
Exploring Genetic Interactions with Telomere Protection Gene pot1 in Fission Yeast

1
Graduate School of Integrated Sciences for Life, Hiroshima University, Higashi-Hiroshima 739-8530, Japan
2
Hiroshima Research Center for Healthy Aging (HiHA), Hiroshima University, Higashi-Hiroshima 739-8530, Japan
Biomolecules 2023, 13(2), 370; https://doi.org/10.3390/biom13020370
Submission received: 22 January 2023
/
Revised: 13 February 2023
/
Accepted: 14 February 2023
/
Published: 15 February 2023
(This article belongs to the Special Issue Yeast Models for Gene Regulation)
Abstract
:The regulation of telomere length has a significant impact on cancer risk and aging in humans. Circular chromosomes are found in humans and are often unstable during mitosis, resulting in genome instability. Some types of cancer have a high frequency of a circular chromosome. Fission yeast is a good model for studying the formation and stability of circular chromosomes as deletion of pot1 (encoding a telomere protection protein) results in rapid telomere degradation and chromosome fusion. Pot1 binds to single-stranded telomere DNA and is conserved from fission yeast to humans. Loss of pot1 leads to viable strains in which all three fission yeast chromosomes become circular. In this review, I will introduce pot1 genetic interactions as these inform on processes such as the degradation of uncapped telomeres, chromosome fusion, and maintenance of circular chromosomes. Therefore, exploring genes that genetically interact with pot1 contributes to finding new genes and/or new functions of genes related to the maintenance of telomeres and/or circular chromosomes.
1. Introduction
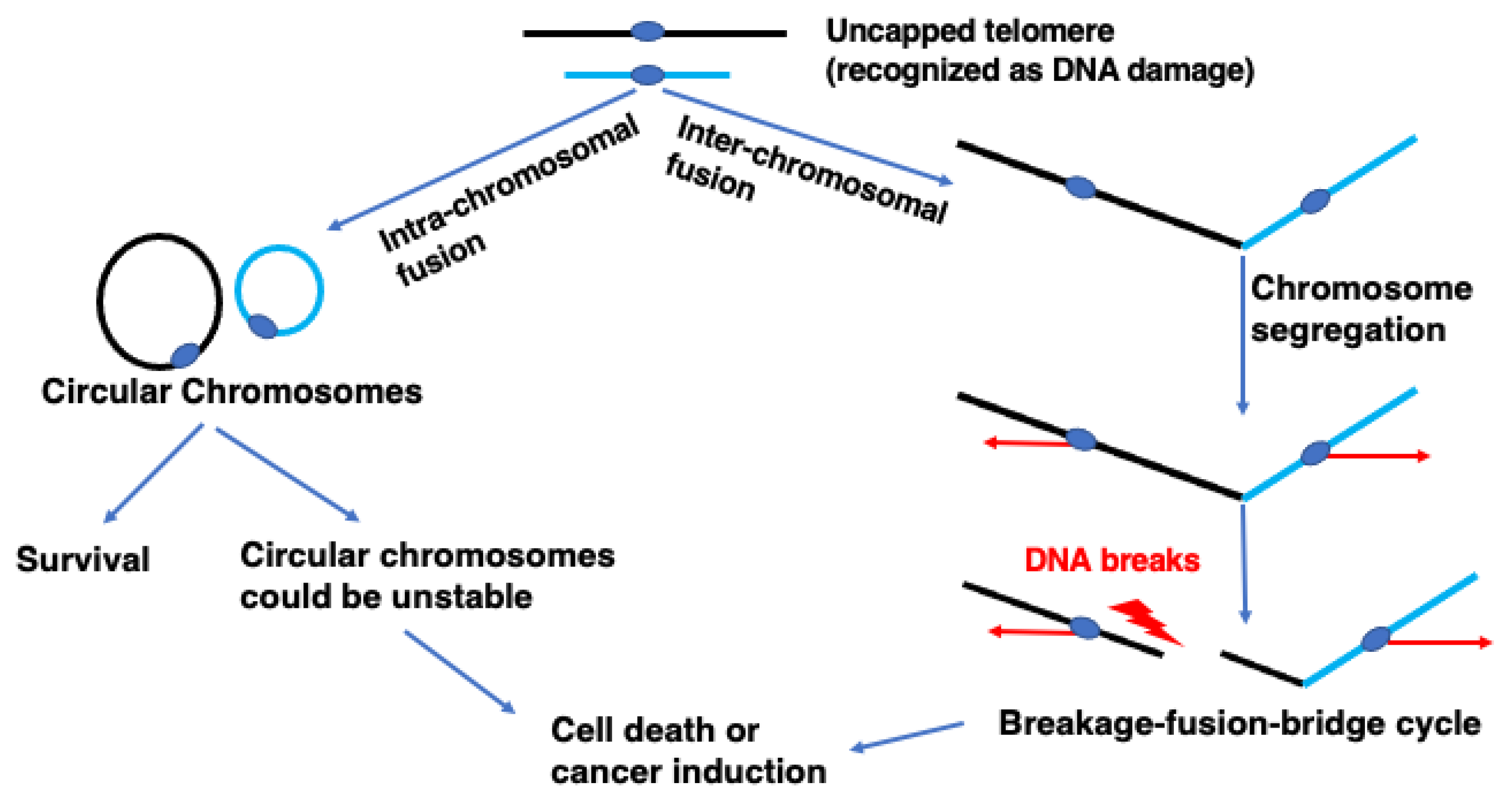
Telomeres cap the end of the chromosome to maintain genome stability [1,2]. They consist of a protein complex called Shelterin, which binds the end of the chromosome [3]. Telomere DNA consists of tandem nucleic acid repeats (TTAGGG) in vertebrates. Repeats of telomere DNA shorten with each cell division in most normal cells, except stem and germ cells. Telomere DNA can be elongated by telomerase, a ribonucleoprotein reverse transcriptase that is active in most cancer cells but inactive in most normal cells. When telomere DNA becomes short and uncapped in normal cells, cells stop dividing through DNA damage checkpoint activation [4]. However, if cells fail to stop cell division due to defects in checkpoint activation, telomere DNA becomes critically short and becomes a substrate for homologous recombination (HR) and/or non-homologous end joining (NHEJ) [5]. HR activity in uncapped telomeres can maintain telomeres without telomerase activity in a process called alternative lengthening of telomeres (ALT), found in ~15% of cancer cells [6]. NHEJ activity in uncapped telomeres results in telomere fusion between the ends of inter-chromosomes or between those of intra-chromosomes (Figure 1). Telomere fusion between inter-chromosome ends results in a dicentric chromosome that forms an anaphase bridge in mitosis. Anaphase bridge breakage results in a DNA double-strand break, which can lead to ectopic fusion and result in another dicentric chromosome. These breakage–fusion–bridge events result in chromosomal instability, which promotes cancer development and telomerase activation [4].
Telomere fusion between intra-chromosome ends results in a circular chromosome. A circular chromosome, also called a ring chromosome, can be found in human cells, although their frequency is very low (approximately 1:50,000) [7]. Many human carriers of a ring chromosome have ring chromosome syndrome, which is characterized by short stature and developmental delay [8]. A ring chromosome induces many secondary chromosome rearrangements, resulting in an increased risk of cancer due to a loss of cancer suppressor genes or other genetic changes. Stabilizing the ring chromosome could reduce the risk of cancer in carriers. A ring chromosome can also be found at a high frequency in some types of cancer cell. In some mesenchymal origin tumors, more than 70% of cells could have a ring chromosome [9]. Therefore, the ring chromosome could be a potential target for cancer therapy. However, the nature of the ring chromosome in humans has not been well studied because of the lack of a model system with which to do so.
2. Telomere Protection by Pot1 in Human and in Fission Yeast Schizosaccharomyces pombe, and by Cdc13 in Budding Yeast Saccharomyces cerevisiae
Human POT1, a component of Shelterin complex, binds to single-stranded telomere DNA [10,11,12]. POT1 forms a subcomplex with TPP1, which recruits telomerase to telomere [13,14]. POT1 knock down in normal human diploid fibroblasts (IMR-90) induces apoptosis, senescence, or telomere fusion, showing that POT1 inhibits the DNA damage response at telomeres [15]. As POT1 blocks the access of telomerase to telomere DNA by the covering of the G-strand overhang, human POT1 acts a negative regulator of telomerase [14,16,17]. Indeed, mutation in POT1 results in telomere elongation [18,19,20].
