- GTGATGAAGTTACTCTGCGTGTTGCCCTGTCTGAAGACGGTACCCGTGTTACTGACGTTTCTTACGACGGTCAGGGTTGCAGCATTTCTCAG
- GCTGCTACTAGCGTTCTGACCGAGCAGGTTATTGGCCAGCGTGTGCCGCGTGCTCTGAACATCGTCGACGCCTTCACTGAAATGGTGAGCTC
- TCGTGGTACCGTGCCGGGCGATGAGGATGTTCTGGGTGATGGCGTGGCGTTCGCAGGTGTTGCAAAGTACCCAGCGCGTGTTAAGTGCGCAC
- TGCTGGGTTGGATGGCTTTCAAAGATGCCCTGGCACAAGCGAGCGAAGCCTTCGAAGAAGTCACGGACGAACGTAACCAACGCACCGGTTGA
- which produces the following 20-kDa protein sequence:
- MGSSHHHHHHSSGLVPRGSHMVTLRLEQIYQDVILDHYKHPQHRGLREPFGAQVYHVNPICGDEVTLRVALSEDGTRVTDVSYDGQGCSISQAATSVLTEQVIGQRVPRALNIVDAFTEMVSSRGTVPGDEDVLGDGVAFAGVAKYPARVKCALLGWMAFKDALAQASEAFEEVTDERNQRTG
- GSHMVTLRLEQIYQDVILDHYKHPQHRGLREPFGAQVYHVNPICGDEVTLRVALSEDGTRVTDVSYDGQGCSISQAATSVLTEQVIGQRVPRALNIVDAFTEMVSSRGTVPGDEDVLGDGVAFAGVAKYPARVKCALLGWMAFKDALAQASEAFEEVTDERNQRTG
2.3. Protein Expression and Purification
2.4. SEC-MALLS-RI
2.5. Cysteine Desulfurase Activity
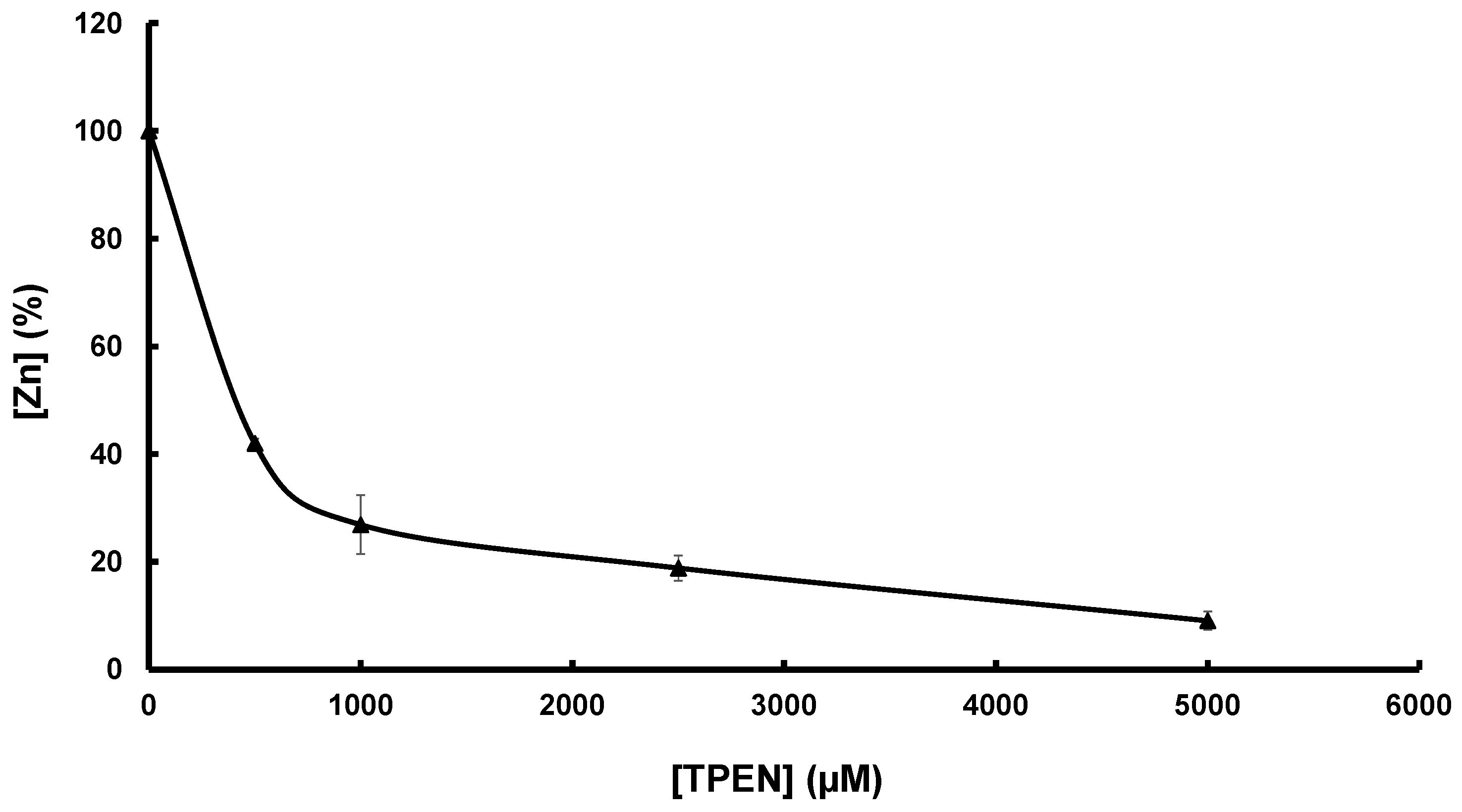
2.6. SufU Affinity for Zinc
2.7. Apo-SufU Preparation
2.8. Reconstitution of SufU with Zinc
2.9. Reconstitution of SufU with Divalent Metal Ions
2.10. ICP-AES
2.11. 6xHis-Tag Cleavage from 6xHis-SufS and 6xHis-SufU
2.12. Fe-S Cluster Reconstitution in SufU
2.13. UV-Visible Absorption Spectroscopy
2.14. Circular Dichroism Analysis
2.15. X-ray Crystallography
3. Results
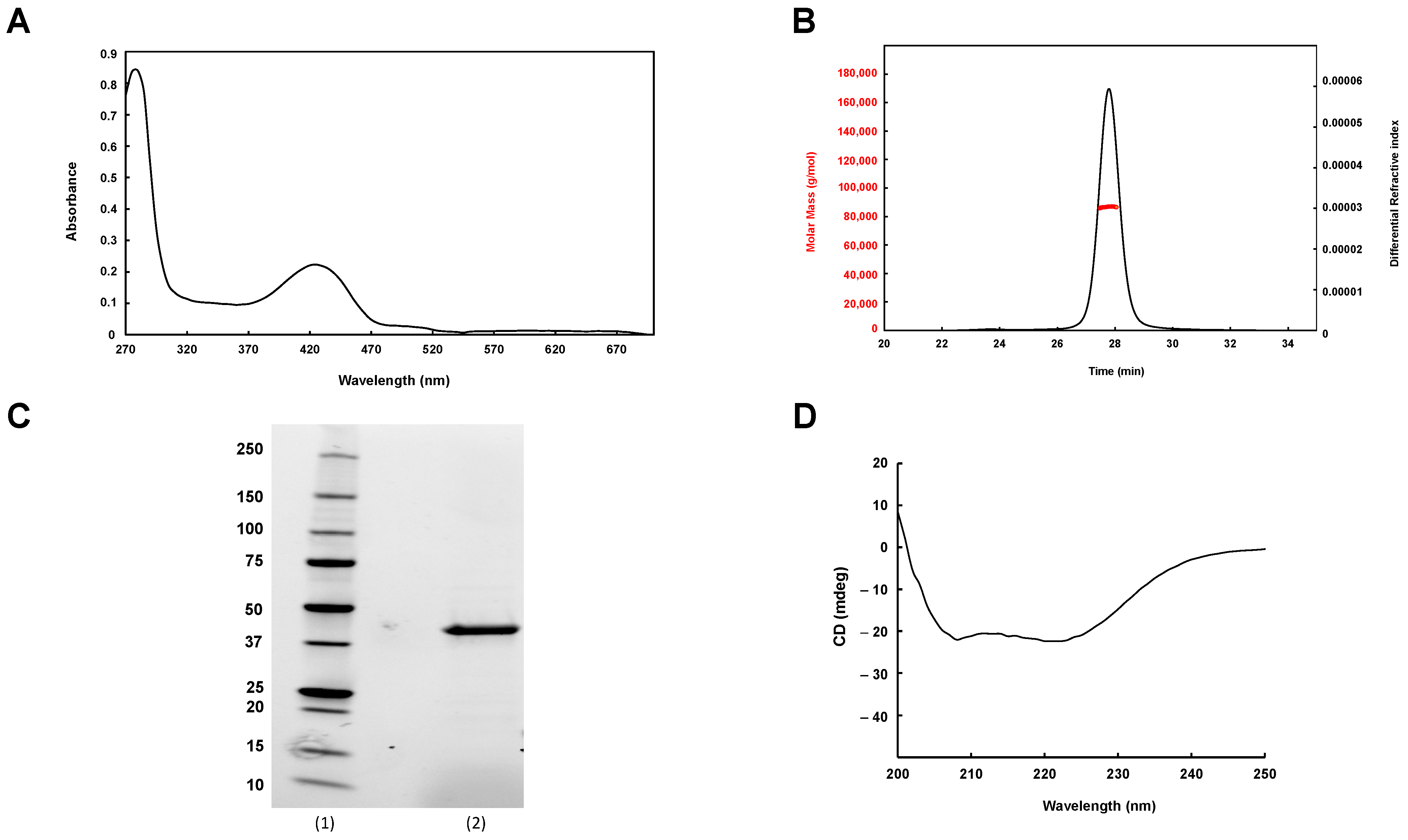
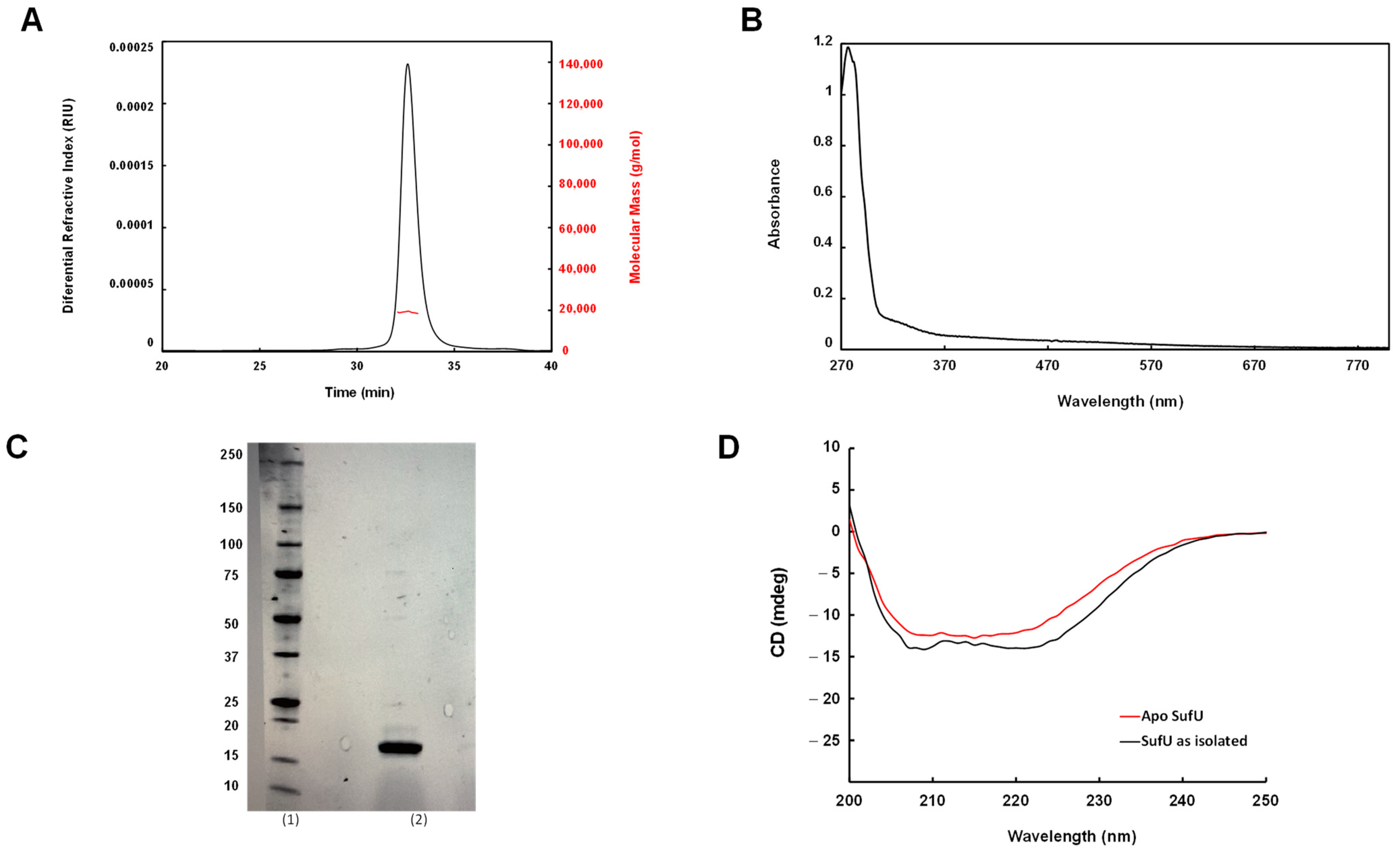
3.1. Purification of Mtb Rv1464 and Rv1465
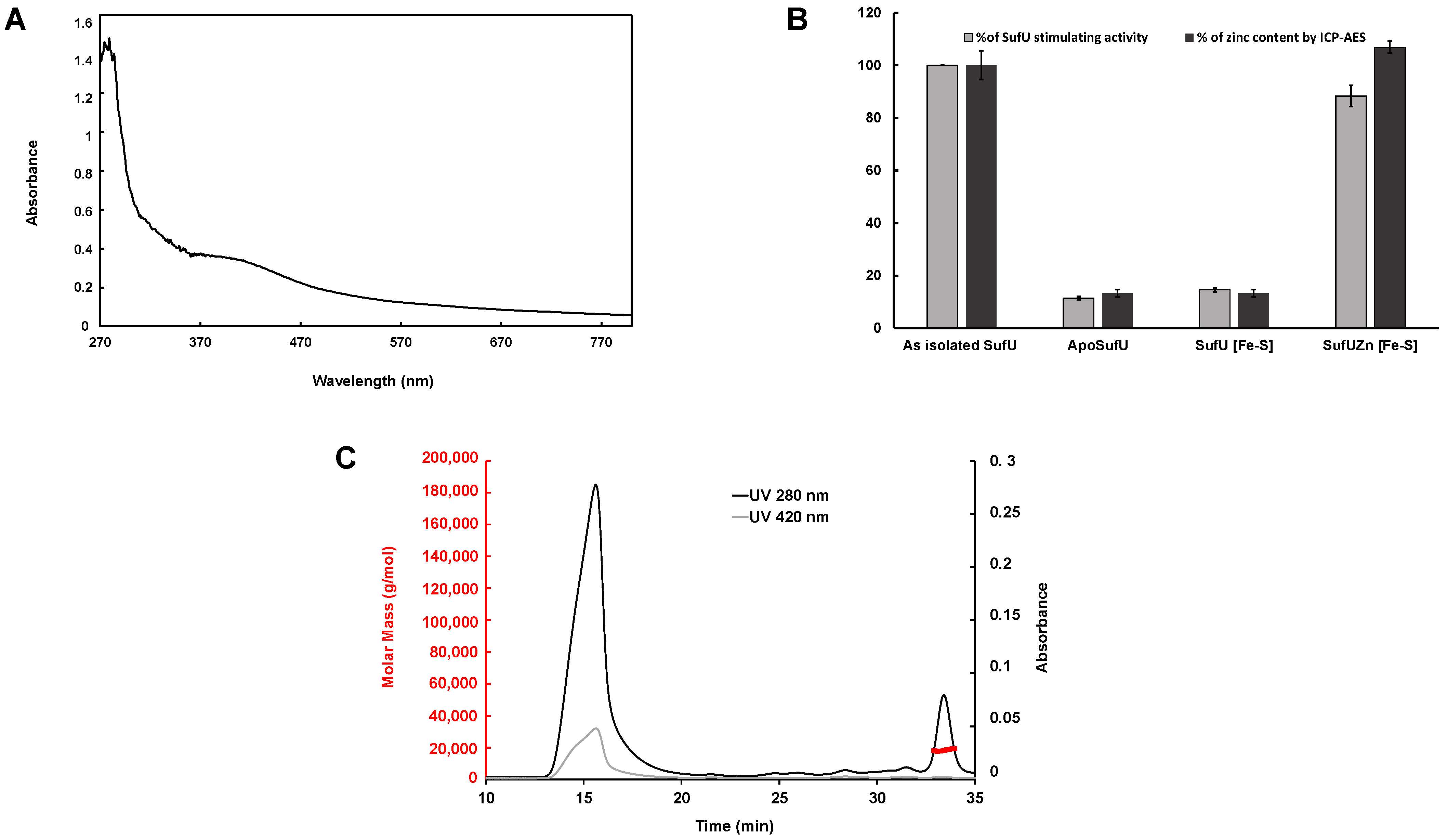
3.2. Zn Is Tightly Bound to Mtb SufU
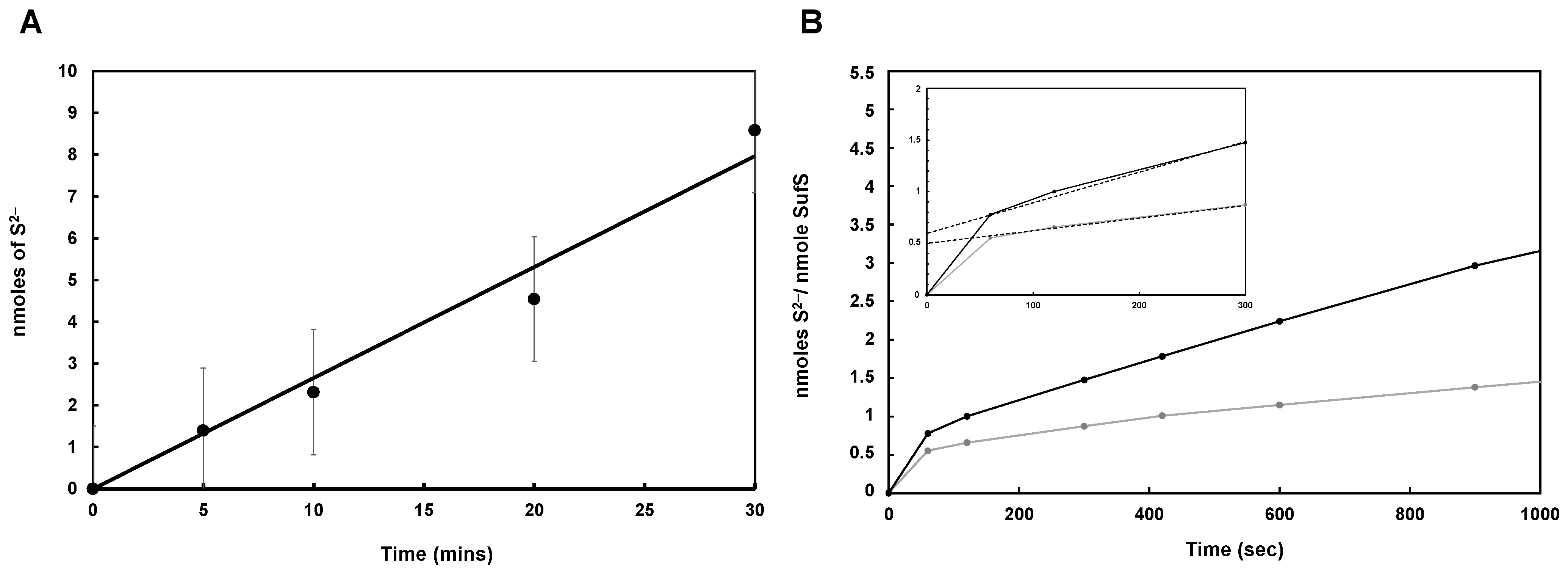
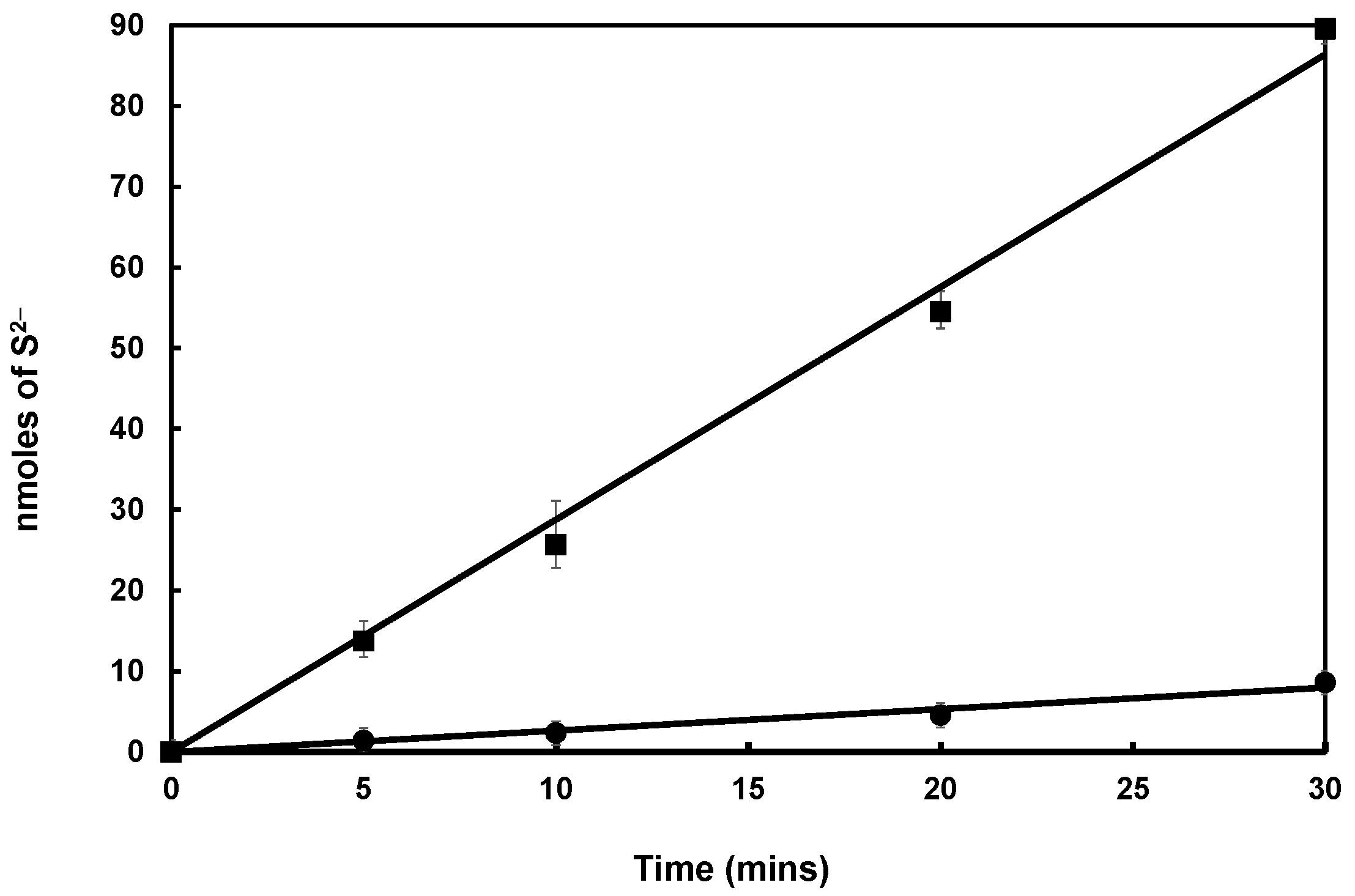
3.3. Mtb SufS Displays a Cysteine Desulfurase Activity and a Half-Site Reactivity
3.4. Mtb Zn-SufU Is a Substrate of SufS
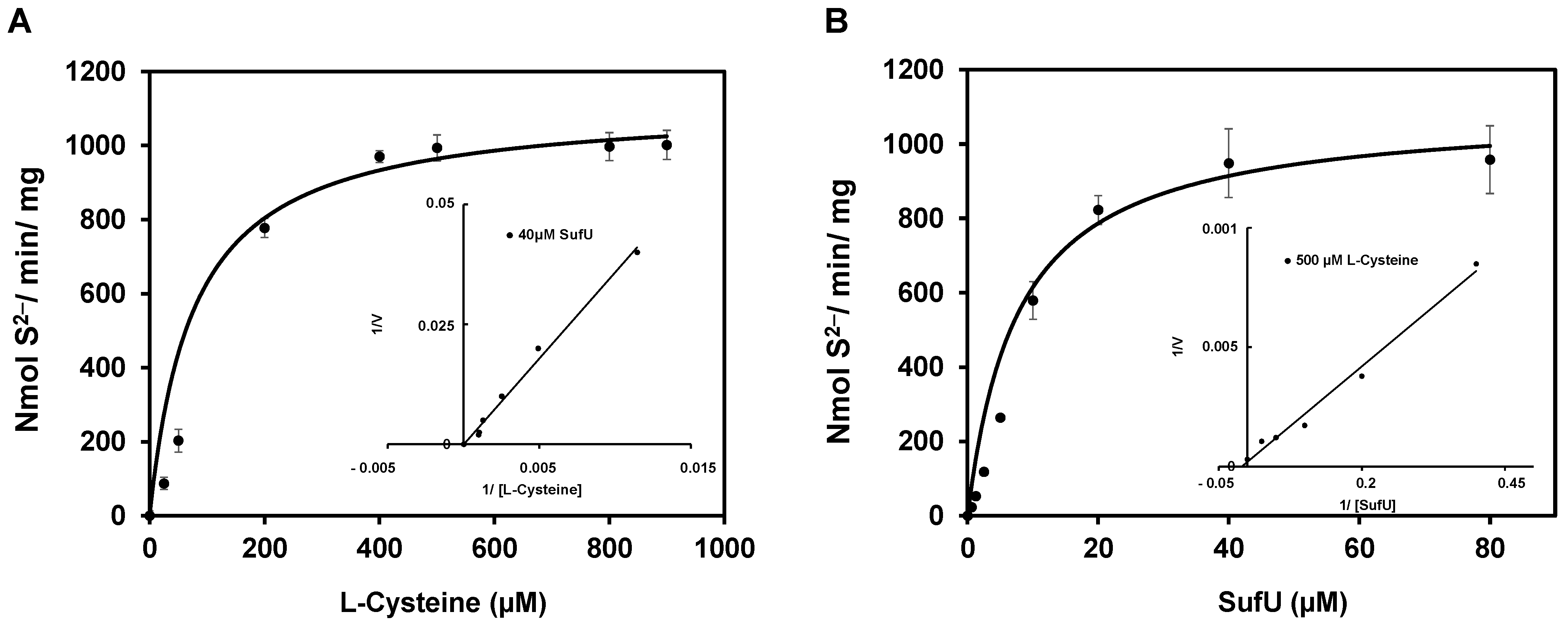
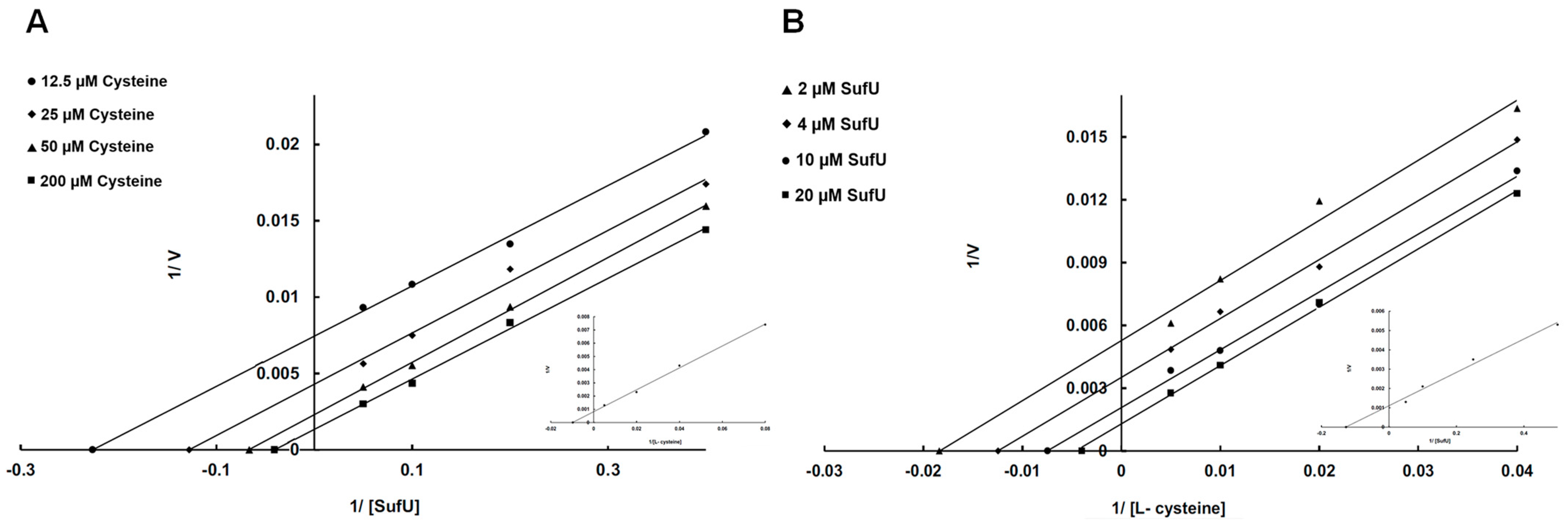
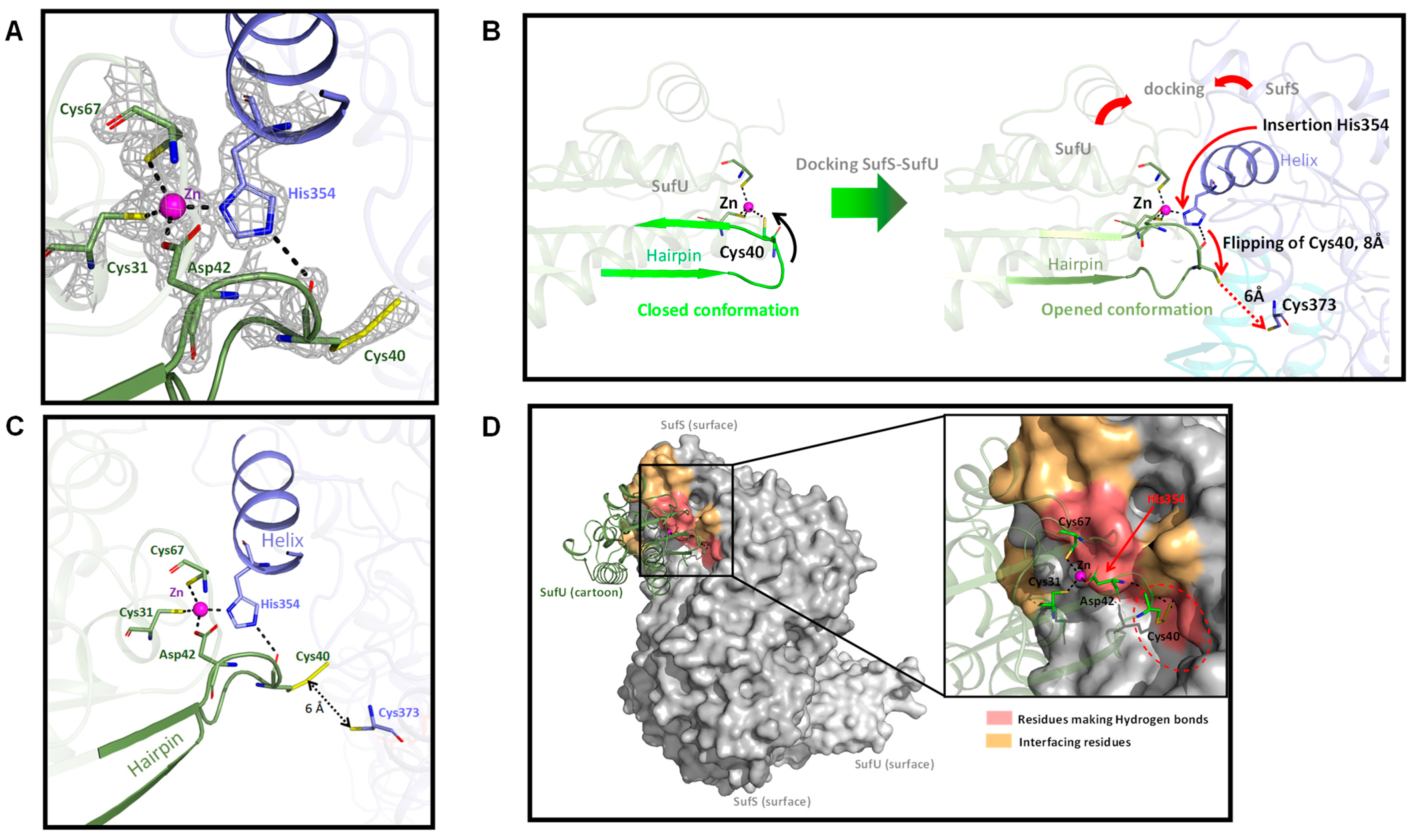
3.5. Mechanism of SufS Activity
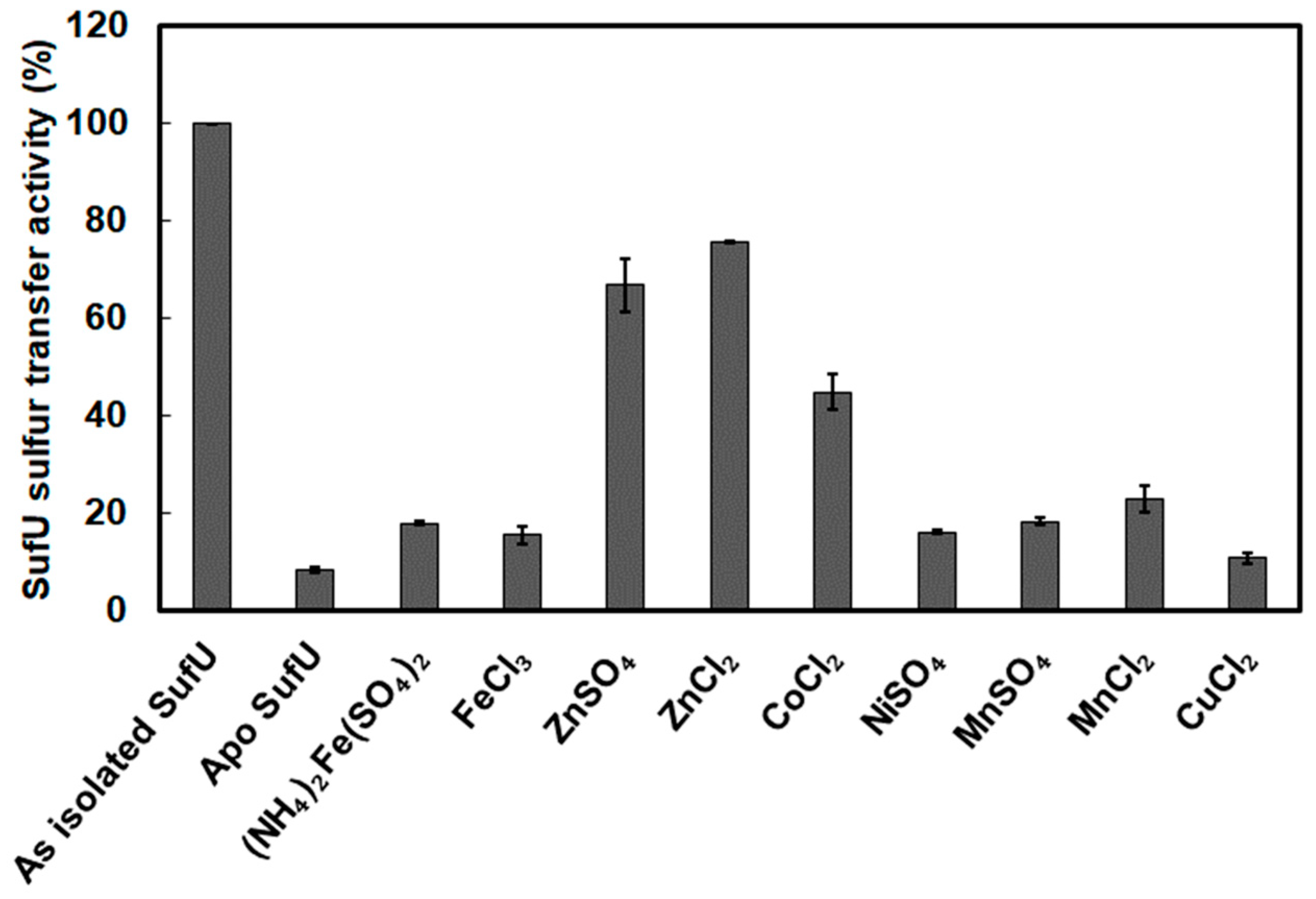
3.6. Zinc Bound to SufU Is Required for SufS Activity
3.7. SufU as a Sulfur Acceptor Protein
3.8. Mtb Zn-SufU Is Not an Intermediate in Fe-S Formation
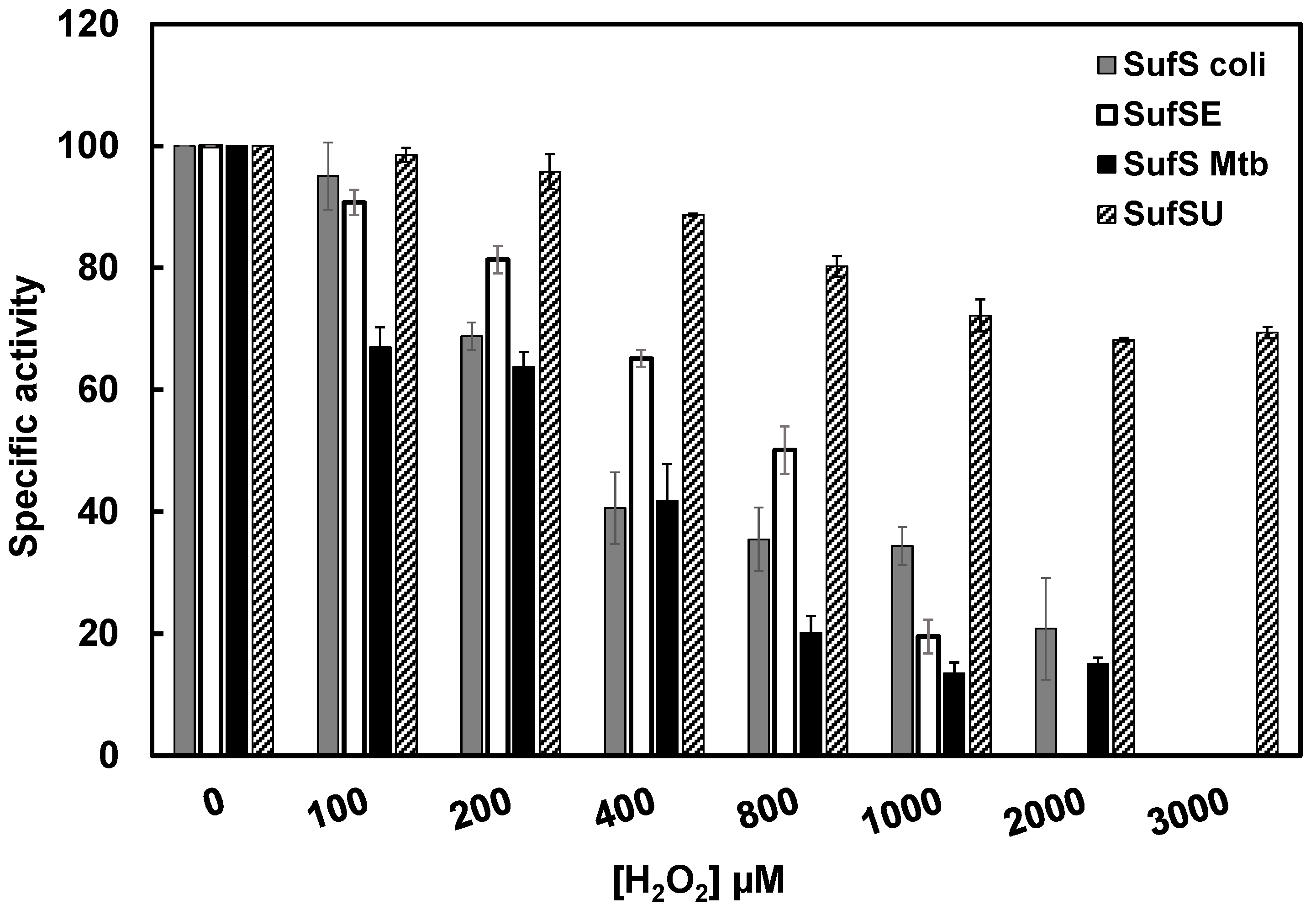
3.9. Mtb SufS-SufU Is More Resistant to H2O2 Than E. coli SufS-SufE
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, M.K. Encyclopedia of Inorganic Chemistry; King, R.B., Ed.; Wiley: New York, NY, USA, 2005; pp. 1896–1915. [Google Scholar]
