
Diversity and Potential Multifunctionality of Archaeal CetZ Tubulin-like Cytoskeletal Proteins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Analysis of Archaeal Tubulin Superfamily Homologues
2.2. Analysis of cetZ Genomic Regions
3. Results
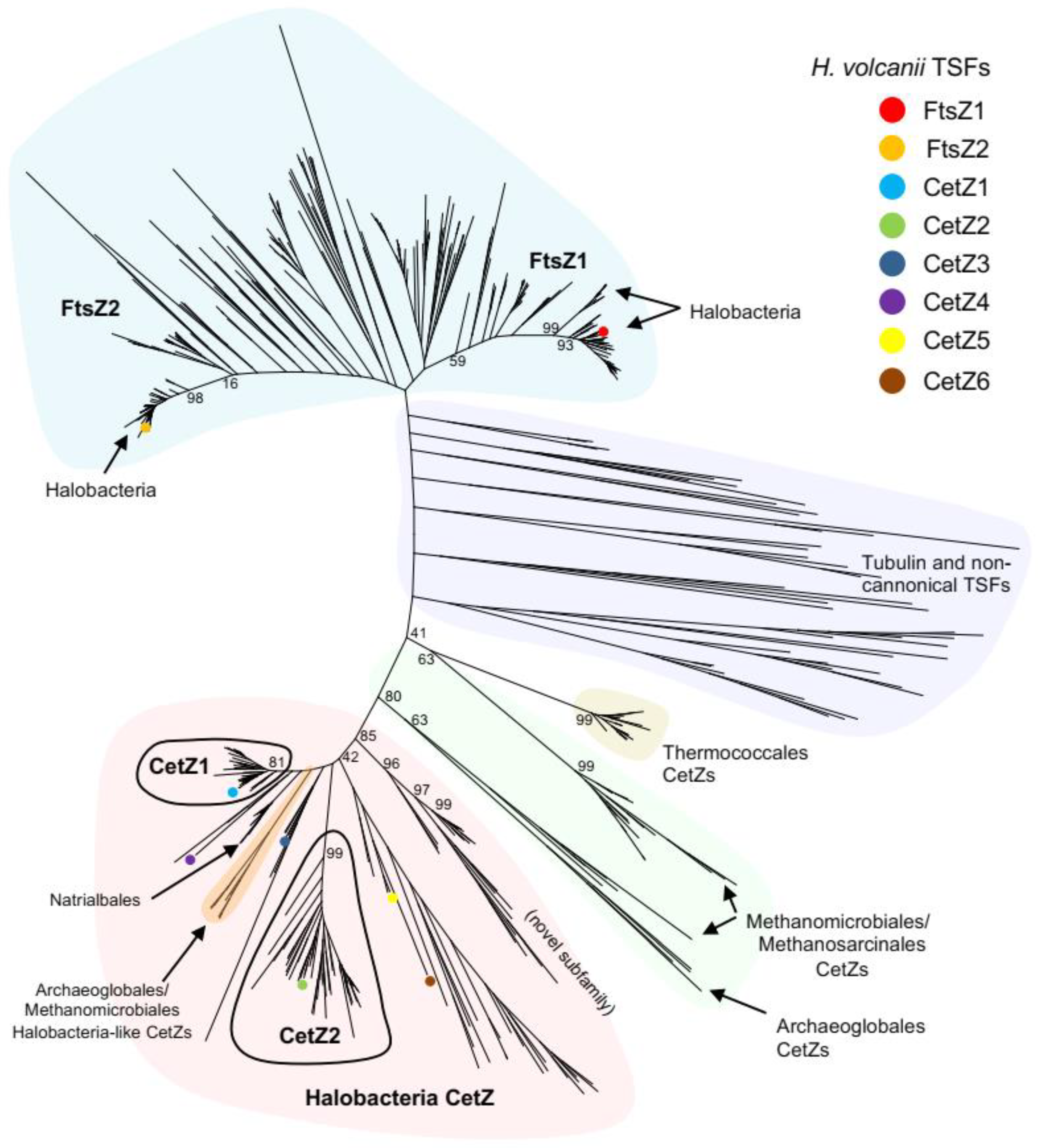
3.1. Identification and Classification of Tubulin Superfamily Proteins in Archaea
3.2. Multiple CetZs Are Abundant in Halobacteria
3.3. Deep Branching CetZs in Thermoccales Define the CetZ Family Boundary
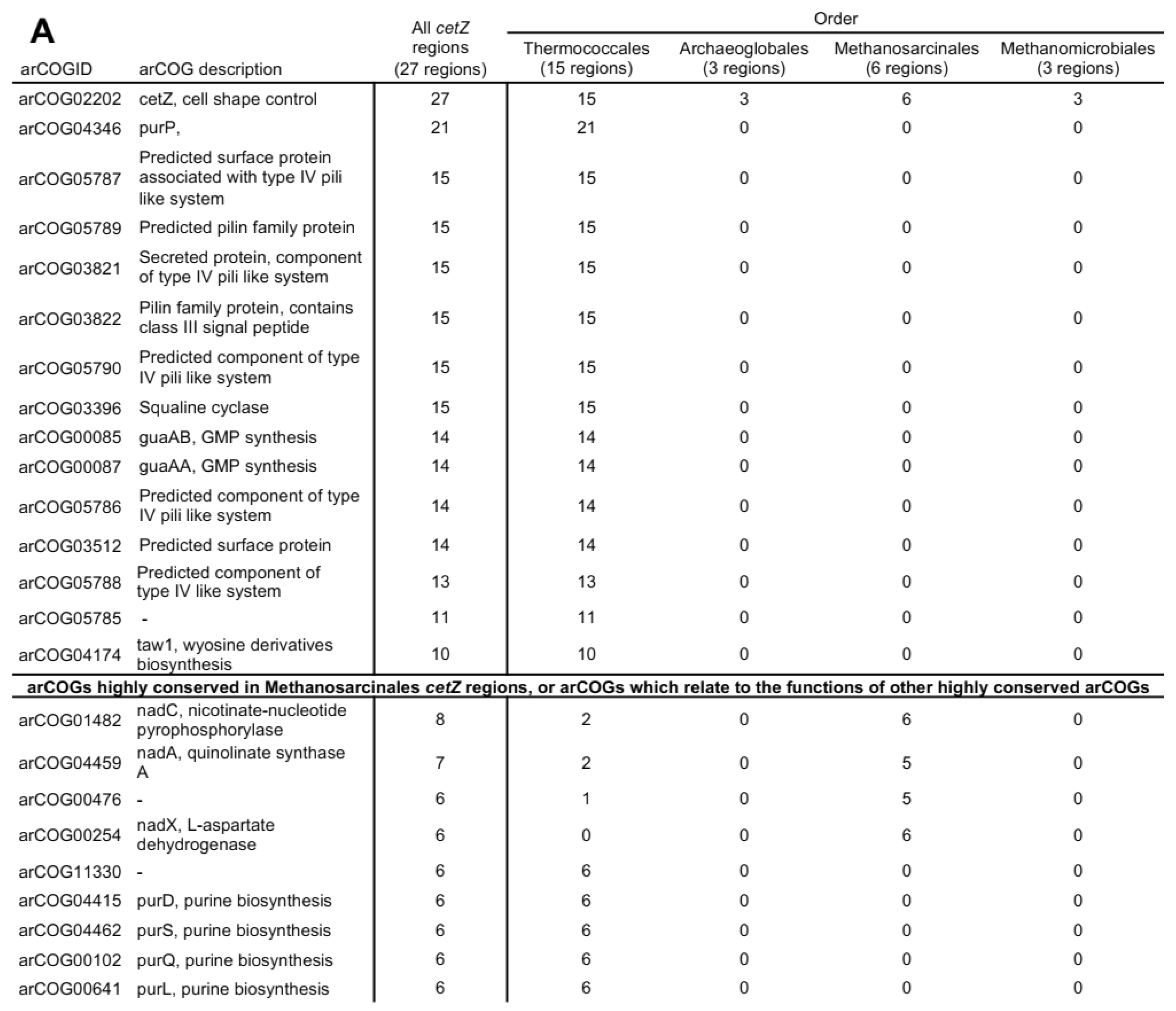
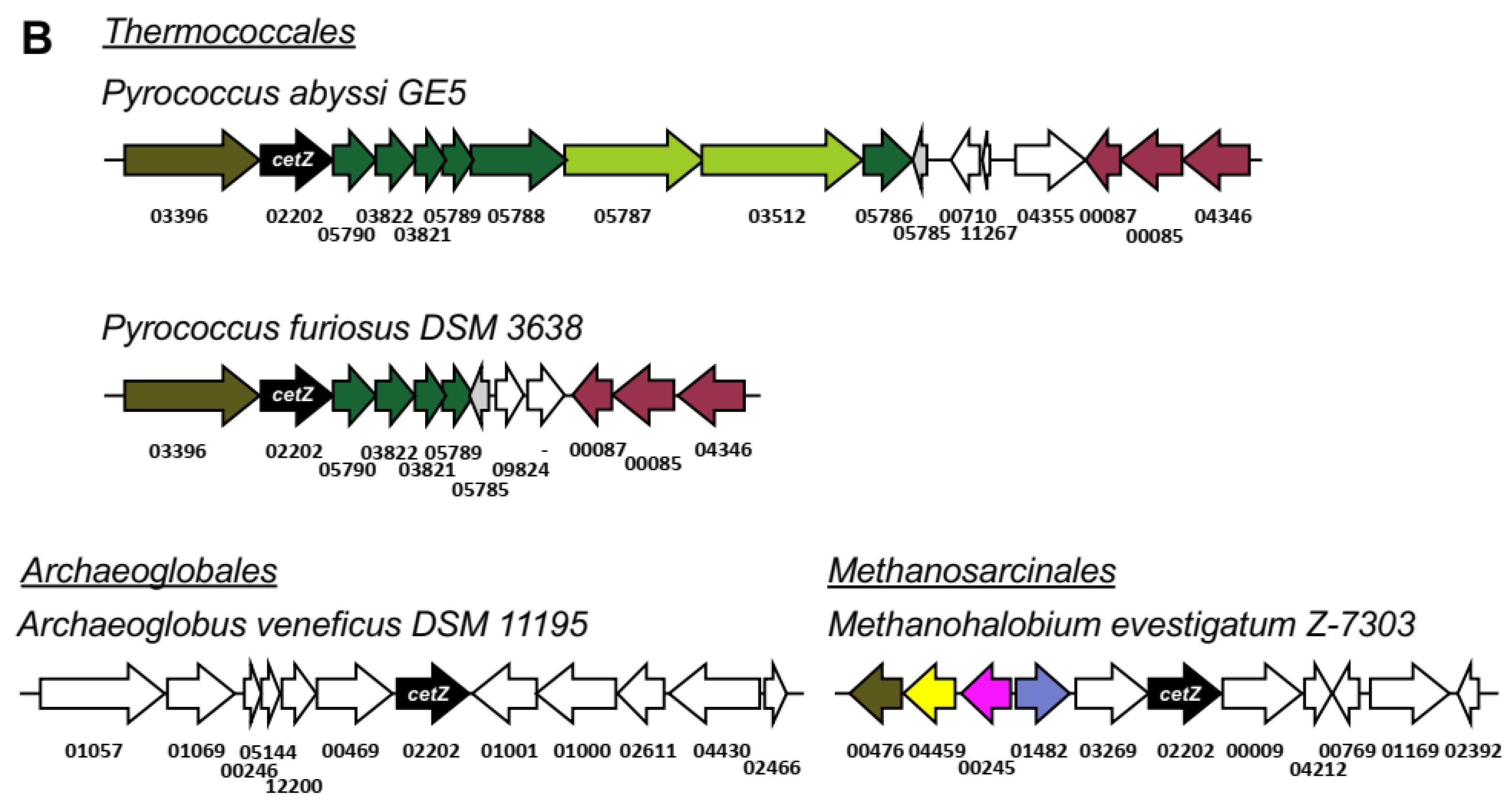
3.4. CetZs in Archaeoglobales, Methanomicrobiales, and Methanosarcinales
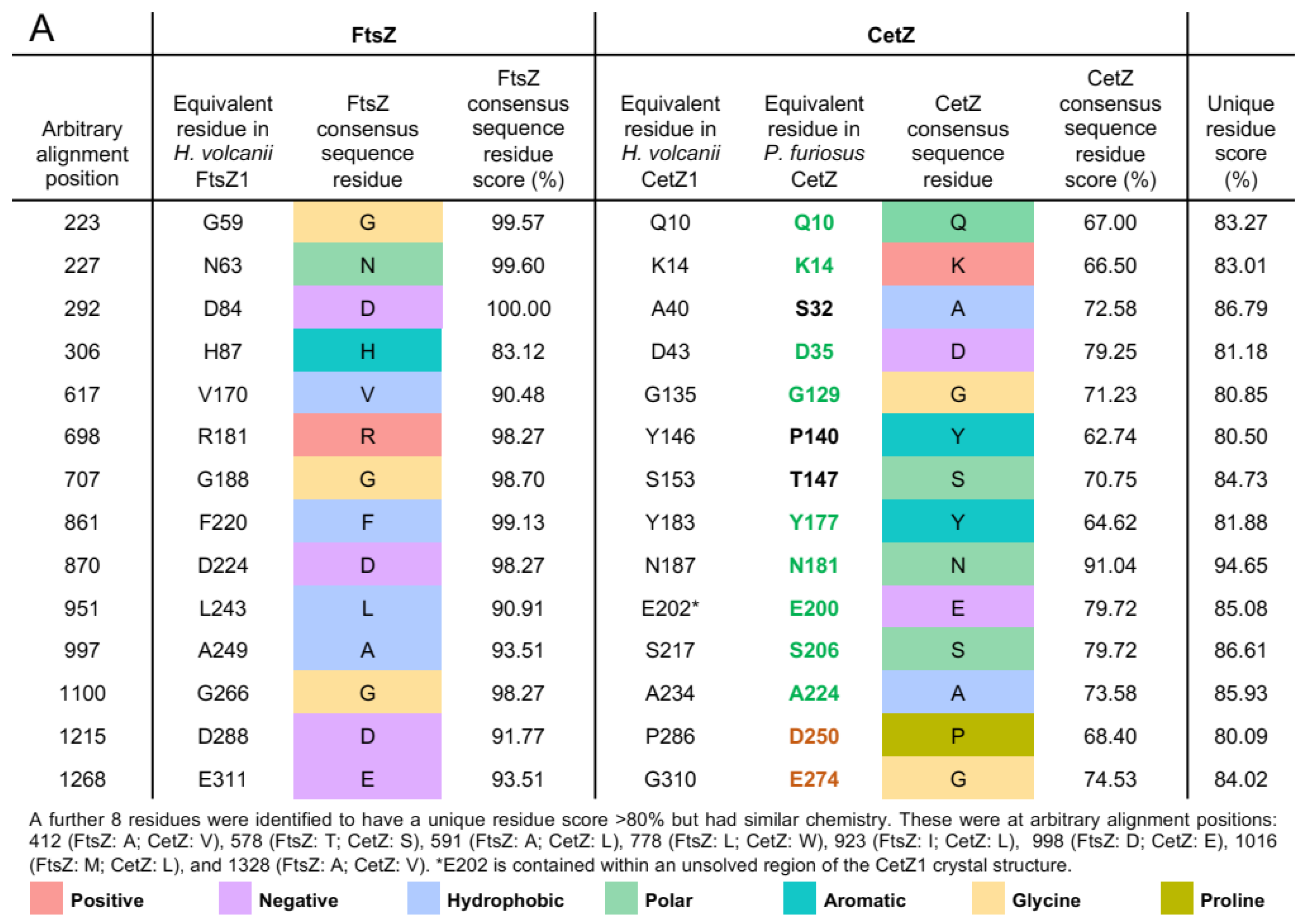
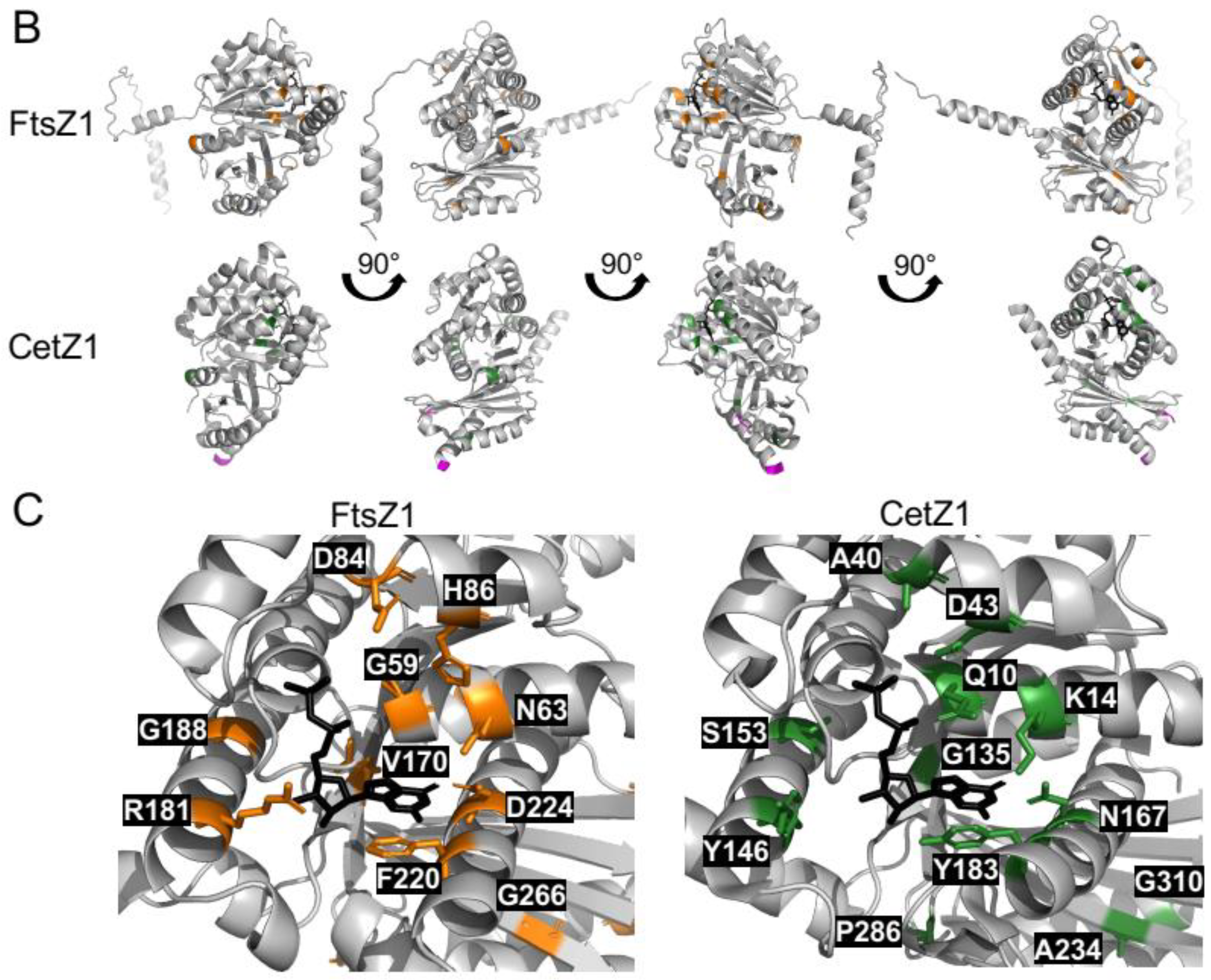
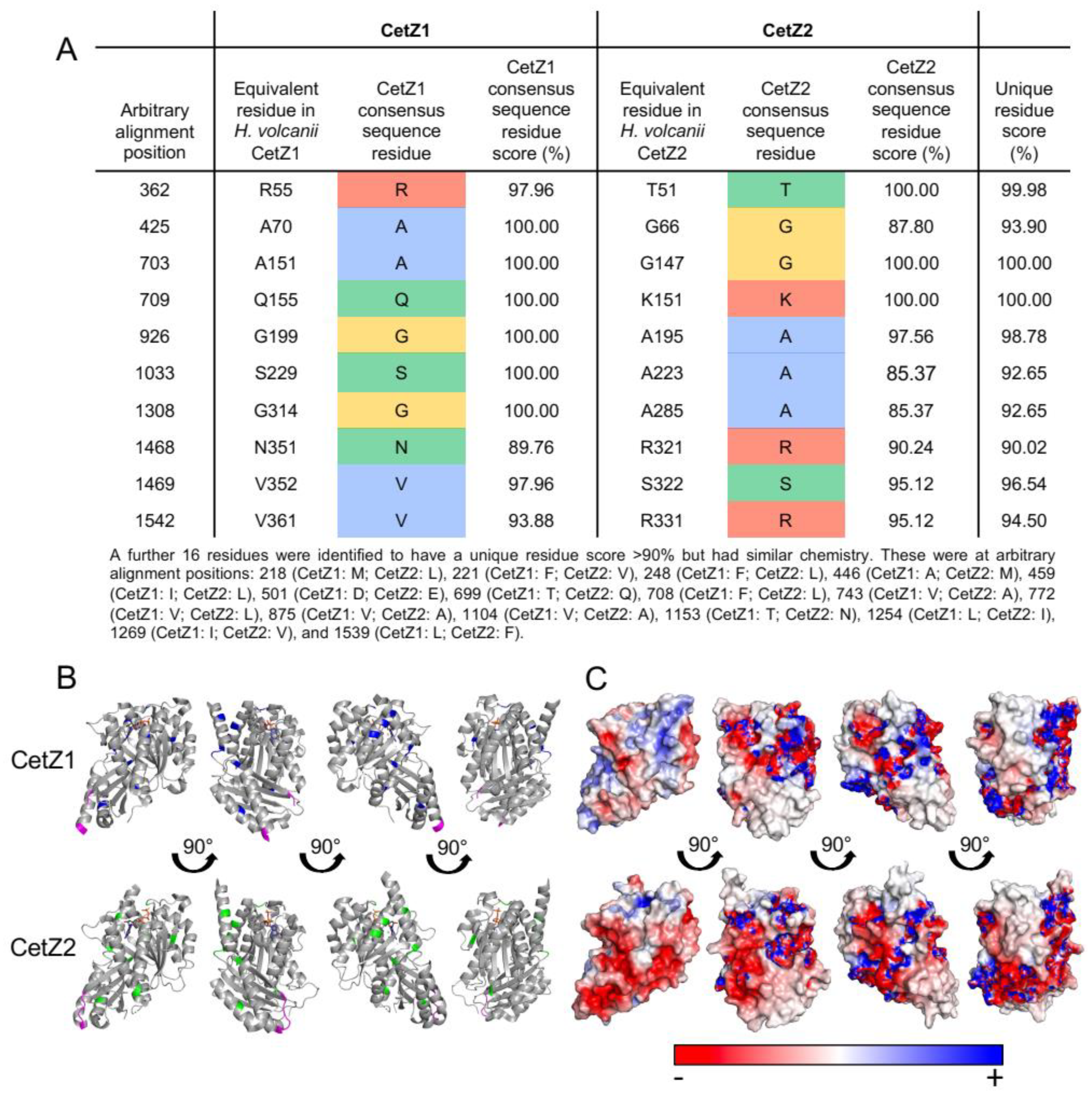
3.5. Halobacteria CetZ1 and CetZ2 Subfamilies Show Distinct Characteristics
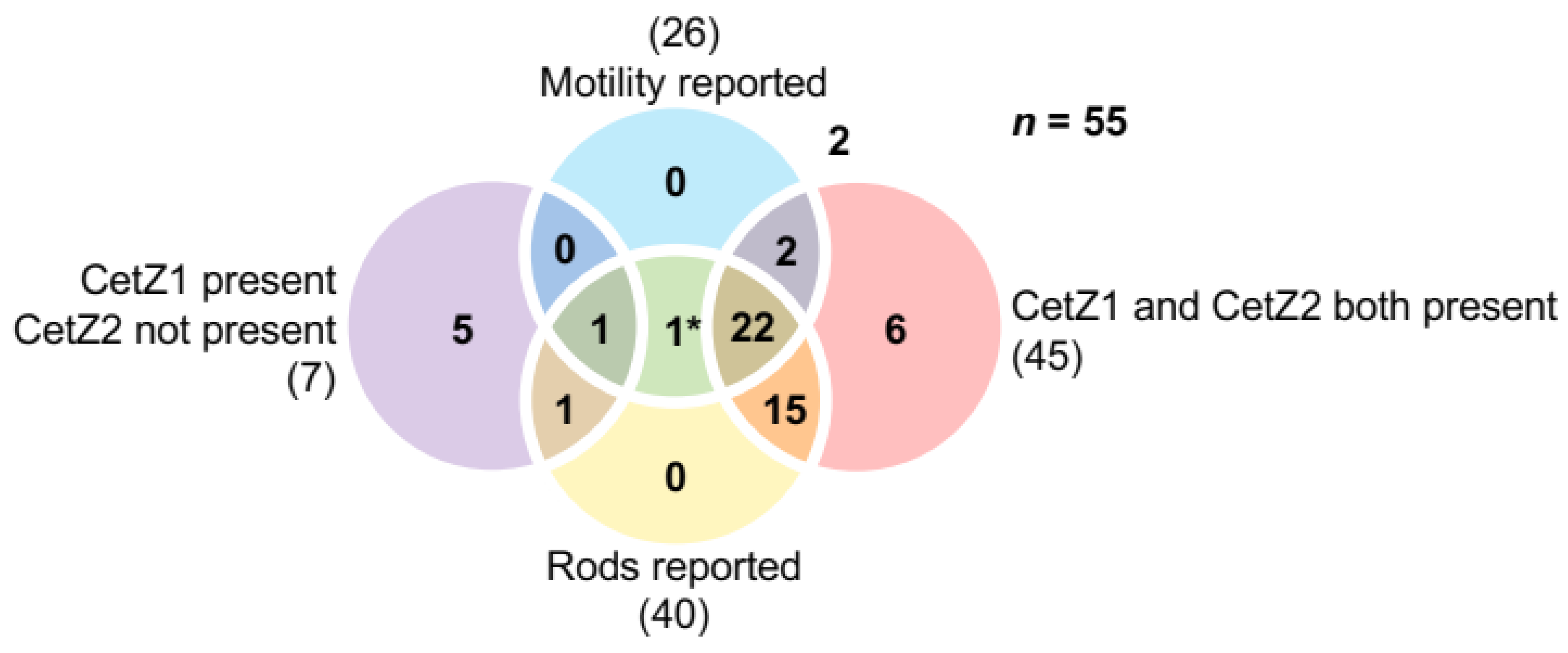
3.6. The Presence of CetZ1 and CetZ2 in Halobacteria Correlates with Rod Shape and Motility
3.7. cetZ1 Genomic Regions Are Associated with Cofactor and Nucleotide Biosynthesis
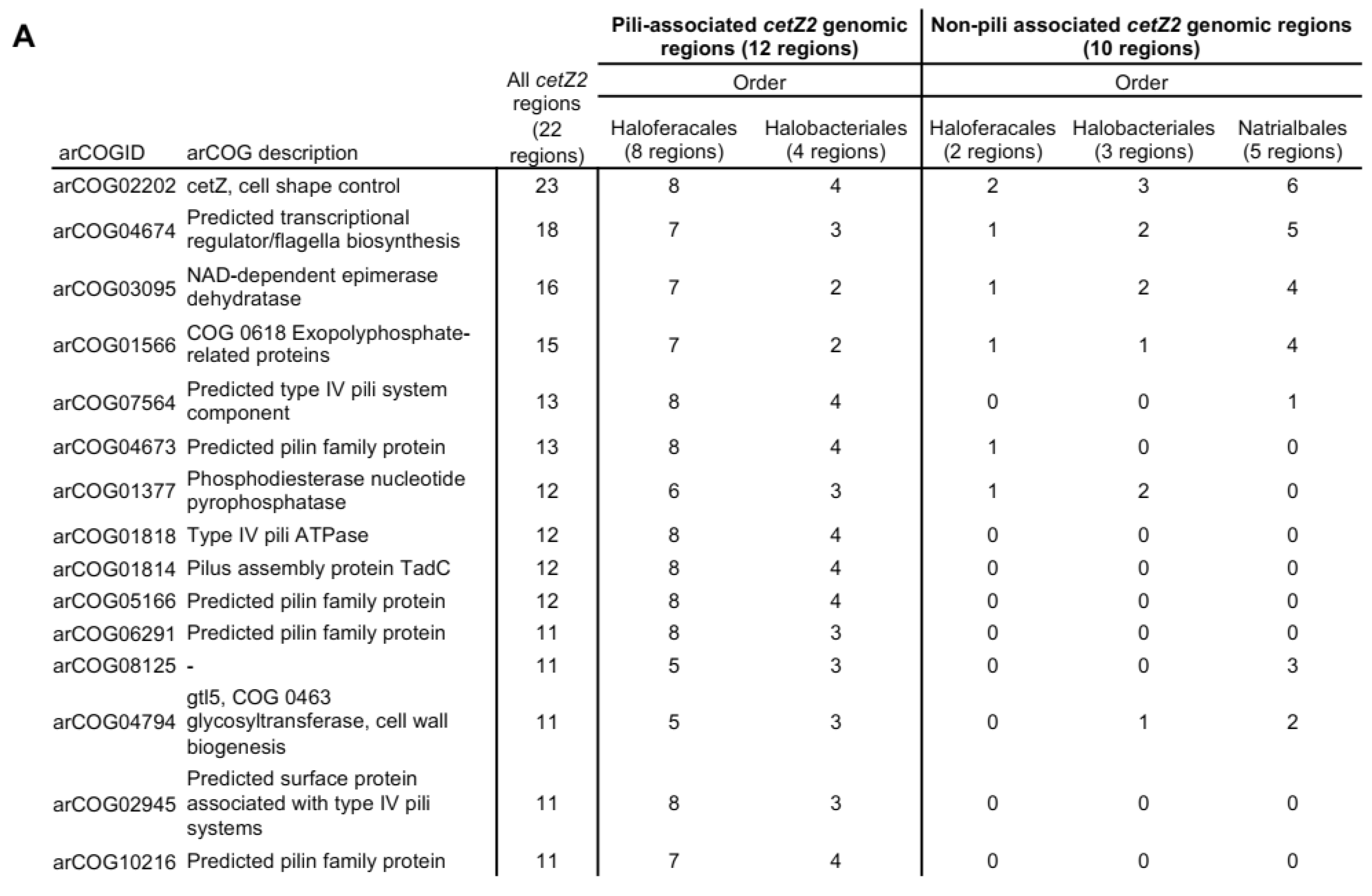
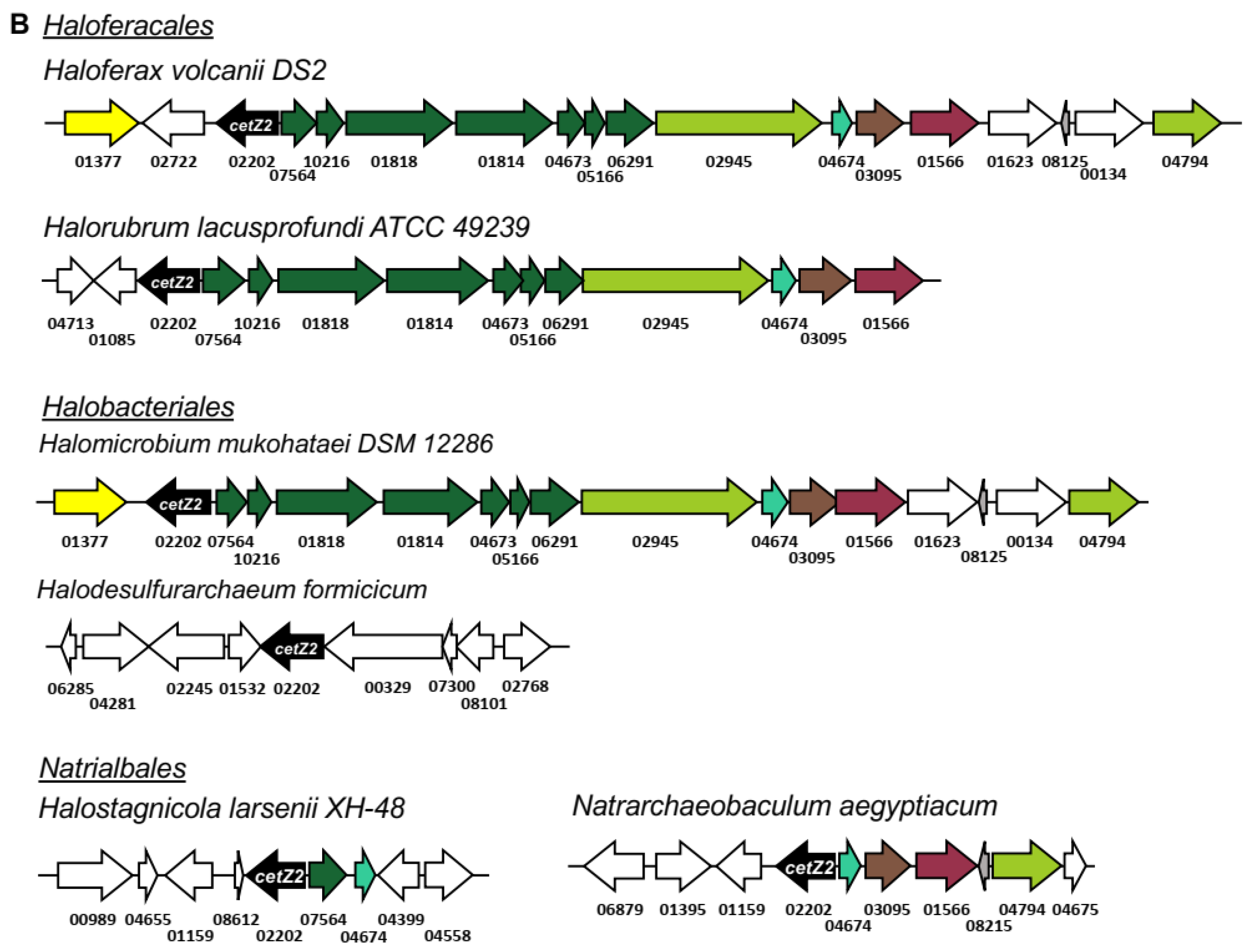
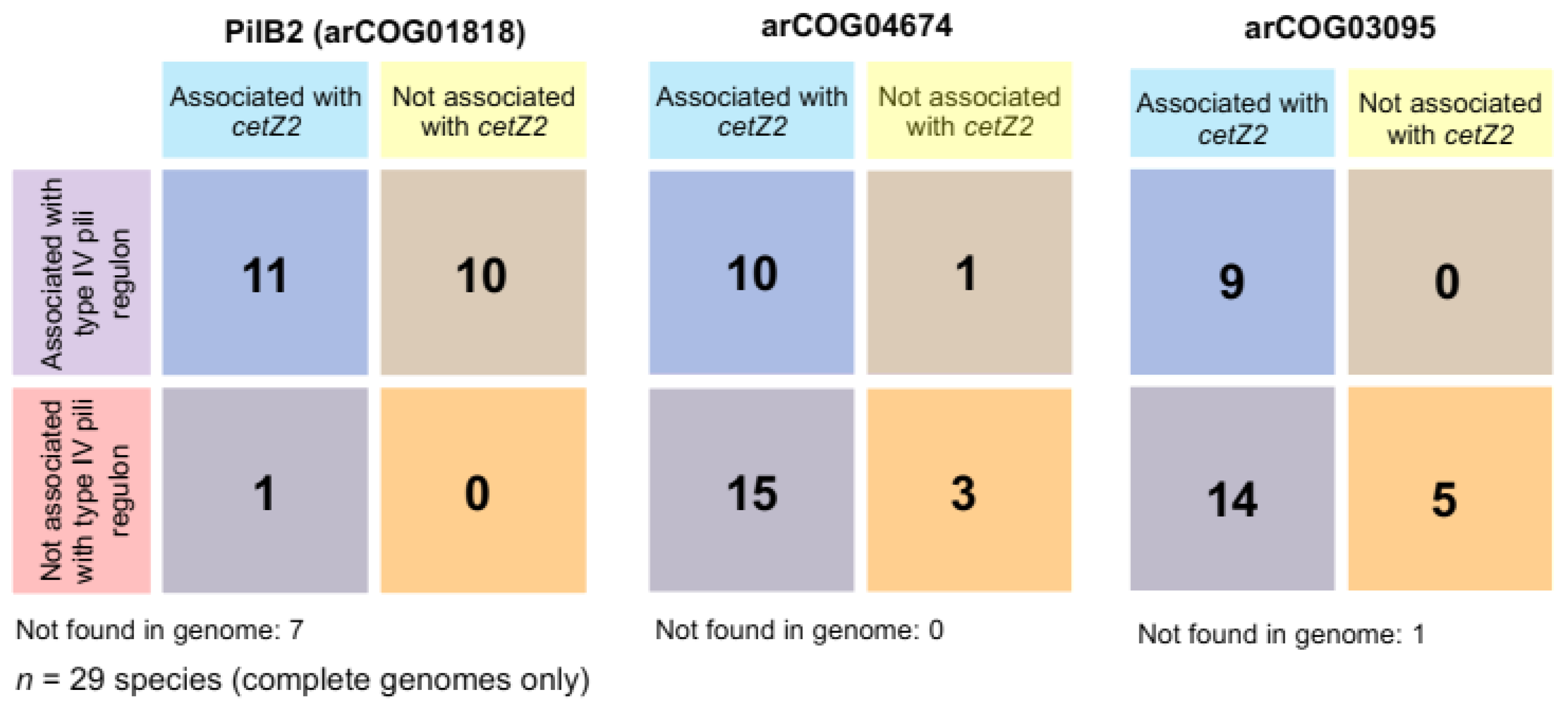
3.8. cetZ2 Is Associated with A Type IV Pili Regulon in Haloferacales and Halobacteriales
3.9. Non-Pili Associated Genes Conserved in cetZ2 Regions
