
Imipramine Treatment Alters Sphingomyelin, Cholesterol, and Glycerophospholipid Metabolism in Isolated Macrophage Lysosomes
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Lysosome Isolation
2.3. Western Blot
2.4. LysoTracker Red and NAG Enzyme Activity
2.5. Cryo-Electron Microscopy
2.6. Lipid Extraction
2.7. Time-Resolved Fluorescence Anisotropy Measurements
2.8. Data Acquisition and Statistical Analysis
2.8.1. LysoTracker Red, NAG Enzyme Activity, and Time-Resolved Fluorescence Anisotropy Data
2.8.2. Lipid Profiling
3. Results
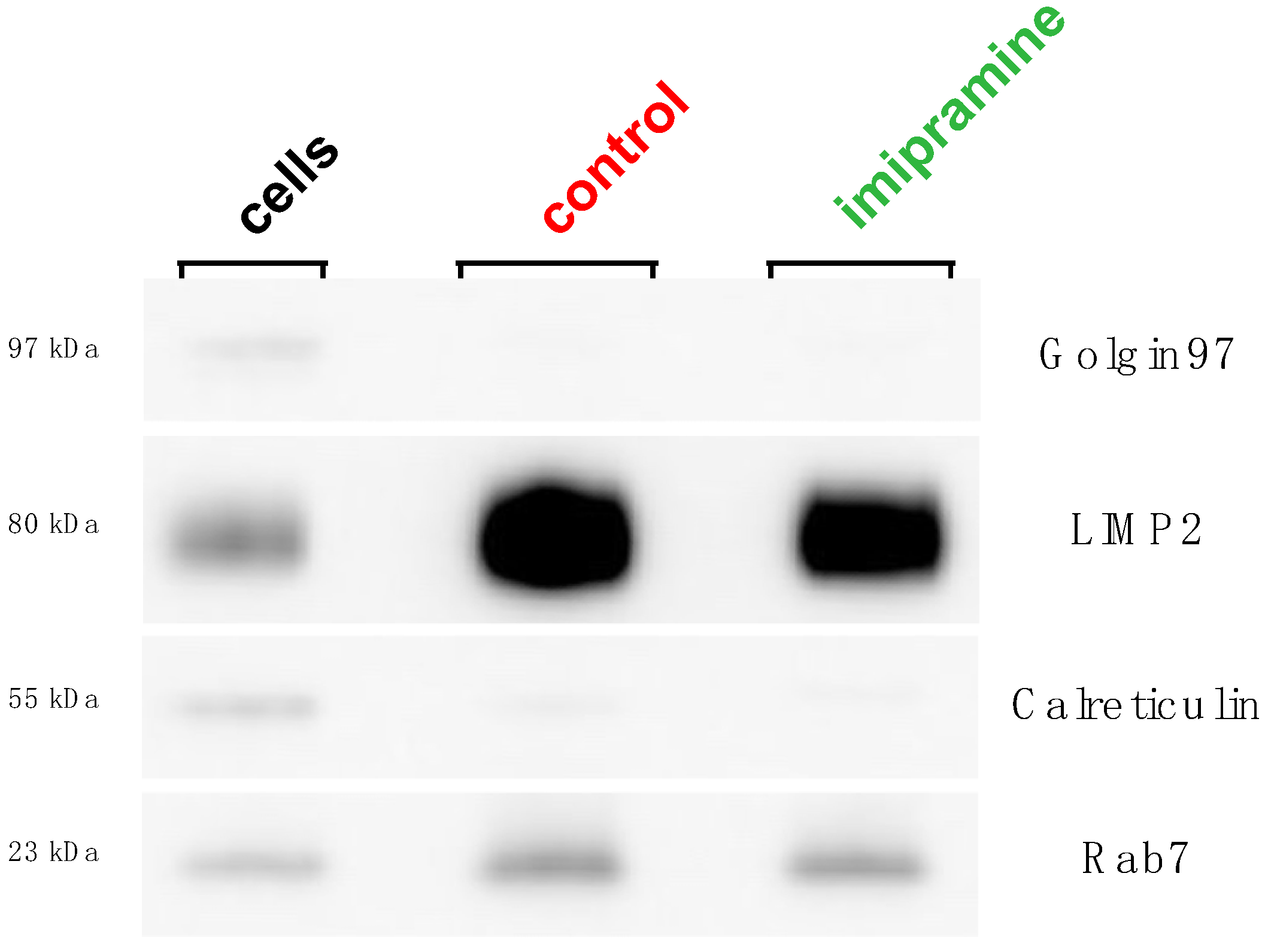
3.1. Validation of Lysosome Purity
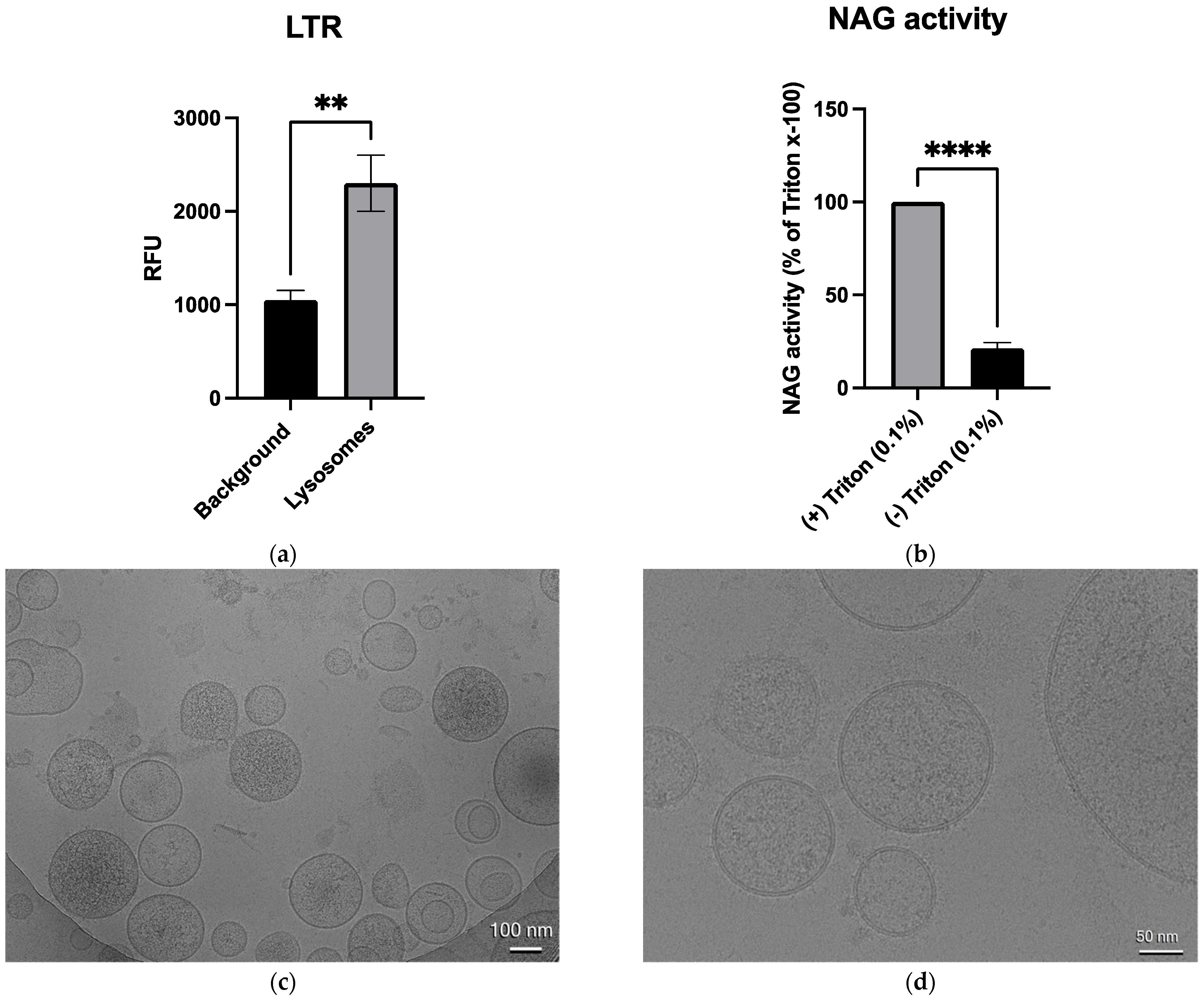
3.2. Validation of Lysosome Integrity, Functionality, and Morphology
3.3. Imipramine Treatment Altered Sphingomyelin, Cholesterol, and Glycerophospholipid Metabolism in Macrophage Lysosomes
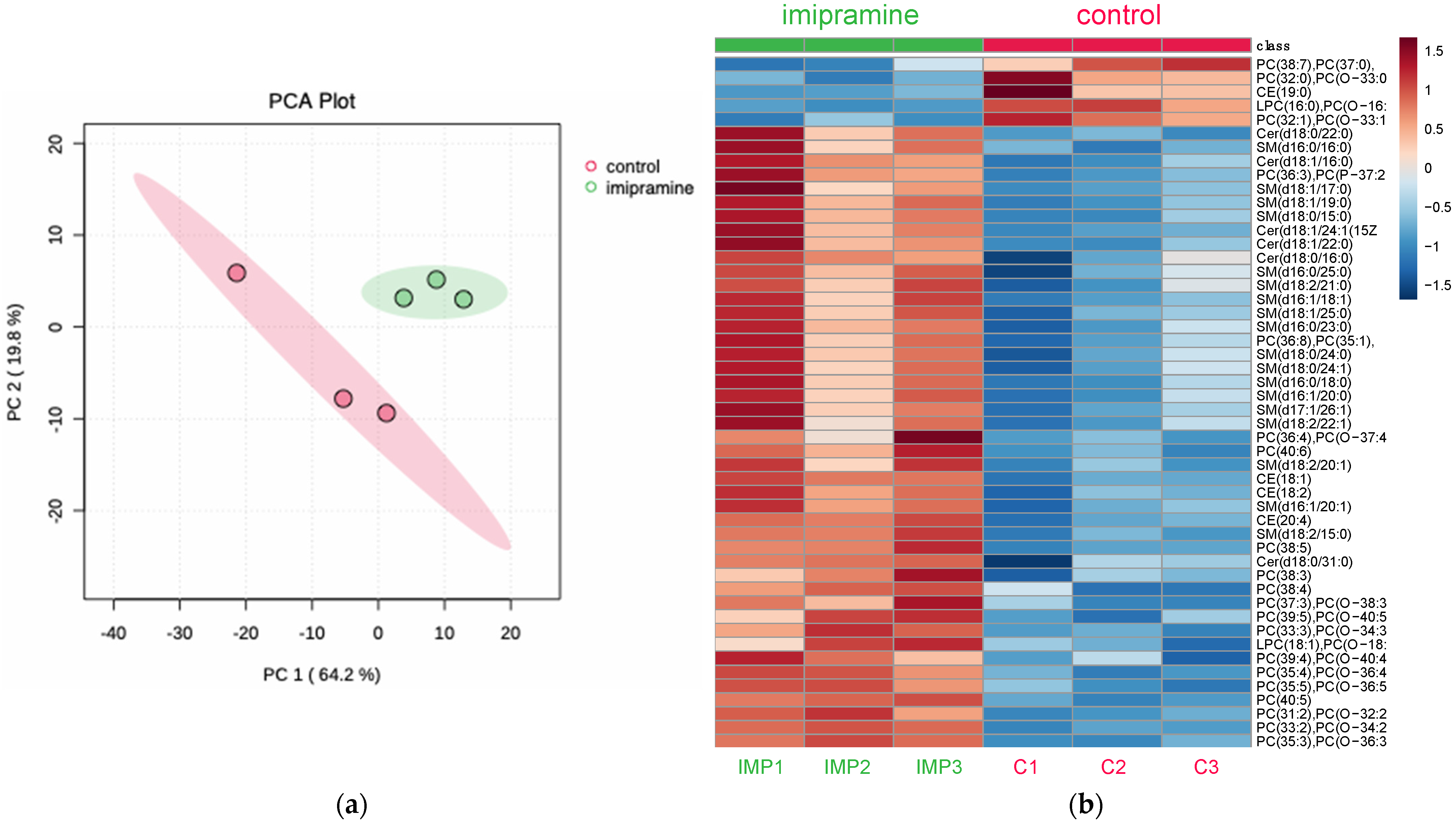
3.3.1. Principal Component Analysis and Heatmap
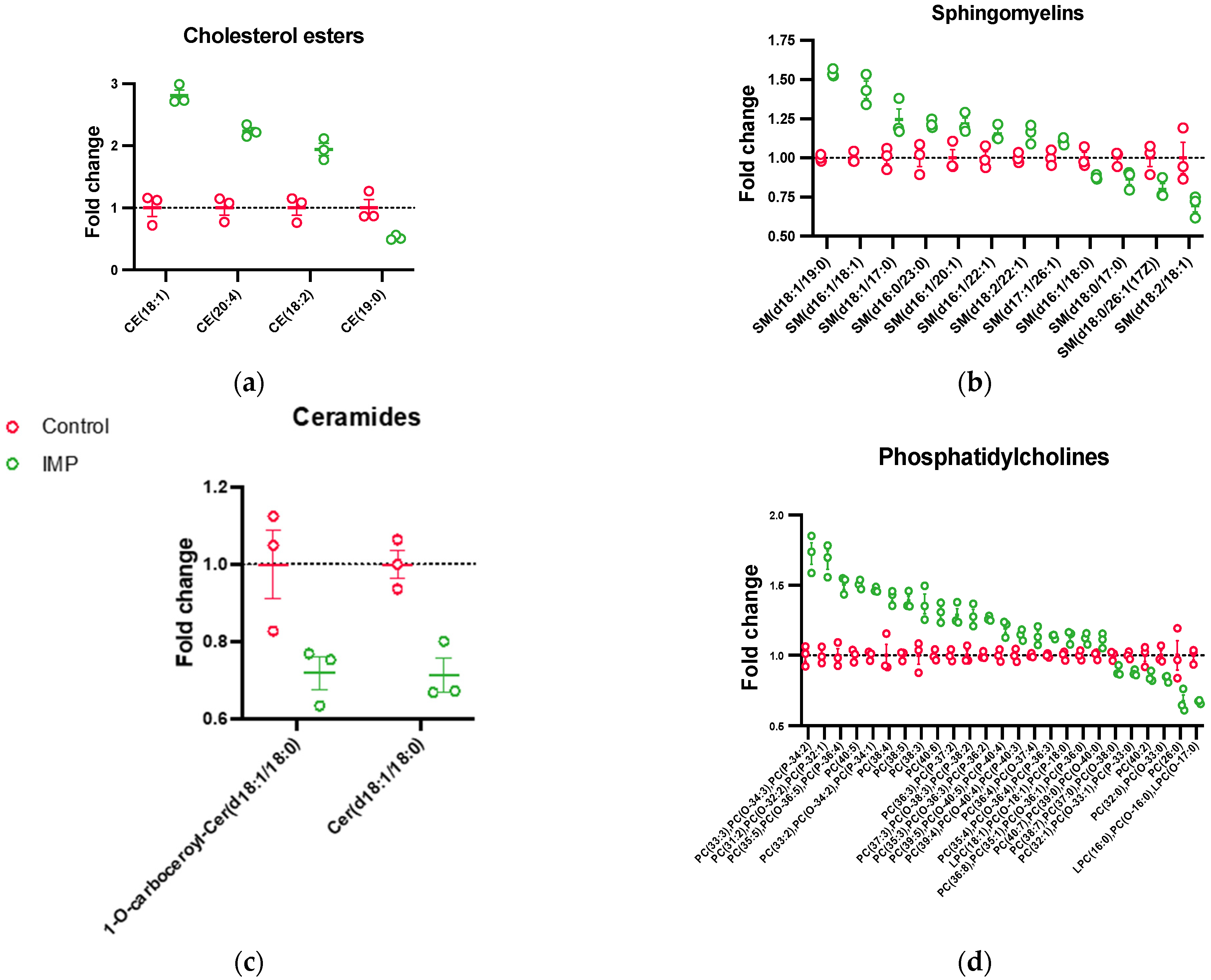
3.3.2. Lipid Species Fold-Changes
3.4. Cholesterol Accumulation Induced Biophysical Changes in Isolated Macrophage Lysosomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nixon, R.A.; Yang, D.S. Autophagy failure in Alzheimer’s disease–locating the primary defect. Neurobiol. Dis. 2011, 43, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Duewell, P.; Kono, H.; Rayner, K.J.; Sirois, C.M.; Vladimer, G.; Bauernfeind, F.G.; Abela, G.S.; Franchi, L.; Nuñez, G.; Schnurr, M.; et al. NLRP3 inflammasomes are required for atherogenesis and activated by cholesterol crystals. Nature 2010, 464, 1357–1361. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Chen, W.Y.; Yu, C.L.; Tsai, C.Y.; Hsieh, S.C. Gouty arthritis involves impairment of autophagic degradation via cathepsin D inactivation-mediated lysosomal dysfunction that promotes apoptosis in macrophages. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166703. [Google Scholar] [CrossRef] [PubMed]
