
Zinc Modulation of Neuronal Calcium Sensor Proteins: Three Modes of Interaction with Different Structural Outcomes

 , , , , ,
, , , , ,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of NCS Proteins
2.2. Isothermal Titration Calorimetry (ITC)
2.3. Nano Differential Scanning Fluorimetry (nanoDSF)
2.4. Thermal Shift Assay
2.5. Dynamic Light Scattering (DLS)
2.6. Structural Bioinformatics Predictions
2.7. Data Analysis and Visualization
3. Results
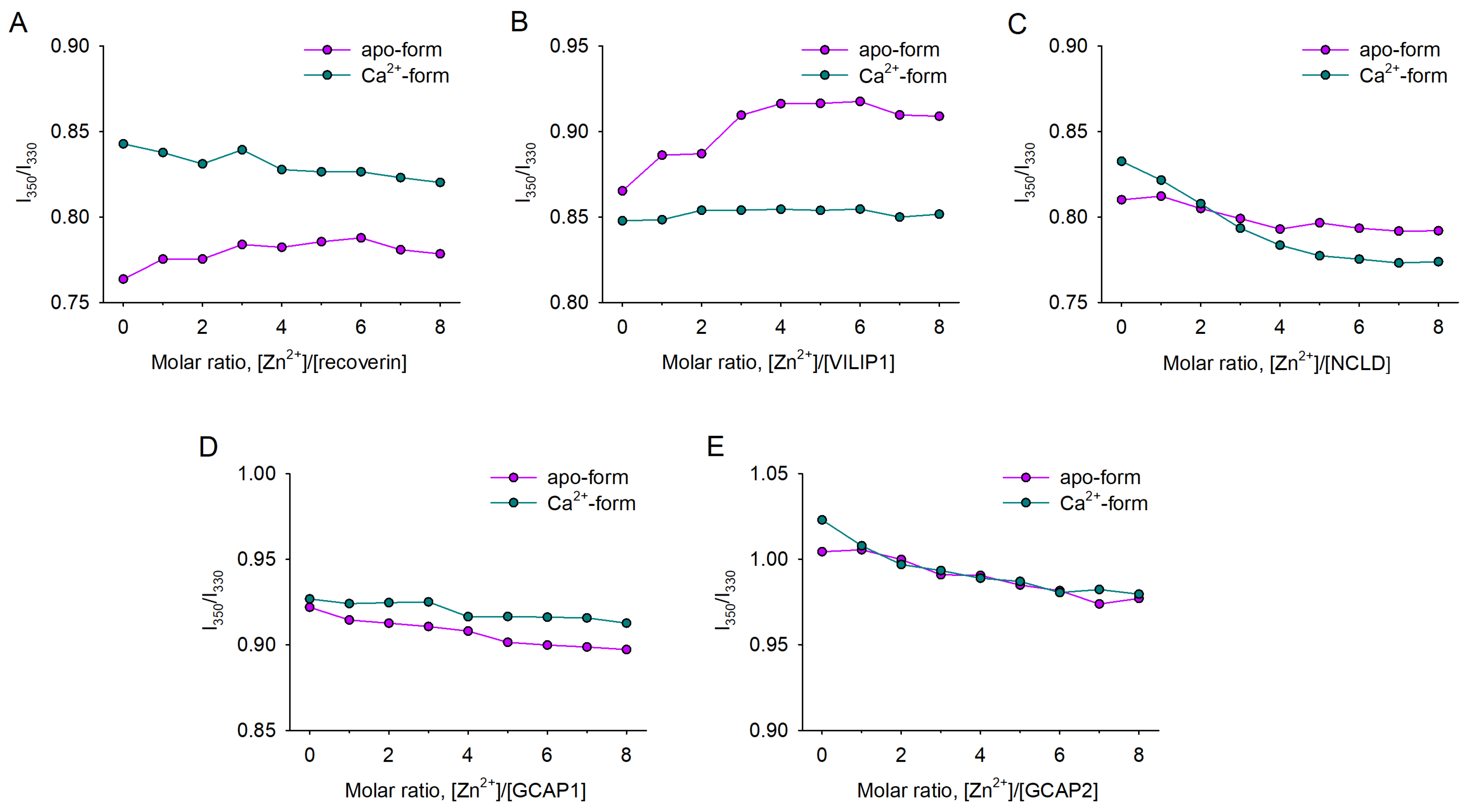
3.1. Coordination of Zn2+ by NCS Proteins
3.2. Zn2+-Induced Conformational Changes in NCS Proteins
3.3. Thermal Stability of Zn2+-bound NCS Proteins
3.4. Zn2+-Dependent Aggregation and Oligomerization of NCS Proteins
3.5. Potential Zn2+-Binding Sites of NCS Proteins
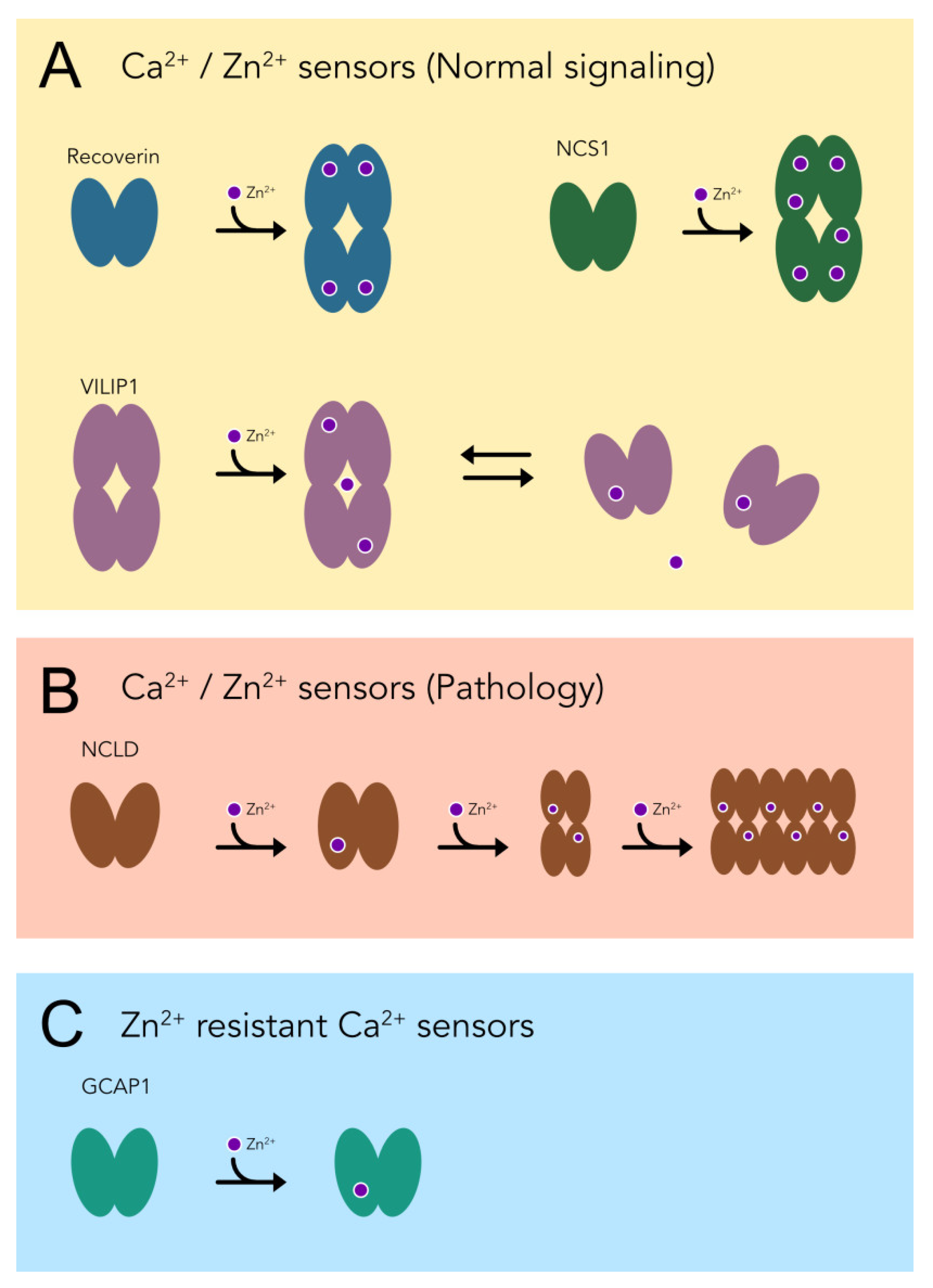
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Burgoyne, R.D. Neuronal calcium sensor proteins: Generating diversity in neuronal Ca2+ signalling. Nat. Rev. Neurosci. 2007, 8, 182–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pchitskaya, E.; Popugaeva, E.; Bezprozvanny, I. Calcium signaling and molecular mechanisms underlying neurodegenerative diseases. Cell Calcium 2018, 70, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.; Hideyama, T.; Yamashita, T.; Aizawa, H. AMPA receptor-mediated neuronal death in sporadic ALS. Neuropathol. Off. J. Jpn. Soc. Neuropathol. 2010, 30, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Zang, J.; Neuhauss, S.C.F. The Binding Properties and Physiological Functions of Recoverin. Front. Mol. Neurosci. 2018, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.W.; Dell’orco, D. A calcium-relay mechanism in vertebrate phototransduction. ACS Chem. Neurosci. 2013, 4, 909–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunewell, K.H.; Klein-Szanto, A.J. Visinin-like proteins (VSNLs): Interaction partners and emerging functions in signal transduction of a subfamily of neuronal Ca2+ -sensor proteins. Cell Tissue Res. 2009, 335, 301–316. [Google Scholar] [CrossRef]
- Burgoyne, R.D.; Helassa, N.; McCue, H.V.; Haynes, L.P. Calcium Sensors in Neuronal Function and Dysfunction. Cold Spring Harb. Perspect. Biol. 2019, 11, a035154. [Google Scholar] [CrossRef]
- Liu, M.; Yang, H.; Fang, D.; Yang, J.J.; Cai, J.; Wan, Y.; Chui, D.H.; Han, J.S.; Xing, G.G. Upregulation of P2X3 receptors by neuronal calcium sensor protein VILIP-1 in dorsal root ganglions contributes to the bone cancer pain in rats. Pain 2013, 154, 1551–1568. [Google Scholar] [CrossRef]