3.10. The Genomic Environments of N. Pharaonis cetZ Genes
3.11. Synteny in cetZ Genomic Regions of Non-Halobacteria Euryarchaeota
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adams, D.W.; Errington, J. Bacterial cell division: Assembly, maintenance and disassembly of the Z ring. Nat. Rev. Genet. 2009, 7, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Barton, N.R.; Goldstein, L. Going mobile: Microtubule motors and chromosome segregation. Proc. Natl. Acad. Sci. USA 1996, 93, 1735–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcin, C.; Straube, A. Microtubules in cell migration. Essays Biochem. 2019, 63, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duggin, I.G.; Aylett, C.H.; Walsh, J.C.; Michie, K.A.; Wang, Q.; Turnbull, L.; Dawson, E.M.; Harry, E.J.; Whitchurch, C.B.; Amos, L.A. CetZ tubulin-like proteins control archaeal cell shape. Nature 2015, 519, 362–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vicente-Manzanares, M.; Choi, C.K.; Horwitz, A.R. Integrins in cell migration–the actin connection. J. Cell Sci. 2009, 122, 1473. [Google Scholar] [CrossRef] [Green Version]
- Kaverina, I.; Straube, A. Regulation of cell migration by dynamic microtubules. Semin. Cell Dev. Biol. 2011, 22, 968–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qualmann, B.; Kessels, M.M.; Kelly, R.B. Molecular links between endocytosis and the actin cytoskeleton. J. Cell Biol. 2000, 150, F111–F116. [Google Scholar] [CrossRef] [Green Version]