- Joshi, G.N.; Knecht, D.A. Silica phagocytosis causes apoptosis and necrosis by different temporal and molecular pathways in alveolar macrophages. Apoptosis 2013, 18, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yue, P.; Lu, T.; Wang, Y.; Wei, Y.; Wei, X. Role of lysosomes in physiological activities, diseases, and therapy. J. Hematol. Oncol. 2021, 14, 79. [Google Scholar] [CrossRef] [PubMed]
- Gros, F.; Muller, S. The role of lysosomes in metabolic and autoimmune diseases. Nat. Rev. Nephrol. 2023, 19, 366–383. [Google Scholar] [CrossRef]
- Lübke, T.; Lobel, P.; Sleat, D.E. Proteomics of the lysosome. Biochim. Biophys. Acta 2009, 1793, 625–635. [Google Scholar] [CrossRef]
- Schröder, B.A.; Wrocklage, C.; Hasilik, A.; Saftig, P. The proteome of lysosomes. Proteomics 2010, 10, 4053–4076. [Google Scholar] [CrossRef]
- Sevin, C.; Deiva, K. Clinical Trials for Gene Therapy in Lysosomal Diseases With CNS Involvement. Front. Mol. Biosci. 2021, 8, 624988. [Google Scholar] [CrossRef]
- Nixon, G.F. Sphingolipids in inflammation: Pathological implications and potential therapeutic targets. Br. J. Pharmacol. 2009, 158, 982–993. [Google Scholar] [CrossRef]
- Norris, G.H.; Porter, C.M.; Jiang, C.; Blesso, C.N. Dietary Milk Sphingomyelin Reduces Systemic Inflammation in Diet-Induced Obese Mice and Inhibits LPS Activity in Macrophages. Beverages 2017, 3, 37. [Google Scholar] [CrossRef]
- Wu, D.; Marko, M.; Claycombe, K.; Paulson, K.E.; Meydani, S.N. Ceramide-induced and age-associated increase in macrophage COX-2 expression is mediated through up-regulation of NF-kappa B activity. J. Biol. Chem. 2003, 278, 10983–10992. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, Z.; Zhang, L.; Kirkwood, C.L.; Kirkwood, K.L.; Lopes-Virella, M.F.; Huang, Y. Inhibition of acid sphingomyelinase by imipramine abolishes the synergy between metabolic syndrome and periodontitis on alveolar bone loss. J. Periodontal Res. 2022, 57, 173–185. [Google Scholar] [CrossRef]
