
Metabolomic Profiling in Patients with Different Hemodynamic Subtypes of Severe Aortic Valve Stenosis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patient Cohort
2.2. Echocardiography
- Normal/preserved ejection fraction, high-gradient AS (NEF HGAS): LVEF ≥ 50%, vmax ≥ 4 m/s or Pmean ≥ 40 mmHg, and aortic valve area (AVA) ≤ 1.0 cm2;
- Low/reduced ejection fraction, high-gradient AS (LEF HGAS): LVEF < 50%, vmax ≥ 4 m/s, or Pmean ≥ 40 mmHg, and AVA ≤ 1.0 cm2;
- Low/reduced ejection fraction, low-gradient AS (‘classic’low-flow, low-gradient AS) (LEF LGAS): LVEF < 50%, vmax < 4 m/s and Pmean < 40 mmHg, AVA ≤ 1.0 cm2, and stroke volume index (SVI) ≤ 35 mL/m2;
- Normal/preserved ejection fraction, low-gradient AS (’paradoxical’low-flow, low-gradient AS) (NEF LGAS): LVEF ≥ 50%, vmax < 4 m/s and Pmean < 40 mmHg, AVA ≤ 1.0 cm2 and indexed AVA ≤ 0.6 cm2/m2, and SVI ≤ 35 mL/m2. For this study, serum from 10 patients per subgroup plus 10 healthy controls were analyzed (Table 1).
2.3. Serum Sample Preparation
2.4. Metabolomic Profiling
2.5. Statistical Analysis
3. Results
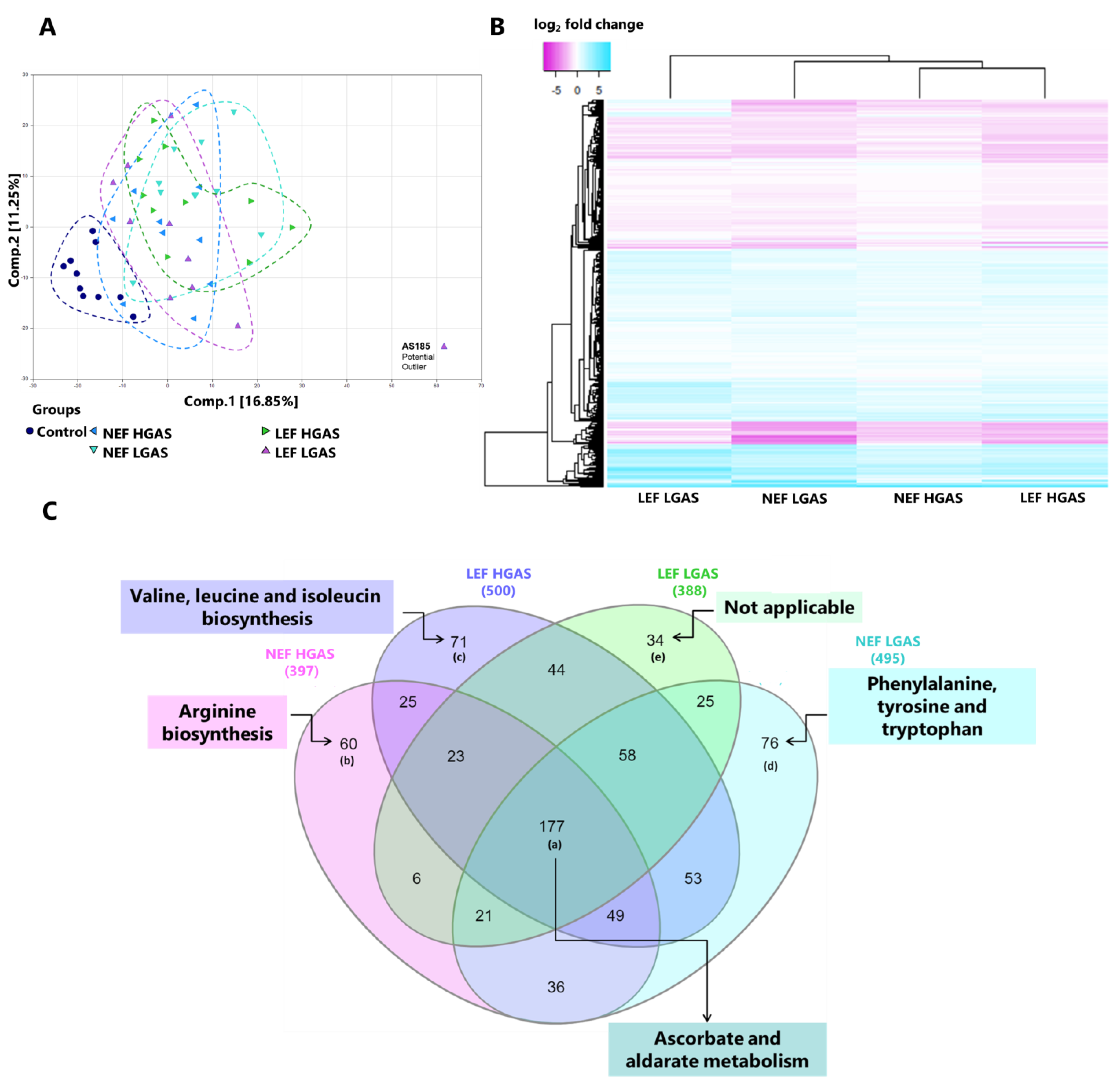
3.1. Exploration with Principal Component Analysis and Hierarchical Clustering
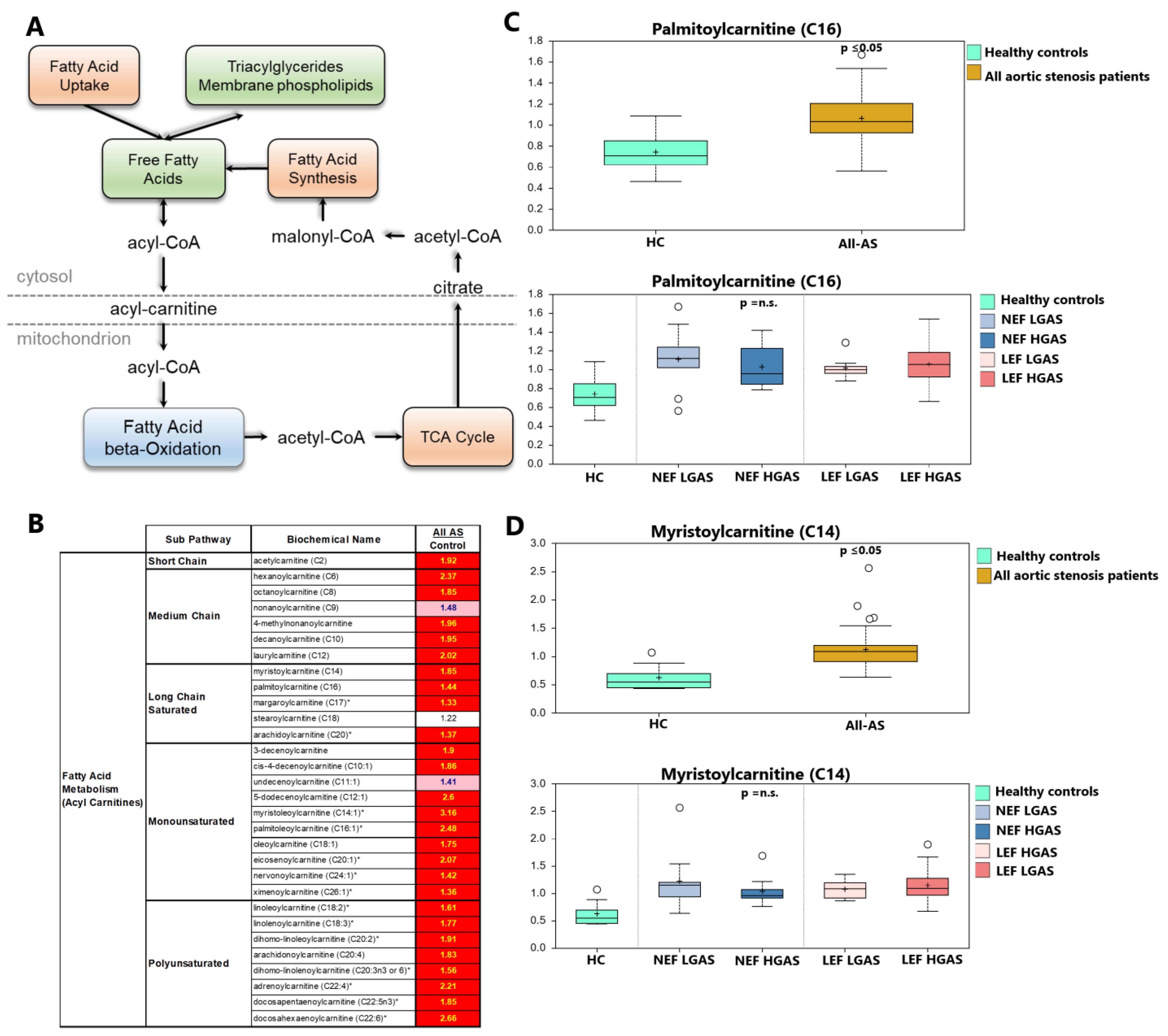
3.2. Alterations in Acylcarnitine Levels
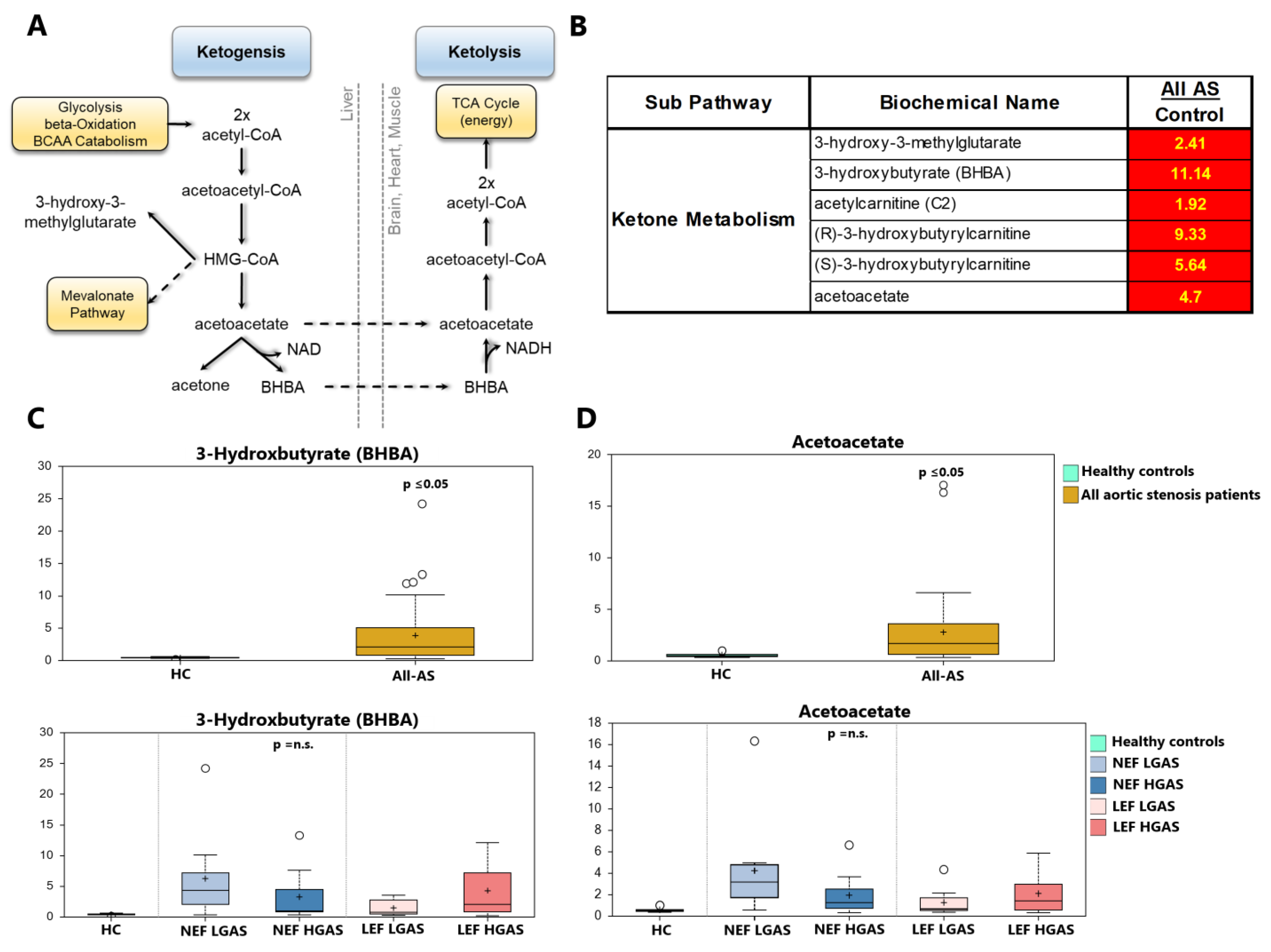
3.3. Alterations in Ketone Body Metabolism
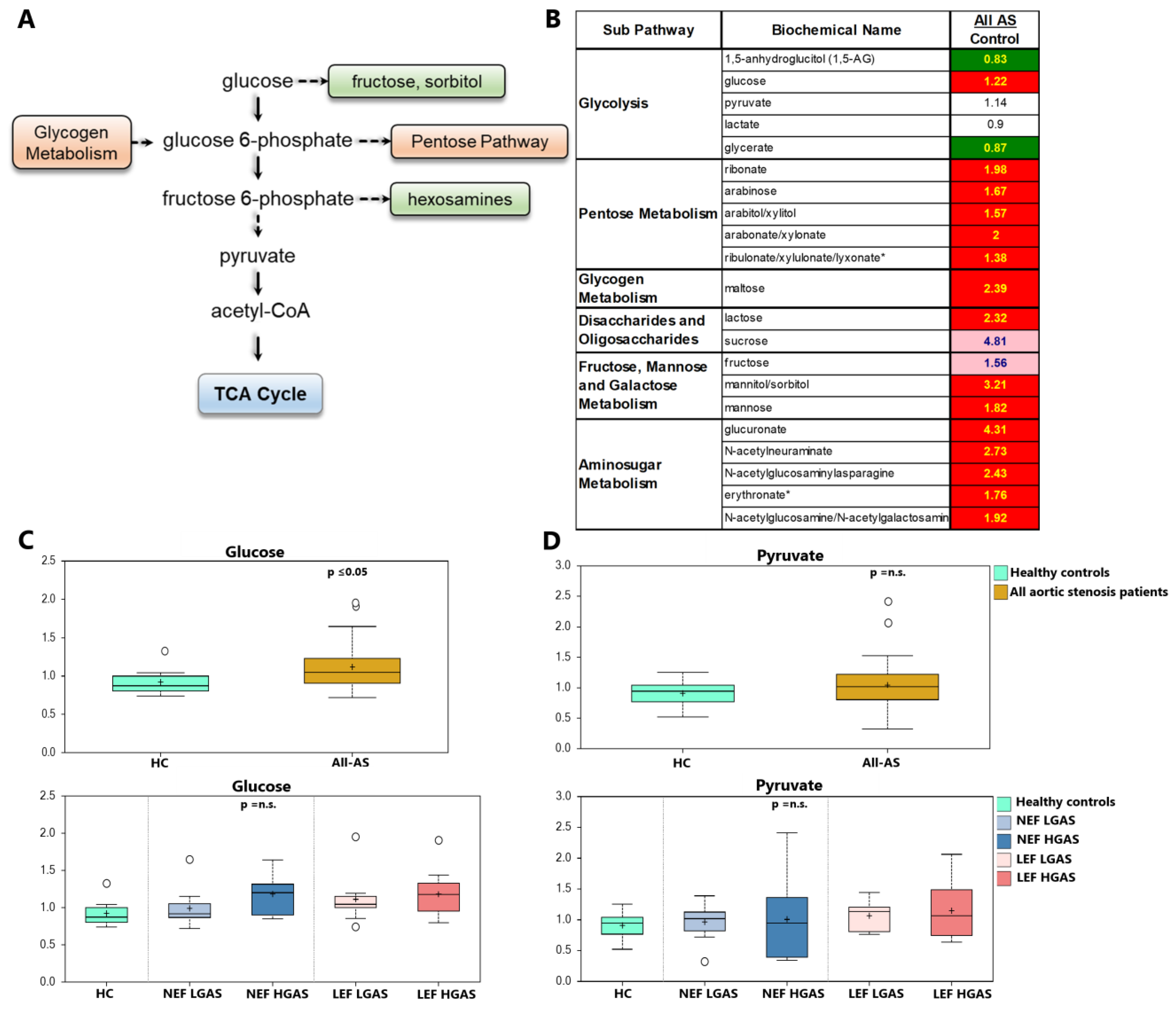
3.4. Alterations in Sugar Metabolism
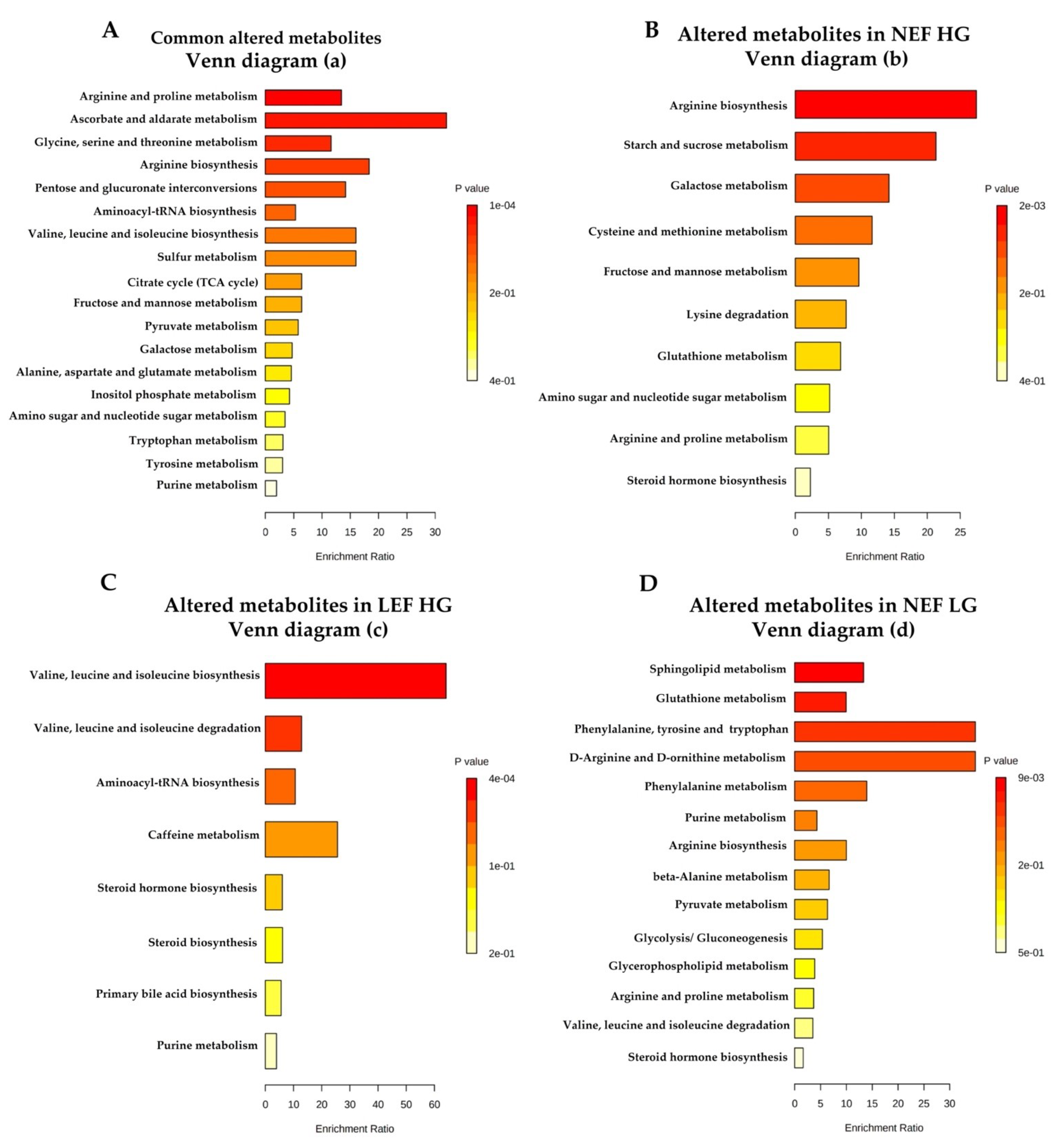
3.5. Differences in Metabolite Levels between AS Subgroups
4. Discussion
4.1. Acylcarnitines
4.2. Ketone Metabolism
4.3. Sugar Metabolism
4.4. Amino Acid Metabolism and Nitrogen Balance
4.5. Observed Biochemical Changes (AS Subtyping)
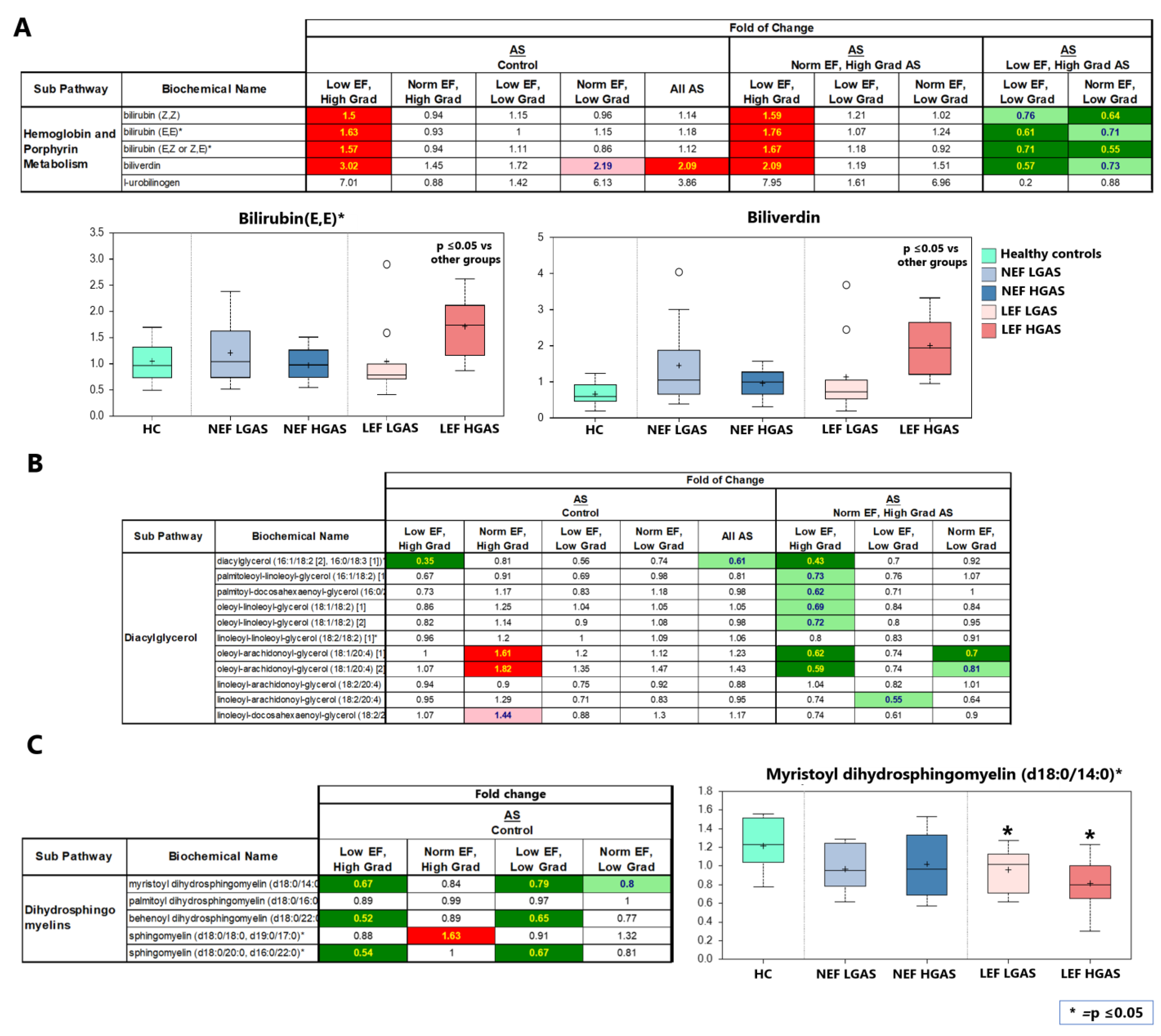
4.5.1. Hemoglobin Metabolism
4.5.2. Diacylglycerols (DAGs)
4.5.3. Dihydrosphingomyelins (DHSMs)
4.6. Study limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baumgartner, H.; Falk, V.; Bax, J.J.; De Bonis, M.; Hamm, C.; Holm, P.J.; Iung, B.; Lancellotti, P.; Lansac, E.; Muñoz, D.R.; et al. 2017 ESC/EACTS guidelines for the management of valvular heart disease. Eur. Heart J. 2017, 38, 2739–2786. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.R.; Leon, M.B.; Mack, M.J.; Miller, D.C.; Moses, J.W.; Svensson, L.G.; Tuzcu, E.M.; Webb, J.G.; Fontana, G.P.; Makkar, R.R.; et al. Transcatheter versus surgical aortic-valve replacement in high-risk patients. N. Engl. J. Med. 2011, 364, 2187–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon, M.B.; Smith, C.R.; Mack, M.J.; Makkar, R.R.; Svensson, L.G.; Kodali, S.K.; Thourani, V.H.; Tuzcu, E.M.; Miller, D.C.; Herrmann, H.C.; et al. Transcatheter or surgical aortic-valve replacement in intermediate-risk patients. N. Engl. J. Med. 2016, 374, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Mack, M.J.; Leon, M.B.; Thourani, V.H.; Makkar, R.; Kodali, S.K.; Russo, M.; Kapadia, S.R.; Chris