- Rivas, M.; Villar, D.; Gonzalez, P.; Dopazo, X.M.; Mellstrom, B.; Naranjo, J.R. Building the DREAM interactome. Sci. China. Life Sci. 2011, 54, 786–792. [Google Scholar] [CrossRef] [Green Version]
- Bandura, J.; Feng, Z.P. Current Understanding of the Role of Neuronal Calcium Sensor 1 in Neurological Disorders. Mol. Neurobiol. 2019, 56, 6080–6094. [Google Scholar] [CrossRef]
- Boeckel, G.R.; Ehrlich, B.E. NCS-1 is a regulator of calcium signaling in health and disease. Biochim. Biophys. Acta. Mol. Cell Res. 2018, 1865, 1660–1667. [Google Scholar] [CrossRef] [PubMed]
- Schnurra, I.; Bernstein, H.G.; Riederer, P.; Braunewell, K.H. The neuronal calcium sensor protein VILIP-1 is associated with amyloid plaques and extracellular tangles in Alzheimer’s disease and promotes cell death and tau phosphorylation in vitro: A link between calcium sensors and Alzheimer’s disease? Neurobiol. Dis. 2001, 8, 900–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGrowder, D.A.; Miller, F.; Vaz, K.; Nwokocha, C.; Wilson-Clarke, C.; Anderson-Cross, M.; Brown, J.; Anderson-Jackson, L.; Williams, L.; Latore, L.; et al. Cerebrospinal Fluid Biomarkers of Alzheimer’s Disease: Current Evidence and Future Perspectives. Brain Sci. 2021, 11, 215. [Google Scholar] [CrossRef] [PubMed]
- Liebl, M.P.; Kaya, A.M.; Tenzer, S.; Mittenzwei, R.; Koziollek-Drechsler, I.; Schild, H.; Moosmann, B.; Behl, C.; Clement, A.M. Dimerization of visinin-like protein 1 is regulated by oxidative stress and calcium and is a pathological hallmark of amyotrophic lateral sclerosis. Free Radic. Biol. Med. 2014, 72, 41–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riessland, M.; Kaczmarek, A.; Schneider, S.; Swoboda, K.J.; Lohr, H.; Bradler, C.; Grysko, V.; Dimitriadi, M.; Hosseinibarkooie, S.; Torres-Benito, L.; et al. Neurocalcin Delta Suppression Protects against Spinal Muscular Atrophy in Humans and across Species by Restoring Impaired Endocytosis. Am. J. Hum. Genet. 2017, 100, 297–315. [Google Scholar] [CrossRef] [Green Version]
- Helassa, N.; Antonyuk, S.V.; Lian, L.Y.; Haynes, L.P.; Burgoyne, R.D. Biophysical and functional characterization of hippocalcin mutants responsible for human dystonia. Hum. Mol. Genet. 2017, 26, 2426–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez-Hurtado, A.; Peraza, D.A.; Cercos, P.; Lagartera, L.; Gonzalez, P.; Dopazo, X.M.; Herranz, R.; Gonzalez, T.; Martin-Martinez, M.; Mellstrom, B.; et al. Targeting the neuronal calcium sensor DREAM with small-molecules for Huntington’s disease treatment. Sci. Rep. 2019, 9, 7260. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, R.; Gonzalez, P.; Lopez-Hurtado, A.; Dopazo, X.M.; Mellstrom, B.; Naranjo, J.R. Inhibition of the Neuronal Calcium Sensor DREAM Modulates Presenilin-2 Endoproteolysis. Front. Mol. Neurosci. 2018, 11, 449. [Google Scholar] [CrossRef] [Green Version]
- Avesani, A.; Marino, V.; Zanzoni, S.; Koch, K.W.; Dell’Orco, D. Molecular properties of human guanylate cyclase-activating protein 2 (GCAP2) and its retinal dystrophy-associated variant G157R. J. Biol. Chem. 2021, 296, 100619. [Google Scholar] [CrossRef]
- Behnen, P.; Dell’Orco, D.; Koch, K.W. Involvement of the calcium sensor GCAP1 in hereditary cone dystrophies. Biol. Chem. 2010, 391, 631–637. [Google Scholar] [CrossRef]
- Morales-Camara, S.; Alexandre-Moreno, S.; Bonet-Fernandez, J.M.; Atienzar-Aroca, R.; Aroca-Aguilar, J.D.; Ferre-Fernandez, J.J.; Mendez, C.D.; Morales, L.; Fernandez-Sanchez, L.; Cuenca, N.; et al. Role of GUCA1C in Primary Congenital Glaucoma and in the Retina: Functional Evaluation in Zebrafish. Genes 2020, 11, 550. [Google Scholar] [CrossRef] [PubMed]
- Ames, J.B.; Lim, S. Molecular structure and target recognition of neuronal calcium sensor proteins. Biochim. Biophys. Acta 2012, 1820, 1205–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Callaghan, D.W.; Burgoyne, R.D. Identification of residues that determine the absence of a Ca(2+)/myristoyl switch in neuronal calcium sensor-1. J. Biol. Chem. 2004, 279, 14347–14354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Callaghan, D.W.; Ivings, L.; Weiss, J.L.; Ashby, M.C.; Tepikin, A.V.; Burgoyne, R.D. Differential use of myristoyl groups on neuronal calcium sensor proteins as a determinant of spatio-temporal aspects of Ca2+ signal transduction. J. Biol. Chem. 2002, 277, 14227–14237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ames, J.B. Structural Insights into Retinal Guanylate Cyclase Activator Proteins (GCAPs). Int. J. Mol. Sci. 2021, 22, 8731. [Google Scholar] [CrossRef]
- Weiergraber, O.H.; Senin, I.I.; Zernii, E.Y.; Churumova, V.A.; Kovaleva, N.A.; Nazipova, A.A.; Permyakov, S.E.; Permyakov, E.A.; Philippov, P.P.; Granzin, J.; et al. Tuning of a neuronal calcium sensor. J. Biol. Chem. 2006, 281, 37594–37602. [Google Scholar] [CrossRef] [Green Version]
