
Spt3 and Spt8 Are Involved in the Formation of a Silencing Boundary by Interacting with TATA-Binding Protein

Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Plasmids
2.3. RNA Extraction and Real-Time PCR Analysis
2.4. Microarray Analysis
2.5. Immunoblotting
2.6. Chromatin Immunoprecipitation (ChIP)
3. Results
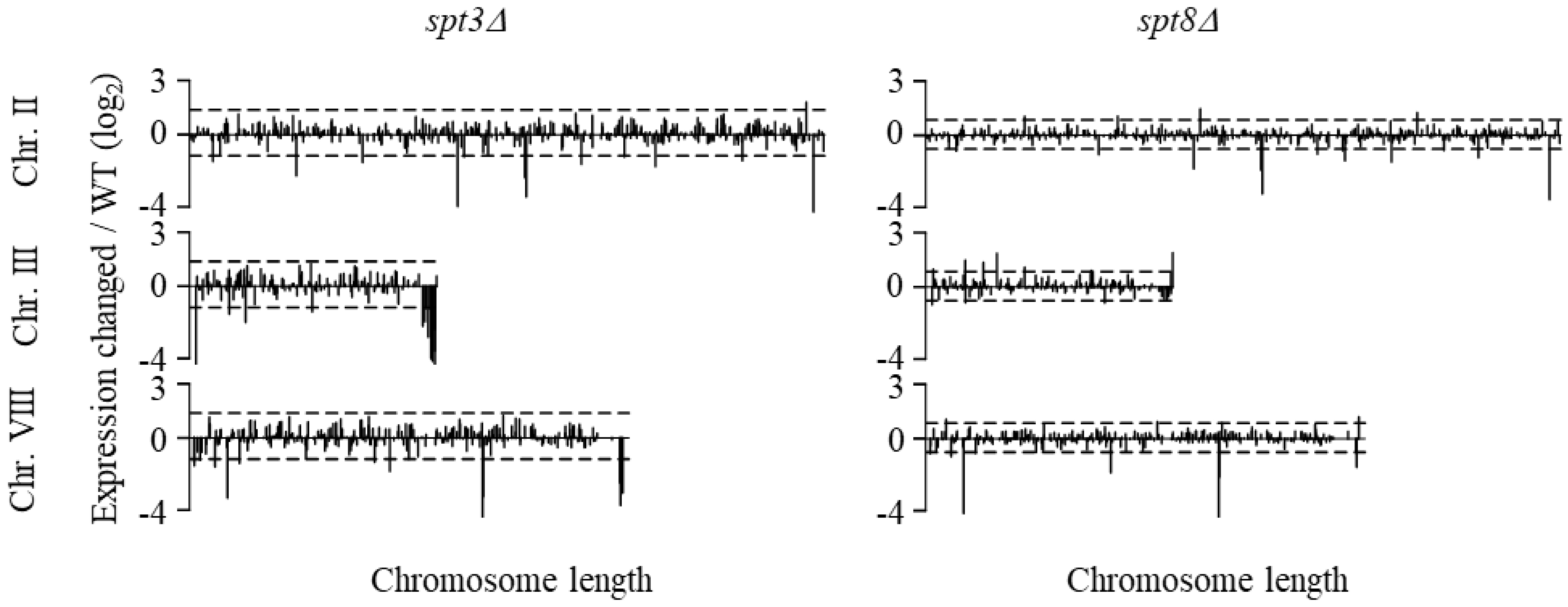
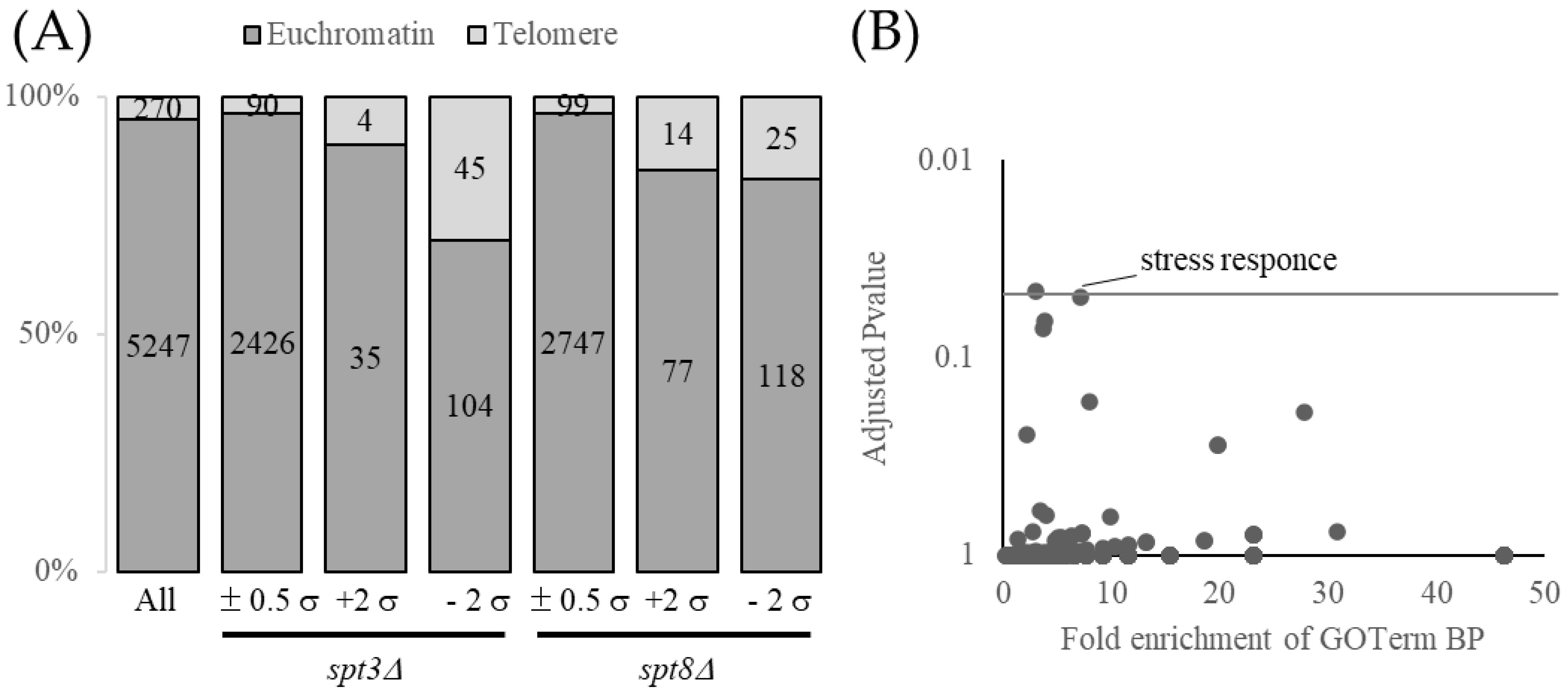
3.1. Spt3 and Spt8 Regulate Subtelomere Genes
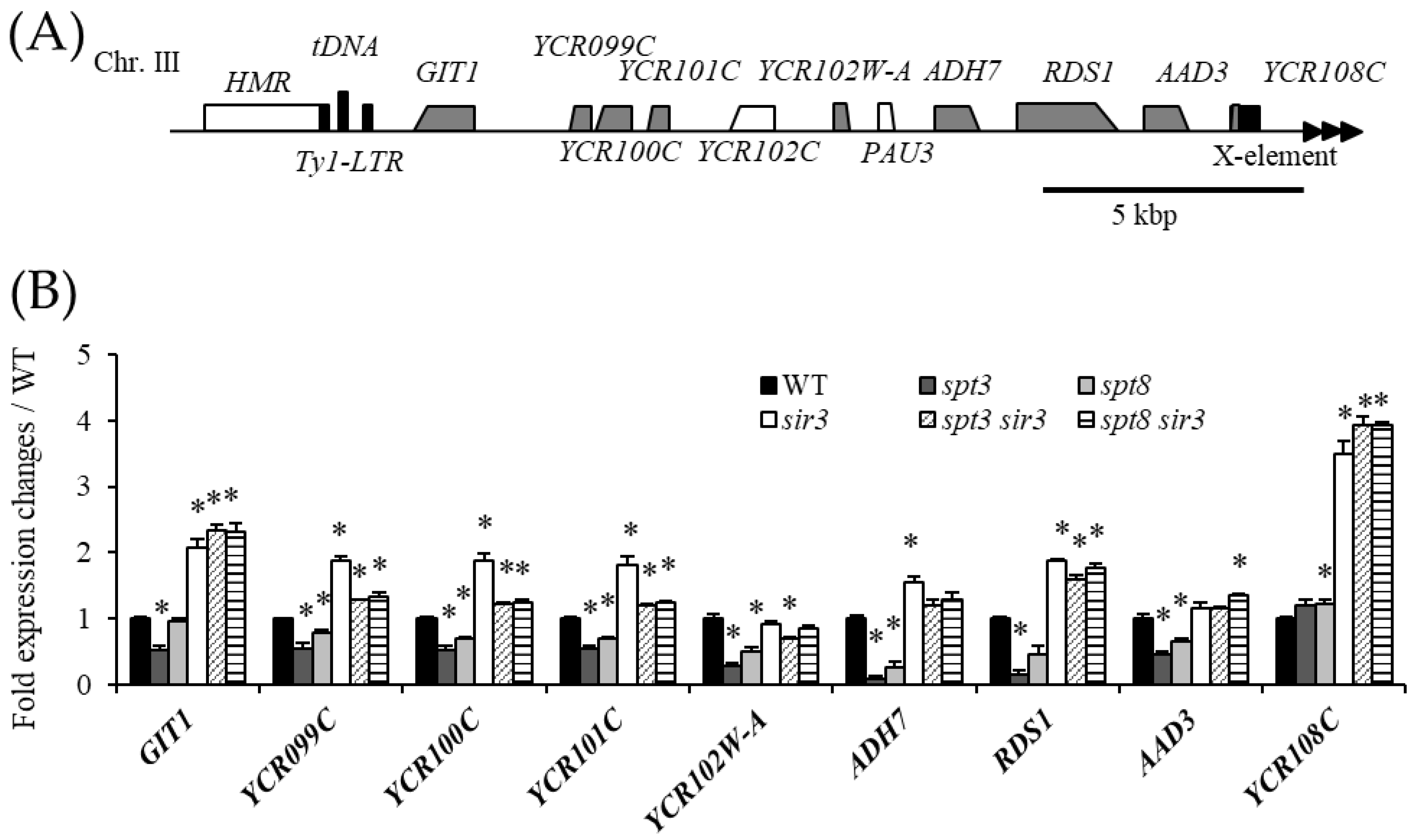
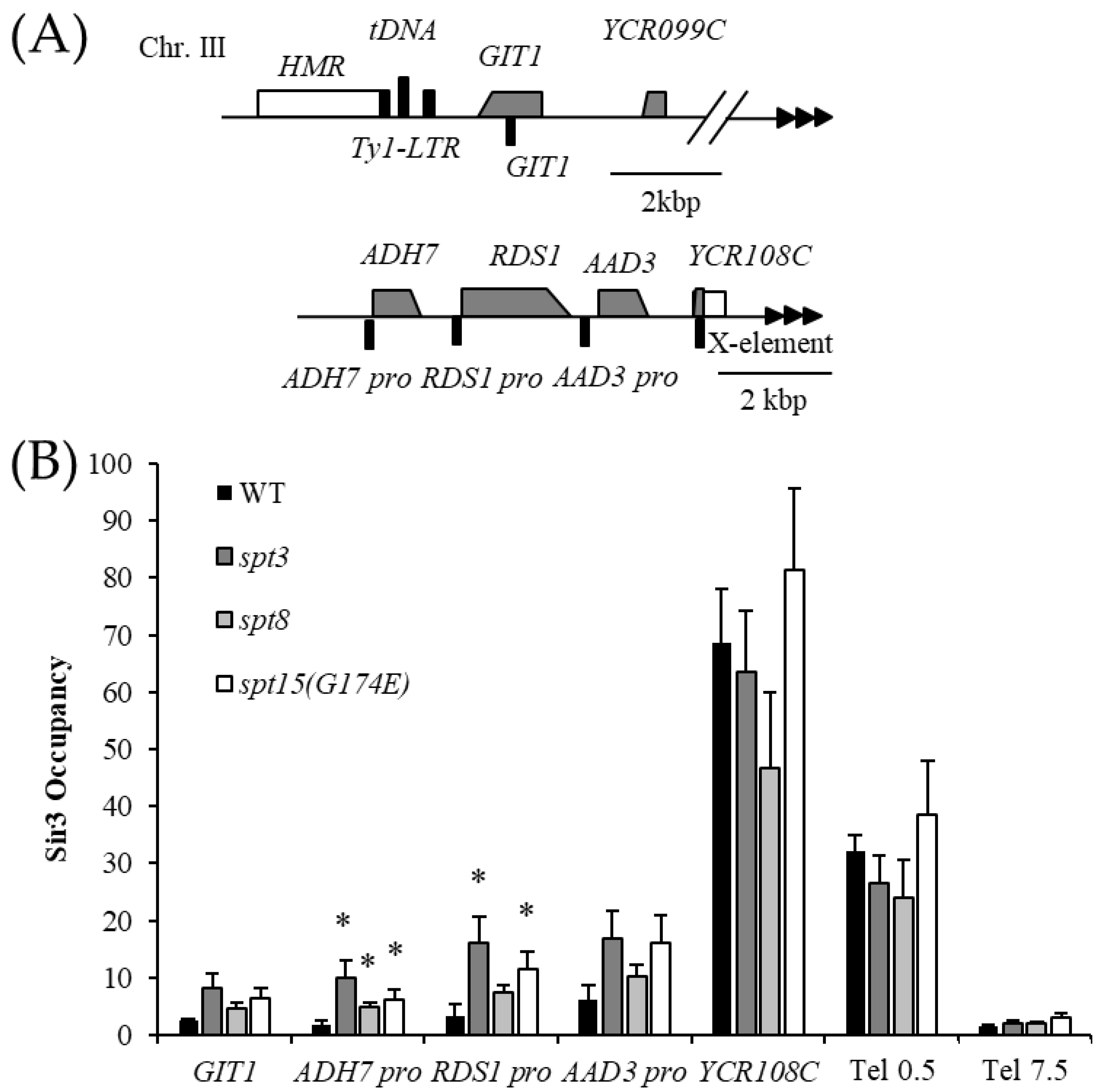
3.2. Spt3 and Spt8 Are Involved in the Subtelomere Silencing Boundary
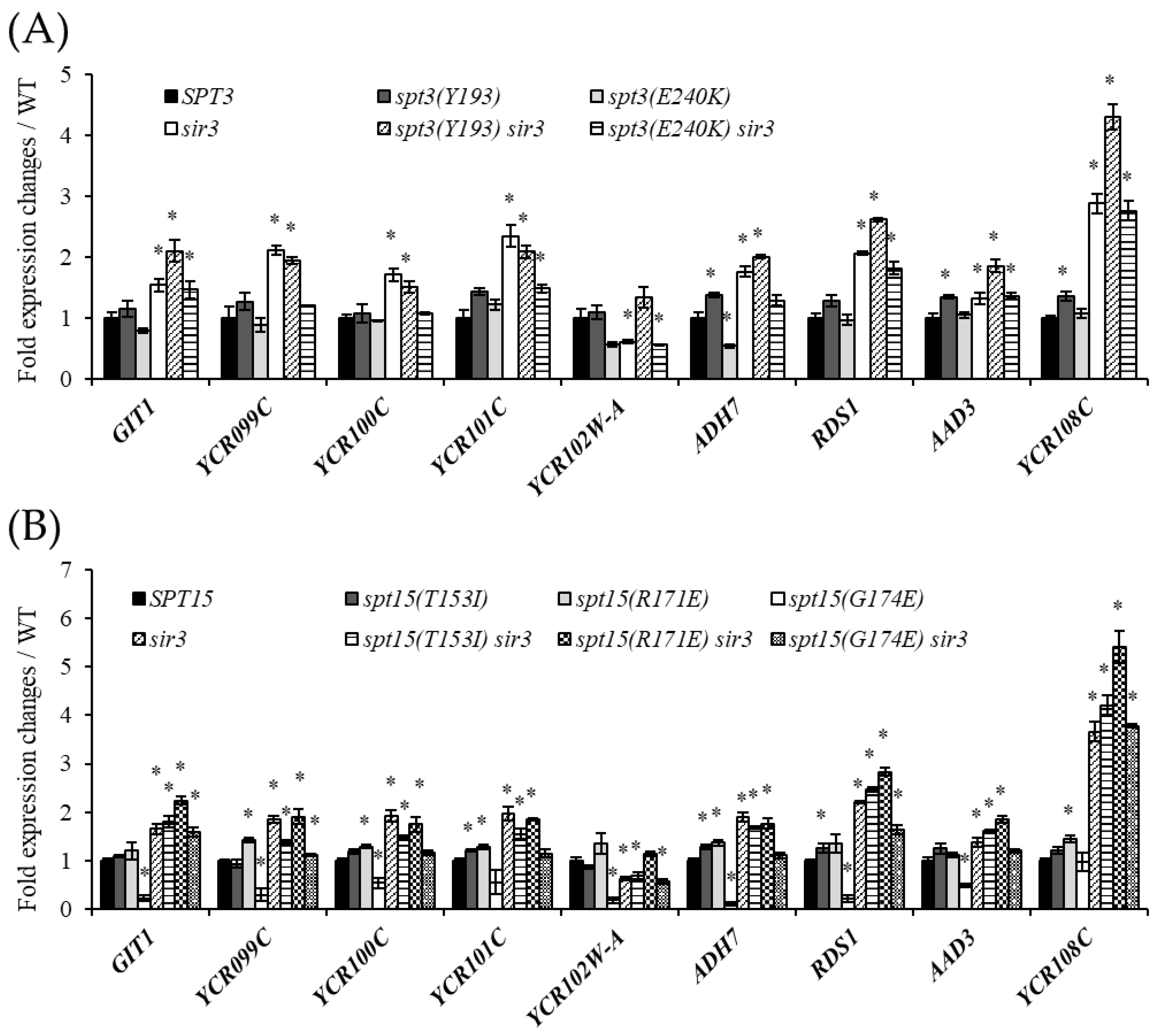
3.3. Interaction between Spt3 and TBPs Is Important for Telomere Silencing Boundary Formation
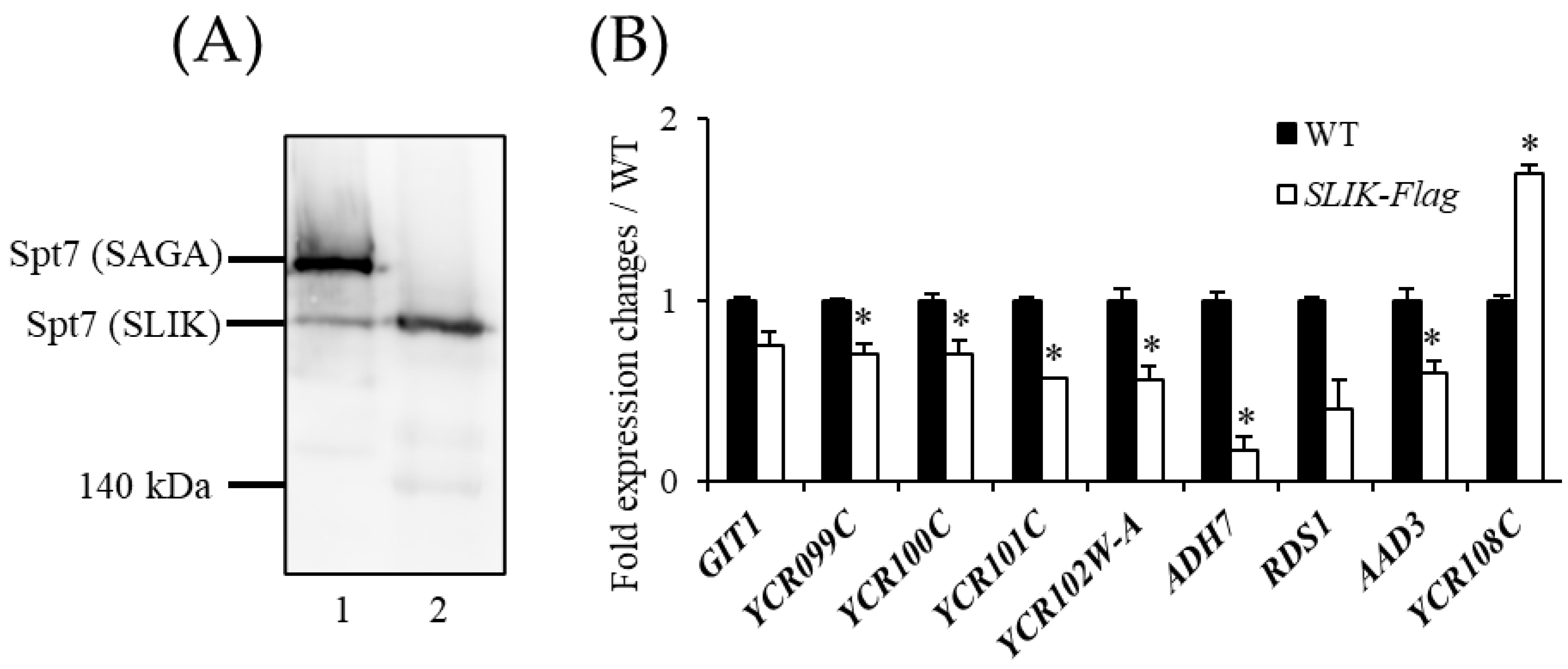
3.4. The SAGA Complex Is Required for Boundary Formation
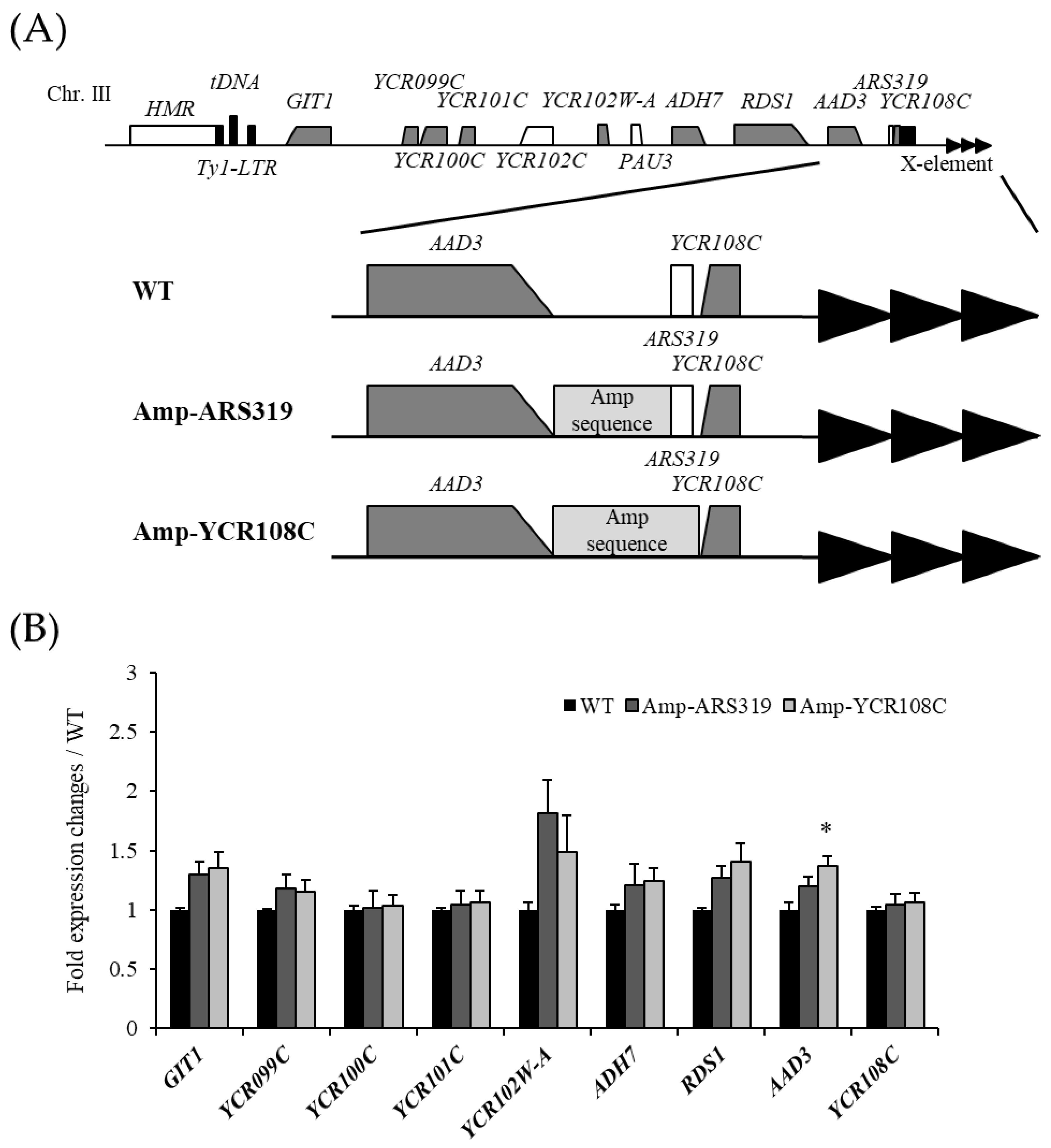
3.5. Spt3 and TBP Mutations Lead to the Spread of the Silencing Region
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imai, S.; Armstrong, C.M.; Kaeberlein, M.; Guarente, L. Transcriptional silencing and longevity protein Sir2 is an NAD-dependent histone deacetylase. Nature 2000, 403, 795–800. [Google Scholar] [CrossRef] [PubMed]
- Rusche, L.N.; Kirchmaier, A.L.; Rine, J. The establishment, inheritance, and function of silenced chromatin in Saccharomyces cerevisiae. Annu. Rev. Biochem. 2003, 72, 481–516. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.Q.; Hatanaka, A.; Oki, M. Boundaries of transcriptionally silent chromatin in Saccharomyces cerevisiae. Genes Genet. Syst. 2011, 86, 73–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartenberg, M.R.; Smith, J.S. The Nuts and Bolts of Transcriptionally Silent Chromatin in Saccharomyces cerevisiae. Genetics 2016, 203, 1563–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellahi, A.; Thurtle, D.M.; Rine, J. The Chromatin and Transcriptional Landscape of Native Saccharomyces cerevisiae Telomeres and Subtelomeric Domains. Genetics 2015, 200, 505–521. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.; Yeom, S.; Park, J.; Lee, J.S. The regional sequestration of heterochromatin structural proteins is critical to form and maintain silent chromatin. Epigenetics Chromatin 2022, 15, 5. [Google Scholar] [CrossRef]