Malaisrie, S.; Cohen, D.J.; Pibarot, P.; et al. Transcatheter aortic-valve replacement with a balloon-expandable valve in low-risk patients. N. Engl. J. Med. 2019, 380, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Makkar, R.R.; Fontana, G.P.; Jilaihawi, H.; Kapadia, S.; Pichard, A.D.; Douglas, P.S.; Thourani, V.H.; Babaliaros, V.C.; Webb, J.G.; Herrmann, H.C.; et al. Transcatheter aortic-valve replacement for inoperable severe aortic stenosis. N. Engl. J. Med. 2012, 366, 1696–1704. [Google Scholar] [CrossRef] [Green Version]
- Leon, M.B.; Smith, C.R.; Mack, M.; Miller, D.C.; Moses, J.W.; Svensson, L.G.; Tuzcu, E.M.; Webb, J.G.; Fontana, G.P.; Makkar, R.R.; et al. Transcatheter aortic-valve implantation for aortic stenosis in patients who cannot undergo surgery. N. Engl. J. Med. 2010, 363, 1597–1607. [Google Scholar] [CrossRef] [Green Version]
- Mangner, N.; Stachel, G.; Woitek, F.; Haussig, S.; Schlotter, F.; Höllriegel, R.; Adam, J.; Lindner, A.; Mohr, F.W.; Schuler, G.; et al. Predictors of mortality and symptomatic outcome of patients with low-flow severe aortic stenosis undergoing transcatheter aortic valve replacement. J. Am. Heart Assoc. 2018, 7, e007977. [Google Scholar] [CrossRef] [Green Version]
- Ahn, Y.; Choi, S.J.; Lim, S.; Kim, J.B.; Song, J.M.; Kang, D.H.; Song, J.K.; Kim, H.J.; Kang, J.W.; Yang, D.H.; et al. Classification of severe aortic stenosis and outcomes after aortic valve replacement. Sci. Rep. 2022, 12, 7506. [Google Scholar] [CrossRef]
- Snir, A.D.; Ng, M.K.; Strange, G.; Playford, D.; Stewart, S.; Celermajer, D.S. Prevalence and outcomes of low-gradient severe aortic stenosis—From the national echo database of Australia. J. Am. Heart Assoc. 2021, 10, 21126. [Google Scholar] [CrossRef]
- Bertero, E.; Maack, C. Metabolic remodelling in heart failure. Nat. Rev. Cardiol. 2018, 15, 457–470. [Google Scholar] [CrossRef]
- Gibb, A.A.; Hill, B.G. Metabolic coordination of physiological and pathological cardiac remodeling. Circ. Res. 2018, 123, 107–128. [Google Scholar] [CrossRef] [PubMed]
- Dávila-Román, V.G.; Vedala, G.; Herrero, P.; De Las Fuentes, L.; Rogers, J.G.; Kelly, D.P.; Gropler, R.J. Altered myocardial fatty acid and glucose metabolism in idiopathic dilated cardiomyopathy. J. Am. Coll. Cardiol. 2002, 40, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neglia, D.; De Caterina, A.; Marraccini, P.; Natali, A.; Ciardetti, M.; Vecoli, C.; Gastaldelli, A.; Ciociaro, D.; Pellegrini, P.; Testa, R.; et al. Impaired myocardial metabolic reserve and substrate selection flexibility during stress in patients with idiopathic dilated cardiomyopathy. Am. J. Physiol.—Heart Circ. Physiol. 2007, 293, 3270–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neubauer, S. The failing heart—An engine out of fuel. N. Engl. J. Med. 2007, 356, 1140–1151. [Google Scholar] [CrossRef] [Green Version]