- Zernii, E.Y.; Komolov, K.E.; Permyakov, S.E.; Kolpakova, T.; Dell’orco, D.; Poetzsch, A.; Knyazeva, E.L.; Grigoriev, I.I.; Permyakov, E.A.; Senin, I.I.; et al. Involvement of the recoverin C-terminal segment in recognition of the target enzyme rhodopsin kinase. Biochem. J. 2011, 435, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Baksheeva, V.E.; Nazipova, A.A.; Zinchenko, D.V.; Serebryakova, M.V.; Senin, I.I.; Permyakov, S.E.; Philippov, P.P.; Li, Y.; Zamyatnin, A.A.; Zernii, E.Y.; et al. Ca2+ -myristoyl switch in neuronal calcium sensor-1: A role of C-terminal segment. CNS Neurol. Disord. Drug Targets 2015, 14, 437–451. [Google Scholar] [CrossRef] [Green Version]
- Zernii, E.Y.; Grigoriev, I.I.; Nazipova, A.A.; Scholten, A.; Kolpakova, T.V.; Zinchenko, D.V.; Kazakov, A.S.; Senin, I.I.; Permyakov, S.E.; Dell’Orco, D.; et al. Regulatory function of the C-terminal segment of guanylate cyclase-activating protein 2. Biochim. Biophys. Acta 2015, 1854, 1325–1337. [Google Scholar] [CrossRef]
- Zhou, W.; Qian, Y.; Kunjilwar, K.; Pfaffinger, P.J.; Choe, S. Structural insights into the functional interaction of KChIP1 with Shal-type K(+) channels. Neuron 2004, 41, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Dizhoor, A.M.; Olshevskaya, E.V.; Peshenko, I.V. Mg2+/Ca2+ cation binding cycle of guanylyl cyclase activating proteins (GCAPs): Role in regulation of photoreceptor guanylyl cyclase. Mol. Cell Biochem. 2010, 334, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Aravind, P.; Chandra, K.; Reddy, P.P.; Jeromin, A.; Chary, K.V.; Sharma, Y. Regulatory and structural EF-hand motifs of neuronal calcium sensor-1: Mg2+ modulates Ca2+ binding, Ca2+-induced conformational changes, and equilibrium unfolding transitions. J. Mol. Biol. 2008, 376, 1100–1115. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Pan, W.; Braunewell, K.H.; Ames, J.B. Structural analysis of Mg2+ and Ca2+ binding, myristoylation, and dimerization of the neuronal calcium sensor and visinin-like protein 1 (VILIP-1). J. Biol. Chem. 2011, 286, 6354–6366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baksheeva, V.E.; Nemashkalova, E.L.; Firsov, A.M.; Zalevsky, A.O.; Vladimirov, V.I.; Tikhomirova, N.K.; Philippov, P.P.; Zamyatnin, A.A., Jr.; Zinchenko, D.V.; Antonenko, Y.N.; et al. Membrane Binding of Neuronal Calcium Sensor-1: Highly Specific Interaction with Phosphatidylinositol-3-Phosphate. Biomolecules 2020, 10, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Permyakov, S.E.; Cherskaya, A.M.; Wasserman, L.A.; Khokhlova, T.I.; Senin, I.I.; Zargarov, A.A.; Zinchenko, D.V.; Zernii, E.Y.; Lipkin, V.M.; Philippov, P.P.; et al. Recoverin is a zinc-binding protein. J. Proteome Res. 2003, 2, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkov, P.O.; Roman, A.Y.; Baksheeva, V.E.; Nazipova, A.A.; Shevelyova, M.P.; Vladimirov, V.I.; Buyanova, M.F.; Zinchenko, D.V.; Zamyatnin, A.A., Jr.; Devred, F.; et al. Functional Status of Neuronal Calcium Sensor-1 Is Modulated by Zinc Binding. Front. Mol. Neurosci. 2018, 11, 459. [Google Scholar] [CrossRef] [Green Version]
- Baksheeva, V.E.; Baldin, A.V.; Zalevsky, A.O.; Nazipova, A.A.; Kazakov, A.S.; Vladimirov, V.I.; Gorokhovets, N.V.; Devred, F.; Philippov, P.P.; Bazhin, A.V.; et al. Disulfide Dimerization of Neuronal Calcium Sensor-1: Implications for Zinc and Redox Signaling. Int. J. Mol. Sci. 2021, 22, 12602. [Google Scholar] [CrossRef] [PubMed]
- Sensi, S.L.; Paoletti, P.; Bush, A.I.; Sekler, I. Zinc in the physiology and pathology of the CNS. Nat. Rev. Neurosci. 2009, 10, 780–791. [Google Scholar] [CrossRef]
- Watt, N.T.; Whitehouse, I.J.; Hooper, N.M. The role of zinc in Alzheimer’s disease. Int. J. Alzheimer’s Dis. 2010, 2011, 971021. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Andereggen, L.; Yuki, K.; Omura, K.; Yin, Y.; Gilbert, H.Y.; Erdogan, B.; Asdourian, M.S.; Shrock, C.; de Lima, S.; et al. Mobile zinc increases rapidly in the retina after optic nerve injury and regulates ganglion cell survival and optic nerve regeneration. Proc. Natl. Acad. Sci. USA 2017, 114, E209–E218. [Google Scholar] [CrossRef] [Green Version]
- Ugarte, M.; Osborne, N.N. Recent advances in the understanding of the role of zinc in ocular tissues. Metallomics 2014, 6, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.P.; Lee, N.M. Role of zinc in ALS. Amyotroph. Lateral Scler. 2007, 8, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Bush, A.I.; Pettingell, W.H.; Multhaup, G.; d Paradis, M.; Vonsattel, J.P.; Gusella, J.F.; Beyreuther, K.; Masters, C.L.; Tanzi, R.E. Rapid induction of Alzheimer A beta amyloid formation by zinc. Science 1994, 265, 1464–1467. [Google Scholar] [CrossRef] [PubMed]
- Garnier, C.; Devred, F.; Byrne, D.; Puppo, R.; Roman, A.Y.; Malesinski, S.; Golovin, A.V.; Lebrun, R.; Ninkina, N.N.; Tsvetkov, P.O. Zinc binding to RNA recognition motif of TDP-43 induces the formation of amyloid-like aggregates. Sci. Rep. 2017, 7, 6812. [Google Scholar] [CrossRef] [Green Version]