- Johnson, M.K. Iron-sulfur proteins: New roles for old clusters. Curr. Opin. Chem. Biol. 1998, 2, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Lill, R.; Srinivasan, V.; Muhlenhoff, U. The role of mitochondria in cytosolic-nuclear iron-sulfur protein biogenesis and in cellular iron regulation. Curr. Opin. Microbiol. 2014, 22, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Py, B.; Barras, F. Building Fe-S proteins: Bacterial strategies. Nat. Rev. Microbiol. 2010, 8, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Garcia, P.S.; D’Angelo, F.; Ollagnier de Choudens, S.; Dussouchaud, M.; Bouveret, E.; Gribaldo, S.; Barras, F. An early origin of iron-sulfur cluster biosynthesis machineries before Earth oxygenation. Nat. Ecol. Evol. 2022, 6, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Huet, G.; Daffe, M.; Saves, I. Identification of the Mycobacterium tuberculosis SUF machinery as the exclusive mycobacterial system of [Fe-S] cluster assembly: Evidence for its implication in the pathogen’s survival. J. Bacteriol. 2005, 187, 6137–6146. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.A.; Al-Tameemi, H.M.; Mashruwala, A.A.; Rosario-Cruz, Z.; Chauhan, U.; Sause, W.E.; Torres, V.J.; Belden, W.J.; Boyd, J.M. The Suf Iron-Sulfur Cluster Biosynthetic System Is Essential in Staphylococcus aureus, and Decreased Suf Function Results in Global Metabolic Defects and Reduced Survival in Human Neutrophils. Infect. Immun. 2017, 85, e00100-17. [Google Scholar] [CrossRef]
- Rybniker, J.; Pojer, F.; Marienhagen, J.; Kolly, G.S.; Chen, J.M.; van Gumpel, E.; Hartmann, P.; Cole, S.T. The cysteine desulfurase IscS of Mycobacterium tuberculosis is involved in iron-sulfur cluster biogenesis and oxidative stress defence. Biochem. J. 2014, 459, 467–478. [Google Scholar] [CrossRef]
- Griffin, J.E.; Gawronski, J.D.; Dejesus, M.A.; Ioerger, T.R.; Akerley, B.J.; Sassetti, C.M. High-resolution phenotypic profiling defines genes essential for mycobacterial growth and cholesterol catabolism. PLoS Pathog. 2011, 7, e1002251. [Google Scholar] [CrossRef] [PubMed]
- Sassetti, C.M.; Boyd, D.H.; Rubin, E.J. Genes required for mycobacterial growth defined by high density mutagenesis. Mol. Microbiol. 2003, 48, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.; De Jesus, M.A.; Poulton, N.C.; Zhang, W.Z.; Engelhart, C.A.; Zaveri, A.; Lavalette, S.; Ruecker, N.; Trujillo, C.; Wallach, J.B.; et al. Genome-wide gene expression tuning reveals diverse vulnerabilities of M. tuberculosis. Cell 2021, 184, 4579–4592. [Google Scholar] [CrossRef]
- Elchennawi, I.; de Choudens, S.O. Iron-Sulfur Clusters toward Stresses: Implication for Understanding and Fighting Tuberculosis. Inorganics 2022, 10, 174. [Google Scholar] [CrossRef]
