
Isolation and Characterization of the Arapaima gigas Growth Hormone (ag-GH) cDNA and Three-Dimensional Modeling of This Hormone in Comparison with the Human Hormone (hGH)

 , , , , ,
, , , , ,
Abstract
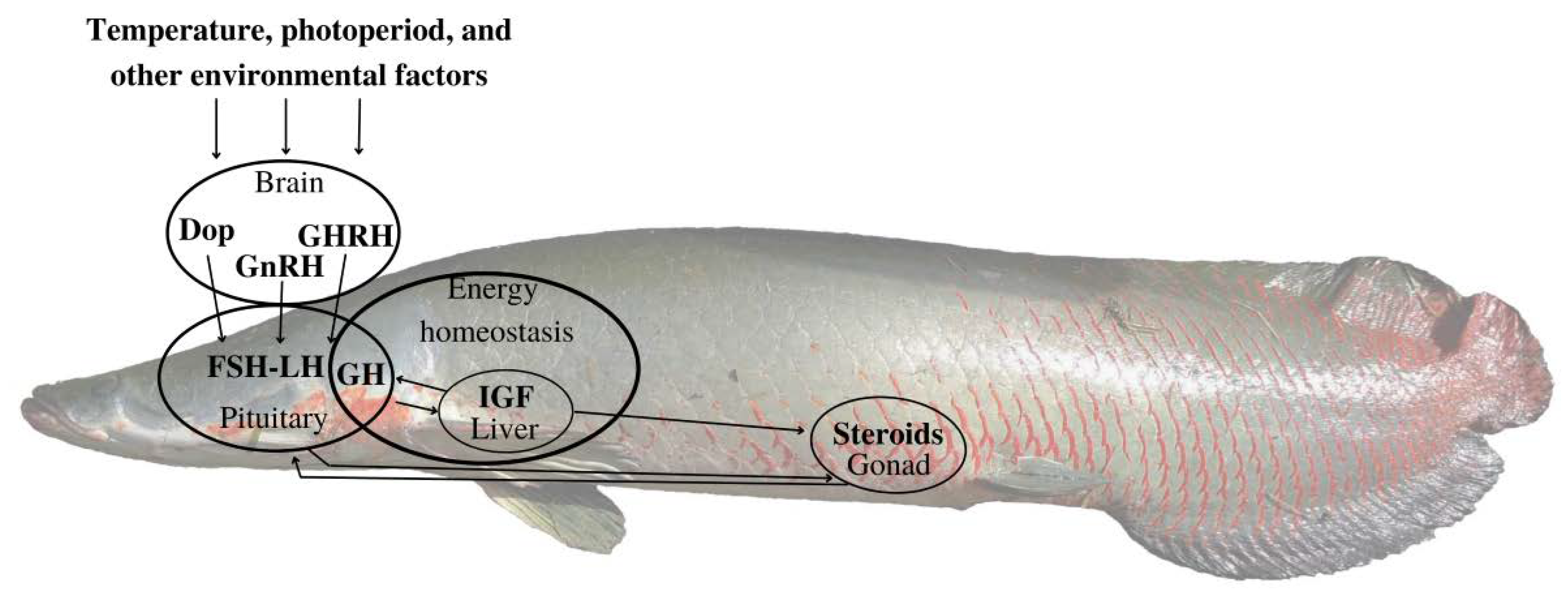
:1. Introduction

2. Materials and Methods
2.1. A. gigas Pituitary Gland Sources and the Detection of a First Partial Region of ag-GH
2.2. In Silico Prediction Analysis
2.3. Obtaining the Coding Region from the Pituitary Gland cDNA of A. gigas
2.4. Rapid Amplification of cDNA Ends (RACE) for Obtaining agGH 3′UTR
2.5. Molecular Modelling
2.6. Molecular Dynamics
2.7. Quantitative Similarity Comparison between the ag-GH Model and the Template (hGH)
3. Results
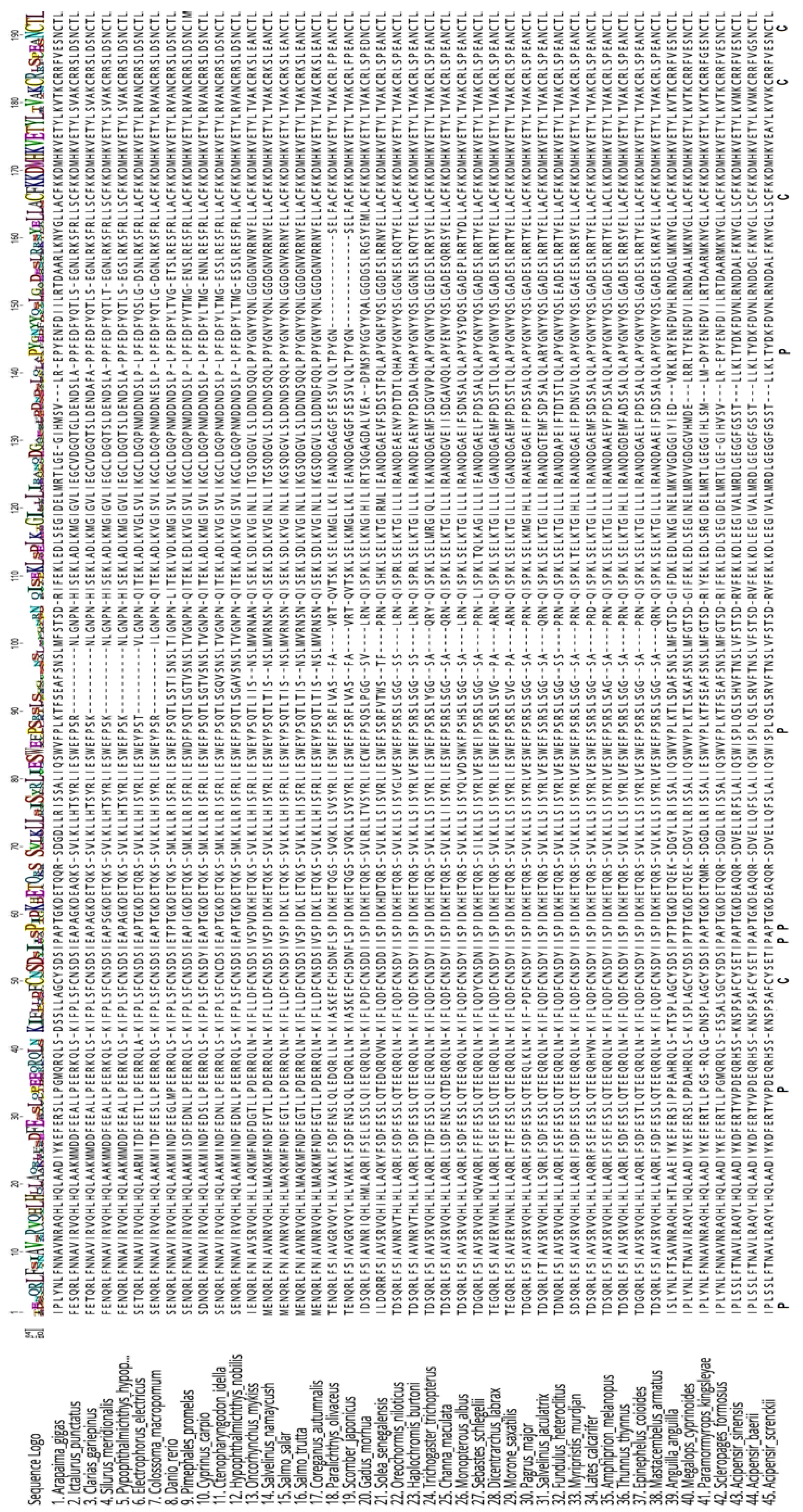
3.1. cDNA Sequencing
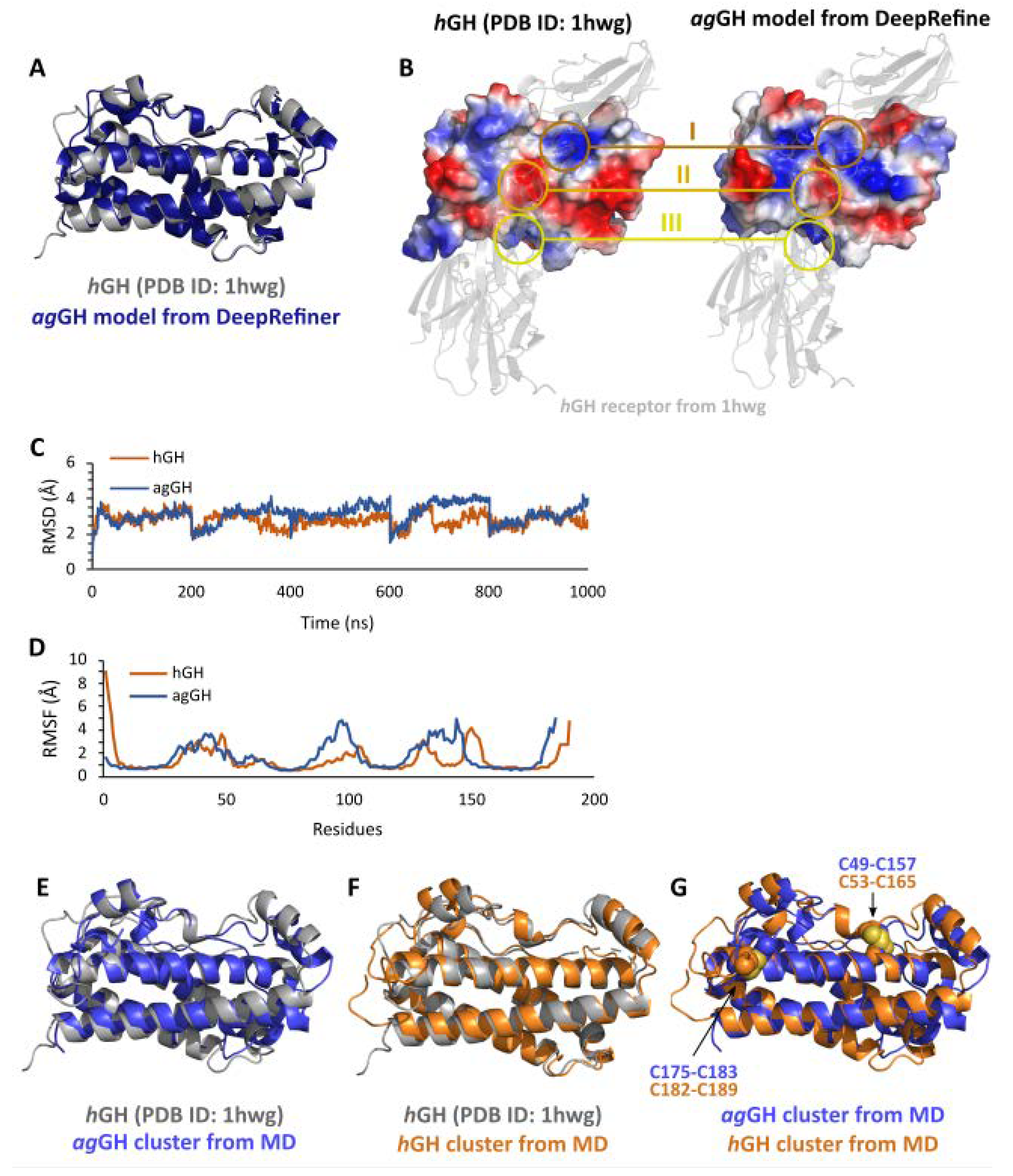
3.2. Three-Dimensional Model of agGH
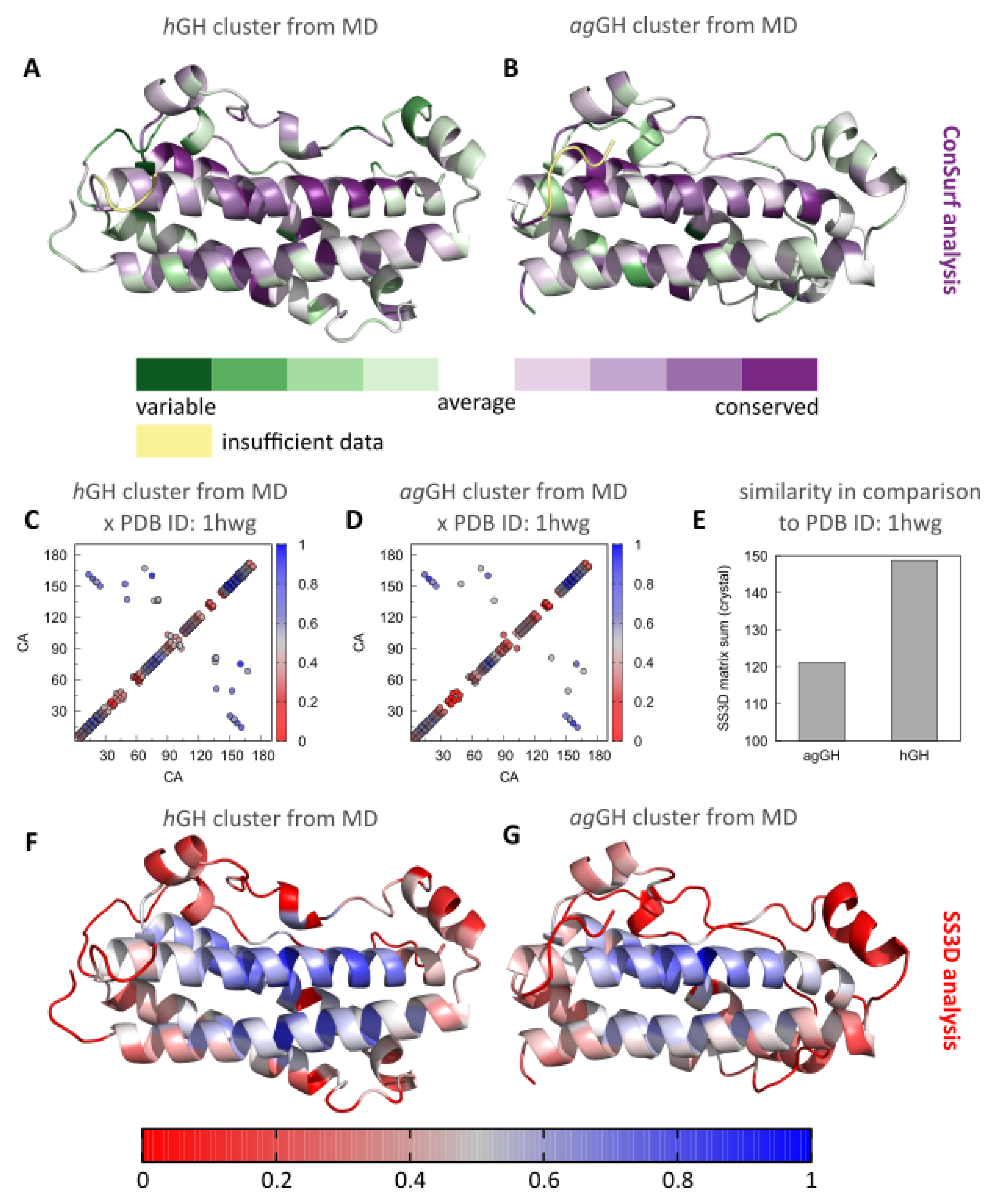
3.3. Evolutionary Implications and Comparison of the Three-Dimensional Contacts of ag-GH and hGH Models
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Borella, M.I.; Venturieri, R.; Mancera, J.M. Immunocytochemical identification of adenohypophyseal cells in the pirarucu (Arapaima gigas), an Amazonian basal teleost. Fish. Physiol. Biochem. 2009, 35, 3–16. [Google Scholar] [CrossRef]
- Cavole, L.M.; Arantes, C.C.; Castello, L. How illegal are tropical small-scale fisheries? An estimate for Arapaima in the Amazon. Fish. Res. 2015, 168, 1–5. [Google Scholar] [CrossRef]
- Watson, L.C.; Stewart, D.J.; Kretzer, A.M. Genetic diversity and population structure of the threatened Giant Arapaima in southwestern Guyana: Implications for their conservation. Copeia 2016, 104, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.; Tello, S.; Duponchelle, F. Patterns of commercial fish landings in the Loreto region (Peruvian Amazon) between 1984 and 2006. Fish. Physiol. Biochem. 2009, 35, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Chu-Koo, F.; Dugue, R.; Aguilar, M.A.; Daza, A.C.; Bocanegra, F.A.; Veintemilla, C.C.; Duponchelle, F.; Renno, J.F.; Tello, S.; Nunes, J. Gender determination in the Paiche or Pirarucu using plasma vitellogenin. 17 beta-estradiol and 11-ketotestosterone levels. Fish. Physiol. Biochem. 