
Vutiglabridin Modulates Paraoxonase 1 and Ameliorates Diet-Induced Obesity in Hyperlipidemic Mice
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
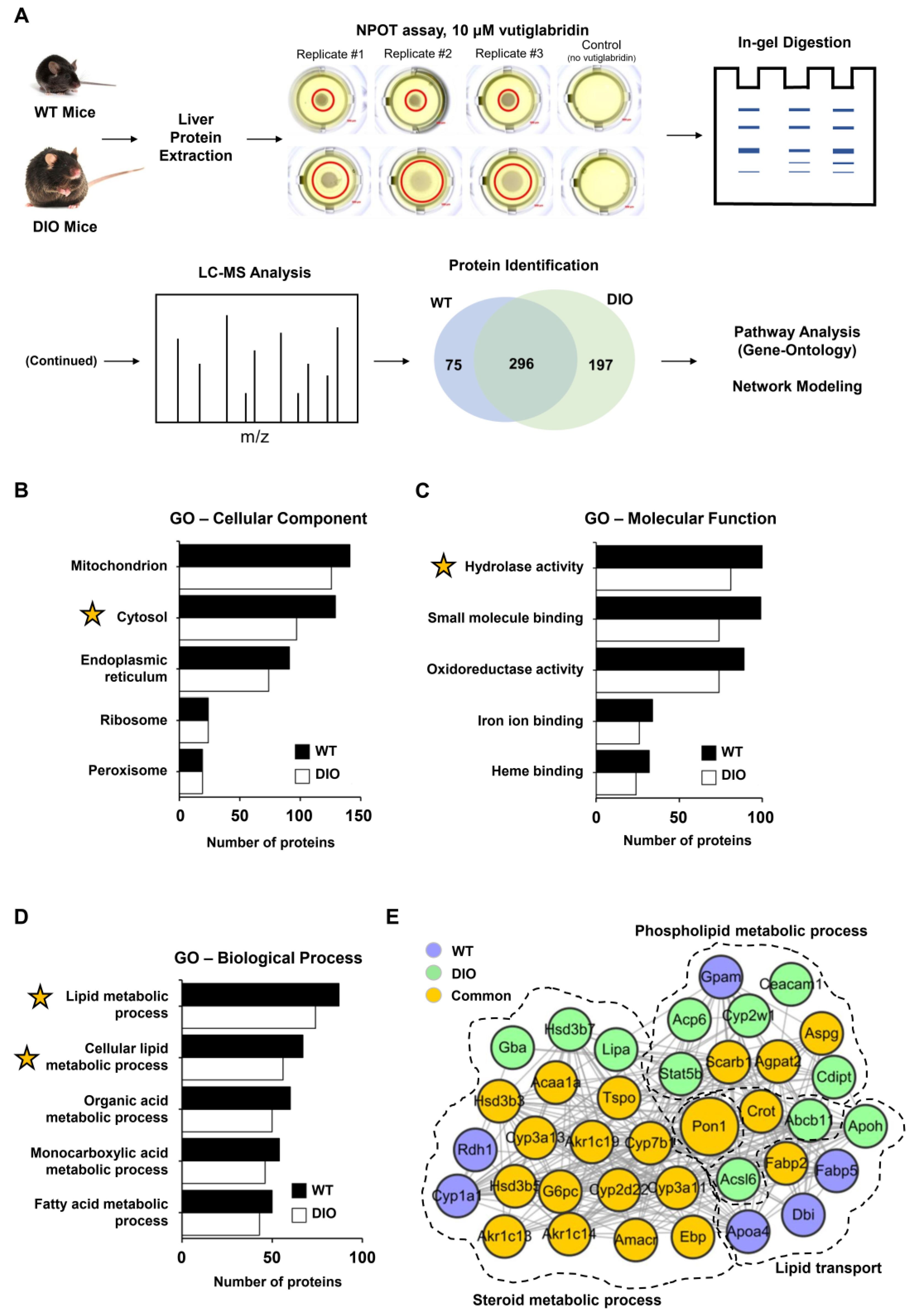
2.1. Nematic Protein Organisation Technique (NPOT®), Heteroassembly Isolation, and Proteomics
2.2. Bioinformatics Analysis
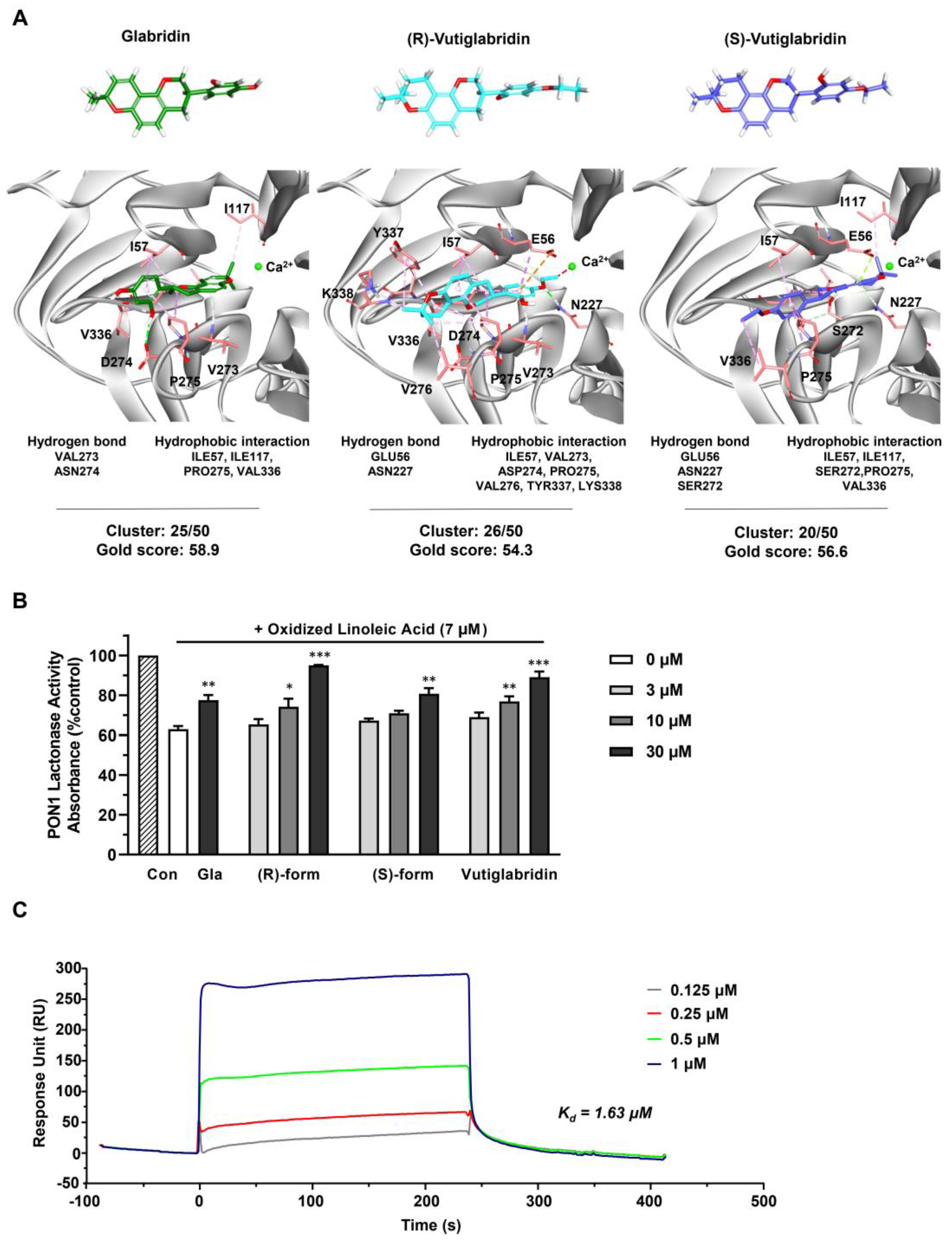
2.3. In Silico Molecular Docking Study
2.4. Recombinant PON1 Lactonase Activity Assay
2.5. Recombinant PON1 Lactonase Kinetic Assay
2.6. Surface Plasmon Resonance (SPR) Analysis
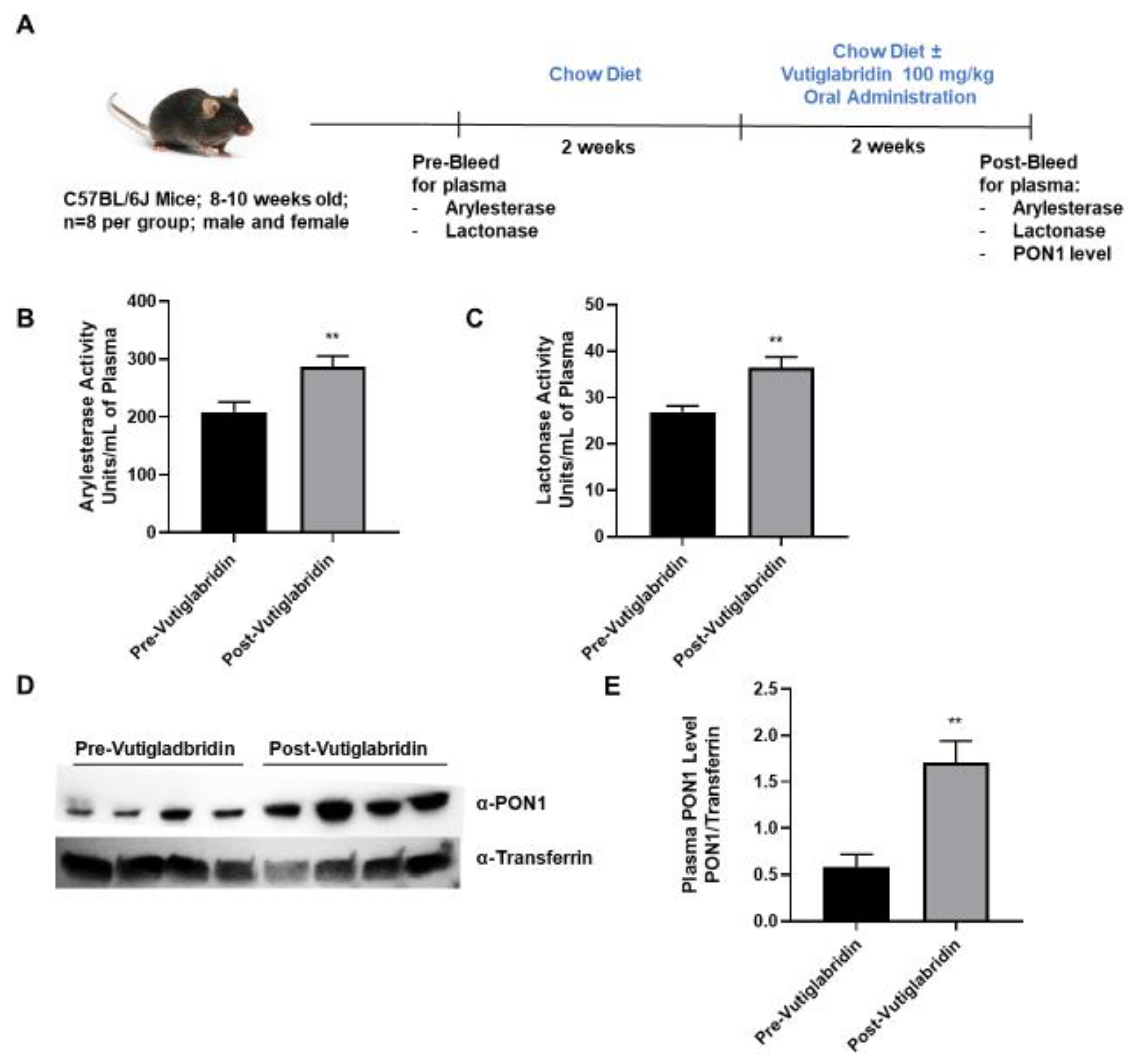
2.7. Mice and Diets
2.8. Plasma Lipids and Glucose Measurement
2.9. Body Composition Measurement
2.10. Immunoblotting
2.11. Lactonase, Arylesterase, and Paraoxonase Assays
2.12. Statistical Analysis
3. Results
3.1. Identification of Potential Target Proteins of Vutiglabridin through NPOT Assay
3.2. Vutiglabridin Binds to rePON1 and Increases Its Enzyme Activity under Oxidative Stress Conditions
3.3. Vutiglabridin Increases PON1 Activity and Expression in the Plasma of WT Mice
3.4. Vutiglabridin Protects against Diet-Induced Obesity in Hyperlipidemic LDLR−/− Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engin, A. The Definition and Prevalence of Obesity and Metabolic Syndrome. Adv. Exp. Med. Biol. 2017, 960, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Klop, B.; Elte, J.W.F.; Cabezas, M.C. Dyslipidemia in Obesity: Mechanisms and Potential Targets. Nutrients 2013, 5, 1218. [Google Scholar] [CrossRef] [PubMed]
- Rakhra, V.; Galappaththy, S.L.; Bulchandani, S.; Cabandugama, P.K. Obesity and the Western Diet: How We Got Here. Mo. Med. 2020, 117, 536. [Google Scholar] [PubMed]
- Boden, G. Obesity and Free Fatty Acids. Endocrinol. Metab. Clin. N. Am. 2008, 37, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.T.; Libby, P. The Unstable Atheroma. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1859–1867. [Google Scholar] [CrossRef]