- Hillhouse, T.M.; Porter, J.H. A brief history of the development of antidepressant drugs: From monoamines to glutamate. Exp. Clin. Psychopharmacol. 2015, 23, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Funk, R.S.; Krise, J.P. Cationic amphiphilic drugs cause a marked expansion of apparent lysosomal volume: Implications for an intracellular distribution-based drug interaction. Mol. Pharm. 2012, 9, 1384–1395. [Google Scholar] [CrossRef]
- Kendall, R.L.; Holian, A. Cholesterol-dependent molecular mechanisms contribute to cationic amphiphilic drugs’ prevention of silica-induced inflammation. Eur. J. Cell Biol. 2023, 102, 151310. [Google Scholar] [CrossRef]
- Appelqvist, H.; Nilsson, C.; Garner, B.; Brown, A.J.; Kågedal, K.; Öllinger, K. Attenuation of the Lysosomal Death Pathway by Lysosomal Cholesterol Accumulation. Am. J. Pathol. 2011, 178, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Sydor, M.J.; Kendall, R.L.; Holian, A. Cholesterol content regulates silica-induced lysosomal membrane permeability. Front. Toxicol. 2023, 5, 1112822. [Google Scholar] [CrossRef]
- Raffy, S.; Teissié, J. Control of lipid membrane stability by cholesterol content. Biophys. J. 1999, 76, 2072–2080. [Google Scholar] [CrossRef]
- Biswas, R.; Trout, K.L.; Jessop, F.; Harkema, J.R.; Holian, A. Imipramine blocks acute silicosis in a mouse model. Part. Fibre Toxicol. 2017, 14, 36. [Google Scholar] [CrossRef]
- Gorki, A.D.; Symmank, D.; Zahalka, S.; Lakovits, K.; Hladik, A.; Langer, B.; Maurer, B.; Sexl, V.; Kain, R.; Knapp, S. Murine Ex Vivo Cultured Alveolar Macrophages Provide a Novel Tool to Study Tissue-Resident Macrophage Behavior and Function. Am. J. Respir. Cell Mol. Biol. 2022, 66, 64–75. [Google Scholar] [CrossRef]
- Kendall, R.L.; Ray, J.L.; Hamilton, R.F., Jr.; Holian, A. Self-replicating murine ex vivo cultured alveolar macrophages as a model for toxicological studies of particle-induced inflammation. Toxicol. Appl. Pharmacol. 2023, 461, 116400. [Google Scholar] [CrossRef] [PubMed]