2009, 35, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Mylonas, C.C.; Fostier, A.; Zanuy, S. Broodstock management and hormonal manipulations of fish reproduction. Gen. Comp. Endocrinol. 2010, 165, 516–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castello, L.; Stewart, D.J.; Arantes, C.C. Modeling population dynamics and conservation of arapaima in the Amazon. Rev. Fish. Biol. Fish. 2011, 21, 623–640. [Google Scholar] [CrossRef]
- Torati, L.S.; Varges, A.P.S.; Galvão, J.A.S.; Mesquita, P.E.C.; Migaud, H. Endoscopy application in broodstock management of Arapaima gigas (Schinz, 1822). J. Appl. Ichthyol. 2016, 32, 353–355. [Google Scholar] [CrossRef]
- Torati, L.S.; Migaud, H.; Doherty, M.K.; Siwy, J.; Mullen, W.; Mesquita, P.E.C.; Albalat, A. Comparative proteome and peptidome analysis of the cephalic fluid secreted by Arapaima gigas (Teleostei: Osteoglossidae) during and outside parental care. PLoS ONE 2017, 12, e0186692. [Google Scholar] [CrossRef] [Green Version]
- Torati, L.S.; Lima, A.F.; Kirschnik, L.N.G.; Migaud, H. Endoscopy and cannulation as non-invasive tools to identify sex and monitor reproductive development in Arapaima gigas. Copeia 2019, 107, 287–296. [Google Scholar] [CrossRef]
- Huang, H.; Zhang, Y.; Huang, W.; Li, S.; Zhu, P.; Liu, Y.; Yin, S.; Liu, X.; Lin, H. Molecular characterization of marbled eel (Anguilla marmorata) gonadotropin subunits and their mRNA expression profiles during artificially induced gonadal development. Gen. Comp. Endocrinol. 2009, 162, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Levavi-Sivan, B.; Bogerd, J.; Mañanós, E.L.; Gómez, A.; Lareyre, J.J. Perspectives on fish gonadotropins and their receptors. Gen. Comp. Endocrinol. 2010, 165, 412–437. [Google Scholar] [CrossRef] [PubMed]
- Schulz, R.W.; de França, L.R.; Larteyre, J.J.; Le Gac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef]
- Senthilkumaran, B. Recent advances in meiotic maturation and ovulation: Comparing mammals and pisces. Front. Biosci. 2011, 16, 1898–1914. [Google Scholar] [CrossRef] [Green Version]