- Selbach, B.P.; Pradhan, P.K.; Dos Santos, P.C. Protected sulfur transfer reactions by the Escherichia coli Suf system. Biochemistry 2013, 52, 4089–4096. [Google Scholar] [CrossRef] [PubMed]
- Loiseau, L.; Ollagnier-de-Choudens, S.; Nachin, L.; Fontecave, M.; Barras, F. Biogenesis of Fe-S cluster by the bacterial Suf system: SufS and SufE form a new type of cysteine desulfurase. J. Biol. Chem. 2003, 278, 38352–38359. [Google Scholar] [CrossRef] [PubMed]
- Outten, F.W.; Wood, M.J.; Munoz, F.M.; Storz, G. The SufE protein and the SufBCD complex enhance SufS cysteine desulfurase activity as part of a sulfur transfer pathway for Fe-S cluster assembly in Escherichia coli. J. Biol. Chem. 2003, 278, 45713–45719. [Google Scholar] [CrossRef] [PubMed]
- Layer, G.; Gaddam, S.A.; Ayala-Castro, C.N.; Ollagnier-de Choudens, S.; Lascoux, D.; Fontecave, M.; Outten, F.W. SufE transfers sulfur from SufS to SufB for iron-sulfur cluster assembly. J. Biol. Chem. 2007, 282, 13342–13350. [Google Scholar] [CrossRef]
- Chahal, H.K.; Dai, Y.; Saini, A.; Ayala-Castro, C.; Outten, F.W. The SufBCD Fe-S scaffold complex interacts with SufA for Fe-S cluster transfer. Biochemistry 2009, 48, 10644–10653. [Google Scholar] [CrossRef] [PubMed]
- Chahal, H.K.; Outten, F.W. Separate FeS scaffold and carrier functions for SufB(2)C(2) and SufA during in vitro maturation of [2Fe2S] Fdx. J. Inorg. Biochem. 2012, 116, 126–134. [Google Scholar] [CrossRef]
- Wollers, S.; Layer, G.; Garcia-Serres, R.; Signor, L.; Clemancey, M.; Latour, J.M.; Fontecave, M.; Ollagnier de Choudens, S. Iron-sulfur (Fe-S) cluster assembly: The SufBCD complex is a new type of Fe-S scaffold with a flavin redox cofactor. J. Biol. Chem. 2010, 285, 23331–23341. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Sendra, M.; Naik, S.G.; Chahal, H.K.; Huynh, B.H.; Outten, F.W.; Fontecave, M.; Ollagnier de Choudens, S. Native Escherichia coli SufA, coexpressed with SufBCDSE, purifies as a [2Fe-2S] protein and acts as an Fe-S transporter to Fe-S target enzymes. J. Am. Chem. Soc. 2009, 131, 6149–6153. [Google Scholar] [CrossRef] [PubMed]
- Selbach, B.; Earles, E.; Dos Santos, P.C. Kinetic analysis of the bisubstrate cysteine desulfurase SufS from Bacillus subtilis. Biochemistry 2010, 49, 8794–8802. [Google Scholar] [CrossRef] [PubMed]