- Korohoda, W.; Oczkowicz, J.; Rolski, K.; Ryszawy, D.; Madeja, Z. Efficacy of Local Anesthetics in Detachment of Normal 3T3 Mouse Fibroblasts and Prostate Cancer AT-2 Cells from Substrata, in Maintenance of Viable Cells in a Non-Adherent State, and in Preservation of Cell Surface Markers Detected with FlowSight Image Cytometry. Folia Biol. 2015, 63, 249–255. [Google Scholar] [CrossRef]
- Aits, S.; Kricker, J.; Liu, B.; Ellegaard, A.M.; Hämälistö, S.; Tvingsholm, S.; Corcelle-Termeau, E.; Høgh, S.; Farkas, T.; Holm Jonassen, A.; et al. Sensitive detection of lysosomal membrane permeabilization by lysosomal galectin puncta assay. Autophagy 2015, 11, 1408–1424. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Minazzo, A.S.; Darlington, R.C.; Ross, J.B. Loop dynamics of the extracellular domain of human tissue factor and activation of factor VIIa. Biophys. J. 2009, 96, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Kinosita, K., Jr.; Ikegami, A.; Kawato, S. On the wobbling-in-cone analysis of fluorescence anisotropy decay. Biophys. J. 1982, 37, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Reis, L.G.; Casey, T.M.; Sobreira, T.J.P.; Cooper, B.R.; Ferreira, C.R. Step-by-Step Approach to Build Multiple Reaction Monitoring (MRM) Profiling Instrument Acquisition Methods for Class-based Lipid Exploratory Analysis by Mass Spectrometry. J. Biomol. Tech. 2023, 34. [Google Scholar] [CrossRef]
- Xie, Z.; Ferreira, C.R.; Virequ, A.A.; Cooks, R.G. Multiple reaction monitoring profiling (MRM profiling): Small molecule exploratory analysis guided by chemical functionality. Chem. Phys. Lipids 2021, 235, 105048. [Google Scholar] [CrossRef]
- Alqahtani, S.; Kobos, L.M.; Xia, L.; Ferreira, C.; Franco, J.; Du, X.; Shannahan, J.H. Exacerbation of Nanoparticle-Induced Acute Pulmonary Inflammation in a Mouse Model of Metabolic Syndrome. Front. Immunol. 2020, 11, 818. [Google Scholar] [CrossRef]
- Ferreira, C.R.; Yannell, K.E.; Mollenhauer, B.; Espy, R.D.; Cordeiro, F.B.; Ouyang, Z.; Cooks, R. Chemical profiling of cerebrospinal fluid by multiple reaction monitoring mass spectrometry. Analyst 2016, 141, 5252–5255. [Google Scholar] [CrossRef] [PubMed]