- Faria, M.T.; Carvalho, R.F.; Sevilhano, T.C.A.; Oliveira, N.A.J.; Silva, C.F.P.; Oliveira, J.E.; Soares, C.R.J.; Garcez, R.; Santo, P.R.; Bartolini, P. Isolation of the pituitary gonadotrophic α-subunit hormone of the giant Amazonian fish: Pirarucu (Arapaima gigas). Fish. Phys. Biochem. 2013, 39, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Sevilhano, T.; Carvalho, R.F.; Oliveira, N.A.; Oliveira, J.E.; Maltarollo, V.G.; Trossini, G.; Garcez, R.; Bartolini, P. Molecular cloning and characterization of pirarucu (Arapaima gigas) follicle-stimulating hormone and luteinizing hormone β-subunit cDNAs. PLoS ONE 2017, 12, e0183545. [Google Scholar] [CrossRef] [Green Version]
- Canosa, L.F.; Chang, J.P.; Peter, R.E. Neuroendocrine control of growth hormone in fish. Gen. Comp. Endocrinol. 2007, 151, 1–26. [Google Scholar] [CrossRef]
- Franz, A.C.; Oliver, F.; Kollner, B.; Shved, N.; Link, K.; Casanova, A.; Wenger, M.; D´Cotta, H.; Baroiller, J.F.; Ullrich, O.; et al. Endocrine and local IGF-I in the bony fish immune system. Biology 2016, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Gopal, R.N.; Kumar, P.; Lal, B. Temperature dependent action of growth hormone on somatic growth and testicular activities of the catfish, Clarias batrachus. Gen. Comp. Endocrinol. 2014, 195, 125–131. [Google Scholar] [CrossRef]
- Vialle, R.A.; de Souza, J.E.S.; Lopes, K.D.; Teixeira, D.G.; Sobrinho, P.D.A.; Ribeiro-dos-Santos, A.M.; Furtado, C.; Sakamoto, T.; Silva, F.A.O.; de Oliveira, E.H.C.; et al. Whole genome sequencing of the pirarucu (Arapaima gigas) supports independent emergence of major teleost clades. Genome Biol. Evol. 2018, 10, 2366–2379. [Google Scholar] [CrossRef]
- González-Villaseñor, L.I.; Zhang, P.; Chen, T.T.; Powers, D.A. Molecular cloning and sequencing of coho salmon growth hormone cDNA. Gene 1988, 65, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Liu, C.S.; Huang, F.L.; Lo, T.B. The primary structures of growth hormones of three cyprinid species: Bighead carp, silver carp and grass carp. Gen. Comp. Endocrinol. 