- Yeom, S.; Oh, J.; Lee, J.S. Spreading-dependent or independent Sir2-mediated gene silencing in budding yeast. Genes Genom. 2022, 44, 359–367. [Google Scholar] [CrossRef]
- Kimura, A.; Umehara, T.; Horikoshi, M. Chromosomal gradient of histone acetylation established by Sas2p and Sir2p functions as a shield against gene silencing. Nat. Genet. 2002, 32, 370–377. [Google Scholar] [CrossRef]
- Suka, N.; Luo, K.; Grunstein, M. Sir2p and Sas2p opposingly regulate acetylation of yeast histone H4 lysine16 and spreading of heterochromatin. Nat. Genet. 2002, 32, 378–383. [Google Scholar] [CrossRef]
- Donze, D.; Adams, C.R.; Rine, J.; Kamakaka, R.T. The boundaries of the silenced HMR domain in Saccharomyces cerevisiae. Genes Dev. 1999, 13, 698–708. [Google Scholar] [CrossRef]
- Donze, D.; Kamakaka, R.T. RNA polymerase III and RNA polymerase II promoter complexes are heterochromatin barriers in Saccharomyces cerevisiae. EMBO J. 2001, 20, 520–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oki, M.; Kamakaka, R.T. Blockers and barriers to transcription: Competing activities? Curr. Opin. Cell Biol. 2002, 14, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Oki, M.; Kamakaka, R.T. Barrier function at HMR. Mol. Cell 2005, 19, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Oki, M.; Valenzuela, L.; Chiba, T.; Ito, T.; Kamakaka, R.T. Barrier proteins remodel and modify chromatin to restrict silenced domains. Mol. Cell. Biol. 2004, 24, 1956–1967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wery, M.; Shematorova, E.; Van Driessche, B.; Vandenhaute, J.; Thuriaux, P.; Van Mullem, V. Members of the SAGA and Mediator complexes are partners of the transcription elongation factor TFIIS. EMBO J. 2004, 23, 4232–4242. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Luo, J.; Ranish, J.; Hahn, S. Architecture of the Saccharomyces cerevisiae SAGA transcription coactivator complex. EMBO J. 2014, 33, 2534–2546. [Google Scholar] [CrossRef] [Green Version]