- Selbach, B.P.; Chung, A.H.; Scott, A.D.; George, S.J.; Cramer, S.P.; Dos Santos, P.C. Fe-S cluster biogenesis in Gram-positive bacteria: SufU is a zinc-dependent sulfur transfer protein. Biochemistry 2014, 53, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, A.G.; Netz, D.J.; Miethke, M.; Pierik, A.J.; Burghaus, O.; Peuckert, F.; Lill, R.; Marahiel, M.A. SufU is an essential iron-sulfur cluster scaffold protein in Bacillus subtilis. J. Bacteriol. 2010, 192, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, T.; Terahata, T.; Kunichika, K.; Yokoyama, N.; Maruyama, C.; Asai, K.; Takahashi, Y. Zinc-Ligand Swapping Mediated Complex Formation and Sulfur Transfer between SufS and SufU for Iron-Sulfur Cluster Biogenesis in Bacillus subtilis. J. Am. Chem. Soc. 2017, 139, 18464–18467. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.C.B. subtilis as a Model for Studying the Assembly of Fe-S Clusters in Gram-Positive Bacteria. Methods Enzymol. 2017, 595, 185–212. [Google Scholar] [CrossRef] [PubMed]
- Willemse, D.; Weber, B.; Masino, L.; Warren, R.M.; Adinolfi, S.; Pastore, A.; Williams, M.J. Rv1460, a SufR homologue, is a repressor of the suf operon in Mycobacterium tuberculosis. PLoS ONE 2018, 13, e0200145. [Google Scholar] [CrossRef] [PubMed]
- Tamuhla, T.; Joubert, L.; Willemse, D.; Williams, M.J. SufT is required for growth of Mycobacterium smegmatis under iron limiting conditions. Microbiology 2020, 166, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Jain, P.; Gulten, G.; Liu, Z.; Feng, Y.; Ganesula, K.; Motiwala, A.S.; Ioerger, T.R.; Alland, D.; Vilcheze, C.; et al. Mycobacterium tuberculosis dihydrofolate reductase is not a target relevant to the antitubercular activity of isoniazid. Antimicrob. Agents Chemother. 2010, 54, 3776–3782. [Google Scholar] [CrossRef] [PubMed]
- Bashiri, G.; Rehan, A.M.; Greenwood, D.R.; Dickson, J.M.; Baker, E.N. Metabolic engineering of cofactor F420 production in Mycobacterium smegmatis. PLoS ONE 2010, 5, e15803. [Google Scholar] [CrossRef]
- Gautam, U.S.; Asrican, R.; Sempowski, G.D. Targeted dose delivery of Mycobacterium tuberculosis in mice using silicon antifoaming agent via aerosol exposure system. PLoS ONE 2022, 17, e0276130. [Google Scholar] [CrossRef]
- Fish, W.W. Rapid colorimetric micromethod for the quantitation of complexed iron in biological samples. Methods Enzymol. 1988, 158, 357–364. [Google Scholar]
- Beinert, H. Semi-micro methods for analysis of labile sulfide and of labile sulfide plus sulfane sulfur in unusually stable iron-sulfur proteins. Anal. Biochem. 1983, 131, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Pernot, P.; Round, A.; Barrett, R.; De Maria Antolinos, A.; Gobbo, A.; Gordon, E.; Huet, J.; Kieffer, J.; Lentini, M.; Mattenet, M.; et al. Upgraded ESRF BM29 beamline for SAXS on macromolecules in solution. J. Synchrotron. Radiat. 