- Rocha, V.Z.; Libby, P. Obesity, Inflammation, and Atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar] [CrossRef]
- Meneses, M.J.; Silvestre, R.; Sousa-Lima, I.; Macedo, M.P. Paraoxonase-1 as a Regulator of Glucose and Lipid Homeostasis: Impact on the Onset and Progression of Metabolic Disorders. Int. J. Mol. Sci. 2019, 20, 4049. [Google Scholar] [CrossRef]
- Véniant, M.M.; Zlot, C.H.; Walzem, R.L.; Pierotti, V.; Driscoll, R.; Dichek, D.; Herz, J.; Young, S.G. Lipoprotein Clearance Mechanisms in LDL Receptor-Deficient “Apo-B48-Only” and “Apo-B100-Only” Mice. J. Clin. Investig. 1998, 102, 1559–1568. [Google Scholar] [CrossRef]
- Fazio, S.; Linton, M.F. Mouse Models of Hyperlipidemia and Atherosclerosis. Front. Biosci. 2001, 6, D515–D525. [Google Scholar] [CrossRef]
- Chan, J.; Karere, G.M.; Cox, L.A.; VandeBerg, J.L. Animal Models of Diet-Induced Hypercholesterolemia. In Hypercholesterolemia; IntechOpen: London, UK, 2015. [Google Scholar] [CrossRef]
- Mackness, M.I.; Arrol, S.; Durrington, P.N. Paraoxonase Prevents Accumulation of Lipoperoxides in Low-Density Lipoprotein. FEBS Lett. 1991, 286, 152–154. [Google Scholar] [CrossRef]
- Gonzalvo, M.C.; Gil, F.; Hernandez, A.F.; Rodrigo, L.; Villanueva, E.; Pla, A. Human Liver Paraoxonase (PON1): Subcellular Distribution and Characterization. J. Biochem. Mol. Toxicol. 1998, 12, 61–69. [Google Scholar] [CrossRef]
- MacKness, M.I.; MacKness, B.; Arrol, S.; Wood, G.; Bhatnagar, D.; Durrington, P.N. Presence of Paraoxonase in Human Interstitial Fluid. FEBS Lett. 1997, 416, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Taler-Verčič, A.; Goličnik, M.; Bavec, A. The Structure and Function of Paraoxonase-1 and Its Comparison to Paraoxonase-2 and -3. Molecules 2020, 25, 5980. [Google Scholar] [CrossRef]