- Setiaputra, D.; Ross, J.D.; Lu, S.; Cheng, D.T.; Dong, M.Q.; Yip, C.K. Conformational flexibility and subunit arrangement of the modular yeast Spt-Ada-Gcn5 acetyltransferase complex. J. Biol. Chem. 2015, 290, 10057–10070. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Zheng, X.; Guan, H.; Cao, Y.; Qu, H.; Kang, J.; Ren, X.; Lei, J.; Dong, M.Q.; Li, X.; et al. Architecture of Saccharomyces cerevisiae SAGA complex. Cell Discov. 2019, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Papai, G.; Frechard, A.; Kolesnikova, O.; Crucifix, C.; Schultz, P.; Ben-Shem, A. Structure of SAGA and mechanism of TBP deposition on gene promoters. Nature 2020, 577, 711–716. [Google Scholar] [CrossRef]
- Wang, H.; Dienemann, C.; Stutzer, A.; Urlaub, H.; Cheung, A.C.M.; Cramer, P. Structure of the transcription coactivator SAGA. Nature 2020, 577, 717–720. [Google Scholar] [CrossRef]
- Ben-Shem, A.; Papai, G.; Schultz, P. Architecture of the multi-functional SAGA complex and the molecular mechanism of holding TBP. FEBS J. 2021, 288, 3135–3147. [Google Scholar] [CrossRef] [PubMed]
- Grant, P.A.; Winston, F.; Berger, S.L. The biochemical and genetic discovery of the SAGA complex. Biochim. Biophys. Acta Gene Regul. Mech. 2021, 1864, 194669. [Google Scholar] [CrossRef] [PubMed]
- Vasyliuk, D.; Felt, J.; Zhong, E.D.; Berger, B.; Davis, J.H.; Yip, C.K. Conformational landscape of the yeast SAGA complex as revealed by cryo-EM. Sci. Rep. 2022, 12, 12306. [Google Scholar] [CrossRef] [PubMed]
- Kamata, K.; Hatanaka, A.; Goswami, G.; Shinmyozu, K.; Nakayama, J.; Urano, T.; Hatashita, M.; Uchida, H.; Oki, M. C-terminus of the Sgf73 subunit of SAGA and SLIK is important for retention in the larger complex and for heterochromatin boundary function. Genes Cells 2013, 18, 823–837. [Google Scholar] [CrossRef]
- Kamata, K.; Goswami, G.; Kashio, S.; Urano, T.; Nakagawa, R.; Uchida, H.; Oki, M. The N-terminus and Tudor domains of Sgf29 are important for its heterochromatin boundary formation function. J Biochem. 2014, 155, 159–171. [Google Scholar] [CrossRef] [PubMed]
- Kamata, K.; Shinmyozu, K.; Nakayama, J.I.; Hatashita, M.; Uchida, H.; Oki, M. Four domains of Ada1 form a heterochromatin boundary through different mechanisms. Genes Cells 2016, 21, 1125–1136. [Google Scholar] [CrossRef]