2013, 20, 660–664. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, A.A.; Barrett, R.; Beteva, A.; Caserotto, H.; Dobias, F.; Felisaz, F.; Giraud, T.; Guijarro, M.; Janocha, R.; Khadrouche, A.; et al. ID30B-a versatile beamline for macromolecular crystallography experiments at the ESRF. J. Synchrotron. Radiat. 2018, 25, 1249–1260. [Google Scholar] [CrossRef]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Winn, M.D.; Ballard, C.C.; Cowtan, K.D.; Dodson, E.J.; Emsley, P.; Evans, P.R.; Keegan, R.M.; Krissinel, E.B.; Leslie, A.G.; McCoy, A.; et al. Overview of the CCP4 suite and current developments. Acta Crystallogr. D Biol. Crystallogr. 2011, 67, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Chojnowski, G.; Pereira, J.; Lamzin, V.S. Sequence assignment for low-resolution modelling of protein crystal structures. Acta Crystallogr. D Struct. Biol. 2019, 75, 753–763. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Mihara, H.; Esaki, N. Bacterial cysteine desulfurases: Their function and mechanisms. Appl. Microbiol. Biotechnol. 2002, 60, 12–23. [Google Scholar]
- Black, K.A.; Dos Santos, P.C. Shared-intermediates in the biosynthesis of thio-cofactors: Mechanism and functions of cysteine desulfurases and sulfur acceptors. Biochim. Biophys. Acta 2015, 1853, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Behshad, E.; Bollinger, J.M., Jr. Kinetic analysis of cysteine desulfurase CD0387 from Synechocystis sp. PCC 6803: Formation of the persulfide intermediate. Biochemistry 2009, 48, 12014–12023. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, E.N.; de Oliveira, J.S.; Nicolet, Y.; Raulfs, E.C.; Amara, P.; Dean, D.R.; Fontecilla-Camps, J.C. (IscS-IscU)2 complex structures provide insights into Fe2S2 biogenesis and transfer. Angew. Chem. Int. Ed. Engl. 2012, 51, 5439–5442. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Proteau, A.; Villarroya, M.; Moukadiri, I.; Zhang, L.; Trempe, J.F.; Matte, A.; Armengod, M.E.; Cygler, M. Structural basis for Fe-S cluster assembly and tRNA thiolation mediated by IscS protein-protein interactions. PLoS Biol. 2010, 8, e1000354. [Google Scholar] [CrossRef]
- Smith, A.D.; Agar, J.N.; Johnson, K.A.; Frazzon, J.; Amster, I.J.; Dean, D.R.; Johnson, M.K. Sulfur transfer from IscS to IscU: The first step in iron-sulfur cluster biosynthesis. J. Am. Chem. Soc. 2001, 123, 11103–11104. [Google Scholar] [CrossRef] [PubMed]