Fission yeast is an excellent model organism for studying telomere maintenance and chromosomal instability induced by telomere dysfunction. The Shelterin complex is similar to that of mammalian cells. For example, the POT1 ortholog, also called Pot1 in Schizosaccharomyces pombe, binds to single-stranded telomere DNA and prevents the DNA damage response [10,21]. S. pombe Tpz1, the TPP1 ortholog, forms a complex with Pot1, and this complex recruits telomerase through Ccq1 [22]. As expression of N-terminal truncated Pot1 results in telomere elongation, S. pombe Pot1 is also thought to function as a negative regulator of telomerase [23]. trt1 encodes a catalytic subunit of telomerase in S. pombe [24]. Telomerase is always active in S. pombe, similar to human cancer cells that have telomerase activity. The deletion of trt1 results in gradual telomere shortening during several cell divisions [24]. Survivors of the trt1 knockout have either three circular chromosomes or three linear chromosomes, in which telomeres are maintained by HR, similar to ALT cells in human cancer [25].
Circularization of one chromosome has been reported in many organisms, including humans, plants, and S. cerevisiae [7,26,27,28,29]. However, the chance of the circularization of all chromosomes is very low if the chromosome number is not low. S. pombe is a unique organism in that there is a high chance of circularization of all three chromosomes. Unlike trt1 knockout, the deletion of pot1 results in rapid telomere shortening and three circular chromosomes in the survivors [10]. It remains unclear why deletion of pot1 does not result in survivors maintaining telomeres by HR. One possible reason is that extensive degradation of chromosome ends results in the complete loss of template DNA, which is necessary for HR at telomeres. Chromosome fusion in the pot1 knockout is mediated by single-strand annealing (SSA) [30]. Both ends of chromosomes I and II have regions of extensive homology, which are used for SSA when pot1 is deleted. Interestingly, knockout of genes required for SSA, such as rad22 (fission yeast homologs of Rad52), srs2, rad16 (ERCC1/XPF endonuclease), and lig4 are synthetically lethal with pot1 deletion. Both ends of chromosome III have long arrays of rDNA repeats adjacent to telomere repeats but lack regions of extensive homology, which are used for SSA. The mechanism of chromosome III circularization in the pot1 disruptant remains unclear. A rad3 tel1 (homologues of ATR and ATM, respectively) double mutant with three circular chromosomes contains microhomologies of three and thirteen nucleotides at the fusion site in the rDNA region of chromosome III, implying that the mechanism of chromosome fusion in chromosome III is different from that in other chromosomes [31]. Degradation of 5′-single-stranded DNA is required for chromosome fusion by SSA in uncapped telomeres. Double deletion of rqh1 (human BLM/S. cerevisiae SGS1) and exo1 slows down the degradation of 5′-single-stranded DNA of the telomere end after pot1 shut-off, suggesting that Rqh1 and Exo1 play redundant roles in the degradation of uncapped telomeres [32]. Deletion of rqh1 and/or exo1 suppresses acute growth defect induced by pot1 shut-off, further supporting this idea [32]. However, the chromosome ends in the pot1 rqh1 exo1 triple mutant were thought to be fused by SSA, implying that unknown nucleases can resect uncapped telomeres in the absence of both Exo1 and Rqh1 [33].
Although there is no clear sequence homology, the functional counterpart of S. pombe Pot1 is Cdc13 in S. cerevisiae [34]. Cdc13 forms a complex with Stn1 and Ten1 and this binds to the single-stranded telomeric DNA [35]. Inactivation of Cdc13 using the cdc13-1 ts mutant results in degradation of telomeric DNA and DNA damage checkpoint dependent cell cycle arrest [36]. The nucleases responsible for the degradation in the cdc13-1 ts mutant are Exo1 and the Pif1 helicase with an unknown nuclease [37]. A common function of human POT1, S. pombe Pot1, and S. cerevisiae Cdc13 is the protection of telomere overhang from DNA damage checkpoint activation. Unlike S. pombe and S. cerevisiae, knock down of human POT1 does not result in rapid telomere degradation. This may be due to high NHEJ preference in humans and high HR preference in S. pombe and S. cerevisiae, which would require high nuclease activity for HR or SSA initiation.
3. Lethality of pot1 rqh1 Double Mutant Is Suppressed by Inactivation of Homologous Recombination
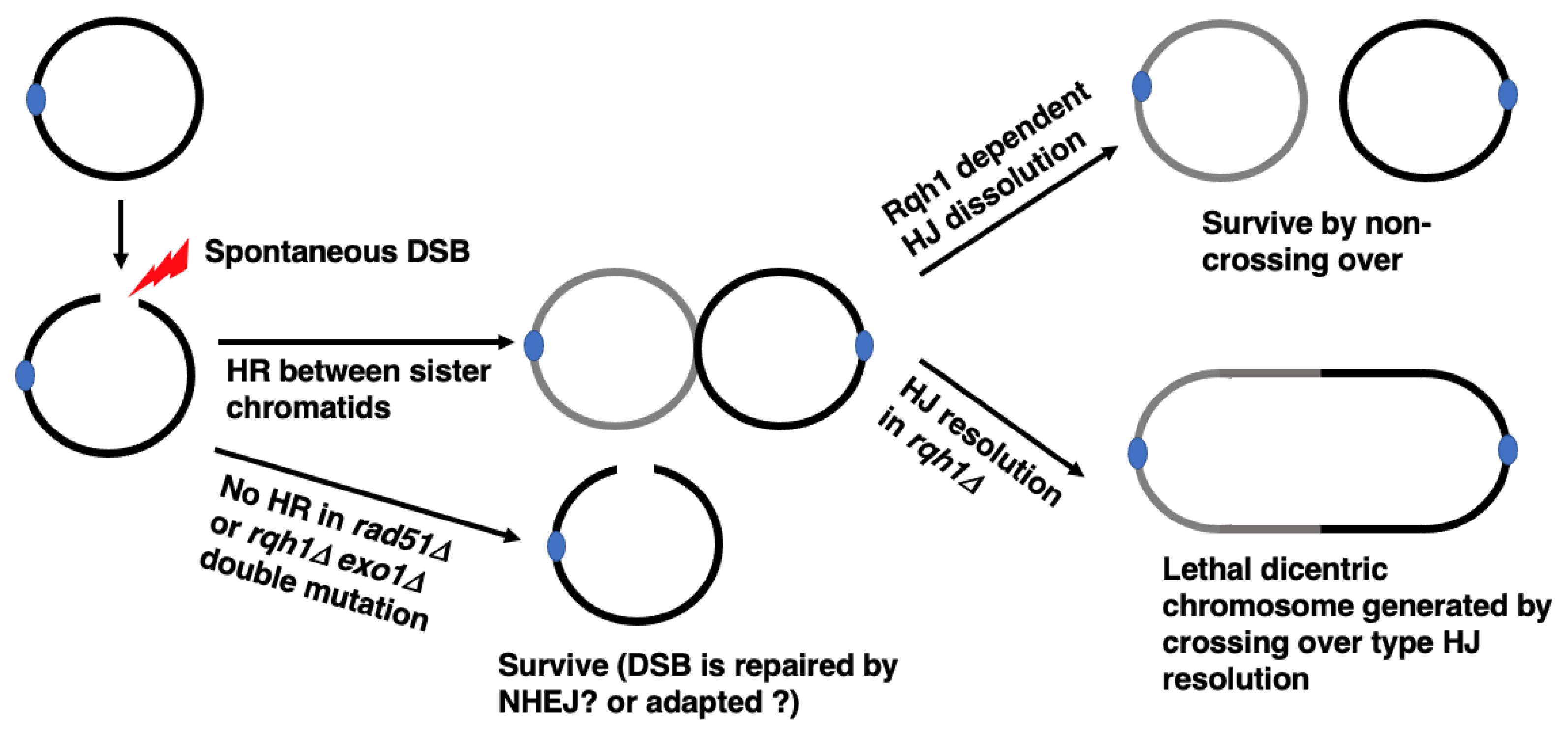
S. pombe Rqh1 is a conserved RecQ helicase involved in several steps of HR [38]. Deletion of rqh1 results in hyperrecombination and the accumulation of recombination intermediates [39,40]. These results and those of other studies suggest that Rqh1 inhibits strand invasion and promotes the resolution of recombination intermediates. Moreover, it has been suggested that Rqh1 is involved in the resection of DNA Double Strand Break (DSB) ends [32,33,41,42]. pot1 and rqh1 are synthetically lethal [30], for two possible reasons, the first being that the chromosome cannot be fused due to defects in resection and SSA [30]. The second reason is that circular chromosomes are not maintained. The chromosome fusion is detected after pot1 shut-off in the rqh1-deletion background, ruling out the first possibility regarding chromosome fusion [33]. To test the second possibility (that the circular chromosomes cannot be maintained in the absence of Rqh1), we screened genes that suppress the lethality of the pot1 rqh1 double mutant. We found that its lethality was suppressed by the deletion of rad51, which plays a central role in HR [33]. The lethality of the pot1 rqh1 double mutant was also suppressed by deletion of exo1, which is thought to play a redundant role with Rqh1 in the resection of DNA DSB ends that is essential for HR initiation [32,33,41,42]. Therefore, it is likely that HR activity was completely blocked in the rqh1 exo1 double mutant (Figure 2). These results suggest that the lethality of pot1 rqh1 is suppressed by HR inactivation. Rqh1 inhibits crossing over, which can produce circular dimers with two centromeres when a DSB in circular chromosomes is repaired by HR between sister chromatids. The shut-off of rqh1 in the pot1 disruptant, which has a circular chromosome, results in chromosome segregation defects. Considered together, we concluded that Rqh1 is required for the maintenance of circular chromosomes by inhibiting the production of circular dimers (Figure 2).