- Birck, C.; Poch, O.; Romier, C.; Ruff, M.; Mengus, G.; Lavigne, A.C.; Davidson, I.; Moras, D. Human TAF(II)28 and TAF(II)18 interact through a histone fold encoded by atypical evolutionary conserved motifs also found in the SPT3 family. Cell 1998, 94, 239–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenmann, D.M.; Arndt, K.M.; Ricupero, S.L.; Rooney, J.W.; Winston, F. SPT3 interacts with TFIID to allow normal transcription in Saccharomyces cerevisiae. Genes Dev. 1992, 6, 1319–1331. [Google Scholar] [CrossRef] [Green Version]
- Dudley, A.M.; Rougeulle, C.; Winston, F. The Spt components of SAGA facilitate TBP binding to a promoter at a post-activator-binding step in vivo. Genes Dev. 1999, 13, 2940–2945. [Google Scholar] [CrossRef]
- Belotserkovskaya, R.; Sterner, D.E.; Deng, M.; Sayre, M.H.; Lieberman, P.M.; Berger, S.L. Inhibition of TATA-binding protein function by SAGA subunits Spt3 and Spt8 at Gcn4-activated promoters. Mol. Cell. Biol. 2000, 20, 634–647. [Google Scholar] [CrossRef] [Green Version]
- Bhaumik, S.R.; Green, M.R. Differential requirement of SAGA components for recruitment of TATA-box-binding protein to promoters in vivo. Mol. Cell. Biol. 2002, 22, 7365–7371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huisinga, K.L.; Pugh, B.F. A genome-wide housekeeping role for TFIID and a highly regulated stress-related role for SAGA in Saccharomyces cerevisiae. Mol. Cell 2004, 13, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, X.; Kuo, M.H. Snf1p regulates Gcn5p transcriptional activity by antagonizing Spt3p. Genetics 2010, 184, 91–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madison, J.M.; Winston, F. Evidence that Spt3 functionally interacts with Mot1, TFIIA, and TATA-binding protein to confer promoter-specific transcriptional control in Saccharomyces cerevisiae. Mol. Cell. Biol. 1997, 17, 287–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampsey, M. Molecular genetics of the RNA polymerase II general transcriptional machinery. Microbiol. Mol. Biol. Rev. 1998, 62, 465–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhorn, J.N.; Winston, F. SPT3 is required for normal levels of a-factor and alpha-factor expression in Saccharomyces cerevisiae. Mol. Cell. Biol. 1988, 8, 822–827. [Google Scholar] [CrossRef] [PubMed]