- Ikeuchi, Y.; Shigi, N.; Kato, J.; Nishimura, A.; Suzuki, T. Mechanistic insights into sulfur relay by multiple sulfur mediators involved in thiouridine biosynthesis at tRNA wobble positions. Mol. Cell. 2006, 21, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Kambampati, R.; Lauhon, C.T. Evidence for the transfer of sulfane sulfur from IscS to ThiI during the in vitro biosynthesis of 4-thiouridine in Escherichia coli tRNA. J. Biol. Chem. 2000, 275, 10727–10730. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Park, S. Structural changes during cysteine desulfurase CsdA and sulfur acceptor CsdE interactions provide insight into the trans-persulfuration. J. Biol. Chem. 2013, 288, 27172–27180. [Google Scholar] [CrossRef]
- Lima, C.D. Analysis of the E. coli NifS CsdB protein at 2.0 A reveals the structural basis for perselenide and persulfide intermediate formation. J. Mol. Biol. 2002, 315, 1199–1208. [Google Scholar] [CrossRef]
- Singh, H.; Dai, Y.; Outten, F.W.; Busenlehner, L.S. Escherichia coli SufE sulfur transfer protein modulates the SufS cysteine desulfurase through allosteric conformational dynamics. J. Biol. Chem. 2013, 288, 36189–36200. [Google Scholar] [CrossRef]
- Dunkle, J.A.; Bruno, M.R.; Outten, F.W.; Frantom, P.A. Structural Evidence for Dimer-Interface-Driven Regulation of the Type II Cysteine Desulfurase, SufS. Biochemistry 2019, 58, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Dunkle, J.A.; Bruno, M.R.; Frantom, P.A. Structural evidence for a latch mechanism regulating access to the active site of SufS-family cysteine desulfurases. Acta Crystallogr. D Struct. Biol. 2020, 76, 291–301. [Google Scholar] [CrossRef]
- Dos Santos, P.C.; Smith, A.D.; Frazzon, J.; Cash, V.L.; Johnson, M.K.; Dean, D.R. Iron-sulfur cluster assembly: NifU-directed activation of the nitrogenase Fe protein. J. Biol. Chem. 2004, 279, 19705–19711. [Google Scholar] [CrossRef] [PubMed]
- Collet, J.F.; D’Souza, J.C.; Jakob, U.; Bardwell, J.C. Thioredoxin 2, an oxidative stress-induced protein, contains a high affinity zinc binding site. J. Biol. Chem. 2003, 278, 45325–45332. [Google Scholar] [CrossRef] [PubMed]
- Jakob, U.; Eser, M.; Bardwell, J.C. Redox switch of hsp33 has a novel zinc-binding motif. J. Biol. Chem. 2000, 275, 38302–38310. [Google Scholar] [CrossRef] [PubMed]
- Hitomi, Y.; Outten, C.E.; O’Halloran, T.V. Extreme zinc-binding thermodynamics of the metal sensor/regulator protein, ZntR. J. Am. Chem. Soc. 2001, 123, 8614–8615. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Outten, F.W. The E. coli SufS-SufE sulfur transfer system is more resistant to oxidative stress than IscS-IscU. FEBS Lett. 2012, 586, 4016–4022. [Google Scholar] [CrossRef] [PubMed]