- Roman, A.Y.; Devred, F.; Byrne, D.; La Rocca, R.; Ninkina, N.N.; Peyrot, V.; Tsvetkov, P.O. Zinc Induces Temperature-Dependent Reversible Self-Assembly of Tau. J. Mol. Biol. 2019, 431, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.; Peto, T.; Lengyel, I.; Emri, E. Zinc Nutrition and Inflammation in the Aging Retina. Mol. Nutr. Food Res. 2019, 63, e1801049. [Google Scholar] [CrossRef]
- Vladimirov, V.I.; Baksheeva, V.E.; Mikhailova, I.V.; Ismailov, R.G.; Litus, E.A.; Tikhomirova, N.K.; Nazipova, A.A.; Permyakov, S.E.; Zernii, E.Y.; Zinchenko, D.V. A Novel Approach to Bacterial Expression and Purification of Myristoylated Forms of Neuronal Calcium Sensor Proteins. Biomolecules 2020, 10, 1025. [Google Scholar] [CrossRef]
- Senin, I.I.; Zargarov, A.A.; Alekseev, A.M.; Gorodovikova, E.N.; Lipkin, V.M.; Philippov, P.P. N-myristoylation of recoverin enhances its efficiency as an inhibitor of rhodopsin kinase. FEBS Lett. 1995, 376, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.Y.; Koch, K.W. Calcium- and myristoyl-dependent properties of guanylate cyclase-activating protein-1 and protein-2. Biochemistry 2002, 41, 13021–13028. [Google Scholar] [CrossRef]
- Baksheeva, V.E.; Roman, A.Y.; Villard, C.; Devred, F.; Byrne, D.; Yatoui, D.; Zalevsky, A.O.; Vologzhannikova, A.A.; Sokolov, A.S.; Permyakov, S.E.; et al. Mechanism of Zn(2+) and Ca(2+) Binding to Human S100A1. Biomolecules 2021, 11, 1823. [Google Scholar] [CrossRef]
- Huynh, K.; Partch, C.L. Analysis of protein stability and ligand interactions by thermal shift assay. Curr. Protoc. Protein Sci. 2015, 79, 28.9.1–28.9.14. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Ames, J.B. Dimerization of Neuronal Calcium Sensor Proteins. Front. Mol. Neurosci. 2018, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Dell’Orco, D.; Behnen, P.; Linse, S.; Koch, K.W. Calcium binding, structural stability and guanylate cyclase activation in GCAP1 variants associated with human cone dystrophy. Cell Mol. Life Sci. 2010, 67, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.S.; Azam, S.; Pham, K.; Leyva, D.; Fouque, K.J.D.; Fernandez-Lima, F.; Miksovska, J. Nanomolar affinity of EF-hands in neuronal calcium sensor 1 for bivalent cations Pb2+, Mn2+ and Hg2+. Metallomics 2022. [Google Scholar] [CrossRef] [PubMed]
- Zernii, E.Y.; Nazipova, A.A.; Nemashkalova, E.L.; Kazakov, A.S.; Gancharova, O.S.; Serebryakova, M.V.; Tikhomirova, N.K.; Baksheeva, V.E.; Vladimirov, V.I.; Zinchenko, D.V.; et al. Light-Induced Thiol Oxidation of Recoverin Affects Rhodopsin Desensitization. Front. Mol. Neurosci. 2018, 11, 474. [Google Scholar] [CrossRef]