- Heggermont, W.A.; Papageorgiou, A.P.; Heymans, S.; van Bilsen, M. Metabolic support for the heart: Complementary therapy for heart failure? Eur. J. Heart Fail. 2016, 18, 1420–1429. [Google Scholar] [CrossRef] [Green Version]
- Piquereau, J.; Boitard, S.E.; Ventura-Clapier, R.; Mericskay, M. Metabolic therapy of heart failure: Is there a future for B vitamins? Int. J. Mol. Sci. 2022, 23, 30. [Google Scholar] [CrossRef]
- McGarrah, R.W.; Crown, S.B.; Zhang, G.F.; Shah, S.H.; Newgard, C.B. Cardiovascular metabolomics. Circ. Res. 2018, 122, 1238–1258. [Google Scholar] [CrossRef]
- Puls, M.; Beuthner, B.E.; Topci, R.; Vogelgesang, A.; Bleckmann, A.; Sitte, M.; Lange, T.; Backhaus, S.J.; Schuster, A.; Seidler, T.; et al. Impact of myocardial fibrosis on left ventricular remodelling, recovery, and outcome after transcatheter aortic valve implantation in different haemodynamic subtypes of severe aortic stenosis. Eur. Heart J. 2020, 41, 1903–1914. [Google Scholar] [CrossRef] [Green Version]
- Lang, R.M.; Badano, L.P.; Victor, M.A.; Afilalo, J.; Armstrong, A.; Ernande, L.; Flachskampf, F.A.; Foster, E.; Goldstein, S.A.; Kuznetsova, T.; et al. Recommendations for cardiac chamber quantification by echocardiography in adults: An update from the American society of echocardiography and the European association of cardiovascular imaging. J. Am. Soc. Echocardiogr. 2015, 28, 1–39.e14. [Google Scholar] [CrossRef] [Green Version]
- Dehaven, C.D.; Evans, A.M.; Dai, H.; Lawton, K.A. Organization of GC/MS and LC/MS metabolomics data into chemical libraries. J. Cheminform. 2010, 2, 9. [Google Scholar] [CrossRef]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Heberle, H.; Meirelles, V.G.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Wishart, D.S. MSEA: A web-based tool to identify biologically meaningful patterns in quantitative metabolomic data. Nucleic Acids Res. 2010, 38, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toischer, K.; Rokita, A.G.; Unsöld, B.; Zhu, W.; Kararigas, G.; Sossalla, S.; Reuter, S.P.; Becker, A.; Teucher, N.; Seidler, T.; et al. Differential cardiac remodeling in preload versus afterload. Circulation 2010, 122, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Yarbrough, W.M.; Mukherjee, R.; Ikonomidis, J.S.; Zile, M.R.; Spinale, F.G. Myocardial remodeling with aortic stenosis and after aortic valve replacement: Mechanisms and future prognostic implications. J. Thorac. Cardiovasc. Surg. 2012, 143, 656–664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seymour, A.-M.L.; Giles, L.; Ball, V.; Miller, J.J.; Clarke, K.; Carr, C.A.; Tyler, D.J. In vivo assessment of cardiac metabolism and function in the abdominal aortic banding model of compensated cardiac hypertrophy. Cardiovasc. Res. 2015, 106, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Schnelle, M.; Chong, M.; Zoccarato, A.; Elkenani, M.; Sawyer, G.J.; Hasenfuss, G.; Ludwig, C.; Shah, A.M. In vivo [U-13C]glucose labeling to assess heart metabolism in murine models of pressure and volume overload. Am. J. Physiol. Circ. Physiol. 2020, 319, H422–H431. [Google Scholar] [CrossRef]