- Litvinov, D.; Mahini, H.; Garelnabi, M. Antioxidant and Anti-Inflammatory Role of Paraoxonase 1: Implication in Arteriosclerosis Diseases. N. Am. J. Med. Sci. 2012, 4, 523. [Google Scholar] [CrossRef] [PubMed]
- Mackness, M.I.; Arrol, S.; Abbott, C.; Durrington, P.N. Protection of Low-Density Lipoprotein against Oxidative Modification by High-Density Lipoprotein Associated Paraoxonase. Atherosclerosis 1993, 104, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Aviram, M.; Rosenblat, M.; Bisgaier, C.L.; Newton, R.S.; Primo-Parmo, S.L.; la Du, B.N. Paraoxonase Inhibits High-Density Lipoprotein Oxidation and Preserves Its Functions. A Possible Peroxidative Role for Paraoxonase. J. Clin. Investig. 1998, 101, 1581–1590. [Google Scholar] [CrossRef]
- Cervellati, C.; Bonaccorsi, G.; Trentini, A.; Valacchi, G.; Sanz, J.M.; Squerzanti, M.; Spagnolo, M.; Massari, L.; Crivellari, I.; Greco, P.; et al. Paraoxonase, Arylesterase and Lactonase Activities of Paraoxonase-1 (PON1) in Obese and Severely Obese Women. Scand. J. Clin. Lab. Investig. 2018, 78, 18–24. [Google Scholar] [CrossRef]
- Ferretti, G.; Bacchetti, T.; Moroni, C.; Savino, S.; Liuzzi, A.; Balzola, F.; Bicchiega, V. Paraoxonase Activity in High-Density Lipoproteins: A Comparison between Healthy and Obese Females. J. Clin. Endocrinol. Metab. 2005, 90, 1728–1733. [Google Scholar] [CrossRef]
- Costa, L.G.; Giordano, G.; Furlong, C.E. Pharmacological and Dietary Modulators of Paraoxonase 1 (PON1) Activity and Expression: The Hunt Goes on. Biochem. Pharmacol. 2011, 81, 337–344. [Google Scholar] [CrossRef]
- Choi, L.S.; Jo, I.G.; Kang, K.S.; Im, J.H.; Kim, J.; Kim, J.; Chung, J.W.; Yoo, S.K. Discovery and Preclinical Efficacy of HSG4112, a Synthetic Structural Analog of Glabridin, for the Treatment of Obesity. Int. J. Obes. 2020, 45, 130–142. [Google Scholar] [CrossRef]
- Simmler, C.; Pauli, G.F.; Chen, S.N. Phytochemistry and Biological Properties of Glabridin. Fitoterapia 2013, 90, 160–184. [Google Scholar] [CrossRef] [PubMed]