- Kobos, L.; Ferreira, C.R.; Sobreira, T.J.P.; Rajwa, B.; Shannahan, J. A novel experimental workflow to determine the impact of storage parameters on the mass spectrometric profiling and assessment of representative phosphatidylethanolamine lipids in mouse tissues. Anal. Bioanal. Chem. 2021, 413, 1837–1849. [Google Scholar] [CrossRef] [PubMed]
- Kobos, L.M.; Alqatani, S.; Ferreira, C.R.; Aryal, U.K.; Hedrick, V.; Sobreira, T.J.P.; Shannahan, J.H. An Integrative Proteomic/Lipidomic Analysis of the Gold Nanoparticle Biocorona in Healthy and Obese Conditions. Appl. In Vitro Toxicol. 2019, 5, 150–166. [Google Scholar] [CrossRef] [PubMed]
- Yannell, K.E.; Ferreira, C.R.; Tichy, S.E.; Cooks, R.G. Multiple reaction monitoring (MRM)-profiling with biomarker identification by LC-QTOF to characterize coronary artery disease. Analyst 2018, 143, 5014–5022. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, A.S.; Rajan, A.; Ferreira, C.R.; Ellis, J.M. Loss of Muscle Carnitine Palmitoyltransferase 2 Prevents Diet-Induced Obesity and Insulin Resistance despite Long-Chain Acylcarnitine Accumulation. Cell Rep. 2020, 33, 108374. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.J.; Hou, J.F.; Jiang, N.; Zhang, G.J. Loss of membrane cholesterol affects lysosomal osmotic stability. Gen. Physiol. Biophys. 2008, 27, 278–283. [Google Scholar] [PubMed]
- Eriksson, I.; Wäster, P.; Öllinger, K. Restoration of lysosomal function after damage is accompanied by recycling of lysosomal membrane proteins. Cell Death Dis. 2020, 11, 370. [Google Scholar] [CrossRef]
- Samsonov, A.V.; Mihalyov, I.; Cohen, F.S. Characterization of cholesterol-sphingomyelin domains and their dynamics in bilayer membranes. Biophys. J. 2001, 81, 1486–1500. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Kennelly, J.P.; Wan, S.; Vance, J.E.; Vance, D.E.; Jacobs, R.L. The critical role of phosphatidylcholine and phosphatidylethanolamine metabolism in health and disease. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1558–1572. [Google Scholar] [CrossRef]