- Elmariah, S.; Farrell, L.A.; Furman, D.; Lindman, B.R.; Shi, X.; Morningstar, J.E.; Rhee, E.P.; Gerszten, R.E. Association of acylcarnitines with left ventricular remodeling in patients with severe aortic stenosis undergoing transcatheter aortic valve replacement. JAMA Cardiol. 2018, 3, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Haase, D.; Bäz, L.; Bekfani, T.; Neugebauer, S.; Kiehntopf, M.; Möbius-Winkler, S.; Franz, M.; Schulze, P.C. Metabolomic profiling of patients with high gradient aortic stenosis undergoing transcatheter aortic valve replacement. Clin. Res. Cardiol. 2021, 110, 399–410. [Google Scholar] [CrossRef]
- van Driel, B.O.; Schuldt, M.; Algül, S.; Levin, E.; Güclü, A.; Germans, T.; van Rossum, A.C.; Pei, J.; Harakalova, M.; Baas, A.; et al. Metabolomics in severe aortic stenosis reveals intermediates of nitric oxide synthesis as most distinctive markers. Int. J. Mol. Sci. 2021, 22, 3569. [Google Scholar] [CrossRef]
- Marcovina, S.M.; Sirtori, C.; Peracino, A.; Gheorghiade, M.; Borum, P.; Remuzzi, G.; Ardehali, H. Translating the basic knowledge of mitochondrial functions to metabolic therapy: Role of L-carnitine. Transl. Res. 2013, 161, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makrecka-Kuka, M.; Sevostjanovs, E.; Vilks, K.; Volska, K.; Antone, U.; Kuka, J.; Makarova, E.; Pugovics, O.; Dambrova, M.; Liepinsh, E. Plasma acylcarnitine concentrations reflect the acylcarnitine profile in cardiac tissues. Sci. Rep. 2017, 7, 17528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strand, E.; Pedersen, E.R.; Svingen, G.F.T.; Olsen, T.; Bjørndal, B.; Karlsson, T.; Dierkes, J.; Njølstad, P.R.; Mellgren, G.; Tell, G.S.; et al. Serum acylcarnitines and risk of cardiovascular death and acute myocardial infarction in patients with stable angina pectoris. J. Am. Heart Assoc. 2017, 6, e003620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, T.; Kelly, J.P.; McGarrah, R.W.; Hellkamp, A.S.; Fiuzat, M.; Testani, J.M.; Wang, T.S.; Verma, A.; Samsky, M.D.; Donahue, M.P.; et al. Prognostic implications of long-chain acylcarnitines in heart failure and reversibility with mechanical circulatory support. J. Am. Coll. Cardiol. 2016, 67, 291–299. [Google Scholar] [CrossRef]
- Chen, W.S.; Liu, M.H.; Cheng, M.L.; Wang, C.H. Decreases in circulating concentrations of short-chain acylcarnitines are associated with systolic function improvement after decompensated heart failure. Int. Heart J. 2020, 61, 1014–1021. [Google Scholar] [CrossRef]