- Gee, M.B.; Smith, P.E. Kirkwood-Buff Theory of Molecular and Protein Association, Aggregation, and Cellular Crowding. J. Chem. Phys. 2009, 131, 10B613. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, S.; Boon, C.L. The Kirkwood-Buff Theory and the Effect of Cosolvents on Biochemical Reactions. J. Chem. Phys. 2004, 121, 9147–9155. [Google Scholar] [CrossRef] [PubMed]
- Kakhlon, O.; Vaknin, H.; Mishra, K.; D’Souza, J.; Marisat, M.; Sprecher, U.; Wald-Altman, S.; Dukhovny, A.; Raviv, Y.; Da’adoosh, B.; et al. Alleviation of a Polyglucosan Storage Disorder by Enhancement of Autophagic Glycogen Catabolism. EMBO Mol. Med. 2021, 13, e14554. [Google Scholar] [CrossRef]
- Wang, L.; Eftekhari, P.; Schachner, D.; Ignatova, I.D.; Palme, V.; Schilcher, N.; Ladurner, A.; Heiss, E.H.; Stangl, H.; Dirsch, V.M.; et al. Novel Interactomics Approach Identifies ABCA1 as Direct Target of Evodiamine, Which Increases Macrophage Cholesterol Efflux. Sci. Rep. 2018, 8, 11061. [Google Scholar] [CrossRef] [PubMed]
- Beyrath, J.; Pellegrini, M.; Renkema, H.; Houben, L.; Pecheritsyna, S.; van Zandvoort, P.; van den Broek, P.; Bekel, A.; Eftekhari, P.; Smeitink, J.A.M. KH176 Safeguards Mitochondrial Diseased Cells from Redox Stress-Induced Cell Death by Interacting with the Thioredoxin System/Peroxiredoxin Enzyme Machinery. Sci. Rep. 2018, 8, 6577. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and Validation of a Genetic Algorithm for Flexible Docking. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef]
- Verdonk, M.L.; Cole, J.C.; Hartshorn, M.J.; Murray, C.W.; Taylor, R.D. Improved Protein-Ligand Docking Using GOLD. Proteins 2003, 52, 609–623. [Google Scholar] [CrossRef]
- Petrič, B.; Goličnik, M.; Bavec, A. The Removal of Time-Concentration Data Points from Progress Curves Improves the Determination of K(m): The Example of Paraoxonase 1. Molecules 2022, 27, 1306. [Google Scholar] [CrossRef]