- Appert-Rolland, C.; Ebbinghaus, M.; Santen, L. Intracellular transport driven by cytoskeletal motors: General mechanisms and defects. Phys. Rep. 2015, 593, 1–59. [Google Scholar] [CrossRef] [Green Version]
- Hirokawa, N.; Noda, Y.; Tanaka, Y.; Niwa, S. Kinesin superfamily motor proteins and intracellular transport. Nat. Rev. Mol. Cell Biol. 2009, 10, 682–696. [Google Scholar] [CrossRef] [PubMed]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [PubMed]
- Ettema, T.J.; Lindås, A.C.; Bernander, R. An actin-based cytoskeleton in archaea. Mol. Microbiol. 2011, 80, 1052–1061. [Google Scholar] [CrossRef]
- Nogales, E.; Downing, K.H.; Amos, L.A.; Löwe, J. Tubulin and FtsZ form a distinct family of GTPases. Nat. Struct. Biol. 1998, 5, 451–458. [Google Scholar] [CrossRef]
- Desai, A.; Mitchison, T.J. Microtubule polymerization dynamics. Annu. Rev. Cell Dev. Biol. 1997, 13, 83–117. [Google Scholar] [CrossRef] [Green Version]
- Mitchison, T. Localization of an exchangeable GTP binding site at the plus end of microtubules. Science 1993, 261, 1044–1047. [Google Scholar] [CrossRef] [PubMed]