- Thompson, D.K.; Sloane, R.; Bain, J.R.; Stevens, R.D.; Newgard, C.B.; Pieper, C.F.; Kraus, V.B. Daily variation of serum acylcarnitines and amino acids. Metabolomics 2012, 8, 556–565. [Google Scholar] [CrossRef] [Green Version]
- Burelle, Y.; Wambolt, R.B.; Grist, M.; Parsons, H.L.; Chow, J.C.F.; Antler, C.; Bonen, A.; Keller, A.; Dunaway, G.A.; Popov, K.M.; et al. Regular exercise is associated with a protective metabolic phenotype in the rat heart. Am. J. Physiol.—Heart Circ. Physiol. 2004, 287, H1055–H1063. [Google Scholar] [CrossRef]
- Voros, G.; Ector, J.; Garweg, C.; Droogne, W.; Van Cleemput, J.; Peersman, N.; Vermeersch, P.; Janssens, S. Increased cardiac uptake of ketone bodies and free fatty acids in human heart failure and hypertrophic left ventricular remodeling. Circ. Heart Fail. 2018, 11, e004953. [Google Scholar] [CrossRef]
- Hunter, W.G.; Kelly, J.P.; Mcgarrah, R.W.; Khouri, M.G.; Craig, D.; Haynes, C.; Ilkayeva, O.; Stevens, R.D.; Bain, J.R.; Muehlbauer, M.J.; et al. Metabolomic profiling identifies novel circulating biomarkers of mitochondrial dysfunction differentially elevated in heart failure with preserved versus reduced ejection fraction: Evidence for shared metabolic impairments in clinical heart failure. J. Am. Heart Assoc. 2016, 5, e003190. [Google Scholar] [CrossRef] [Green Version]
- Adams, S.H.; Hoppel, C.L.; Lok, K.H.; Zhao, L.; Wong, S.W.; Minkler, P.E.; Hwang, D.H.; Newman, J.W.; Garvey, W.T. Plasma acylcarnitine profiles suggest incomplete long-chain fatty acid β-oxidation and altered tricarboxylic acid cycle activity in type 2 diabetic African-American women. J. Nutr. 2009, 139, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Flam, E.; Jang, C.; Murashige, D.; Yang, Y.; Morley, M.P.; Jung, S.; Kantner, D.S.; Pepper, H.; Bedi, K.C.; Brandimarto, J.; et al. Integrated landscape of cardiac metabolism in end-stage human nonischemic dilated cardiomyopathy. Nat. Cardiovasc. Res. 2022, 1, 817–829. [Google Scholar] [CrossRef]
- Bedi, K.C.; Snyder, N.W.; Brandimarto, J.; Aziz, M.; Mesaros, C.; Worth, A.J.; Wang, L.L.; Javaheri, A.; Blair, I.A.; Margulies, K.B.; et al. Evidence for intramyocardial disruption of lipid metabolism and increased myocardial ketone utilization in advanced human heart failure. Circulation 2016, 133, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubert, G.; Martin, O.J.; Horton, J.L.; Lai, L.; Vega, R.B.; Leone, T.C.; Koves, T.; Gardell, S.J.; Krüger, M.; Hoppel, C.L.; et al. The Failing heart relies on ketone bodies as a fuel. Circulation 2016, 133, 698–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lommi, J.; Kupari, M.; Koskinen, P.; Näveri, H.; Leinonen, H.; Pulkki, K.; Härkönen, M. Blood ketone bodies in congestive heart failure. J. Am. Coll. Cardiol. 1996, 28, 665–672. [Google Scholar] [CrossRef]