- Pray-Grant, M.G.; Schieltz, D.; McMahon, S.J.; Wood, J.M.; Kennedy, E.L.; Cook, R.G.; Workman, J.L.; Yates, J.R., 3rd; Grant, P.A. The novel SLIK histone acetyltransferase complex functions in the yeast retrograde response pathway. Mol. Cell. Biol. 2002, 22, 8774–8786. [Google Scholar] [CrossRef] [Green Version]
- Sterner, D.E.; Belotserkovskaya, R.; Berger, S.L. SALSA, a variant of yeast SAGA, contains truncated Spt7, which correlates with activated transcription. Proc. Natl. Acad. Sci. USA 2002, 99, 11622–11627. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.K.; Workman, J.L. Histone acetyltransferase complexes: One size doesn’t fit all. Nat. Rev. Mol. Cell Biol. 2007, 8, 284–295. [Google Scholar] [CrossRef]
- Mischerikow, N.; Spedale, G.; Altelaar, A.F.; Timmers, H.T.; Pijnappel, W.W.; Heck, A.J. In-depth profiling of post-translational modifications on the related transcription factor complexes TFIID and SAGA. J. Proteome Res. 2009, 8, 5020–5030. [Google Scholar] [CrossRef]
- Rothstein, R. Targeting, disruption, replacement, and allele rescue: Integrative DNA transformation in yeast. Methods Enzymol. 1991, 194, 281–301. [Google Scholar] [CrossRef] [PubMed]
- Longtine, M.S.; Fares, H.; Pringle, J.R. Role of the yeast Gin4p protein kinase in septin assembly and the relationship between septin assembly and septin function. J. Cell Biol. 1998, 143, 719–736. [Google Scholar] [CrossRef]
- Boeke, J.D.; LaCroute, F.; Fink, G.R. A positive selection for mutants lacking orotidine-5′-phosphate decarboxylase activity in yeast: 5-fluoro-orotic acid resistance. Mol. Gen. Genet. 1984, 197, 345–346. [Google Scholar] [CrossRef] [PubMed]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Mitsumori, R.; Ohashi, T.; Kugou, K.; Ichino, A.; Taniguchi, K.; Ohta, K.; Uchida, H.; Oki, M. Analysis of novel Sir3 binding regions in Saccharomyces cerevisiae. J. Biochem. 2016, 160, 11–17. [Google Scholar] [CrossRef]