Top3 functions with Rqh1 to inhibit crossover by dissolution of double Holliday junctions (HJ) [43], suggesting that Top3 is required for the maintenance of circular chromosomes by inhibiting the production of circular dimers. top3 is an essential gene in S. pombe, but the top3 rqh1-helicase dead (rqh1-hd) double mutant is viable [44]. As discussed later, the pot1 rqh1-hd double mutant is also viable [45]. We found that the pot1 rqh1-hd top3 triple mutant was lethal, suggesting that Top3 is required for the maintenance of circular chromosomes. Interestingly, the lethality of the pot1 rqh1-hd top3 triple mutant was also suppressed by rad51 deletion [33]. These facts further suggest that the inhibition of crossover is critical for maintaining circular chromosomes.
4. pot1 rqh1-hd Cells Maintain Telomeres by HR and Accumulate Recombination Intermediates at Telomeres
The pot1 rqh1 double mutant is lethal, whereas the pot1 rqh1-hd double mutant is not [45]. This is surprising because the helicase activity of Rqh1 is thought to constitute its main function [46]. To study why the pot1 rqh1-hd double mutant was not lethal, we characterized the phenotype of the pot1 rqh1-hd double mutant. The pot1 rqh1-hd double mutant had a stronger subtelomere signal than wild-type cells, suggesting that chromosome ends are maintained by HR using subtelomeric DNA as a template [45]. This subtelomere signal is completely abolished through the deletion of rad51, further suggesting that chromosome ends are maintained by HR in the pot1 rqh1-hd double mutant. Next, we analyzed chromosome ends using PFGE to confirm that the chromosomes of the pot1 rqh1-hd double mutant were linear. We could not detect any bands corresponding to the chromosome end fragments using PFGE. DNA with branched structures such as recombination intermediates cannot enter a pulsed-field gel. As Rqh1 is required for the resolution of recombination intermediates [40], these facts suggest that recombination intermediates accumulate at the chromosome ends in the pot1 rqh1-hd double mutant. It remains unclear why the pot1 rqh1 null double mutant cannot maintain telomeres by HR. I assume that the Rqh1-hd protein may prevent nuclease-dependent rapid telomere degradation as seen in the pot1 disruptant, which could increase the chance of HR-dependent telomere maintenance.
The phenotype of the pot1 rqh1-hd double mutant was unique compared to that of the trt1 disruptant that maintains telomeres by HR, because the recombination intermediates do not accumulate at the telomere in the trt1 disruptant. The pot1 rqh1-hd double mutant had defects in chromosome segregation and was sensitive to the anti-microtubule drug thiabendazole (TBZ) [45]. These phenotypes can be explained as follows: the presence of recombination intermediates at telomeres prevents chromosome segregation. Inhibition in mitotic spindles by TBZ increases chromosome segregation defect in the pot1 rqh1-hd double mutant. TBZ sensitivity of pot1 rqh1-hd double mutant is suppressed by mutation in chk1, cdc2-3w, or wee1 mik1 double mutation [47,48]. All of these mutations shorten G2 phase length, suggesting that long G2 accumulates recombination intermediates in pot1 rqh1-hd double mutant that make cells sensitive to TBZ.
Surprisingly, the spindle checkpoint was activated in the pot1 rqh1-hd double mutant [47]. The spindle checkpoint monitors defects in kinetochore–microtubule attachment or lack of tension at the kinetochores. Therefore, our results suggest that accumulation of recombination intermediates at the telomere (or elsewhere) somehow induces defects in kinetochore–microtubule attachment or a lack of tension at the kinetochores. Indeed, the growth of the pot1 rqh1-hd double mutant is made worse by the deletion of gene encoding spindle checkpoint proteins Mad2 or Bub1 [47]. Moreover, Mad2 foci persisted longer than usual at kinetochores in the double mutant, suggesting a defect in kinetochore–microtubule attachment in the pot1 rqh1-hd double mutant. It remains unclear how the accumulation of recombination intermediates at the telomere induces defects in kinetochore-microtubule attachment.
Centromeric adaptor, shugoshin Sgo2, mainly binds to subtelomeres during interphase, but binds to centromeres during mitosis [49], implying that the function of proteins that localize to both telomeres and centromeres may be affected in the pot1 rqh1 double mutant. CENP–S and CENP–X, also called Mhf1 and Mhf2, respectively, localize to the centromere and play a role in recombination and repair [50]. This implies that the function of proteins that localize to both recombination sites and centromeres may be affected in the pot1 rqh1-hd double mutant. Further investigation will reveal the mechanism by which the accumulation of recombination intermediates at the telomere induces defects in kinetochore–microtubule attachment.
5. pot1 and Genes Encoding the Chromosomal Passenger Complex (CPC) Are Lethal
The chromosomal passenger complex (CPC) comprises Aurora-B protein kinase, the inner centromere protein INCENP, survivin, and borealin. The CPC plays a crucial role in mitosis [51]. The S. pombe CPC consists of the aurora-related kinase Ark1, survivin homolog Bir1, and inner centromere protein Pic1 [52]. The S. pombe CPC is important for proper bipolar attachment of sister chromatids during metaphase [53]. Interestingly, we found that genes encoding the CPC, ark1, bir1, and pic1 genetically interact with pot1 in fission yeast [54]. ark1, bir1, and pic1 are essential for growth but temperature-sensitive (ts) mutants can grow at a semi-permissive temperature. We found that the double mutants of pot1 and ark1 or bir1 or pic1 ts alleles were synthetically lethal at semi-permissive temperatures [54]. The pot1 pic1 double mutant, which has circular chromosomes, is viable at a permissive temperature (25 °C). This strain lost viability completely at a semi-permissive temperature (33 °C), whereas the pic1 single mutant could survive at 33 °C. These facts suggest that Pic1 and most likely the CPC are required for the growth of strains that have circular chromosomes. The trt1 pic1 double mutant, which has circular chromosomes at a permissive temperature (25 °C), showed the same phenotype as the pot1 pic1 double mutant, further demonstrating that the CPC is required for the growth of strains with circular chromosomes.
Sgo2 is required for centromeric localization of the Aurora kinase complex [55]. However, the sgo2 pot1 double mutant is not synthetically lethal [54]. Therefore, the function of CPC, which is not related to Sgo2 function, is important for the growth of cells with circular chromosomes. The pot1 pic1 double mutant displayed elevated rates of chromosome segregation defects at the permissive temperature (33 °C), suggesting that the role of the CPC in chromosome segregation is important for the growth of cells with circular chromosomes. One possible candidate for this function is condensin, because the condensin subunit Cnd2 is phosphorylated by the CPC subunit Ark1 throughout mitosis [56]. Circular chromosomes may have difficulty with chromosome condensation because they are topologically constrained. Identifying the target of Ark1, which is essential for the growth of cells that have circular chromosomes, will contribute to the understanding of why the pot1 cpc complex is synthetically lethal.
We also found interesting genetic interaction between pot1 and sgo2 and swi6 [54]. Both pot1 sgo2 double mutant and pot1 swi6 double mutant show higher chromosome segregation defects than each single mutant [54]. Both Sgo2 and Swi6 localize to centromere and (sub) telomere [57], implying that telomere loss may affect centromere function. Defect of dcr1 disruptant in pericentric heterochromatin silencing is suppressed by deletion of pot1, further suggesting that telomere loss may affect centromere function [58].