- Leong, H.S.; Brownsey, R.W.; Kulpa, J.E.; Allard, M.F. Glycolysis and pyruvate oxidation in cardiac hypertrophy—Why so unbalanced? In Comparative Biochemistry and Physiology—A Molecular and Integrative Physiology; Elsevier Inc.: Amsterdam, The Netherlands, 2003; Volume 135, pp. 499–513. [Google Scholar]
- Sansbury, B.E.; DeMartino, A.M.; Xie, Z.; Brooks, A.C.; Brainard, R.E.; Watson, L.J.; DeFilippis, A.P.; Cummins, T.D.; Harbeson, M.A.; Brittian, K.R.; et al. Metabolomic analysis of pressure-overloaded and infarcted mouse hearts. Circ. Heart Fail. 2014, 7, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jaswal, J.S.; Ussher, J.R.; Sankaralingam, S.; Wagg, C.; Zaugg, M.; Lopaschuk, G.D. Cardiac insulin-resistance and decreased mitochondrial energy production precede the development of systolic heart failure after pressure-overload hypertrophy. Circ. Heart Fail. 2013, 6, 1039–1048. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.H.; Wang, Z.V. Glucose metabolism in cardiac hypertrophy and heart failure. J. Am. Heart Assoc. 2019, 8, e012673. [Google Scholar] [CrossRef] [Green Version]
- McCommis, K.S.; Douglas, D.L.; Krenz, M.; Baines, C.P. Cardiac-specific hexokinase 2 overexpression attenuates hypertrophy by increasing pentose phosphate pathway flux. J. Am. Heart Assoc. 2013, 2, e000355. [Google Scholar] [CrossRef]
- Gupte, S.A.; Levine, R.J.; Gupte, R.S.; Young, M.E.; Lionetti, V.; Labinskyy, V.; Floyd, B.C.; Ojaimi, C.; Bellomo, M.; Wolin, M.S.; et al. Glucose-6-phosphate dehydrogenase-derived NADPH fuels superoxide production in the failing heart. J. Mol. Cell. Cardiol. 2006, 41, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Drake, K.J.; Sidorov, V.Y.; McGuinness, O.P.; Wasserman, D.H.; Wikswo, J.P. Amino acids as metabolic substrates during cardiac ischemia. Exp. Biol. Med. 2012, 237, 1369–1378. [Google Scholar] [CrossRef]
- Schwartz, R.G.; Barrett, E.J.; Francis, C.K.; Jacob, R.; Zaret, B.L. Regulation of myocardial amino acid balance in the conscious dog. J. Clin. Investig. 1985, 75, 1204–1211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudge, G.H.; Mills, R.M.; Taegtmeyer, H.; Gorlin, R.; Lesch, M. Alterations of myocardial amino acid metabolism in chronic ischemic heart disease. J. Clin. Investig. 1976, 58, 1185–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinagawa, H.; Inomata, T.; Koitabashi, T.; Nakano, H.; Takeuchi, I.; Osaka, T.; Nishii, M.; Takehana, H.; Izumi, T. Increased serum bilirubin levels coincident with heart failure decompensation indicate the need for intravenous inotropic agents. Int. Heart J. 2007, 48, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, L.A.; Felker, G.M.; Pocock, S.; McMurray, J.J.V.; Pfeffer, M.A.; Swedberg, K.; Wang, D.; Yusuf, S.; Michelson, E.L.; Granger, C.B. Liver function abnormalities and outcome in patients with chronic heart failure: Data from the candesartan in heart failure: Assessment of Reduction in Mortality and Morbidity (CHARM) program. Eur. J. Heart Fail. 2009, 11, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Vítek, L.; Jirsa, M.; Brodanová, M.; Kaláb, M.; Mareček, Z.; Danzig, V.; Novotný, L.; Kotal, P. Gilbert syndrome and ischemic heart disease: A protective effect of elevated bilirubin levels. Atherosclerosis 2002, 160, 449–456. [Google Scholar] [CrossRef]