- Maret, W. Metals on the move: Zinc ions in cellular regulation and in the coordination dynamics of zinc proteins. Biometals 2011, 24, 411–418. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Bush, A.I. Synaptically released zinc: Physiological functions and pathological effects. Biometals 2001, 14, 353–366. [Google Scholar] [CrossRef]
- Redenti, S.; Ripps, H.; Chappell, R.L. Zinc release at the synaptic terminals of rod photoreceptors. Exp. Eye Res. 2007, 85, 580–584. [Google Scholar] [CrossRef]
- Lee, J.Y.; Cole, T.B.; Palmiter, R.D.; Koh, J.Y. Accumulation of zinc in degenerating hippocampal neurons of ZnT3-null mice after seizures: Evidence against synaptic vesicle origin. J. Neurosci. 2000, 20, RC79. [Google Scholar] [CrossRef]
- Kiedrowski, L. Cytosolic acidification and intracellular zinc release in hippocampal neurons. J. Neurochem. 2012, 121, 438–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, A.; Hosseinibarkooie, S.; Schneider, S.; Kaczmarek, A.; Torres-Benito, L.; Mendoza-Ferreira, N.; Overhoff, M.; Rombo, R.; Grysko, V.; Kye, M.J.; et al. Neurocalcin Delta Knockout Impairs Adult Neurogenesis Whereas Half Reduction Is Not Pathological. Front. Mol. Neurosci. 2019, 12, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zernii, E.Y.; Nazipova, A.A.; Gancharova, O.S.; Kazakov, A.S.; Serebryakova, M.V.; Zinchenko, D.V.; Tikhomirova, N.K.; Senin, I.I.; Philippov, P.P.; Permyakov, E.A.; et al. Light-induced disulfide dimerization of recoverin under ex vivo and in vivo conditions. Free Radic. Biol. Med. 2015, 83, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Maret, W.; Vallee, B.L. Thiolate ligands in metallothionein confer redox activity on zinc clusters. Proc. Natl. Acad. Sci. USA 1998, 95, 3478–3482. [Google Scholar] [CrossRef] [Green Version]
- Aravindakumar, C.T.; Ceulemans, J.; De Ley, M. Nitric oxide induces Zn2+ release from metallothionein by destroying zinc-sulphur clusters without concomitant formation of S-nitrosothiol. Biochem. J. 1999, 344 Pt 1, 253–258. [Google Scholar] [CrossRef]
- Hayashi, M.; Miyata, R.; Tanuma, N. Oxidative stress in developmental brain disorders. Adv. Exp. Med. Biol. 2012, 724, 278–290. [Google Scholar] [CrossRef]