1992, 87, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Scharlt, M. Sequence of the growth hormone (GH) gene from the silver carp (Hypophthalmichthys molitrix) and evolution of GH genes in vertebrates. Biochim. Biophys. Acta 1993, 1174, 285–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, J.S.; Wolff, J.L.C.; Araujo, R.D.; Hilsdorf, A.W.S. Molecular cloning and sequence analysis of growth hormone cDNA of Neotropical freshwater fish Pacu (Piaractus mesopotamicus). Genet. Mol. Biol. 2008, 31, 381–384. [Google Scholar] [CrossRef] [Green Version]
- Sekine, S.; Mizukami, T.; Nishi, T.; Kuwana, Y.; Saito, A.; Sato, M.; Itoh, S.; Kawauchi, H. Cloning and expression of cDNA for salmon growth hormone in Escherichia coli. Proc. Natl. Acad. Sci. USA 1985, 82, 4306–4310. [Google Scholar] [CrossRef] [Green Version]
- Saito, A.; Sekine, S.; Komatsu, Y.; Sato, M.; Hirano, T.; Itoh, S. Molecular cloning of eel growth hormone and its expression in Escherichia coli. Gene 1988, 73, 545–551. [Google Scholar]
- Tsai, H.J.; Chen, H.M.; Lo, C.F. Secretory synthesis of active recombinant fish growth hormone by insect cells using a baculovirus vector. Can. J. Fish. Aquat. Sci. 1994, 51, 1–7. [Google Scholar] [CrossRef]
- Tsai, H.J.; Hsih, M.H.; Kuo, J.C. Escherichia coli-produced fish growth hormone as a feed additive to enhance the growth of juvenile black seabream (Acanthopagrus schlegeli). J. Appl. Ichthyol. 1997, 13, 79–82. [Google Scholar] [CrossRef]
- Sciara, A.A.; Rubiolo, J.A.; Somoza, G.M.; Arranz, S.E. Molecular cloning, expression and immunological characterization of pejerrey (Odontesthes bonariensis) growth hormone. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 284–292. [Google Scholar] [CrossRef]
- Li, C.; Chen, X.; Zhang, Y.; Ye, H.; Liu, T. Molecular and expression characterization of growth hormone/prolactin family genes in the Prenant’s schizothoracin. Mol. Biol. Rep. 2011, 38, 4595–4602. [Google Scholar] [CrossRef]
- Lundberg, J.G.; Chernoff, B. A Miocene Fossil of the Amazonian Fish Araipama (Teleostei, Arapaimidae) from the Magdalena River Region of Colombia-Biogeographic and Evolutionary Implications. Biotropica 1992, 24, 2–14. [Google Scholar] [CrossRef]
- Rainer, F.; Pauly, D. (Eds.) Arapaimidae. FishBase. August 2013. Available online: http://www.fishbase.org/Summary/FamilySummary.php?Family=Arapaimidae (accessed on 1 June 2022).