- Bulmer, A.C.; Bakrania, B.; Du Toit, E.F.; Boon, A.C.; Clark, P.J.; Powell, L.W.; Wagner, K.H.; Headrick, J.P. Bilirubin acts as a multipotent guardian of cardiovascular integrity: More than just a radical idea. Am. J. Physiol.—Heart Circ. Physiol. 2018, 315, H429–H447. [Google Scholar] [CrossRef] [Green Version]
- Hosoda, J.; Ishikawa, T.; Matsumoto, K.; Iguchi, K.; Matsushita, H.; Ogino, Y.; Taguchi, Y.; Sugano, T.; Ishigami, T.; Kimura, K.; et al. Significance of change in serum bilirubin in predicting left ventricular reverse remodeling and outcomes in heart failure patients with cardiac resynchronization therapy. J. Cardiol. 2017, 70, 416–419. [Google Scholar] [CrossRef] [Green Version]
- Gotzmann, M.; Rahlmann, P.; Hehnen, T.; Müller, P.; Lindstaedt, M.; Mügge, A.; Ewers, A. Heart failure in severe aortic valve stenosis: Prognostic impact of left ventricular ejection fraction and mean gradient on outcome after transcatheter aortic valve implantation. Eur. J. Heart Fail. 2012, 14, 1155–1162. [Google Scholar] [CrossRef]
- Marrocco, V.; Bogomolovas, J.; Ehler, E.; dos Remedios, C.G.; Yu, J.; Gao, C.; Lange, S. PKC and PKN in heart disease. J. Mol. Cell. Cardiol. 2019, 128, 212–226. [Google Scholar] [CrossRef] [Green Version]
- Bednarski, T.K.; Duda, M.K.; Dobrzyn, P. Alterations of lipid metabolism in the heart in spontaneously hypertensive rats precedes left ventricular hypertrophy and cardiac dysfunction. Cells 2022, 11, 3032. [Google Scholar] [CrossRef]
- Epand, R.M. Cholesterol in bilayers of sphingomyelin or dihydrosphingomyelin at concentrations found in ocular lens membranes. Biophys. J. 2003, 84, 3102–3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Sun, A.; Zou, Y.; Ge, J.; Lazar, J.M.; Jiang, X.C. Impact of sphingomyelin levels on coronary heart disease and left ventricular systolic function in humans. Nutr. Metab. 2011, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, R.N.; Jensen, P.N.; Hoofnagle, A.; Mcknight, B.; Fretts, A.M.; King, I.B.; Siscovick, D.S.; Psaty, B.M.; Heckbert, S.R.; Mozaffarian, D.; et al. Plasma ceramides and sphingomyelins in relation to heart failure risk: The cardiovascular health study. Circ. Heart Fail. 2019, 12, e005708. [Google Scholar] [CrossRef] [PubMed]
- Hua, T.; Bao, Q.; He, X.; Cai, W.; He, J. Lipidomics revealed alteration of sphingolipid metabolism during the reparative phase after myocardial infarction injury. Front. Physiol. 2021, 12, 663480. [Google Scholar] [CrossRef] [PubMed]