- Lim, S.; Scholten, A.; Manchala, G.; Cudia, D.; Zlomke-Sell, S.K.; Koch, K.W.; Ames, J.B. Structural Characterization of Ferrous Ion Binding to Retinal Guanylate Cyclase Activator Protein 5 from Zebrafish Photoreceptors. Biochemistry 2017, 56, 6652–6661. [Google Scholar] [CrossRef]
- He, F.; Seryshev, A.B.; Cowan, C.W.; Wensel, T.G. Multiple zinc binding sites in retinal rod cGMP phosphodiesterase, PDE6alpha beta. J. Biol. Chem. 2000, 275, 20572–20577. [Google Scholar] [CrossRef] [Green Version]
- Shuster, T.A.; Nagy, A.K.; Conly, D.C.; Farber, D.B. Direct zinc binding to purified rhodopsin and disc membranes. Biochem. J. 1992, 282 Pt 1, 123–128. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Ca2+ | ITC | TS (or nanoDSF) | DLS | ||
|---|---|---|---|---|---|---|
| N * | KD, μM | ΔH *, kcal/mol | Tm *, °C (−/+ Zn2+) | Dh *, nm (−/+ Zn2+) | ||
| recoverin | − | 1.8 | (1.0 ± 0.2) × 10−6 | −11.4 | 50.3/54.8 | 4.6/5.7 |
| + | 1.3 | (2.7 ± 0.3) × 10−6 | 3.0 | 78.4/69.3 | 4.6/4.7 | |
| VILIP1 | − | 1.3 0.6 | (4.3 ± 1.0) × 10−8, (1.7 ± 0.2) × 10−6 | −2.7 −1.8 | 68.6/63.7; 38.1 | 6.3/6.6 |
| + | no binding | n.d./38.2 | 6.7/6.9 | |||
| NCLD | − | 0.9 | (2.6 ± 0.5) × 10−5 | −2.4 | 61.1/38.8 | 5.4/5.8 |
| + | 2.6 | (2.0 ± 0.5) × 10−5 | −1.4 | 84.5/51.2 | 5.8/6.6 | |
| GCAP1 | − | 0.7 | (1.4 ± 0.2) × 10−5 | −9.3 | n.d. | 5.9/6.8 |
| + | 1.1 | (1.1 ± 0.1) × 10−5 | −6.1 | n.d. | 5.9/7.3 | |
| GCAP2 | − | n.d. | n.d. | n.d. | n.d. | 6.1/6.4 |
| + | n.d. | n.d. | n.d. | n.d. | 6.1/6.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baksheeva, V.E.; Tsvetkov, P.O.; Zalevsky, A.O.; Vladimirov, V.I.; Gorokhovets, N.V.; Zinchenko, D.V.; Permyakov, S.E.; Devred, F.; Zernii, E.Y. Zinc Modulation of Neuronal Calcium Sensor Proteins: Three Modes of Interaction with Different Structural Outcomes. Biomolecules 2022, 12, 956. https://doi.org/10.3390/biom12070956
Baksheeva VE, Tsvetkov PO, Zalevsky AO, Vladimirov VI, Gorokhovets NV, Zinchenko DV, Permyakov SE, Devred F, Zernii EY. Zinc Modulation of Neuronal Calcium Sensor Proteins: Three Modes of Interaction with Different Structural Outcomes. Biomolecules. 2022; 12(7):956. https://doi.org/10.3390/biom12070956
Chicago/Turabian StyleBaksheeva, Viktoriia E., Philipp O. Tsvetkov, Arthur O. Zalevsky, Vasiliy I. Vladimirov, Neonila V. Gorokhovets, Dmitry V. Zinchenko, Sergei E. Permyakov, François Devred, and Evgeni Yu. Zernii. 2022. "Zinc Modulation of Neuronal Calcium Sensor Proteins: Three Modes of Interaction with Different Structural Outcomes" Biomolecules 12, no. 7: 956. https://doi.org/10.3390/biom12070956