
Cu+/Ag+ Competition in Type I Copper Proteins (T1Cu)
 , ,
, , 
Abstract
:
1. Introduction
2. Methods
2.1. DFT Methodology
2.2. Calibration and Validation of the Chosen Methodology
3. Results and Discussion
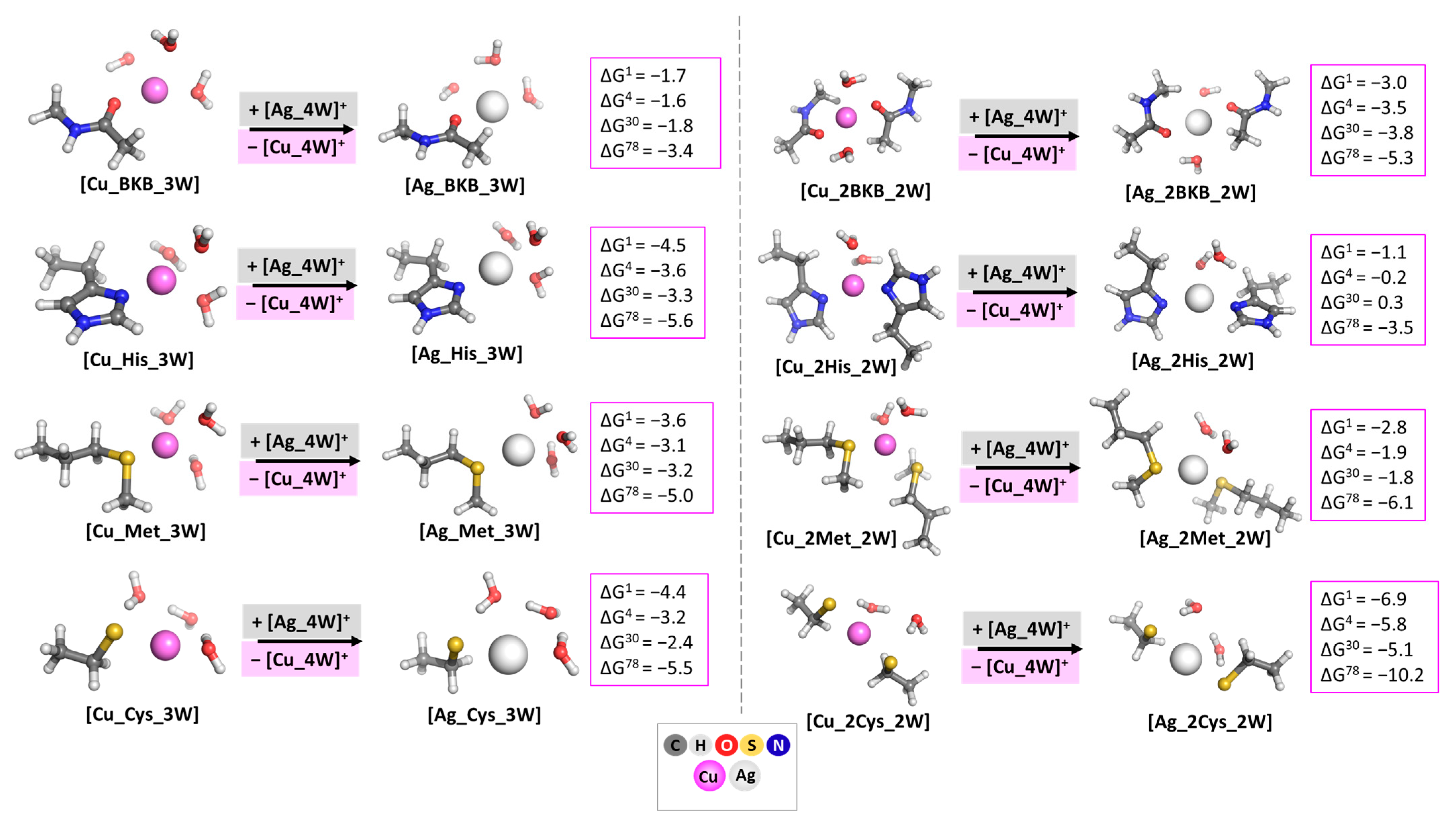
3.1. Evaluating the Cu+/Ag+ Competition in Model Systems with One and Two Identical Amino Acid Residues
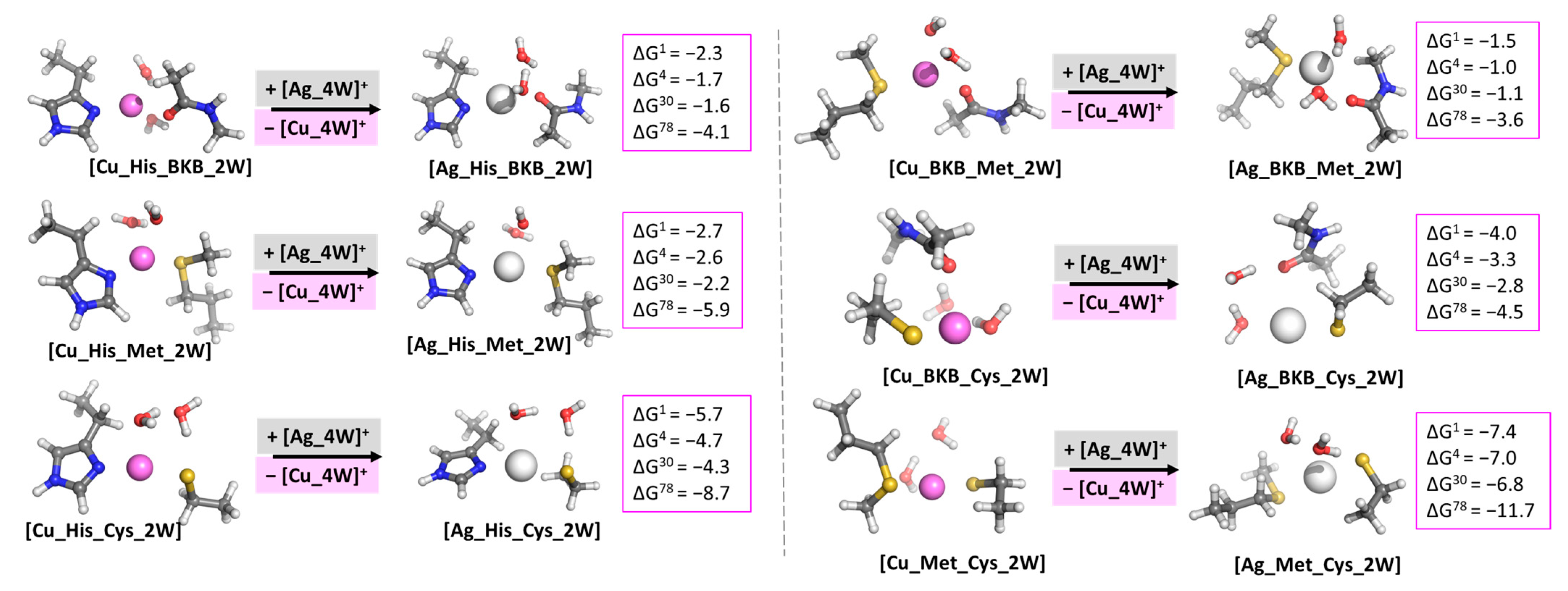
3.2. Evaluating the Cu+/Ag+ Competition in Model Systems with Two Different Amino acid Residues
3.3. Evaluating the Cu+/Ag+ Competition in Model Systems with Two Identical His and a Different Amino Acid Residue (Model of the Trigonal Planar Structure in T1Cu)
3.4. Evaluating the Cu+/Ag+ Competition in Model Systems of the Metal-Binding Centers in Plastocyanin (Class 1), Ceruloplasmin (Class 2), and Azurin (Class 3)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bhamidimarri, S.P.; Young, T.R.; Shanmugam, M.; Soderholm, S.; Basle, A.; Bumann, D.; Van Den Berg, B. Acquisition of Ionic Copper by the Bacterial Outer Membrane Protein OprC through a Novel Binding Site. PLoS Biol. 2021, 19, e3001446. [Google Scholar] [CrossRef] [PubMed]
- Rubino, J.T.; Franz, K.J. Coordination Chemistry of Copper Proteins: How Nature Handles a Toxic Cargo for Essential Function. J. Inorg. Biochem. 2012, 107, 129–143. [Google Scholar] [CrossRef]
- Williams, R.J.P. Energised (Entatic) States of Groups and of Secondary Structures in Proteins and Metalloproteins. Eur. J. Biochem. 1995, 234, 363–381. [Google Scholar] [CrossRef] [PubMed]
- Vallee, B.L.; Williams, R.J.P. Metalloenzymes: The Entatic Nature of Their Active Sites. Proc. Natl. Acad. Sci. USA 1968, 59, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Shannon, R.D. Revised Effective Ionic Radii and Systematic Studies of Interatomic Distances in Halides and Chalcogenides. Acta Crystallogr. Sect. A 1976, 32, 751–767. [Google Scholar] [CrossRef]
- Panzner, M.J.; Bilinovich, S.M.; Parker, J.A.; Bladholm, E.L.; Ziegler, C.J.; Berry, S.M.; Leeper, T.C. Isomorphic Deactivation of a Pseudomonas Aeruginosa Oxidoreductase: The Crystal Structure of Ag(I) Metallated Azurin at 1.7 Å. J. Inorg. Biochem. 2013, 128, 11–16. [Google Scholar] [CrossRef]
- Blasie, C.A.; Berg, J.M. Kinetics and Thermodynamics of Copper (II) Binding to Apoazurin. J. Am. Chem. Soc. 2003, 1, 6866–6867. [Google Scholar] [CrossRef]