- Thomas, J.M.; Hullin, F.; Chap, H.; Douste-Blazy, L. Phosphatidylcholine is the major phospholipid providing arachidonic acid for prostacyclin synthesis in thrombin-stimulated human endothelial cells. Thromb. Res. 1984, 34, 117–123. [Google Scholar] [CrossRef]
- Law, S.H.; Chan, M.L.; Marathe, G.K.; Parveen, F.; Chen, C.H.; Ke, L.Y. An Updated Review of Lysophosphatidylcholine Metabolism in Human Diseases. Int. J. Mol. Sci. 2019, 20, 1149. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.; Qiu, C.; Zhao, L. Lysophosphatidylcholine perpetuates macrophage polarization toward classically activated phenotype in inflammation. Cell. Immunol. 2014, 289, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Kilin, V.; Glushonkov, O.; Herdly, L.; Klymchenko, A.; Richert, L.; Mely, Y. Fluorescence lifetime imaging of membrane lipid order with a ratiometric fluorescent probe. Biophys. J. 2015, 108, 2521–2531. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Millard, A.C.; Wuskell, J.P.; Clark, H.A.; Loew, L.M. Cholesterol-Enriched Lipid Domains Can Be Visualized by di-4-ANEPPDHQ with Linear and Nonlinear Optics. Biophys. J. 2005, 89, L04–L06. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Liang, Q.; Abi-Mosleh, L.; Das, A.; De Brabander, J.K.; Goldstein, J.L.; Brown, M.S. Identification of NPC1 as the target of U18666A, an inhibitor of lysosomal cholesterol export and Ebola infection. eLife 2015, 4, e12177. [Google Scholar] [CrossRef] [PubMed]
- Lorey, M.B.; Öörni, K.; Kovanen, P.T. Modified Lipoproteins Induce Arterial Wall Inflammation During Atherogenesis. Front. Cardiovasc. Med. 2022, 9, 841545. [Google Scholar] [CrossRef] [PubMed]
- Cockburn, C.L.; Green, R.S.; Damle, S.R.; Martin, R.K.; Ghahrai, N.N.; Colonne, P.M.; Fullerton, M.S.; Conrad, D.H.; Chalfant, C.E.; Voth, D.E.; et al. Functional inhibition of acid sphingomyelinase disrupts infection by intracellular bacterial pathogens. Life Sci. Alliance 2019, 2, e201800292. [Google Scholar] [CrossRef]