6. Histone H4 Acetylation and the Bromodomain Protein Bdf2 Are Required for the Growth of Cells with Circular Chromosomes
A ring chromosome is a cytogenetic hallmark of atypical lipomatous tumors and can be used to determine a proper diagnosis [59]. Chemicals that inhibit growth of cells that have circular chromosomes can be used as anti-cancer drugs for cancers in which most cells have a circular chromosome, such as dermatofibrosarcoma protuberans (70%) and atypical lipomatous tumors (85%) [60]. A chemical that inhibits the growth of fission yeast pot1 disruptant with circular chromosomes could be a candidate for this purpose. We found that (+)-JQ1, which is a potent, high-affinity, selective BET bromodomain inhibitor, inhibits the growth of fission yeast pot1 disruptant more than the control strain with linear chromosomes [61]. (+)-JQ1 displaces BRD4 from chromatin [62]. Human BRD4 binds to acetylated lysine in histone tails via a bromodomain [63]. The orthologs of human BRD4 are Bfd1 and Bfd2 in fission yeast. Therefore, one possible explanation for the growth inhibition of pot1 disruptant by (+)-JQ1 could be that (+)-JQ1 inhibits fission yeast Bfd1 and/or Bfd2, and fission yeast bfd1 and/or bfd2 are important for the growth of pot1 disruptants with circular chromosomes. pot1 bdf2, but not pot1 bdf1, was found to be synthetically lethal [61]. These results suggest that (+)-JQ1 inhibits fission yeast Bdf2. To study why the pot1 bdf2 double mutant is synthetically lethal, we created a bdf2-ts mutant that loses Bdf2 function at a high temperature (36 °C). The pot1 bdf2-ts double mutant with circular chromosomes is viable at a permissive temperature (25 °C) but shows a synthetic growth defect at 36 °C. These results demonstrated that Bdf2 plays an important role in the growth of cells with circular chromosomes.
mst1 encodes a histone acetyltransferase in fission yeast. Both Bdf2 and Mst1 are required for maintaining proper histone H4 acetylation level, suggesting functional overlap between Bdf2 and Mst1 [64,65]. Because Mst1 is essential for growth, we used the mst1-ts mutant [66]. We found that the pot1 mst1-ts double mutant, which has circular chromosomes at a permissive temperature (25 °C), showed a synthetic growth defect at a semi-permissive temperature (31 °C). These results suggested that maintaining proper histone H4 acetylation levels is important for the growth of cells with circular chromosomes. The change in histone H4 acetylation levels in bdf2 and mst1 mutants could affect whole-genome gene expression profiling. Therefore, analysis of changes in gene expression profiling in the bdf2 and mst1 mutants would improve our understanding of the importance of Bdf2 and Mst1 for the growth of cells that have circular chromosomes.
7. pot1 and Gene Encoding Phosphatidylinositol 4-Kinase Pik1 Is Lethal
Genes that genetically interact with pot1 can be screened using forward genetics. We found that a mutation in gene encoding phosphatidylinositol 4-kinase Pik1, called pik1-1, is synthetically lethal with pot1 [67,68]. Pik1 localizes to the Golgi and phosphorylates PI to produce PI(4)P [69]. The shut-off of pot1 in the pik1-1 mutant resulted in chromosome fusion, demonstrating that Pik1 is not required for chromosome circularization, suggesting that Pik1 is required for the growth of cells with circular chromosomes.
We found that the PI(4)P level in the Golgi was reduced in pik1-1, suggesting that PI(4)P in the Golgi is required for the growth of the pot1 disruptant. However, it has been argued that PI(4)P has a nuclear role in humans [70]. Moreover, S. cerevisiae Pik1 localizes to the nucleus in addition to the Golgi apparatus [71]. Therefore, although it remains unclear whether S. pombe Pik1 localizes to the nucleus [72], S. pombe Pik1 may function to regulate PI(4)P levels in the nucleus and/or in the nuclear membrane, and this may contribute to the growth of cells with circular chromosomes. It is also possible that the pik1 mutation may affect the sorting of proteins necessary for the maintenance of chromosome circularization due to defects in Golgi functions.
8. The Effect of Chromosome Circularization in Torsional Constraint, DNA Damage, and Chromosome Organization
DNA replication and transcription generate a DNA topological strain that can inhibit DNA replication and transcription if the DNA topological strain is not removed [73]. Excessive supercoiling is mitigated by topoisomerases (I and II). In S. cerevisiae, DNA is not torsionally constrained at the chromosomal ends, making it difficult to evaluate the impact of torsional constraints when the chromosome is linear [74]. The circular chromosome provides an excellent model system to study torsional constraints because the chromosome ends are physically fused. Cells with circular chromosomes are highly sensitive to the topoisomerase I inhibitor, camptothecin CPT [61,75]. We found that DNA topoisomerase II poisons, such as mitoxantrone and its analog, inhibit the growth of cells with circular chromosomes more efficiently than cells with linear chromosomes (manuscript in preparation). These results demonstrate that cells with circular chromosomes are useful for studying the importance of torsional constraint and screening novel topoisomerase inhibitors and proteins that are involved in the regulation of torsional constraint.
Five percent of the pot1 disruptants have RPA foci (a marker of DNA damage), compared to ~two percent of wild-type cells, suggesting that circular chromosomes have more risk of spontaneous DNA damage [76]. This could be because of the higher torsional constraint in the pot1 disruptant. Consistently, the pot1 disruptant is more sensitive to the alkylating agent methyl-methanesulfonate MMS than the wild-type strain [77]. The Rad9-Rad1-Hus1 (9-1-1) complex involved in DNA damage checkpoint is loaded onto chromatin when DNA replication is halted [78]. We found that double mutants between pot1 and genes encoding the 9-1-1 checkpoint complex (Rad9-Rad1-Hus1) are more sensitive to DNA replication inhibitors, such as hydroxyurea (HU), than each single mutant [76]. These results demonstrate that DNA integrity checkpoints play an essential role in maintaining the viability of strains with circular chromosomes in the presence of DNA replication inhibitors. In other words, strains with circular chromosomes have a higher risk of genomic instability during DNA replication. These findings also demonstrate that screening of genes that genetically interact with pot1 in the presence of DNA replication inhibitors will be useful for identifying and studying proteins involved in DNA damage checkpoint.
Telomeres are attached to the nuclear envelope by the interaction between the nuclear envelope protein Bqt4 and the telomere-associated protein Rap1 in fission yeast [79]. When telomere-binding proteins are visualized using a fluorescent protein, one or two dots can be detected, indicating that telomeres are clustered [80]. However, the importance of telomere clustering and tethering to the nuclear envelope remains unclear. It is possible that the pot1 disruptant, which has circular chromosomes, loses the tethering of telomeres to the nuclear envelope because more than 10 kb of the telomere ends are lost [30]. Moreover, 3D genome organization may be affected by pot1 disruption. Therefore, screening of genes that genetically interact with pot1 will be useful for studying the importance of telomere tethering to the nuclear envelope and telomere clustering.
Recently, single-chromosome fission yeast models have been reported [81]. The chromosome fusions in these strains caused few defects in cell morphology and growth and had a limited impact on gene expression. As these strains have a linear chromosome, it would be interesting to compare the phenotype of the pot1 disruptant that has a single circular chromosome with the strain that has a single liner chromosome.
9. Biology of Yeast Carrying Ring Chromosomes and Comparison of the Difference between Six Types of S. pombe Mutants That Have Circular Chromosomes
There are six types of mutant that have circular chromosomes in S. pombe, namely, pot1/tpz1 disruptant [22], trt1 disruptant [25], rad3 tel1 double mutant [31], poz1 ccq1 double mutant [22], ten1/ste1 disruptant [82], and taz1 rad11 double mutant [83]. A common feature of these survivors is the complete loss of the telomeric repeat sequence. Sexual reproduction is severely affected with reduced spore viability in trt1 disruptant and rad3 tel1 double mutant [31], demonstrating that circular chromosomes without telomere sequence cause problems during meiosis in S. pombe [25,31,84].