- Kędziora, A.; Speruda, M.; Krzyżewska, E.; Rybka, J.; Łukowiak, A.; Bugla-Płoskońska, G. Similarities and Differences between Silver Ions and Silver in Nanoforms as Antibacterial Agents. Int. J. Mol. Sci. 2018, 19, 444. [Google Scholar] [CrossRef]
- Eckhardt, S.; Brunetto, P.S.; Gagnon, J.; Priebe, M.; Giese, B.; Fromm, K.M. Nanobio Silver: Its Interactions with Peptides and Bacteria, and Its Uses in Medicine. Chem. Rev. 2013, 113, 4708–4754. [Google Scholar] [CrossRef] [PubMed]
- Li, W.R.; Xie, X.B.; Shi, Q.S.; Zeng, H.Y.; Ou-Yang, Y.S.; Chen, Y. Ben Antibacterial Activity and Mechanism of Silver Nanoparticles on Escherichia Coli. Appl. Microbiol. Biotechnol. 2010, 85, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Hecel, A.; Kolkowska, P.; Krzywoszynska, K.; Szebesczyk, A.; Rowinska-Zyrek, M.; Kozlowski, H. Ag+ Complexes as Potential Therapeutic Agents in Medicine and Pharmacy. Curr. Med. Chem. 2019, 26, 624–647. [Google Scholar] [PubMed]
- Betts, H.D.; Whitehead, C.; Harris, H.H. Silver in Biology and Medicine: Opportunities for Metallomics Researchers. Metallomics 2021, 13, mfaa001. [Google Scholar] [CrossRef] [PubMed]
- Lachiewicz, A.M.; Hauck, C.G.; Weber, D.J.; Cairns, B.A.; Van Duin, D. Bacterial Infections after Burn Injuries: Impact of Multidrug Resistance. Clin. Infect. Dis. 2017, 65, 2130–2136. [Google Scholar] [CrossRef] [PubMed]
- Mussin, J.E.; Roldán, M.V.; Rojas, F.; Sosa, M.d.l.Á.; Pellegri, N.; Giusiano, G. Antifungal Activity of Silver Nanoparticles in Combination with Ketoconazole against Malassezia Furfur. AMB Express 2019, 9, 131–140. [Google Scholar] [CrossRef]
- Jo, Y.K.; Kim, B.H.; Jung, G. Antifungal Activity of Silver Ions and Nanoparticles on Phytopathogenic Fungi. Plant Dis. 2009, 93, 1037–1043. [Google Scholar] [CrossRef]
- Galdiero, S.; Falanga, A.; Vitiello, M.; Cantisani, M.; Marra, V.; Galdiero, M. Silver Nanoparticles as Potential Antiviral Agents. Molecules 2011, 16, 8894–8918. [Google Scholar] [CrossRef]
- Kircheva, N.; Dobrev, S.; Nikolova, V.; Angelova, S.; Dudev, T. Theoretical Insight into the Phosphate-Targeted Silver’s Antibacterial Action: Differentiation between Gram (+) and Gram (−) Bacteria. Inorg. Chem. 2022, 61, 10089–10100. [Google Scholar] [CrossRef]