- Jerome, W.G.; Cox, B.E.; Griffin, E.E.; Ullery, J.C. Lysosomal cholesterol accumulation inhibits subsequent hydrolysis of lipoprotein cholesteryl ester. Microsc. Microanal. 2008, 14, 138–149. [Google Scholar] [CrossRef]
- Tharkeshwar, A.K.; Trekker, J.; Vermeire, W.; Pauwels, J.; Sannerud, R.; Priestman, D.A.; Te Vruchte, D.; Vints, K.; Baatsen, P.; Decuypere, J.P.; et al. A novel approach to analyze lysosomal dysfunctions through subcellular proteomics and lipidomics: The case of NPC1 deficiency. Sci. Rep. 2017, 7, 41408. [Google Scholar] [CrossRef]
- Deffieu, M.S.; Pfeffer, S.R. Niemann-Pick type C 1 function requires lumenal domain residues that mediate cholesterol-dependent NPC2 binding. Proc. Natl. Acad. Sci. USA 2011, 108, 18932–18936. [Google Scholar] [CrossRef]
- Oninla, V.O.; Breiden, B.; Babalola, J.O.; Sandhoff, K. Acid sphingomyelinase activity is regulated by membrane lipids and facilitates cholesterol transfer by NPC2. J. Lipid Res. 2014, 55, 2606–2619. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, H. Lysosomal Acid Lipase in Lipid Metabolism and Beyond. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 850–856. [Google Scholar] [CrossRef]
- Abe, A.; Hiraoka, M.; Wild, S.; Wilcoxen, S.E.; Paine, R., 3rd; Shayman, J.A. Lysosomal phospholipase A2 is selectively expressed in alveolar macrophages. J. Biol. Chem. 2004, 279, 42605–42611. [Google Scholar] [CrossRef] [PubMed]
- Bartolf, M.; Franson, R.C. pH-dependent modulation of phospholipase A2 activity by alkaline cations and catecholamines in a granule-enriched fraction of adrenal medulla. Biochim. Biophys. Acta 1984, 793, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Abe, A.; Shayman, J.A. The role of negatively charged lipids in lysosomal phospholipase A2 function. J. Lipid Res. 2009, 50, 2027–2035. [Google Scholar] [CrossRef] [PubMed]