- Mitchison, T.; Kirschner, M. Dynamic instability of microtubule growth. Nature 1984, 312, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Nogales, E. Structural insights into microtubule function. Annu. Rev. Biochem. 2000, 30, 397–420. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, A.; Lutkenhaus, J. Guanine nucleotide-dependent assembly of FtsZ into filaments. J. Bacteriol. 1994, 176, 2754–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, A.; Lutkenhaus, J. Dynamic assembly of FtsZ regulated by GTP hydrolysis. EMBO J. 1998, 17, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Erickson, H.P.; Taylor, D.W.; Taylor, K.A.; Bramhill, D. Bacterial cell division protein FtsZ assembles into protofilament sheets and minirings, structural homologs of tubulin polymers. Proc. Natl. Acad. Sci. USA 1996, 93, 519–523. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, L.S.; Yang, Z. Microtubule-based transport systems in neurons: The roles of kinesins and dyneins. Annu. Rev. Neurosci. 2000, 23, 39–71. [Google Scholar] [CrossRef] [PubMed]
- Kulić, I.M.; Brown, A.E.; Kim, H.; Kural, C.; Blehm, B.; Selvin, P.R.; Nelson, P.C.; Gelfand, V.I. The role of microtubule movement in bidirectional organelle transport. Proc. Natl. Acad. Sci. USA 2008, 105, 10011–10016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, M.A.; Maxfield, F.R. Ca2+-and calcineurin-dependent recycling of an integrin to the front of migrating neutrophils. Nature 1995, 377, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Johnsson, A.