- Dobrev, S.; Kircheva, N.; Nikolova, V.; Angelova, S.; Dudev, T. Competition between Ag+ and Ni2+ in Nickel Enzymes: Implications for the Ag+ Antibacterial Activity. Comput. Biol. Chem. 2022, 101, 107785. [Google Scholar] [CrossRef]
- Puchkova, L.V.; Broggini, M.; Polishchuk, E.V.; Ilyechova, E.Y.; Polishchuk, R.S. Silver Ions as a Tool for Understanding Different Aspects of Copper Metabolism. Nutrients 2019, 11, 1364. [Google Scholar] [CrossRef]
- Veronesi, G.; Gallon, T.; Deniaud, A.; Boff, B.; Gateau, C.; Lebrun, C.; Vidaud, C.; Rollin-Genetet, F.; Carrière, M.; Kieffer, I.; et al. XAS Investigation of Silver(I) Coordination in Copper(I) Biological Binding Sites. Inorg. Chem. 2015, 54, 11688–11696. [Google Scholar] [CrossRef]
- Viček, A. Electron Tunneling in Single Crystals of Pseudomonas Aeruginosa Azurins. J. Am. Chem. Soc. 2001, 123, 11623–11631. [Google Scholar]
- Choi, M.; Davidson, V.L. Cupredoxins—A Study of How Proteins May Evolve to Use Metals for Bioenergetic Processes. Metallomics 2011, 3, 140–151. [Google Scholar] [CrossRef]
- Latorre, R.; Baez-nieto, D. Encyclopedia of Biophysics; Springer: Berlin, Heidelberg, 2013; ISBN 9783642167126. [Google Scholar]
- Colman, P.M.; Freeman, H.C.; Guss, J.M.; Murata, M.; Norris, V.A.; Ramshaw, J.A.M.; Venkatappa, M.P. X-ray Crystal Structure Analysis of Plastocyanin at 2.7 A Resolution. Nature 1978, 272, 319–324. [Google Scholar] [CrossRef]
- Ilyechova, E.Y.; Saveliev, A.N.; Skvortsov, A.N.; Babich, P.S.; Zatulovskaia, Y.A.; Pliss, M.G.; Korzhevskii, D.E.; Tsymbalenko, N.V.; Puchkova, L.V. The Effects of Silver Ions on Copper Metabolism in Rats. Metallomics 2014, 6, 1970–1987. [Google Scholar] [CrossRef] [PubMed]
- Nar, H.; Messerschmidt, A.; Huber, R.; van de Kamp, M.; Canters, G.W. Crystal Structure Analysis of Oxidized Pseudomonas Aeruginosa Azurin at PH 5·5 and PH 9·0. A PH-Induced Conformational Transition Involves a Peptide Bond Flip. J. Mol. Biol. 1991, 221, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Solomon, E.I.; Szilagyi, R.K.; DeBeer George, S.; Basumallick, L. Electronic Structures of Metal Sites in Proteins and Models: Contributions to Function in Blue Copper Proteins. Chem. Rev. 2004, 104, 419–458. [Google Scholar] [CrossRef]
- Holm, R.H.; Kennepohl, P.; Solomon, E.I. Structural and Functional Aspects of Metal Sites in Biology. Chem. Rev. 1996, 96, 2239–2314. [Google Scholar] [CrossRef]