- Steele, H.B.B.; Sydor, M.J.; Anderson, D.S.; Holian, A.; Ross, J.B.A. Using Time-Resolved Fluorescence Anisotropy of di-4-ANEPPDHQ and F2N12S to Analyze Lipid Packing Dynamics in Model Systems. J. Fluoresc. 2019, 29, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Sydor, M.J.; Anderson, D.S.; Steele, H.B.B.; Ross, J.B.A.; Holian, A. Effects of titanium dioxide and zinc oxide nano-materials on lipid order in model membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2020, 1862, 183313. [Google Scholar] [CrossRef]
- Rog, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. Ordering effects of cholesterol and its analogues. Biochim. Biophys. Acta 2009, 1788, 97–121. [Google Scholar] [CrossRef]
- Urbina, J.A.; Pekerar, S.; Le, H.-b.; Patterson, J.; Montez, B.; Oldfield, E. Molecular order and dynamics of phosphatidylcholine bilayer membranes in the presence of cholesterol, ergosterol and lanosterol: A comparative study using 2H-, 13C- and 31P-NMR spectroscopy. Biochim. Biophys. Acta (BBA)-Biomembr. 1995, 1238, 163–176. [Google Scholar] [CrossRef]
- Boggs, J.M.; Hsia, J.C. Effect of cholesterol and water on the rigidity and order of phosphatidylcholine bilayers. Biochim. Biophys. Acta 1972, 290, 32–42. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albright, J.M.; Sydor, M.J.; Shannahan, J.; Ferreira, C.R.; Holian, A. Imipramine Treatment Alters Sphingomyelin, Cholesterol, and Glycerophospholipid Metabolism in Isolated Macrophage Lysosomes. Biomolecules 2023, 13, 1732. https://doi.org/10.3390/biom13121732
Albright JM, Sydor MJ, Shannahan J, Ferreira CR, Holian A. Imipramine Treatment Alters Sphingomyelin, Cholesterol, and Glycerophospholipid Metabolism in Isolated Macrophage Lysosomes. Biomolecules. 2023; 13(12):1732. https://doi.org/10.3390/biom13121732
Chicago/Turabian StyleAlbright, Jacob M., Matthew J. Sydor, Jonathan Shannahan, Christina R. Ferreira, and Andrij Holian. 2023. "Imipramine Treatment Alters Sphingomyelin, Cholesterol, and Glycerophospholipid Metabolism in Isolated Macrophage Lysosomes" Biomolecules 13, no. 12: 1732. https://doi.org/10.3390/biom13121732