-K.; Karlsson, R. Microtubule-dependent localization of profilin I mRNA to actin polymerization sites in serum-stimulated cells. Eur. J. Cell Biol. 2010, 89, 394–401. [Google Scholar] [CrossRef]
- Bergmann, J.E.; Kupfer, A.; Singer, S. Membrane insertion at the leading edge of motile fibroblasts. Proc. Natl. Acad. Sci. USA 1983, 80, 1367–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezratty, E.J.; Partridge, M.A.; Gundersen, G.G. Microtubule-induced focal adhesion disassembly is mediated by dynamin and focal adhesion kinase. Nat. Cell Biol. 2005, 7, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Laan, L.; Husson, J.; Munteanu, E.L.; Kerssemakers, J.W.; Dogterom, M. Force-generation and dynamic instability of microtubule bundles. Proc. Natl. Acad. Sci. USA 2008, 105, 8920–8925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoué, S.; Salmon, E.D. Force Generation by Microtubule Assembly/Disassembly in Mitosis and Related Movements. Mol. Biol. Cell 1995, 6, 1619–1640. [Google Scholar] [CrossRef]
- Liao, Y.; Ithurbide, S.; de Silva, R.T.; Erdmann, S.; Duggin, I.G. Archaeal cell biology: Diverse functions of tubulin-like cytoskeletal proteins at the cell envelope. Emerg. Top Life Sci. 2018, 2, 547–559. [Google Scholar] [CrossRef]