There are similarities and differences between the six types of S. pombe mutant that have circular chromosomes. The trt1 disruptant and rad3 tel1 double mutant lose telomere gradually before chromosome circularization, while the pot1/tpz1 disruptant, poz1 ccq1 double mutant, taz1 rad11 double mutant, and ten1/ste1 disruptant lose telomere rapidly [22,25,31,82,83]. These facts allow us to categorize these mutants into two groups (rapid telomere loss group and gradual telomere loss group). Regarding Rad3 and Tel1 phosphorylates Ccq1 which recruit telomerase, the Trt1 recruitment is mostly eliminated in the rad3 tel1 double mutant [85], suggesting that the mechanism of telomere loss in the rad3 tel1 double mutant is due to a defect in the recruitment of telomerase, explaining why Rad3 and Tel1 are in the gradual telomere loss group. Pot1, Tpz1, Poz1, and Ccq1 form a protein complex to protect the telomere end [22], explaining why they are in the rapid telomere loss group. Overexpression of Pot1 suppresses rapid telomere loss in the taz1 rad11 double mutant, suggesting that the mechanism of telomere loss in the taz1 rad11 double mutant is related to a defect in the Pot1 function [83]. Although deletion of stn1/ten1 also results in rapid telomere loss, the phenotype of the stn1-1 ts mutant is frequent replication fork collapses specifically in the subtelomere regions, suggesting that the mechanism of telomere loss in stn1 disruptant is different from that in the pot1 disruptant [75].
10. Conclusions and Perspectives on Genes which Genetically Interact with the Telomere Protection Gene pot1 in Fission Yeast
This review demonstrates that the screening of genes that genetically interact with fission yeast pot1 provides many potential genes that play important roles in chromosome maintenance. Genetic interactions with pot1 are summarized in Table 1. Proteins involved in SSA are essential for the growth of pot1 disruptants because no chromosomal fusion would result in the extensive degradation of uncapped telomeres. The Rqh1-Top3 complex is essential for the maintenance of circular chromosomes, probably because spontaneous DNA breaks are repaired by HR, producing circular chromosome dimers that induce breakage–fusion–bridge cycles, resulting in cell death. The genes encoding CPC are also synthetically lethal with pot1, suggesting that the CPC is important for chromosome segregation in strains with circular chromosomes. Moreover, Bdf2 and Mst1, which are required for maintaining proper histone H4 acetylation levels, are required for the growth of cells with circular chromosomes. Changes in histone H4 acetylation levels may affect whole-genome gene expression. Therefore, the analysis of gene expression in bdf2 and mst1 mutants will be the next step in understanding the importance of Bdf2 and Mst1 in the growth of cells with circular chromosomes.
A strain with a linear chromosome should deal with topological stress during DNA replication, transcription, and chromosome segregation. A strain with circular chromosomes is more sensitive to topoisomerase I and II inhibition, suggesting that circular chromosomes are topologically constrained. These results demonstrate that a strain with circular chromosomes is a useful model system for studying how cells deal with topological stress. The growth of the strain that has circular chromosomes depends on the 9-1-1 DNA damage checkpoint function when DNA replication is inhibited, suggesting that circular chromosomes are more vulnerable to replication inhibition than linear chromosomes. Finally, phosphatidylinositol 4-kinase Pik1 is essential for the growth of pot1 disruptants, probably because of its role in the growth of cells with circular chromosomes. Although the mechanism of this finding remains unclear, our results suggest the importance of the regulation of PI(4)P levels in the Golgi and possibly in the nucleus. In addition to genetic interaction discussed here, genome-wide genetic interaction analysis for S. pombe revealed that pot1 has negative genetic interactome with atp10 and prz1 and positive genetic interaction with SPAC227.17c [86]. However, these interactions are not studied in detail. Further research will be necessary to understand the mechanism of these genetic interactions.
Prokaryotes such as E. coli have a circular chromosome. In contrast, eukaryotes, including yeasts and humans, have linear chromosomes. One evolutionary question is why natural selection chooses linear chromosomes in eukaryotes, even though linear chromosomes have DNA end replication problems. The studies presented in this review clearly demonstrate that the stable maintenance of circular chromosomes is more difficult than that of linear chromosomes during mitosis. Prokaryotes only have one circular chromosome, whereas eukaryotes have multiple linear chromosomes. The HR between sister chromatids in circular chromosomes can create circular chromosome dimers, which in E. coli are converted to monomers by Xer site-specific recombination [87]. The resolution of dimeric chromosomes using Xer site-specific recombination is possible for a single circular chromosome, but it may be more difficult if E. coli has more than two circular chromosomes. This may be the reason why prokaryotes have a single circular chromosome and it makes it difficult to increase the genome size in prokaryotes. In contrast, linear chromosomes do not require an Xer site-specific recombination system, making it possible to maintain more than two linear chromosomes without problems. This could be the reason why eukaryotes can maintain multiple linear chromosomes that can easily increase their genome size. Therefore, during evolution, eukaryotes might have acquired linear chromosomes with a telomere maintenance system before they acquire a sexual reproduction system. The importance of telomeres in meiosis also could be the reason why natural selection chooses linear chromosomes in eukaryotes, which is discussed in the review by the Ishikawa group [88].
In conclusion, the screening of genes that genetically interact with fission yeast pot1 is a powerful method for studying the role of proteins required for telomere fusion, telomere degradation, and the maintenance of circular chromosomes. These studies are also informative for understanding the mechanism underlying the maintenance of linear chromosomes.
Funding
This work was supported by MEXT/JSPS KAKENHI Grant Number 20K06488.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I thank Jo Murray (University of Sussex) for critical reading and comments on the manuscript.
Conflicts of Interest
No potential conflict of interest were disclosed.
References
- Turner, K.J.; Vasu, V.; Griffin, D.K. Telomere Biology and Human Phenotype. Cells 2019, 8, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armanios, M. The Role of Telomeres in Human Disease. Annu. Rev. Genom. Hum. Genet. 2022, 23, 363–381. [Google Scholar] [CrossRef] [PubMed]
- de Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feijoo, P.; Dominguez, D.; Tusell, L.; Genesca, A. Telomere-dependent genomic integrity: Evolution of the fusion-bridge-breakage cycle concept. Curr. Pharm. Des. 2014, 20, 6375–6385. [Google Scholar] [CrossRef]
- Casari, E.; Gnugnoli, M.; Rinaldi, C.; Pizzul, P.; Colombo, C.V.; Bonetti, D.; Longhese, M.P. To Fix or Not to Fix: Maintenance of Chromosome Ends Versus Repair of DNA Double-Strand Breaks. Cells 2022, 11, 3224. [Google Scholar] [CrossRef]
- Reddel, R.R. Telomere maintenance mechanisms in cancer: Clinical implications. Curr. Pharm. Des. 2014, 20, 6361–6374. [Google Scholar] [CrossRef] [Green Version]
- Pristyazhnyuk, I.E.; Menzorov, A.G. Ring chromosomes: From formation to clinical potential. Protoplasma 2018, 255, 439–449. [Google Scholar] [CrossRef]
- Yip, M.Y. Autosomal ring chromosomes in human genetic disorders. Transl. Pediatr. 2015, 4, 164–174. [Google Scholar] [CrossRef]