- Migaud, H.; Davie, A.; Taylor, J.F. Current knowledge on the photoneuroendocrine regulation of reproduction in temperature fish species. J. Fish Biol. 2010, 76, 27–68. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souvorov, Y.; Kapustin, B.; Kiryitin, V.; Chetvernin, T.; Tatusova, T.; Lipman, D. Gnomon—NCBI eukaryotic gene predicyion tool. National Center for Biotechnology Information; NCBI: Bethesda, MD, USA, 25 February 2010. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of highquality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2008, 46, W296–W303 (2018). [Google Scholar] [CrossRef] [Green Version]
- Biener, S.; Waterhouse, A.; de Beer, T.A.P.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository—New features and functionality. Nucleic Acids Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef] [Green Version]
- De Vos, A.M.; Ultsch, M.; Kossiakoff, A.A. Human growth hormone and extracellular domain of its receptor—Crystal-structure of the complex. Science 1992, 255, 306–312. [Google Scholar] [CrossRef]
- Sundstrom, M.; Lundqvist, T.; Rodin, J.; Giebel, L.B.; Milligan, D.; Norstedt, G. Crystal structure of an antagonist mutant of human growth hormone, G120R, in complex with its receptor at 2.9 angstrom resolution. J. Biol. Chem. 1996, 271, 32197–32203. [Google Scholar] [CrossRef] [Green Version]
- Clakson, T.; Ultsch, M.H.; Wells, J.A.; de Vos, A.M. Structural and functional analysis of the 1:1 growth hormone: Receptor complex reveals the molecular basis for receptor affinity. J. Mol. Biol. 1998, 277, 1111–1128. [Google Scholar] [CrossRef]
- Somers, W.; Ultsch, M.; De Vos, A.M.; Kossiakoff, A.A. The X-ray structure of a growth hormone-prolactin receptor complex. Nature 1994, 372, 478–484. [Google Scholar] [CrossRef]
- Atwell, S.; Ultsch, M.; De Vos, A.M.; Wells, J.A. Structural plasticity in a remodeled protein-protein interface. Science 1997, 278, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo—Distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Shuvo, M.H.; Gulfam, M.; Bhattacharya, D. DeepRefiner: High-accuracy protein structure refinement by deep network calibration. Nucleic Acids Res. 2021, 49, W147–W152. [Google Scholar] [CrossRef] [PubMed]
- Fiser, A.; Sali, A. ModLoop: Automated modeling of loops in protein structures. Bioinformatics 2003, 19, 2500–2501. [Google Scholar] [CrossRef] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Varadi, M.; Anyango, S.; Deshpande, M.; Sreenath, N.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2021, 50, D439–D444. [Google Scholar] [CrossRef]
- Maciejewwski, M.W.; Schuyler, A.D.; Gryk, M.R.; Moraru, I.I.; Romero, P.R.; Ulrich, E.L.; Eghbalnia, H.R.; Livny, M.; Delaglio, F.; Hoch, J.C. NMRbox: A resourche for biomolecular NMR computation. Biophys. J. 2017, 112, 1529–1534. [Google Scholar] [CrossRef] [Green Version]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [Green Version]
- Hooft, R.W.W.; Sander, C.; Vriend, G. Objectively judging the quality of a protein structure from a Ramachandran plot. Comput. Appl. Biosci. 1997, 13, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Shelley, J.C.; Cholleti, A.; Frye, L.L.; Greenwood, J.R.; Timlin, M.R.; Uchimaya, M. Epik: A software program for pK(a) prediction and protonation state generation for drug-like molecules. J. Comp. Aided Mol. Des. 2007, 21, 681–691. [Google Scholar] [CrossRef]
- Lu, C.; Wu, C.J.; Ghoreishi, D.; Chen, W.; Wang, L.L.; Damm, W.; Ross, G.A.; Dahlgren, M.K.; Russell, E.; Von Barger, C.D.; et al. OPLS4: Improving force field accuracy on challenging regimes of chemical space. J. Chem. Theory Comput. 2021, 17, 4291–4300. [Google Scholar] [CrossRef] [PubMed]
- Bowers, K.J.; Chow, E.; Xu, H.; Dror, R.O.; Eastwood, M.P.; Gregersen, B.A.; Klepeis, J.L.; Kolossvary, I.; Moraes, M.A.; Sacerdoti, F.D.; et al. Scalable algorithms for molecular dynamics simulations on commodity clustersTitle. In Proceedings of the 2006 ACM/IEEE Conference on Supercomputing, Tampa, FL, USA, 11–17 November 2006; ACM: New York, NY, USA, 2006. [Google Scholar] [CrossRef]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald—An n.log(n) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Ashkenazy, H.; Abadi, S.; Martz, E.; Chay, O.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf 2016: An improved methodology to estimate and visualize evolutionary conservation in macromolecules. Nucleic Acids Res. 2016, 44, W344–W350. [Google Scholar] [CrossRef] [Green Version]
- Celniker, G.; Nimrod, G.; Ashkenazy, H.; Glaser, F.; Martz, E.; Mayrose, I.; Pupko, T.; Ben-Tal, N. ConSurf: Using evolutionary data to raise testable hypotheses about protein function. Isr. J. Chem. 2013, 53, 199–206. [Google Scholar] [CrossRef]