- Liao, Y.; Ithurbide, S.; Evenhuis, C.; Löwe, J.; Duggin, I.G. Cell division in the archaeon Haloferax volcanii relies on two FtsZ proteins with distinct functions in division ring assembly and constriction. Nat. Microbiol. 2021, 6, 594–605. [Google Scholar] [CrossRef]
- Bi, E.; Dai, K.; Subbarao, S.; Beall, B.; Lutkenhaus, J. FtsZ and cell division. Res. Microbiol. 1991, 142, 249–252. [Google Scholar] [CrossRef]
- Goehring, N.W.; Beckwith, J. Diverse paths to midcell: Assembly of the bacterial cell division machinery. Curr. Biol. 2005, 15, R514–R526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haeusser, D.P.; Margolin, W. Splitsville: Structural and functional insights into the dynamic bacterial Z ring. Nat. Rev. Genet. 2016, 14, 305–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Lyu, Z.; Miguel, A.; McQuillen, R.; Huang, K.C.; Xiao, J. GTPase activity–coupled treadmilling of the bacterial tubulin FtsZ organizes septal cell wall synthesis. Science 2017, 355, 744–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osawa, M.; Erickson, H.P. Liposome division by a simple bacterial division machinery. Proc. Natl. Acad. Sci. USA 2013, 110, 11000–11004. [Google Scholar] [CrossRef] [Green Version]