- Dudev, T.; Lim, C. Competition among Metal Ions for Protein Binding Sites: Determinants of Metal Ion Selectivity in Proteins. Chem. Rev. 2014, 114, 538–556. [Google Scholar] [CrossRef]
- Dudev, T.; Grauffel, C.; Hsu, S.T.D.; Lim, C. How Native and Non-Native Cations Bind and Modulate the Properties of GTP/ATP. J. Chem. Theory Comput. 2018, 14, 3311–3320. [Google Scholar] [CrossRef]
- Dudev, T.; Lim, C. Competition between Li+ and Mg2+ in Metalloproteins. Implications for Lithium Therapy. J. Am. Chem. Soc. 2011, 133, 9506–9515. [Google Scholar] [CrossRef]
- Dudev, T.; Chang, L.Y.; Lim, C. Factors Governing the Substitution of La3+ for Ca2+ and Mg2+ in Metalloproteins: A DFT/CDM Study. J. Am. Chem. Soc. 2005, 127, 4091–4103. [Google Scholar] [CrossRef] [PubMed]
- Kircheva, N.; Toshev, N.; Dudev, T. Holo-Chromodulin: Competition between the Native Cr3+ and Other Biogenic Cations (Fe3+, Fe2+, Mg2+, and Zn2+) for the Binding Sites. Metallomics 2022, 14, mfac082. [Google Scholar] [CrossRef] [PubMed]
- Kircheva, N.; Dudev, T. Gallium as an Antibacterial Agent: A DFT/SMD Study of the Ga3+/Fe3+ Competition for Binding Bacterial Siderophores. Inorg. Chem. 2020, 59, 6242–6254. [Google Scholar] [CrossRef] [PubMed]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal Solvation Model Based on Solute Electron Density and on a Continuum Model of the Solvent Defined by the Bulk Dielectric Constant and Atomic Surface Tensions. J.Phys.Chem B 2009, 113, 6378–6396. [Google Scholar] [CrossRef] [PubMed]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Montgomery, J.A., Jr.; Vreven, T.; Kudin, K.N.; Burant, J.C.; et al. Gaussian 09; Gaussian Inc.: Wallingford, CT, USA, 2013. [Google Scholar]
- Andrae, D.; Häußermann, U.; Dolg, M.; Stoll, H.; Preuß, H. Energy-Adjusted Ab Initio Pseudopotentials for the Second and Third Row Transition Elements. Theor. Chim. Acta 1990, 77, 123–141. [Google Scholar] [CrossRef]
- The PyMol Molecular Graphics System, Version 2.2.3; Schrödinger, LLC: New York, NY, USA, 2018.
- Betts, H.D.; Neville, S.L.; McDevitt, C.A.; Sumby, C.J.; Harris, H.H. The Biochemical Fate of Ag+ Ions in Staphylococcus Aureus, Escherichia Coli, and Biological Media. J. Inorg. Biochem. 2021, 225, 111598. [Google Scholar] [CrossRef]
- Penner-Hahn, J.E. Characterization of “Spectroscopically Quiet” Metals in Biology. Coord. Chem. Rev. 2005, 249, 161–177. [Google Scholar] [CrossRef]