- Ashkenazy, H.; Erez, E.; Martz, E.; Pupko, T.; Ben-Tal, N. ConSurf 2010: Calculating evolutionary conservation in sequence and structure of proteins and nucleic acids. Nucleic Acids Res. 2010, 38, W529–W533. [Google Scholar] [CrossRef] [Green Version]
- Lima, I.; Cino, E.A. Sequence similarity in 3D for comparison of protein families. J. Mol. Graph. Model. 2021, 106, 107906. [Google Scholar] [CrossRef] [PubMed]
- Teufel, F.; Armenteros, J.J.A.; Johansen, A.R.; Gislason, M.H.; Pihl, S.I.; Tsirigos, K.D.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 6.0 predicts all five types of signal peptides using protein language models. Nat. Biotechnol. 2022, 40, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Venugopal, T.; Anathy, V.; Pandian, T.J.; Gong, G.Z.; Mathavan, S. Molecular cloning of growth hormone-encoding cDNA of an Indian major carp, Labeo rohita, and its expression in Escherichia coli and zebrafish. Gen. Comp. Endocrinol. 2002, 125, 236–247. [Google Scholar] [CrossRef]
- Morgan, A.A.; Rubenstein, E. Proline: The distribution, frequency, positioning, and common functional roles of proline and polyproline sequences in the human proteome. PLoS ONE 2013, 8, e53785. [Google Scholar] [CrossRef]
- Bellini, M.H.; Bartolini, P. In vivo bioassay for the potency determination of human growth hormone in dwarf “little” mice. Endocrinology 1993, 132, 2051–2055. [Google Scholar] [CrossRef] [PubMed]
- Maltarollo, V.G.; Shevchenko, E.; de Miranda Lima, I.D.; Cino, E.A.; Ferreira, G.M.; Poso, A.; Kronenberger, T. Do go chasing waterfalls: Enoyl reductase (FabI) in complex with 6inhibitors stabilize tetrameric structure and opens water channels. J. Comp. Inf. Model. 2022, 62, 5746–5761. [Google Scholar] [CrossRef] [PubMed]
- Kronenberger, T.; Serafim, M.S.M.; Tonduru, A.K.; Maltarollo, V.G.; Poso, A. Ligand accessibility insights to the dengue virus NS3-NS2B protease assessed by long-timescale molecular dynamics simulations. ChemMedChem 2021, 16, 2524–2534. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.M.; Pillaiyar, T.; Hirata, M.H.; Poso, A.; Kronenberger, T. Inhibitor induced conformational changes in SARS-CoV-2 papain-like protease. Sci. Rep. 2022, 12, 11585. [Google Scholar] [CrossRef]
- Fan, H.; Mark, A.E. Refinement of homology-based protein structures by molecular dynamics simulation techniques. Protein Sci. 2022, 13, 211–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayano, A.M.; Simoes-Silva, R.; Medeiros, P.S.M.; Maltarollo, V.G.; Honorio, K.M.; Oliveira, E.; Albericio, F.; da Silva, S.L.; Aguiar, A.C.C.; Krettli, A.U.; et al. BpMP-1, a new metalloproteinase isolated from Bothrops brazili snake venom with in vitro antiplasmodial properties. Toxicon 2015, 106, 30–41. [Google Scholar] [CrossRef]
- Zaqueo, K.D.; Kayano, A.M.; Simoes-Silva, R.; Moreira-Dill, L.S.; Fernandes, C.F.C.; Fuly, A.L.; Maltarollo, V.G.; Honorio, K.M.; da Silva, S.L.; Acosta, G.; et al. Isolation and biochemical characterization of a new thrombin-like serine protease from Bothrops pirajai snake venom. Biomed. Res. Int. 2014, 595186. [Google Scholar] [CrossRef] [Green Version]
- Martins, W.; Baldasso, P.A.; Honorio, K.M.; Maltarollo, V.G.; Ribeiro, R.I.M.A.; Carvalho, B.M.A.; Soares, A.M.; Calderon, L.A.; Stabeli, R.G.; Caballol, M.A.O.; et al. A novel phospholipase A(2) (D49) from the venom of the Crotalus oreganus abyssus (North American Grand Canyon rattlesnake). Biomed. Res. Int. 2014, 654170. [Google Scholar] [CrossRef]
- Cooper, M.; Brown, C.D. Qualifying the relationship between sequence conservation and molecular function. Genome Res. 2008, 18, 201–205. [Google Scholar] [CrossRef] [Green Version]
- Naidu, C.K.; Suneetha, Y.; Reddy, P.S. Computational analysis of molt-inhibiting hormone from selected crustaceans. Comp. Biochem. Physiol. Part D Genom. Proteom. 2013, 8, 292–299. [Google Scholar] [CrossRef]
- Cecchi, C.R.; Higuti, E.; Oliveira, N.A.J.; Lima, E.R.; Jakobsen, M.; Dagnaes-Hansen, F.; Gissel, H.; Aagaard, L.; Jensen, T.G.; Jorge, A.A.L.; et al. A novel homologous model for gene therapy of dwarfism by non-viral transfer of the mouse growth hormone into immunocompetent dwarf mice. Curr. Gene Ther. 2014, 14, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Cecchi, C.R.; Higuti, E.; Lima, E.R.; Vieira, D.P.; Squair, P.L.; Peroni, C.N.; Bartolini, P. Efficient non-invasive plasmid-DNA administration into tibialis cranialis muscle of “little” mice. Curr. Gene Ther. 2017, 17, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Lima, E.R.; Cecchi, C.R.; Higuti, E.; de Jesus, G.P.P.; Gomes, A.M.; Zacarias, E.A.; Bartolini, P.; Peroni, C.N. Optimization of mouse growth hormone plasmid DNA electrotransfer into tibialis cranialis muscle of “little” mice. Molecules 2020, 25, 5034. [Google Scholar] [CrossRef] [PubMed]