- Petrič, B.; Goličnik, M.; Bavec, A. iFIT: An automated web tool for determining enzyme-kinetic parameters based on the high-curvature region of progress curves. Acta Chim. Slov. 2022, 69, 478–482. [Google Scholar] [CrossRef]
- Shih, D.M.; Meng, Y.; Sallam, T.; Vergnes, L.; Shu, M.L.; Reue, K.; Tontonoz, P.; Fogelman, A.M.; Lusis, A.J.; Reddy, S.T. PON2 Deficiency Leads to Increased Susceptibility to Diet-Induced Obesity. Antioxidants 2019, 8, 19. [Google Scholar] [CrossRef]
- Aviram, M.; Rosenblat, M. Paraoxonases (PON1, PON2, PON3) Analyses in Vitro and in Vivo in Relation to Cardiovascular Diseases. Methods Mol. Biol. 2008, 477, 259–276. [Google Scholar] [CrossRef] [PubMed]
- Humbert, R.; Adler, D.A.; Disteche, C.M.; Hassett, C.; Omiecinski, C.J.; Furlong, C.E. The Molecular Basis of the Human Serum Paraoxonase Activity Polymorphism. Nat. Genet 1993, 3, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Charles-Schoeman, C.; Wang, J.; Shahbazian, A.; Lee, Y.Y.; Wang, X.; Grijalva, V.; Brahn, E.; Shih, D.M.; Devarajan, A.; Montano, C.; et al. Suppression of inflammatory arthritis in human serum paraoxonase 1 transgenic mice. Sci. Rep. 2020, 10, 16848. [Google Scholar] [CrossRef]
- Atrahimovich, D.; Vaya, J.; Tavori, H.; Khatib, S. Glabridin Protects Paraoxonase 1 from Linoleic Acid Hydroperoxide Inhibition via Specific Interaction: A Fluorescence-Quenching Study. J. Agric. Food Chem. 2012, 60, 3679–3685. [Google Scholar] [CrossRef] [PubMed]
- Tavori, H.; Rosenblat, M.; Vaya, J.; Aviram, M. Paraoxonase 1 Interactions with Atherosclerotic Lesions and Arterial Macrophages Protect against Foam Cell Formation and Atherosclerosis Development. Clin. Lipidol. 2010, 5, 685–697. [Google Scholar] [CrossRef]
- Jung, H.J.; Kwon, H.J. Target Deconvolution of Bioactive Small Molecules: The Heart of Chemical Biology and Drug Discovery. Arch. Pharm. Res. 2015, 38, 1627–1641. [Google Scholar] [CrossRef]