- Nußbaum, P.; Gerstner, M.; Dingethal, M.; Erb, C.; Albers, S.-V. The archaeal protein SepF is essential for cell division in Haloferax volcanii. Nat. Commun. 2021, 12, 3469. [Google Scholar] [CrossRef]
- Liao, Y.; Vogel, V.; Hauber, S.; Bartel, J.; Alkhnbashi, O.S.; Maaß, S.; Schwarz, T.S.; Backofen, R.; Becher, D.; Duggin, I.G. CdrS is a global transcriptional regulator influencing cell division in Haloferax volcanii. Mbio 2021, 12, e0141621. [Google Scholar] [CrossRef]
- de Silva, R.T.; Abdul-Halim, M.F.; Pittrich, D.A.; Brown, H.J.; Pohlschroder, M.; Duggin, I.G. Improved growth and morphological plasticity of Haloferax volcanii. Microbiology 2021, 167, 001012. [Google Scholar] [CrossRef]
- Mullakhanbhai, M.F.; Larsen, H. Halobacterium volcanii spec. nov., a Dead Sea halobacterium with a moderate salt requirement. Arch. Microbiol. 1975, 104, 207–214. [Google Scholar] [CrossRef]
- Li, Z.; Kinosita, Y.; Rodriguez-Franco, M.; Nußbaum, P.; Braun, F.; Delpech, F.; Quax, T.E.; Albers, S.-V. Positioning of the motility machinery in halophilic archaea. Mbio 2019, 10, e00377-19. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.; Potter, S.C.; Finn, R.D. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, D439–D444. [Google Scholar] [CrossRef]
- The PyMOL Molecular Graphics System, version 1.8; Schrödinger, LLC: New York, NY, USA, 2015.