- Higuti, E.; Cecchi, C.R.; Oliveira, N.A.J.; Vieira, D.P.; Jensen, T.G.; Jorge, A.A.L.; Bartolini, P.; Peroni, C.N. Growth responses following a single intra-muscular hGH plasmid administration compared to daily injections of hGH in dwarf mice. Curr. Gene Ther. 2012, 12, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Higuti, E.; Cecchi, C.R.; Oliveira, N.A.J.; Lima, E.R.; Vieira, D.P.; Vieira, D.P.; Jensen, T.G.; Jorge, A.A.L.; Bartolini, P.; Peroni, C.N.; et al. Partial correction of the dwarf phenotype by non-viral transfer of the growth hormone gene in mice: Treatment age is critical. Growth Horm. IGF Res. 2016, 26, 1–7. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Direction | Name | Sequence |
|---|---|---|---|
| Primer 1 | Sense | ag-GH1 | 5′ TCA GCA AGA CCT CCC CA 3′ |
| Primer 2 | Antisense | ag-GH2 | 5′ TTG AAG CTG CAT GTG GGA TCC TTG 3′ |
| Primer 3 | Sense | GSP1 | 5′ GAG GCT TCT TAC TGG GCT CC 3′ |
| Primer 4 | Antisense | GSP6 | 5′ AAG TGC AGT TGC TCT CCA CAA 3′ |
| Primer 5 | Antisense | AP | 5′ GGC CAC GCG TCG ACT AGT ACT TTT TTT TTT TTT TTT T 3′ |
| Primer 6 | Sense or Antisense | AUAP | 5′ GGC CAC GCG TCG ACT AGT AC 3′ |
| Primer 7 | Sense | GSP8 | 5′ CTC TCA AGA CCT TCA GCG AGG 3′ |
| Organism | Nucleotide (mRNA) | Amino Acid | Superorder, Order, and Family |
|---|---|---|---|
| Arapaima gigas | Osteoglossomorpha, Osteoglossiformes, Arapaimidae | ||
| Acipenser baerii | JX947839.1 | AFX71645.1 | Chondrostei, Acipenseriformes, Acipenseridae |
| Anguilla Japonica | M24066.1 | AAA48535.1 | Elopomorpha, Anguilliformes, Anguillidae |
| Anguilla anguilla | AY148493.1 | AAN61122.1 | Elopomorpha, Anguilliformes, Anguillidae |
| Chanos chanos | XM_030794045.1 | XP_030649905.1 | Ostariophysi, Cypriniformes, Cyprinidae |
| Colossoma macropomum | XM_036564141.1 | XP_036420034.1 | Ostariophysi, Characiformes, Serrasalmidae |
| Coregonus clupeaformis | XM_041852232.2 | XP_041708166.2 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Electrophorus electricus | XM_027026729.2 | XP_026882530.1 | Ostariophysi, Gymnotiformes, Gymnotidae |
| Lates calcarifer | X59378.1 | CAA42022.1 | Percoidei, Perciformes, Latidae |
| Megalops cyprinoides | XM_036552661.1 | XP_036408554.1 | Elopomorpha, Elopiformes, Megalopidae |
| Myripristis murdjan | XM_030057829.1 | XP_029913689.1 | Berycoidei, Beryciformes, Holocentridae |
| Pangasianodon hypophthalmus | JF303887.1 | ADX99243.1 | Ostariophysi, Siluriformes, Pangasiidae |
| Paramormyrops kingsleyae | XM_023835741.1 | XP_023691509.1 | Osteoglossomorpha, Osteoglossiformes, Mormyridae |
| Scleropages formosus | XM_018764341.2 | XP_018619857.1 | Osteoglossomorpha, Osteoglossiformes, Osteoglossidae |
| Salmo trutta | XM_029727357.1 | XP_029583217.1 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Salmo salar | X14305.1 | CAA32481.1 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Salvelinus namaycush | XM_038989124.1 | XP_038845052.1 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Toxotes jaculatrix | XM_041062256.1 | XP_040918190.1 | Acanthopterygii, Perciformes, Toxotidae |
| Salvellinus alpinus | XM_024010963.1 | XP_023866731.1 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Monopterus albus | AY265351.1 | AAP42271.1 | Acanthopterygii, Synbranchiformes, Synbranchidae |
| Sebastes schlegelii | U89917.1 | AAB49492.1 | Acanthopterygii, Scorpaeniformes, Sebastidae |
| Solea senegalensis | U01143.1 | AAA60372.1 | Acanthopterygii, Pleuronectiformes, Soleidae |
| Paralichthys olivaceus | D29737.1 | BAA06159.1 | Acanthopterygii, Pleuronectiformes, Paralichthyidae |
| Thunnus thynnus | X06735.1 | CAA29914.1 | Acanthopterygii, Perciformes, Scombridae |
| Scomber japonicus | GU138643.1 | ACZ04987.1 | Acanthopterygii, Perciformes, Scombridae |
| Chrysophrys major | AB904715.1 | BAR88081.1 | Acanthopterygii, Perciformes, Sparidae |
| Epinephelus coioides | AY513647.1 | AAR97729.1 | Acanthopterygii, Perciformes, Serranidae |
| Morone saxatilis | XM_035674082.1 | XP_035529975.1 | Acanthopterygii, Perciformes, Moronidae |
| Dicentrarchus labrax | GQ918491.1 | ADB23477.1 | Acanthopterygii, Perciformes, Moronidae |
| Amphiprion melanopus | HM135406.1 | ADJ57589.1 | Acanthopterygii, Perciformes, Pomacentridae |
| Oreochromis niloticus | KT387598.1 | AMB21545.1 | Acanthopterygii, Perciformes, Cichlidae |
| Acipenser screnckii | KC460212.2 | AGI96360.2 | Chondrostei, Acipenseriformes, Acipenseridae |
| Acipenser sinensis | EU599640.2 | ACC86123.1 | Chondrostei, Acipenseriformes, Acipenseridae |