- Mitsumori, R.; Shinmyozu, K.; Nakayama, J.I.; Uchida, H.; Oki, M. Gic1 is a novel heterochromatin boundary protein in vivo. Genes Genet. Syst. 2016, 91, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, S.; Pugh, B.F. Sequential recruitment of SAGA and TFIID in a genomic response to DNA damage in Saccharomyces cerevisiae. Mol. Cell. Biol. 2011, 31, 190–202. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.J.; Miller, L.R.; Kreisberg, R.; Vazquez, L.; Wan, Y.; Aitchison, J.D. Environment-responsive transcription factors bind subtelomeric elements and regulate gene silencing. Mol. Syst. Biol. 2011, 7, 455. [Google Scholar] [CrossRef]
- Laprade, L.; Rose, D.; Winston, F. Characterization of new Spt3 and TATA-binding protein mutants of Saccharomyces cerevisiae: Spt3 TBP allele-specific interactions and bypass of Spt8. Genetics 2007, 177, 2007–2017. [Google Scholar] [CrossRef] [Green Version]
- Stargell, L.A.; Struhl, K. A new class of activation-defective TATA-binding protein mutants: Evidence for two steps of transcriptional activation in vivo. Mol. Cell. Biol. 1996, 16, 4456–4464. [Google Scholar] [CrossRef] [Green Version]
- Larschan, E.; Winston, F. The, S. cerevisiae SAGA complex functions in vivo as a coactivator for transcriptional activation by Gal4. Genes Dev. 2001, 15, 1946–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sermwittayawong, D.; Tan, S. SAGA binds TBP via its Spt8 subunit in competition with DNA: Implications for TBP recruitment. EMBO J. 2006, 25, 3791–3800. [Google Scholar] [CrossRef] [PubMed]
- Basehoar, A.D.; Zanton, S.J.; Pugh, B.F. Identification and distinct regulation of yeast TATA box-containing genes. Cell 2004, 116, 699–709. [Google Scholar] [CrossRef] [Green Version]
- de Jonge, W.J.; O’Duibhir, E.; Lijnzaad, P.; van Leenen, D.; Groot Koerkamp, M.J.; Kemmeren, P.; Holstege, F.C. Molecular mechanisms that distinguish TFIID housekeeping from regulatable SAGA promoters. EMBO J. 2017, 36, 274–290. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain No. | Genotype | Source |
|---|---|---|
| FUY 837 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 | This study |
| FUY 1621 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt3Δ::KanMX | This study |