- Gebhart, E. Ring chromosomes in human neoplasias. Cytogenet. Genome Res. 2008, 121, 149–173. [Google Scholar] [CrossRef] [Green Version]
- Baumann, P.; Cech, T.R. Pot1, the putative telomere end-binding protein in fission yeast and humans. Science 2001, 292, 1171–1175. [Google Scholar] [CrossRef] [Green Version]
- Baumann, P.; Podell, E.; Cech, T.R. Human Pot1 (protection of telomeres) protein: Cytolocalization, gene structure, and alternative splicing. Mol. Cell. Biol. 2002, 22, 8079–8087. [Google Scholar] [CrossRef] [Green Version]
- Aramburu, T.; Plucinsky, S.; Skordalakes, E. POT1-TPP1 telomere length regulation and disease. Comput. Struct. Biotechnol. J. 2020, 18, 1939–1946. [Google Scholar] [CrossRef]
- Abreu, E.; Aritonovska, E.; Reichenbach, P.; Cristofari, G.; Culp, B.; Terns, R.M.; Lingner, J.; Terns, M.P. TIN2-tethered TPP1 recruits human telomerase to telomeres in vivo. Mol. Cell. Biol. 2010, 30, 2971–2982. [Google Scholar] [CrossRef] [Green Version]
- Xin, H.; Liu, D.; Wan, M.; Safari, A.; Kim, H.; Sun, W.; O’Connor, M.S.; Songyang, Z. TPP1 is a homologue of ciliate TEBP-beta and interacts with POT1 to recruit telomerase. Nature 2007, 445, 559–562. [Google Scholar] [CrossRef]
- Veldman, T.; Etheridge, K.T.; Counter, C.M. Loss of hPot1 function leads to telomere instability and a cut-like phenotype. Curr. Biol. 2004, 14, 2264–2270. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Stock, A.J.; Liu, Y. The enigma of excessively long telomeres in cancer: Lessons learned from rare human POT1 variants. Curr. Opin. Genet. Dev. 2020, 60, 48–55. [Google Scholar] [CrossRef]
- Wang, F.; Podell, E.R.; Zaug, A.J.; Yang, Y.; Baciu, P.; Cech, T.R.; Lei, M. The POT1-TPP1 telomere complex is a telomerase processivity factor. Nature 2007, 445, 506–510. [Google Scholar] [CrossRef] [Green Version]
- Robles-Espinoza, C.D.; Harland, M.; Ramsay, A.J.; Aoude, L.G.; Quesada, V.; Ding, Z.; Pooley, K.A.; Pritchard, A.L.; Tiffen, J.C.; Petljak, M.; et al. POT1 loss-of-function variants predispose to familial melanoma. Nat. Genet. 2014, 46, 478–481. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.T.; Hennick, K.; Johnson, J.; Finnerty, B.; Choo, S.; Short, S.B.; Drubin, C.; Forster, R.; McMaster, M.L.; Hockemeyer, D. Cancer-associated POT1 mutations lead to telomere elongation without induction of a DNA damage response. Embo J. 2021, 40, e107346. [Google Scholar] [CrossRef]
- Ramsay, A.J.; Quesada, V.; Foronda, M.; Conde, L.; Martínez-Trillos, A.; Villamor, N.; Rodríguez, D.; Kwarciak, A.; Garabaya, C.; Gallardo, M.; et al. POT1 mutations cause telomere dysfunction in chronic lymphocytic leukemia. Nat. Genet. 2013, 45, 526–530. [Google Scholar] [CrossRef]
- Jain, D.; Cooper, J.P. Telomeric strategies: Means to an end. Annu. Rev. Genet. 2010, 44, 243–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Kanoh, J.; Saito, M.; Ishikawa, F. Fission yeast Pot1-Tpp1 protects telomeres and regulates telomere length. Science 2008, 320, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Bunch, J.T.; Bae, N.S.; Leonardi, J.; Baumann, P. Distinct requirements for Pot1 in limiting telomere length and maintaining chromosome stability. Mol. Cell. Biol. 2005, 25, 5567–5578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase catalytic subunit homologs from fission yeast and human. Science 1997, 277, 955–959. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.M.; Cooper, J.P.; Cech, T.R. Two modes of survival of fission yeast without telomerase. Science 1998, 282, 493–496. [Google Scholar] [CrossRef] [Green Version]
- McClintock, B. A Correlation of Ring-Shaped Chromosomes with Variegation in Zea Mays. Proc. Natl. Acad. Sci. USA 1932, 18, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Yang, R.; Li, Y.; Su, Y.; Shen, Y.; Tang, D.; Luo, Q.; Cheng, Z. A functional centromere lacking CentO sequences in a newly formed ring chromosome in rice. J. Genet. Genom. 2016, 43, 694–701. [Google Scholar] [CrossRef]
- Warmington, J.R.; Anwar, R.; Newlon, C.S.; Waring, R.B.; Davies, R.W.; Indge, K.J.; Oliver, S.G. A ‘hot-spot’ for Ty transposition on the left arm of yeast chromosome III. Nucleic Acids Res. 1986, 14, 3475–3485. [Google Scholar] [CrossRef] [Green Version]
- Haber, J.E.; Thorburn, P.C.; Rogers, D. Meiotic and mitotic behavior of dicentric chromosomes in Saccharomyces cerevisiae. Genetics 1984, 106, 185–205. [Google Scholar] [CrossRef]
- Wang, X.; Baumann, P. Chromosome fusions following telomere loss are mediated by single-strand annealing. Mol. Cell. 2008, 31, 463–473. [Google Scholar] [CrossRef]
- Naito, T.; Matsuura, A.; Ishikawa, F. Circular chromosome formation in a fission yeast mutant defective in two ATM homologues. Nat. Genet. 1998, 20, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Nanbu, T.; Nguyễn, L.C.; Habib, A.G.; Hirata, N.; Ukimori, S.; Tanaka, D.; Masuda, K.; Takahashi, K.; Yukawa, M.; Tsuchiya, E.; et al. Fission Yeast Exo1 and Rqh1-Dna2 Redundantly Contribute to Resection of Uncapped Telomeres. PLoS ONE 2015, 10, e0140456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanbu, T.; Takahashi, K.; Murray, J.M.; Hirata, N.; Ukimori, S.; Kanke, M.; Masukata, H.; Yukawa, M.; Tsuchiya, E.; Ueno, M. Fission yeast RecQ helicase Rqh1 is required for the maintenance of circular chromosomes. Mol. Cell. Biol. 2013, 33, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mersaoui, S.Y.; Wellinger, R.J. Fine tuning the level of the Cdc13 telomere-capping protein for maximal chromosome stability performance. Curr. Genet. 2019, 65, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.; Skordalakes, E. Structure and function of the telomeric CST complex. Comput. Struct. Biotechnol. J. 2016, 14, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Garvik, B.; Carson, M.; Hartwell, L. Single-stranded DNA arising at telomeres in cdc13 mutants may constitute a specific signal for the RAD9 checkpoint. Mol. Cell. Biol. 1995, 15, 6128–6138. [Google Scholar] [CrossRef] [Green Version]
- Dewar, J.M.; Lydall, D. Similarities and differences between “uncapped” telomeres and DNA double-strand breaks. Chromosoma 2012, 121, 117–130. [Google Scholar] [CrossRef]
- Gupta, S.V.; Schmidt, K.H. Maintenance of Yeast Genome Integrity by RecQ Family DNA Helicases. Genes 2020, 11, 205. [Google Scholar] [CrossRef] [Green Version]
- Stewart, E.; Chapman, C.R.; Al-Khodairy, F.; Carr, A.M.; Enoch, T. rqh1+, a fission yeast gene related to the Bloom’s and Werner’s syndrome genes, is required for reversible S phase arrest. Embo J. 1997, 16, 2682–2692. [Google Scholar] [CrossRef] [Green Version]
- Doe, C.L.; Dixon, J.; Osman, F.; Whitby, M.C. Partial suppression of the fission yeast rqh1(−) phenotype by expression of a bacterial Holliday junction resolvase. Embo J. 2000, 19, 2751–2762. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Huang, J. DNA End Resection: Facts and Mechanisms. Genom. Proteom. Bioinform. 2016, 14, 126–130. [Google Scholar] [CrossRef] [Green Version]
- Tisi, R.; Vertemara, J.; Zampella, G.; Longhese, M.P. Functional and structural insights into the MRX/MRN complex, a key player in recognition and repair of DNA double-strand breaks. Comput. Struct. Biotechnol. J. 2020, 18, 1137–1152. [Google Scholar] [CrossRef]
- Bizard, A.H.; Hickson, I.D. The dissolution of double Holliday junctions. Cold Spring Harb. Perspect Biol. 2014, 6, a016477. [Google Scholar] [CrossRef]
- Ahmad, F.; Stewart, E. The N-terminal region of the Schizosaccharomyces pombe RecQ helicase, Rqh1p, physically interacts with Topoisomerase III and is required for Rqh1p function. Mol. Genet. Genom. 2005, 273, 102–114. [Google Scholar] [CrossRef]