- Jurrus, E.; Engel, D.; Star, K.; Monson, K.; Brandi, J.; Felberg, L.E.; Brookes, D.H.; Wilson, L.; Chen, J.; Liles, K. Improvements to the APBS biomolecular solvation software suite. Protein Sci. 2018, 27, 112–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantalapiedra, C.P.; Hernández-Plaza, A.; Letunic, I.; Bork, P.; Huerta-Cepas, J. eggNOG-mapper v2: Functional Annotation, Orthology Assignments, and Domain Prediction at the Metagenomic Scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [Green Version]
- Aylett, C.H.; Duggin, I.G. The tubulin superfamily in archaea. Subcell. Biochem. 2017, 84, 393–417. [Google Scholar] [CrossRef] [PubMed]
- Kocur, M.; Hodgkiss, W. Taxonomic status of the genus Halococcus Schoop. Int. J. Syst. Evol. Microbiol. 1973, 23, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Montero, C.G.; Ventosa, A.; Rodriguez-Valera, F.; Kates, M.; Moldoveanu, N.; Ruiz-Berraquero, F. Halococcus saccharolyticus sp. nov., a new species of extremely halophilic non-alkaliphilic cocci. Syst. Appl. Microbiol. 1989, 12, 167–171. [Google Scholar] [CrossRef]
- Gonzalez, O.; Oberwinkler, T.; Mansueto, L.; Pfeiffer, F.; Mendoza, E.; Zimmer, R.; Oesterhelt, D. Characterization of growth and metabolism of the haloalkaliphile Natronomonas pharaonis. PLoS Comput. Biol. 2010, 6, e1000799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Koonin, E.V.; Kelman, Z. The CMG (CDC45/RecJ, MCM, GINS) complex is a conserved component of the DNA replication system in all archaea and eukaryotes. Biol. Direct 2012, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Archaeal clusters of orthologous genes (arCOGs): An update and application for analysis of shared features between Thermococcales, Methanococcales, and Methanobacteriales. Life 2015, 5, 818–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Rueda, E.; Janga, S.C. Identification and genomic analysis of transcription factors in archaeal genomes exemplifies their functional architecture and evolutionary origin. Mol. Biol. Evol. 2010, 27, 1449–1459. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, L.; Maklad, H.R.; Bervoets, I.; Peeters, E. Transcription regulators in archaea: Homologies and differences with bacterial regulators. J. Mol. Biol. 2019, 431, 4132–4146. [Google Scholar] [CrossRef] [PubMed]
- Schwaiger, R.; Schwarz, C.; Furtwängler, K.; Tarasov, V.; Wende, A.; Oesterhelt, D. Transcriptional control by two leucine-responsive regulatory proteins in Halobacterium salinarum R1. BMC Mol. Biol. 2010, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Ettema, T.J.; Brinkman, A.B.; Tani, T.H.; Rafferty, J.B.; Van Der Oost, J. A novel ligand-binding domain involved in regulation of amino acid metabolism in prokaryotes. J. Biol. Chem. 2002, 277, 37464–37468. [Google Scholar] [CrossRef] [Green Version]
- Peeters, E.; Albers, S.V.; Vassart, A.; Driessen, A.J.; Charlier, D. Ss-LrpB, a transcriptional regulator from Sulfolobus solfataricus, regulates a gene cluster with a pyruvate ferredoxin oxidoreductase-encoding operon and permease genes. Mol. Microbiol. 2009, 71, 972–988. [Google Scholar] [CrossRef] [Green Version]
- Islam, R.; Brown, S.; Taheri, A.; Dumenyo, C.K. The gene encoding NAD-dependent epimerase/dehydratase, wcaG, affects cell surface properties, virulence, and extracellular enzyme production in the soft rot phytopathogen, Pectobacterium carotovorum. Microorganisms 2019, 7, 172. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Koonin, E.V.; Albers, S.-V. Diversity and evolution of type IV pili systems in archaea. Front. Microbiol. 2016, 7, 667. [Google Scholar] [CrossRef]
- Jevtić, Ž.; Stoll, B.; Pfeiffer, F.; Sharma, K.; Urlaub, H.; Marchfelder, A.; Lenz, C. The Response of Haloferax volcanii to Salt and Temperature Stress: A Proteome Study by Label-Free Mass Spectrometry. Proteomics 2019, 19, 1800491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.M.; Hoopes, S.L.; White, R.H.; Sarisky, C.A. Purine biosynthesis in archaea: Variations on a theme. Biol. Direct 2011, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, S.; Deng, Y.; Tomsho, J.W.; Kyoung, M.; Benkovic, S.J. Microtubule-assisted mechanism for functional metabolic macromolecular complex formation. Proc. Natl. Acad. Sci. USA 2010, 107, 12872–12876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinter, R.; Greening, C. Cofactor F420: An expanded view of its distribution, biosynthesis and roles in bacteria and archaea. FEMS Microbiol. Rev. 2021, 45, fuab021. [Google Scholar] [CrossRef] [PubMed]
- Cassimeris, L.; Silva, V.C.; Miller, E.; Ton, Q.; Molnar, C.; Fong, J. Fueled by microtubules: Does tubulin dimer/polymer partitioning regulate intracellular metabolism? Cytoskeleton 2012, 69, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Szwedziak, P.; Wang, Q.; Bharat, T.A.; Tsim, M.; Löwe, J. Architecture of the ring formed by the tubulin homologue FtsZ in bacterial cell division. Elife 2014, 3, e04601. [Google Scholar] [CrossRef]
- Abdul Halim, M.F.; Schulze, S.; DiLucido, A.; Pfeiffer, F.; Bisson Filho, A.W.; Pohlschroder, M. Lipid Anchoring of Archaeosortase Substrates and Midcell Growth in Haloarchaea. Mbio 2020, 11, e00349-20. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brown, H.J.; Duggin, I.G. Diversity and Potential Multifunctionality of Archaeal CetZ Tubulin-like Cytoskeletal Proteins. Biomolecules 2023, 13, 134. https://doi.org/10.3390/biom13010134
Brown HJ, Duggin IG. Diversity and Potential Multifunctionality of Archaeal CetZ Tubulin-like Cytoskeletal Proteins. Biomolecules. 2023; 13(1):134. https://doi.org/10.3390/biom13010134
Chicago/Turabian StyleBrown, Hannah J., and Iain G. Duggin. 2023. "Diversity and Potential Multifunctionality of Archaeal CetZ Tubulin-like Cytoskeletal Proteins" Biomolecules 13, no. 1: 134. https://doi.org/10.3390/biom13010134