- Shumilina, E.; Dobrovolska, O.; Del Conte, R.; Holen, H.W.; Dikiy, A. Competitive cobalt for zinc substitution in mammalian methionine sulfoxide reductase B1 overexpressed in E. coli: Structural and functional insight. J. Biol. Inorg. Chem. 2014, 19, 85–95. [Google Scholar] [CrossRef]
- Anderson, R.A.; Vallee, B.L. Cobalt(III), a probe of metal binding sites of Escherichia coli alkaline phosphatase. Proc. Natl. Acad. Sci. USA 1975, 72, 394–397. [Google Scholar] [CrossRef]
- Bertini, I.; Gray, H.B.; Lippard, S.J.; Valentine, J.S. Bioinorganic Chemistry; Mill Valley, C.A., Ed.; University Science Books: Herndon, VA, USA, 1994. [Google Scholar]
- Sourice, M.; Askenasy, I.; Garcia, P.S.; Denis, Y.; Brasseur, G.; Kiley, P.J.; Py, B.; Aubert, C. A Diverged Transcriptional Network for Usage of Two Fe-S Cluster Biogenesis Machineries in the Delta-Proteobacterium Myxococcus xanthus. Mbio 2023, 14, e0300122. [Google Scholar] [CrossRef]
- Hudspeth, J.D.; Boncella, A.E.; Sabo, E.T.; Andrews, T.; Boyd, J.M.; Morrison, C.N. Structural and Biochemical Characterization of Staphylococcus aureus Cysteine Desulfurase Complex SufSU. ACS Omega 2022, 7, 44124–44133. [Google Scholar] [CrossRef] [PubMed]
- Riboldi, G.P.; de Oliveira, J.S.; Frazzon, J. Enterococcus faecalis SufU scaffold protein enhances SufS desulfurase activity by acquiring sulfur from its cysteine-153. Biochim. Biophys. Acta 2011, 1814, 1910–1918. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Imlay, J.A. Hydrogen peroxide inactivates the Escherichia coli Isc iron-sulphur assembly system, and OxyR induces the Suf system to compensate. Mol. Microbiol. 2010, 78, 1448–1467. [Google Scholar] [CrossRef]
- Ehrt, S.; Schnappinger, D. Mycobacterial survival strategies in the phagosome: Defence against host stresses. Cell. Microbiol. 2009, 11, 1170–1178. [Google Scholar] [CrossRef] [PubMed]
- Shastri, M.D.; Shukla, S.D.; Chong, W.C.; Dua, K.; Peterson, G.M.; Patel, R.P.; Hansbro, P.M.; Eri, R.; O’Toole, R.F. Role of Oxidative Stress in the Pathology and Management of Human Tuberculosis. Oxid. Med. Cell. Longev. 2018, 2018, 7695364. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C.; Metodiewa, D. Reactivity of biologically important thiol compounds with superoxide and hydrogen peroxide. Free Radic. Biol. Med. 1999, 27, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Ortiz de Orue Lucana, D.; Wedderhoff, I.; Groves, M.R. ROS-Mediated Signalling in Bacteria: Zinc-Containing Cys-X-X-Cys Redox Centres and Iron-Based Oxidative Stress. J. Signal Transduct. 2012, 2012, 605905. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, S.W.; King, J.; Cooney, C.L. The reactivity and oxidation pathway of cysteine 232 in recombinant human alpha 1-antitrypsin. J. Biol. Chem. 2002, 277, 25486–25492. [Google Scholar] [CrossRef] [PubMed]