- Takahashi, K.; Imano, R.; Kibe, T.; Seimiya, H.; Muramatsu, Y.; Kawabata, N.; Tanaka, G.; Matsumoto, Y.; Hiromoto, T.; Koizumi, Y.; et al. Fission yeast Pot1 and RecQ helicase are required for efficient chromosome segregation. Mol. Cell. Biol. 2011, 31, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Laursen, L.V.; Ampatzidou, E.; Andersen, A.H.; Murray, J.M. Role for the fission yeast RecQ helicase in DNA repair in G2. Mol. Cell. Biol. 2003, 23, 3692–3705. [Google Scholar] [CrossRef] [Green Version]
- Nakano, A.; Masuda, K.; Hiromoto, T.; Takahashi, K.; Matsumoto, Y.; Habib, A.G.; Darwish, A.G.; Yukawa, M.; Tsuchiya, E.; Ueno, M. Rad51-dependent aberrant chromosome structures at telomeres and ribosomal DNA activate the spindle assembly checkpoint. Mol. Cell. Biol. 2014, 34, 1389–1397. [Google Scholar] [CrossRef] [Green Version]
- Habib, A.G.; Masuda, K.; Yukawa, M.; Tsuchiya, E.; Ueno, M. Long G2 accumulates recombination intermediates and disturbs chromosome segregation at dysfunction telomere in Schizosaccharomyces pombe. Biochem. Biophys. Res. Commun. 2015, 464, 140–146. [Google Scholar] [CrossRef]
- Kanoh, J. Unexpected roles of a shugoshin protein at subtelomeres. Genes Genet. Syst. 2018, 92, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Osman, F.; Feeney, L.; Lorenz, A.; Bryer, C.; Whitby, M.C. MHF1-2/CENP-S-X performs distinct roles in centromere metabolism and genetic recombination. Open Biol. 2013, 3, 130102. [Google Scholar] [CrossRef] [Green Version]
- Ruchaud, S.; Carmena, M.; Earnshaw, W.C. Chromosomal passengers: Conducting cell division. Nat. Rev. Mol. Cell. Biol. 2007, 8, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Leverson, J.D.; Huang, H.K.; Forsburg, S.L.; Hunter, T. The Schizosaccharomyces pombe aurora-related kinase Ark1 interacts with the inner centromere protein Pic1 and mediates chromosome segregation and cytokinesis. Mol. Biol. Cell. 2002, 13, 1132–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y. Temporal and spatial regulation of targeting aurora B to the inner centromere. Cold Spring Harb. Symp. Quant. Biol. 2010, 75, 419–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, A.G.K.; Sugiura, K.; Ueno, M. Chromosome passenger complex is required for the survival of cells with ring chromosomes in fission yeast. PLoS ONE 2018, 13, e0190523. [Google Scholar] [CrossRef] [Green Version]
- Vanoosthuyse, V.; Prykhozhij, S.; Hardwick, K.G. Shugoshin 2 regulates localization of the chromosomal passenger proteins in fission yeast mitosis. Mol. Biol. Cell. 2007, 18, 1657–1669. [Google Scholar] [CrossRef] [Green Version]
- Nakazawa, N.; Mehrotra, R.; Ebe, M.; Yanagida, M. Condensin phosphorylated by the Aurora-B-like kinase Ark1 is continuously required until telophase in a mode distinct from Top2. J. Cell. Sci. 2011, 124, 1795–1807. [Google Scholar] [CrossRef] [Green Version]
- Kanoh, J.; Sadaie, M.; Urano, T.; Ishikawa, F. Telomere binding protein Taz1 establishes Swi6 heterochromatin independently of RNAi at telomeres. Curr. Biol. 2005, 15, 1808–1819. [Google Scholar] [CrossRef] [Green Version]
- Tadeo, X.; Wang, J.; Kallgren, S.P.; Liu, J.; Reddy, B.D.; Qiao, F.; Jia, S. Elimination of shelterin components bypasses RNAi for pericentric heterochromatin assembly. Genes Dev. 2013, 27, 2489–2499. [Google Scholar] [CrossRef] [Green Version]
- Gisselsson, D.; Höglund, M.; Mertens, F.; Johansson, B.; Dal Cin, P.; Van den Berghe, H.; Earnshaw, W.C.; Mitelman, F.; Mandahl, N. The structure and dynamics of ring chromosomes in human neoplastic and non-neoplastic cells. Hum. Genet. 1999, 104, 315–325. [Google Scholar] [CrossRef]
- Trombetta, D.; Mertens, F.; Lonoce, A.; D’Addabbo, P.; Rennstam, K.; Mandahl, N.; Storlazzi, C.T. Characterization of a hotspot region on chromosome 12 for amplification in ring chromosomes in atypical lipomatous tumors. Genes Chromosomes Cancer 2009, 48, 993–1001. [Google Scholar] [CrossRef]
- Yasuda, M.; Habib, A.G.K.; Sugiura, K.; Shamim, H.M.; Ueno, M. The fission yeast bromodomain protein Bdf2 is required for the growth of cells with circular chromosomes. Biosci. Biotechnol. Biochem. 2022, 86, 224–230. [Google Scholar] [CrossRef]
- Pérez-Salvia, M.; Esteller, M. Bromodomain inhibitors and cancer therapy: From structures to applications. Epigenetics 2017, 12, 323–339. [Google Scholar] [CrossRef] [Green Version]
- Dey, A.; Chitsaz, F.; Abbasi, A.; Misteli, T.; Ozato, K. The double bromodomain protein Brd4 binds to acetylated chromatin during interphase and mitosis. Proc. Natl. Acad. Sci. USA 2003, 100, 8758–8763. [Google Scholar] [CrossRef] [Green Version]
- Garabedian, M.V.; Noguchi, C.; Ziegler, M.A.; Das, M.M.; Singh, T.; Harper, L.J.; Leman, A.R.; Khair, L.; Moser, B.A.; Nakamura, T.M.; et al. The double-bromodomain proteins Bdf1 and Bdf2 modulate chromatin structure to regulate S-phase stress response in Schizosaccharomyces pombe. Genetics 2012, 190, 487–500. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tadeo, X.; Hou, H.; Tu, P.G.; Thompson, J.; Yates, J.R., 3rd; Jia, S. Epe1 recruits BET family bromodomain protein Bdf2 to establish heterochromatin boundaries. Genes Dev. 2013, 27, 1886–1902. [Google Scholar] [CrossRef] [Green Version]
- Gómez, E.B.; Nugent, R.L.; Laria, S.; Forsburg, S.L. Schizosaccharomyces pombe histone acetyltransferase Mst1 (KAT5) is an essential protein required for damage response and chromosome segregation. Genetics 2008, 179, 757–771. [Google Scholar] [CrossRef] [Green Version]
- Sugihara, A.; Nguyen, L.C.; Shamim, H.M.; Iida, T.; Nakase, M.; Takegawa, K.; Senda, M.; Jida, S.; Ueno, M. Mutation in fission yeast phosphatidylinositol 4-kinase Pik1 is synthetically lethal with defect in telomere protection protein Pot1. Biochem. Biophys. Res. Commun. 2018, 496, 1284–1290. [Google Scholar] [CrossRef]
- Iida, N.; Yamao, F.; Nakamura, Y.; Iida, T. Mudi, a web tool for identifying mutations by bioinformatics analysis of whole-genome sequence. Genes Cells 2014, 19, 517–527. [Google Scholar] [CrossRef] [Green Version]
- Matsuyama, A.; Arai, R.; Yashiroda, Y.; Shirai, A.; Kamata, A.; Sekido, S.; Kobayashi, Y.; Hashimoto, A.; Hamamoto, M.; Hiraoka, Y.; et al. ORFeome cloning and global analysis of protein localization in the fission yeast Schizosaccharomyces pombe. Nat. Biotechnol. 2006, 24, 841–847. [Google Scholar] [CrossRef]
- Fáberová, V.; Kalasová, I.; Krausová, A.; Hozák, P. Super-Resolution Localisation of Nuclear PI(4)P and Identification of Its Interacting Proteome. Cells 2020, 9, 1191. [Google Scholar] [CrossRef]
- Strahl, T.; Hama, H.; DeWald, D.B.; Thorner, J. Yeast phosphatidylinositol 4-kinase, Pik1, has essential roles at the Golgi and in the nucleus. J. Cell. Biol. 2005, 171, 967–979. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Steinbach, S.K.; Desautels, M.; Hemmingsen, S.M. Essential role for Schizosaccharomyces pombe pik1 in septation. PLoS ONE 2009, 4, e6179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levens, D.; Baranello, L.; Kouzine, F. Controlling gene expression by DNA mechanics: Emerging insights and challenges. Biophys. Rev. 2016, 8, 259–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, R.S.; Piña, B.; Roca, J. Positional dependence of transcriptional inhibition by DNA torsional stress in yeast chromosomes. Embo J. 2010, 29, 740–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takikawa, M.; Tarumoto, Y.; Ishikawa, F. Fission yeast Stn1 is crucial for semi-conservative replication at telomeres and subtelomeres. Nucleic Acids Res. 2017, 45, 1255–1269. [Google Scholar] [CrossRef] [Green Version]
- Shamim, H.M.; Minami, Y.; Tanaka, D.; Ukimori, S.; Murray, J.M.; Ueno, M. Fission yeast strains with circular chromosomes require the 9-1-1 checkpoint complex for the viability in response to the anti-cancer drug 5-fluorodeoxyuridine. PLoS ONE 2017, 12, e0187775. [Google Scholar] [CrossRef] [Green Version]
- Jain, D.; Hebden, A.K.; Nakamura, T.M.; Miller, K.M.; Cooper, J.P. HAATI survivors replace canonical telomeres with blocks of generic heterochromatin. Nature 2010, 467, 223–227. [Google Scholar] [CrossRef]