- Mielke, M.M.; Haughey, N.J.; Bandaru, V.V.R.; Weinberg, D.D.; Darby, E.; Zaidi, N.; Pavlik, V.; Doody, R.S.; Lyketsos, C.G. Plasma sphingomyelins are associated with cognitive progression in alzheimer’s disease. J. Alzheimer’s Dis. 2011, 27, 259–269. [Google Scholar] [CrossRef]
- Ussher, J.R.; Elmariah, S.; Gerszten, R.E.; Dyck, J.R.B. The Emerging Role of Metabolomics in the Diagnosis and Prognosis of Cardiovascular Disease. J. Am. Coll. Cardiol. 2016, 68, 2850–2870. [Google Scholar] [CrossRef]
- Lindman, B.R.; Patel, J.N. Multimorbidity in Older Adults with Aortic Stenosis. Clin. Geriatr. Med. 2016, 32, 305–314. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group ID | n | Description |
|---|---|---|
| Ctrl | 10 | Healthy controls |
| NEF HGAS | 10 | Normal EF, high gradient AS |
| NEF LGAS | 10 | Normal EF, low gradient AS |
| LEF HGAS | 10 | Low EF, high gradient AS |
| LEF LGAS | 10 | Low EF, low gradient AS |
| Control | NEF HGAS | LEF HGAS | LEF LGAS | NEF LGAS | |
|---|---|---|---|---|---|
| Demographics | |||||
| Male gender, n (%) | 5 (50%) | 5 (50%) | 6 (60%) | 9 (90%) | 6 (60%) |
| Age (years) | 60–70 | 79.4 ± 4.1 | 77.1 ± 9.0 | 79.5 ± 6.4 | 80.4 ± 5.9 |
| BMI | 29.66 ± 5.2 | 28.31 ± 8.2 | 25.82± 3.9 | 30.38 ± 7.3 | |
| Laboratory measures | |||||
| Hemoglobin (g/dL) | 13.2 ± 1.9 | 13.1 ± 2.2 | 12.7 ± 1.6 | 11.8 ± 1.9 | |
| Creatinine (mg/dL) | 1.27 ± 0.41 | 0.97 ± 0.14 | 1.13 ± 0.48 | 1.23 ± 0.40 | |
| NT-proBNP (ng/L) | 2717 ± 4412 | 10964 ± 12751 | 7069 ± 7388 | 2210 ± 1317 | |
| Echocardiography | |||||
| LVEF (%) | 63.4 ± 8.8 | 30.1 ± 9.1 | 33.3 ± 7.1 | 61.9 ±7.6 | |
| LVEDD (mm) | 40. 7 ± 3.3 | 54.6 ± 9.6 | 51.8 ± 7.3 | 43.1 ± 5.6 | |
| Aortic valve orifice area (cm2) | 0.70 ± 0.16 | 0.59 ± 0.11 | 0.71 ± 0.18 | 0.77 ± 0.14 | |
| Peak gradient (mmHg) | 52.9 ± 10.4 | 48.4 ± 11.3 | 26.7 ± 5.0 | 26.3 ± 5.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bengel, P.; Elkenani, M.; Beuthner, B.E.; Pietzner, M.; Mohamed, B.A.; Pollok-Kopp, B.; Krätzner, R.; Toischer, K.; Puls, M.; Fischer, A.; et al. Metabolomic Profiling in Patients with Different Hemodynamic Subtypes of Severe Aortic Valve Stenosis. Biomolecules 2023, 13, 95. https://doi.org/10.3390/biom13010095
Bengel P, Elkenani M, Beuthner BE, Pietzner M, Mohamed BA, Pollok-Kopp B, Krätzner R, Toischer K, Puls M, Fischer A, et al. Metabolomic Profiling in Patients with Different Hemodynamic Subtypes of Severe Aortic Valve Stenosis. Biomolecules. 2023; 13(1):95. https://doi.org/10.3390/biom13010095
Chicago/Turabian StyleBengel, Philipp, Manar Elkenani, Bo E. Beuthner, Maik Pietzner, Belal A. Mohamed, Beatrix Pollok-Kopp, Ralph Krätzner, Karl Toischer, Miriam Puls, Andreas Fischer, and et al. 2023. "Metabolomic Profiling in Patients with Different Hemodynamic Subtypes of Severe Aortic Valve Stenosis" Biomolecules 13, no. 1: 95. https://doi.org/10.3390/biom13010095