- Primo-Parmo, S.L.; Sorenson, R.C.; Teiber, J.; la Du, B.N. The Human Serum Paraoxonase/Arylesterase Gene (PON1) Is One Member of a Multigene Family. Genomics 1996, 33, 498–507. [Google Scholar] [CrossRef]
- Draganov, D.I.; Stetson, P.L.; Watson, C.E.; Billecke, S.S.; la Du, B.N. Rabbit Serum Paraoxonase 3 (PON3) Is a High Density Lipoprotein-Associated Lactonase and Protects Low Density Lipoprotein against Oxidation. J. Biol. Chem. 2000, 275, 33435–33442. [Google Scholar] [CrossRef]
- Priyanka, K.; Singh, S.; Gill, K. Paraoxonase 3: Structure and Its Role in Pathophysiology of Coronary Artery Disease. Biomolecules 2019, 9, 817. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PON1 | |||
|---|---|---|---|
| - | OX-LA (7µM) | ||
| - | Vutiglabridin (30 µM) | ||
| Km (µM) | 155.78 | 102.11 | 134.52 |
| Vmax (µM/min) | 92.34 | 46.71 | 69.14 |
| Vmax/Km | 0.59 | 0.46 | 0.51 |
| Fold change of Vmax/Km | 1 | 0.77 | 0.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sulaiman, D.; Choi, L.S.; Lee, H.M.; Shin, J.; Kim, D.H.; Lee, K.W.; Eftekhari, P.; Quartier, A.; Park, H.S.; Reddy, S.T. Vutiglabridin Modulates Paraoxonase 1 and Ameliorates Diet-Induced Obesity in Hyperlipidemic Mice. Biomolecules 2023, 13, 687. https://doi.org/10.3390/biom13040687
Sulaiman D, Choi LS, Lee HM, Shin J, Kim DH, Lee KW, Eftekhari P, Quartier A, Park HS, Reddy ST. Vutiglabridin Modulates Paraoxonase 1 and Ameliorates Diet-Induced Obesity in Hyperlipidemic Mice. Biomolecules. 2023; 13(4):687. https://doi.org/10.3390/biom13040687
Chicago/Turabian StyleSulaiman, Dawoud, Leo Sungwong Choi, Hyeong Min Lee, Jaejin Shin, Dong Hwan Kim, Keun Woo Lee, Pierre Eftekhari, Angélique Quartier, Hyung Soon Park, and Srinivasa T. Reddy. 2023. "Vutiglabridin Modulates Paraoxonase 1 and Ameliorates Diet-Induced Obesity in Hyperlipidemic Mice" Biomolecules 13, no. 4: 687. https://doi.org/10.3390/biom13040687