| FUY 1622 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt8Δ::KanMX | This study |
| FUY 1624 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 sir3Δ::HphMX | This study |
| FUY 1657 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt3Δ::KanMX sir3Δ::HphMX | This study |
| FUY 1658 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt8Δ::KanMX sir3Δ::HphMX | This study |
| FUY 1662 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 SPT3::3xFlag-SPT3 | This study |
| FUY 1663 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 SPT7::3xFlag-SPT7 | This study |
| FUY 1664 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 SPT8::3xFlag-SPT8 | This study |
| FUY 1665 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 TBP(SPT15)::3xFlag-TBP(SPT15) | This study |
| FUY 1757 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt3Δ::Flag-spt3(Y193C) | This study |
| FUY 1758 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt3Δ::Flag-spt3(E240K) | This study |
| FUY 1759 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt15Δ::Flag-spt15(T153I) | This study |
| FUY 1760 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt15Δ::Flag-spt15(R171E) | This study |
| FUY 1761 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt15Δ::Flag-spt15(G174E) | This study |
| FUY 1762 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt3Δ::Flag-spt3(Y193C) sir3Δ::HphMX | This study |
| FUY 1763 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt3Δ::Flag-spt3(E240K) sir3Δ::HphMX | This study |
| FUY 1764 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt15Δ::Flag-spt15(T153I) sir3Δ::HphMX | This study |
| FUY 1765 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt15Δ::Flag-spt15(R171E) sir3Δ::HphMX | This study |
| FUY 1766 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt15Δ::Flag-spt15(G174E) sir3Δ::HphMX | This study |
| FUY 1840 | MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 spt7Δ::SPT7(1-1141)-3xFlag | This study |
| Primer No. | Sequence |
|---|---|
| 832-GIT1/YCR098C-F | CCAAAAGAGGTGGTATCCTGGTT |
| 833-GIT1/YCR098C-Rv | TGGACCACCGAAGGCTAGTG |
| 834-YCR099C-F | AATGCAAAAAGCCCATGGAA |
| 835-YCR099C-Rv | CTCTCCCTCAGGATTTTTTCACA |