- Ramelot, T.A.; Cort, J.R.; Goldsmith-Fischman, S.; Kornhaber, G.J.; Xiao, R.; Shastry, R.; Acton, T.B.; Honig, B.; Montelione, G.T.; Kennedy, M.A. Solution NMR structure of the iron-sulfur cluster assembly protein U (IscU) with zinc bound at the active site. J. Mol. Biol. 2004, 344, 567–583. [Google Scholar] [CrossRef]
- Chandrangsu, P.; Helmann, J.D. Intracellular Zn(II) Intoxication Leads to Dysregulation of the PerR Regulon Resulting in Heme Toxicity in Bacillus subtilis. PLoS Genet. 2016, 12, e1006515. [Google Scholar] [CrossRef] [PubMed]
- Krissinel, E.; Henrick, K. Inference of macromolecular assemblies from crystalline state. J. Mol. Biol. 2007, 372, 774–797. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Concentration (µM/µM as-Isolated SufU) |
|---|---|
| Fe | 0 |
| Ni | 0.1 |
| Zn | 1.1 ± 0.1 |
| Substrates | Km (µM) | Vmax (nmol/min/mg) | kcat (s−1) | kcat/Km (s−1 M−1) |
|---|---|---|---|---|
| Cysteine | 103 | 1250 | 0.94 | 9.1 × 103 |
| SufU | 7.8 | 909 | 0.68 | 8.7 × 104 |
| Element | Concentration (µM/µM Protein) |
|---|---|
| Fe | 3.4 ± 0.14 |
| S | 2.7 ± 0.18 |
| Zn | 1.1 ± 0.10 |
| Cysteine Desulfurase | IscS E. coli | SufSE E. coli | SufSU B. subtilis | SufSU S. aureus | SufSU E. faecalis | SufSU Mtb |
|---|---|---|---|---|---|---|
| Velocity (nmol/min/mg) | 90 | 600 | 1000 | 6 | 150 | 900 |
| Stimulating factor | / | 150 | 40–130 | 1.5 | 37 | 180 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elchennawi, I.; Carpentier, P.; Caux, C.; Ponge, M.; Ollagnier de Choudens, S. Structural and Biochemical Characterization of Mycobacterium tuberculosis Zinc SufU-SufS Complex. Biomolecules 2023, 13, 732. https://doi.org/10.3390/biom13050732
Elchennawi I, Carpentier P, Caux C, Ponge M, Ollagnier de Choudens S. Structural and Biochemical Characterization of Mycobacterium tuberculosis Zinc SufU-SufS Complex. Biomolecules. 2023; 13(5):732. https://doi.org/10.3390/biom13050732
Chicago/Turabian StyleElchennawi, Ingie, Philippe Carpentier, Christelle Caux, Marine Ponge, and Sandrine Ollagnier de Choudens. 2023. "Structural and Biochemical Characterization of Mycobacterium tuberculosis Zinc SufU-SufS Complex" Biomolecules 13, no. 5: 732. https://doi.org/10.3390/biom13050732