- Klotchenko, S.A.; Tsymbalenko, N.V.; Solov’ev, K.V.; Skvortsov, A.N.; Zatulovskii, E.A.; Babich, P.S.; Platonova, N.A.; Shavlovskii, M.M.; Puchkova, L.V.; Broggini, M. The Effect of Silver Ions on Copper Metabolism and Expression of Genes Encoding Copper Transport Proteins in Rat Liver. Dokl. Biochem. Biophys. 2008, 418, 24–27. [Google Scholar] [CrossRef]
- Vermes, I.; Dohanics, J.; Toth, G.; Pongracz, J. Maturation of the Circadian Rhythm of the Adrenocortical Functions in Human Neonates and Infants. Horm. Res. 1980, 12, 237–244. [Google Scholar] [CrossRef]
- Hill, C.H.; Starcher, B.; Matrone, G. Mercury and Silver Interrelationships with Copper. J. Nutr. 1964, 83, 107–110. [Google Scholar] [CrossRef]
- Persson, I.; Penner-Hahn, J.E.; Hodgson, K.O. An EXAFS Spectroscopic Study of Solvates of Copper(I) and Copper(II) in Acetonitrile, Dimethyl Sulfoxide, Pyridine, and Tetrahydrothiophene Solutions and a Large-Angle X-ray Scattering Study of the Copper(II) Acetonitrile Solvate in Solution. Inorg. Chem. 1993, 32, 2497–2501. [Google Scholar] [CrossRef]
- Persson, I. Structures of Hydrated Metal Ions in Solid State and Aqueous Solution. Liquids 2022, 2, 210–242. [Google Scholar] [CrossRef]
- Burda, J.V.; Pavelka, M.; Šimánek, M. Theoretical Model of Copper Cu(I)/Cu(II) Hydration. DFT and Ab Initio Quantum Chemical Study. J. Mol. Struct. THEOCHEM 2004, 683, 183–193. [Google Scholar] [CrossRef]
- Lee, H.M.; Min, S.K.; Lee, E.C.; Min, J.H.; Odde, S.; Kim, K.S. Hydrated Copper and Gold Monovalent Cations: Ab Initio Study. J. Chem. Phys. 2005, 122, 064314. [Google Scholar] [CrossRef]
- Persson, I.; Nilsson, K.B. Coordination Chemistry of the Solvated Silver(I) Ion in the Oxygen Donor Solvents Water, Dimethyl Sulfoxide, and N,N′-Dimethylpropyleneurea. Inorg. Chem. 2006, 45, 7428–7434. [Google Scholar] [CrossRef] [PubMed]
- Texter, J.; Hastreiter, J.J.; Hall, J.L. Spectroscopic Confirmation of the Tetrahedral Geometry of Ag(H2O)4+. J. Phys. Chem. 1983, 87, 4690–4693. [Google Scholar] [CrossRef]
- Johansson, G. Structures of Complexes in Solution Derived from X-ray Diffraction Measurements. Adv. Inorg. Chem. 1992, 39, 159–232. [Google Scholar]
- Busato, M.; Melchior, A.; Migliorati, V.; Colella, A.; Persson, I.; Mancini, G.; Veclani, D.; D’Angelo, P. Elusive Coordination of the Ag+Ion in Aqueous Solution: Evidence for a Linear Structure. Inorg. Chem. 2020, 59, 17291–17302. [Google Scholar] [CrossRef]