| Ctenopharyngodon idella | M27094.1 | AAA58724.1 | Ostariophysi, Cypriniformes, Cyprinidae |
| Danio rerio | AJ937858.1 | CAI79040.1 | Ostariophysi, Cypriniformes, Cyprinidae |
| Hypophthalmichthys nobilis | GQ497221.1 | ADD62434.1 | Ostariophysi, Cypriniformes, Cyprinidae |
| Pimephales promelas | AY643399.1 | AAT91088.1 | Ostariophysi, Cypriniformes, Cyprinidae |
| Clarias gariepinus | FJ823972.1 | ACN97175.1 | Ostariophysi, Siluriformes, Clariidae |
| Ictalurus punctatus | EU009499.1 | ABS70461.1 | Ostariophysi, Siluriformes, Ictaluridae |
| Silurus meridionalis | AF530481.2 | AAP82934.1 | Ostariophysi, Siluriformes, Siluridae |
| Coregonus autumnalis | X77245.1 | CAA54461.1 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Oncorhynchus mykiss | NM_001124689.1 | NP_001118161.1 | Protacanthopterygii, Salmoniformes, Salmonidae |
| Gadus morhua | EU676171.1 | ACD46080.1 | Paracanthopterygii, Gadiformes, Gadidae |
| Fundulus heteroclitus | XM_012872083.3 | XP_012727537.1 | Acanthopterygii, Cyprinodontiformes, Fundulidae |
| Trichogaster trichopterus | AF157633.1 | AAG60346.1 | Acanthopterygii, Perciformes, Osphronemidae |
| Superorder | Order | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. | Osteoglossomorpha | Osteoglossiformes (3) | 86.4 | 53.8 | 62.7 | 63.1 | 63.2 | 74.5 | 82.0 | 79.7 | 62.6 | 68.2 | 66.3 | 65.6 | 62.3 | 70.8 |
| 2. | Acanthopterygii | Pleuronectiformes (2) | - | 63.7 | 68.1 | 67.6 | 68.3 | 55.7 | 45.7 | 45.2 | 63.0 | 68.5 | 53.5 | 80.7 | 59.5 | 78.0 |
| 3. | Acanthopterygii | Scorpaeniformes (1) | - | 100 | 84.8 | 84.3 | 66.0 | 40.1 | 40.1 | 72.6 | 78.2 | 52.4 | 84.9 | 65.7 | 86.3 | |
| 4. | Acanthopterygii | Synbranchiformes (1) | - | 100 | 86.3 | 67.2 | 40.1 | 40.1 | 72.1 | 77.7 | 52.9 | 84.9 | 66.7 | 86.5 | ||
| 5. | Berycoidei | Beryciformes (1) | - | 100 | 66.9 | 41.5 | 41.0 | 74.1 | 79.5 | 55.8 | 84.7 | 71.6 | 87.4 | |||
| 6. | Chondrostei | Acipenseriformes (3) | - | 97.2 | 81.5 | 79.4 | 62.2 | 66.6 | 69.0 | 64.8 | 65.7 | 68.4 | ||||
| 7. | Elopomorpha | Elopiformes (1) | - | 100 | 86.6 | 69.5 | 78.6 | 49.1 | 75.5 | 38.4 | 82.8 | |||||
| 8. | Elopomorpha | Anguilliformes (1) | - | 100 | 68.0 | 78.1 | 46.7 | 75.5 | 37.9 | 82.7 | ||||||
| 9. | Ostaryophysi | Siluriformes (3) | - | 95.2 | 82.7 | 88.8 | 71.0 | 71.9 | 78.8 | |||||||
| 10. | Ostaryophysi | Cypriniformes (5) | - | 92.9 | 89.2 | 68.5 | 77.6 | 78.3 | ||||||||
| 11. | Ostaryophysi | Characiformes (1) | - | 100 | 78.3 | 49.0 | 87.2 | |||||||||
| 12. | Acanthopterygii | Perciformes (9) | - | 84.8 | 62.3 | 70.8 | ||||||||||
| 13. | Paracanthopterygii | Gadiformes (1) | - | 100 | 85.6 | |||||||||||
| 14. | Protacanthopterygii | Salmoniformes (6) | - | 97.0 |
| Common name | Scientific Name | Identity (%) | GenBank/NCBI |
|---|---|---|---|
| Pirarucu | Arapaima gigas | 100.0 | OP575308 |
| Crocodile | Crocodylus porosus | 61.5 | AAB34999.1 |
| Manatee | Trichechus manatus | 60.9 | XP_004374274.1 |
| Australian parakeet | Melopsittacus undulatus | 59.4 | XP_005140349.2 |
| Whale | Balaenoptera physalus | 58.9 | CAH42020.1 |
| Dolphin | Tursiops truncatus | 58.9 | XP_033703592.1 |
| Orca | Orcinus orca | 58.9 | XP_004275687.2 |
| Mouse | Mus musculus | 57.8 | AAH61157.1 |
| Ox | Bos taurus | 56.3 | AAX07134.1 |
| Chimpanzee | Pan troglodytes | 44.7 | AAL72284.1 |
| Human | Homo sapiens | 44.7 | KAI2584577.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, E.R.; Freire, R.P.; Suzuki, M.F.; Oliveira, J.E.; Yosidaki, V.L.; Peroni, C.N.; Sevilhano, T.; Zorzeto, M.; Torati, L.S.; Soares, C.R.J.; et al. Isolation and Characterization of the Arapaima gigas Growth Hormone (ag-GH) cDNA and Three-Dimensional Modeling of This Hormone in Comparison with the Human Hormone (hGH). Biomolecules 2023, 13, 158. https://doi.org/10.3390/biom13010158
Lima ER, Freire RP, Suzuki MF, Oliveira JE, Yosidaki VL, Peroni CN, Sevilhano T, Zorzeto M, Torati LS, Soares CRJ, et al. Isolation and Characterization of the Arapaima gigas Growth Hormone (ag-GH) cDNA and Three-Dimensional Modeling of This Hormone in Comparison with the Human Hormone (hGH). Biomolecules. 2023; 13(1):158. https://doi.org/10.3390/biom13010158
Chicago/Turabian StyleLima, Eliana Rosa, Renan Passos Freire, Miriam Fussae Suzuki, João Ezequiel Oliveira, Vanessa Luna Yosidaki, Cibele Nunes Peroni, Thaís Sevilhano, Moisés Zorzeto, Lucas Simon Torati, Carlos Roberto Jorge Soares, and et al. 2023. "Isolation and Characterization of the Arapaima gigas Growth Hormone (ag-GH) cDNA and Three-Dimensional Modeling of This Hormone in Comparison with the Human Hormone (hGH)" Biomolecules 13, no. 1: 158. https://doi.org/10.3390/biom13010158