| 837-YCR100C-C | GGGCCACCCTCCATGTTAG |
| 838-YCR101C-F | TGGGAAACGGTCAAAGAAATTG |
| 839-YCR101C-Rv | CCATGGAAAGGATCAACAGTAAATC |
| 842-YCR102W-A-F | GAGGAAAAGTTTGGAAGAACAAAAA |
| 843-YCR102W-A-Rv | CTCCCCGTAAAGAATGCTTGAT |
| 852-AAD3/YCR107W-F | GCGCCTCCGAACAAACAG |
| 853-AAD3/YCR107W-Rv | AGCAATCTTGGCCAATGCTT |
| 854-YCR108C-F | CCATGGCCCATTCTCACTAAA |
| 855-YCR108C-Rv | CAAGTGCCGTGCATAATGATG |
| 1503-836-YCR100C-F | CGATCGGAAGGACCGAAAA |
| 1707-851-RDS1/YCR106W-2Rv | GGACATAGCGGTATTGGCTTTT |
| 1708-oki264 GIT1-F | TGGATGTGCGTACGACCAAT |
| 1709-oki265 GIT1-Rv | ACCTGGTCCAGCATTACCTAACA |
| 1719-847-ADH7/YCR105W-Rv | TCTCCGCTTTCCATCCTTGT |
| 2882-846-ADH7/YCR105W-F | AAACTTCCGATCAGCGAAGAAG |
| 2883-850-RDS1/YCR106W-2F | GCCAGATGGAGGATGCAGTT |
| Primer No. | Sequence |
|---|---|
| 854-YCR108C-F | CCATGGCCCATTCTCACTAAA |
| 855-YCR108C-Rv | CAAGTGCCGTGCATAATGATG |
| 1708-oki264 GIT1-F | TGGATGTGCGTACGACCAAT |
| 1709-oki265 GIT1-Rv | ACCTGGTCCAGCATTACCTAACA |
| 3055-AAD3(-1)-R | TTTAAGCACGATGGATATGCTTC |
| 3056-AAD3(-82)-F | CTTTTGCTGGTTTCGATGATG |
| 3061-RDS1(-1)-R | CAGAGCATTTCAGCAGCCAA |
| 3062-RDS1(-93)-F | GACATCATTACTAATAATGTTACTC |
| 4827-ADH7 pro 1 F | CGGCCGCATAATAAAATGGA |
| 4828-ADH7 pro 1 Rv | TCTACTCAGAGTTTTGGTGCTCAATT |
| 4850-tel(VI)0.5(F) | CCTTTTTTGATATAACTGTCGGAGAGT |
| 4851-tel(VI)0.5(Rv) | TCCGAACGCTATTCCAGAAAGT |
| 4852-tel(VI)7.5(F) | TGTAGACTTCCCACTGTATTTGAATGA |
| 4853-tel(VI)7.5(Rv) | CGTGAAAGTTCAGCGCAACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamata, K.; Ayano, T.; Oki, M. Spt3 and Spt8 Are Involved in the Formation of a Silencing Boundary by Interacting with TATA-Binding Protein. Biomolecules 2023, 13, 619. https://doi.org/10.3390/biom13040619
Kamata K, Ayano T, Oki M. Spt3 and Spt8 Are Involved in the Formation of a Silencing Boundary by Interacting with TATA-Binding Protein. Biomolecules. 2023; 13(4):619. https://doi.org/10.3390/biom13040619
Chicago/Turabian StyleKamata, Kazuma, Takahito Ayano, and Masaya Oki. 2023. "Spt3 and Spt8 Are Involved in the Formation of a Silencing Boundary by Interacting with TATA-Binding Protein" Biomolecules 13, no. 4: 619. https://doi.org/10.3390/biom13040619