- Marcus, Y. Thermodynamics of Solvation of Ions. J. Chem. Soc. Faraday Trans. 1991, 89, 2995–2999. [Google Scholar] [CrossRef]
- Awoonor-williams, E.; Rowley, C.N. Evaluation of Methods for the Calculation of the PKa of Cysteine Residues in Proteins. J. Chem. Theory Comput. 2016, 12, 4662–4673. [Google Scholar] [CrossRef]
- Bulaj, G.; Kortemme, T.; Goldenberg, D.P. Ionization—Reactivity Relationships for Cysteine Thiols in Polypeptides. Biochemistry 1998, 37, 8965–8972. [Google Scholar] [CrossRef] [PubMed]
- Van Campen, D.R. Effects of Zinc, Cadmium, Silver and Mercury on the Absorption and Distribution of Copper-64 in Rats. J. Nutr. 1966, 88, 125–130. [Google Scholar] [CrossRef]
- Marenich, A.V.; Jerome, S.V.; Cramer, C.J.; Truhlar, D.G. Charge Model 5: An Extension of Hirshfeld Population Analysis for the Accurate Description of Molecular Interactions in Gaseous and Condensed Phases. J. Chem. Theory Comput. 2012, 8, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Hirshfeld, F.L. Bonded-Atom Fragments for Describing Molecular Charge Densities. Theor. Chim. Acta 1977, 44, 129–138. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bond Length in Å/ Method | M+-N (His1) | M+-N (His2) | M+-S (Cys) | M+-S (Met) | M+-O (BKB) | |
|---|---|---|---|---|---|---|
| Experiment | ∆(Ag+-Cu+) | 0.33 | 0.29 | 0.14 | 0.22 | 0.22 |
| B3LYP/6-31+G(3d,p)-D3 | ∆(Ag+-Cu+) | 0.07 | 0.84 | 0.18 | 0.00 | 0.55 |
| M062X/6-31+G(d,p) | ∆(Ag+-Cu+) | 0.35 | 0.19 | 0.16 | 0.33 | 0.07 |
| M062X/6-311++(d,p) | ∆(Ag+-Cu+) | 0.33 | 0.19 | 0.15 | 0.30 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kircheva, N.; Angelova, S.; Dobrev, S.; Petkova, V.; Nikolova, V.; Dudev, T. Cu+/Ag+ Competition in Type I Copper Proteins (T1Cu). Biomolecules 2023, 13, 681. https://doi.org/10.3390/biom13040681
Kircheva N, Angelova S, Dobrev S, Petkova V, Nikolova V, Dudev T. Cu+/Ag+ Competition in Type I Copper Proteins (T1Cu). Biomolecules. 2023; 13(4):681. https://doi.org/10.3390/biom13040681
Chicago/Turabian StyleKircheva, Nikoleta, Silvia Angelova, Stefan Dobrev, Vladislava Petkova, Valya Nikolova, and Todor Dudev. 2023. "Cu+/Ag+ Competition in Type I Copper Proteins (T1Cu)" Biomolecules 13, no. 4: 681. https://doi.org/10.3390/biom13040681