- Kai, M.; Wang, T.S. Checkpoint responses to replication stalling: Inducing tolerance and preventing mutagenesis. Mutat. Res. 2003, 532, 59–73. [Google Scholar] [CrossRef]
- Matsuda, A.; Asakawa, H.; Haraguchi, T.; Hiraoka, Y. Spatial organization of the Schizosaccharomyces pombe genome within the nucleus. Yeast 2017, 34, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Chikashige, Y.; Yamane, M.; Okamasa, K.; Tsutsumi, C.; Kojidani, T.; Sato, M.; Haraguchi, T.; Hiraoka, Y. Membrane proteins Bqt3 and -4 anchor telomeres to the nuclear envelope to ensure chromosomal bouquet formation. J. Cell. Biol. 2009, 187, 413–427. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Ye, T.; Zhang, X.R.; Nie, L.; Wang, H.; Li, W.; Lu, R.; Fu, C.; Du, L.L.; Zhou, J.Q. Single-chromosome fission yeast models reveal the configuration robustness of a functional genome. Cell. Rep. 2022, 40, 111237. [Google Scholar] [CrossRef]
- Martín, V.; Du, L.L.; Rozenzhak, S.; Russell, P. Protection of telomeres by a conserved Stn1-Ten1 complex. Proc. Natl. Acad. Sci. USA 2007, 104, 14038–14043. [Google Scholar] [CrossRef] [Green Version]
- Kibe, T.; Ono, Y.; Sato, K.; Ueno, M. Fission yeast Taz1 and RPA are synergistically required to prevent rapid telomere loss. Mol. Biol. Cell. 2007, 18, 2378–2387. [Google Scholar] [CrossRef] [Green Version]
- Sadaie, M.; Naito, T.; Ishikawa, F. Stable inheritance of telomere chromatin structure and function in the absence of telomeric repeats. Genes Dev. 2003, 17, 2271–2282. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, H.; Tarumoto, Y.; Ishikawa, F. Tel1(ATM) and Rad3(ATR) phosphorylate the telomere protein Ccq1 to recruit telomerase and elongate telomeres in fission yeast. Genes Dev. 2012, 26, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Ryan, C.J.; Roguev, A.; Patrick, K.; Xu, J.; Jahari, H.; Tong, Z.; Beltrao, P.; Shales, M.; Qu, H.; Collins, S.R.; et al. Hierarchical modularity and the evolution of genetic interactomes across species. Mol. Cell. 2012, 46, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Sciochetti, S.A.; Piggot, P.J. A tale of two genomes: Resolution of dimeric chromosomes in Escherichia coli and Bacillus subtilis. Res. Microbiol. 2000, 151, 503–511. [Google Scholar] [CrossRef]
- Ishikawa, F.; Naito, T. Why do we have linear chromosomes? A matter of Adam and Eve. Mutat. Res. 1999, 434, 99–107. [Google Scholar] [CrossRef]
Figure 1.
Uncapping of telomeres leads to chromosome abnormalities. Telomere uncapping results in breakage–fusion–bridge (BFB) cycles or chromosome circularization. Inter-chromosomal fusion generates dicentric chromosomes. Segregation of the two centromeres towards opposite poles creates chromatin bridges, which will result in DSB. The DSB ends will be fused, generating dicentric chromosomes. This breakage–fusion–bridge (BFB) cycle results in translocations and aneuploid nuclei, which can drive cancer. Intra-chromosomal fusion generates circular chromosomes, which can be unstable, and result in chromosome breaks and/or rearrangement that could induce cancer.
Figure 1.
Uncapping of telomeres leads to chromosome abnormalities. Telomere uncapping results in breakage–fusion–bridge (BFB) cycles or chromosome circularization. Inter-chromosomal fusion generates dicentric chromosomes. Segregation of the two centromeres towards opposite poles creates chromatin bridges, which will result in DSB. The DSB ends will be fused, generating dicentric chromosomes. This breakage–fusion–bridge (BFB) cycle results in translocations and aneuploid nuclei, which can drive cancer. Intra-chromosomal fusion generates circular chromosomes, which can be unstable, and result in chromosome breaks and/or rearrangement that could induce cancer.

Figure 2.
Consequence of HR in cells with circular chromosomes in the absence or presence of Rqh1. Spontaneous DSB in a circular chromosome would result in Exo1 and/or Rqh1 dependent DSB end processing and Rad51 dependent generation of recombination intermediates between sister chromatids. This intermediate can be resolved without crossing over by Rqh1-Top3 dependent double Holliday junction (HJ) dissolution. In the absence of Rqh1, this intermediate will be resolved by nuclease(s), which could generate dicentric circular chromosome dimer by crossing over. Even if spontaneous DSB in a circular chromosome is not repaired by HR due to the defect in HR activity, this DSB would not result in cell death, possibly due to adaptation or to NHEJ dependent DSB repair.
Figure 2.
Consequence of HR in cells with circular chromosomes in the absence or presence of Rqh1. Spontaneous DSB in a circular chromosome would result in Exo1 and/or Rqh1 dependent DSB end processing and Rad51 dependent generation of recombination intermediates between sister chromatids. This intermediate can be resolved without crossing over by Rqh1-Top3 dependent double Holliday junction (HJ) dissolution. In the absence of Rqh1, this intermediate will be resolved by nuclease(s), which could generate dicentric circular chromosome dimer by crossing over. Even if spontaneous DSB in a circular chromosome is not repaired by HR due to the defect in HR activity, this DSB would not result in cell death, possibly due to adaptation or to NHEJ dependent DSB repair.


{kind=link}
{kind=link}
Table 1.
Summary of pot1′s genetic interactions.
| Gene Name (Related Functions) | Phenotype (References) |
|---|---|
| Lethal genetic interaction with pot1 | |
| lig4, rad16, rad22, srs2 (Single strand annealing (SSA)) | No chromosome fusion by deletion of SSA genes [30] |
| rqh1, top3 (dissolution of double holiday junction) | Circular chromosome is not maintained by deletion of rqh1 or mutation in top3 [33] |
| ark1, bir1, pic1 (CPC) (chromosome segregation) | Circular chromosome is not maintained by mutation of CPC [54] |
| bdf2, mst1 (maintaining proper histone H4 acetylation level) | Circular chromosome causes growth defect by bdf2 or mst1 mutation [61] |
| pik1 (regulation of PI(4)P level in Golgi and possibly in nucleus?) | Circular chromosome causes growth defect by pik1 mutation? [67] |
| Suppresion of pot1, pot1 rqh1 double and pot1 rqh1-hd double mutant phenotype | |
| rqh1 (Helicase) | rqh1-hd mutation rescues telomere loss of pot1Δ cells [45] |
| chk1, wee1 mik1 double mutation, cdc2-3w (G2 length regulation) | chk1 or cdc2-3w mutation, or wee1 mik1 double mutation rescue TBZ sensitivity of pot1 rqh1-hd cells [47,48] |
| rad51, exo1 rqh1 double mutation (HR) | Lethality of pot1 rqh1 is suppressed by deletion of rad51 or exo1 [33] |
| exo1, rqh1 (processing of uncapped telomere) | Acute growth defect by pot1 shut-off is suppressed by exo1 and/or rqh1 mutation [32] |
| Enhancement of pot1 and pot1 rqh1 double mutant phenotype | |
| bub1, mad2 (Spindle checkpoint) | Growth of pot1 rqh1-hd cells becomes worse by bub1 or mad2 mutation [47] |
| hus1, rad1, rad9 (9-1-1 complex) (DNA damage checkpoint) | Mutation in 9-1-1 complex increases HU sensitivity of pot1Δ cells [76] |
| swi6 (Heterochromatin), sgo2 (Sister chromatid biorientation) | Mutation of swi6 or sgo2 increases chromosome segregation defect of pot1Δ cells [54] |
| Phenotype suppression by deletion of pot1 | |
| dcr1 (RNA interference machinery) | Pericentric silencing defect in dcr1Δ cells is suppressed by pot1 deletion [58] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ueno, M. Exploring Genetic Interactions with Telomere Protection Gene pot1 in Fission Yeast. Biomolecules 2023, 13, 370. https://doi.org/10.3390/biom13020370
AMA Style
Ueno M. Exploring Genetic Interactions with Telomere Protection Gene pot1 in Fission Yeast. Biomolecules. 2023; 13(2):370. https://doi.org/10.3390/biom13020370
Chicago/Turabian StyleUeno, Masaru. 2023. "Exploring Genetic Interactions with Telomere Protection Gene pot1 in Fission Yeast" Biomolecules 13, no. 2: 370. https://doi.org/10.3390/biom13020370
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.