
The Role of Pyroptosis and Autophagy in Ischemia Reperfusion Injury


Abstract
:1. Introduction
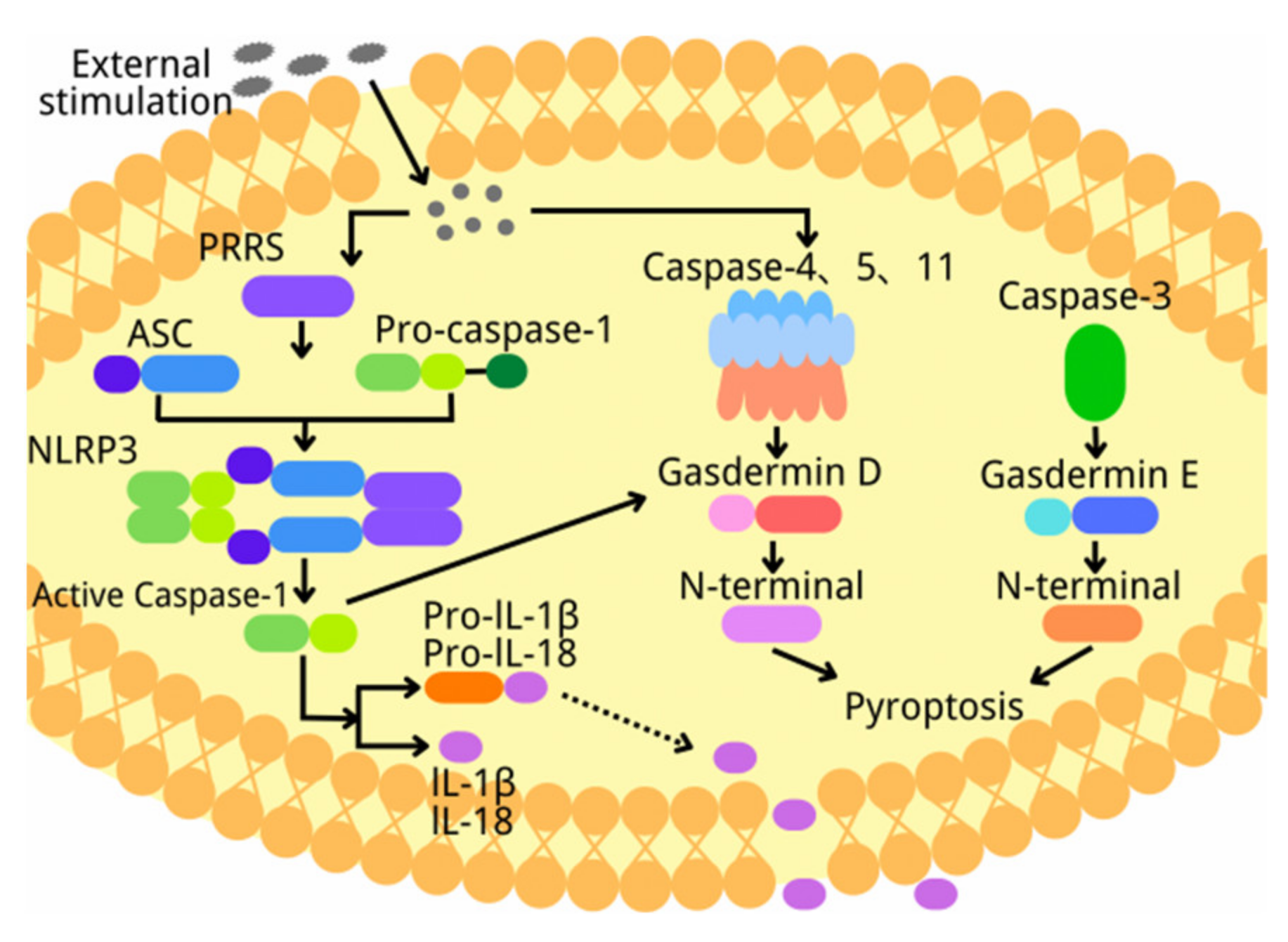
2. Overview of Pyroptosis
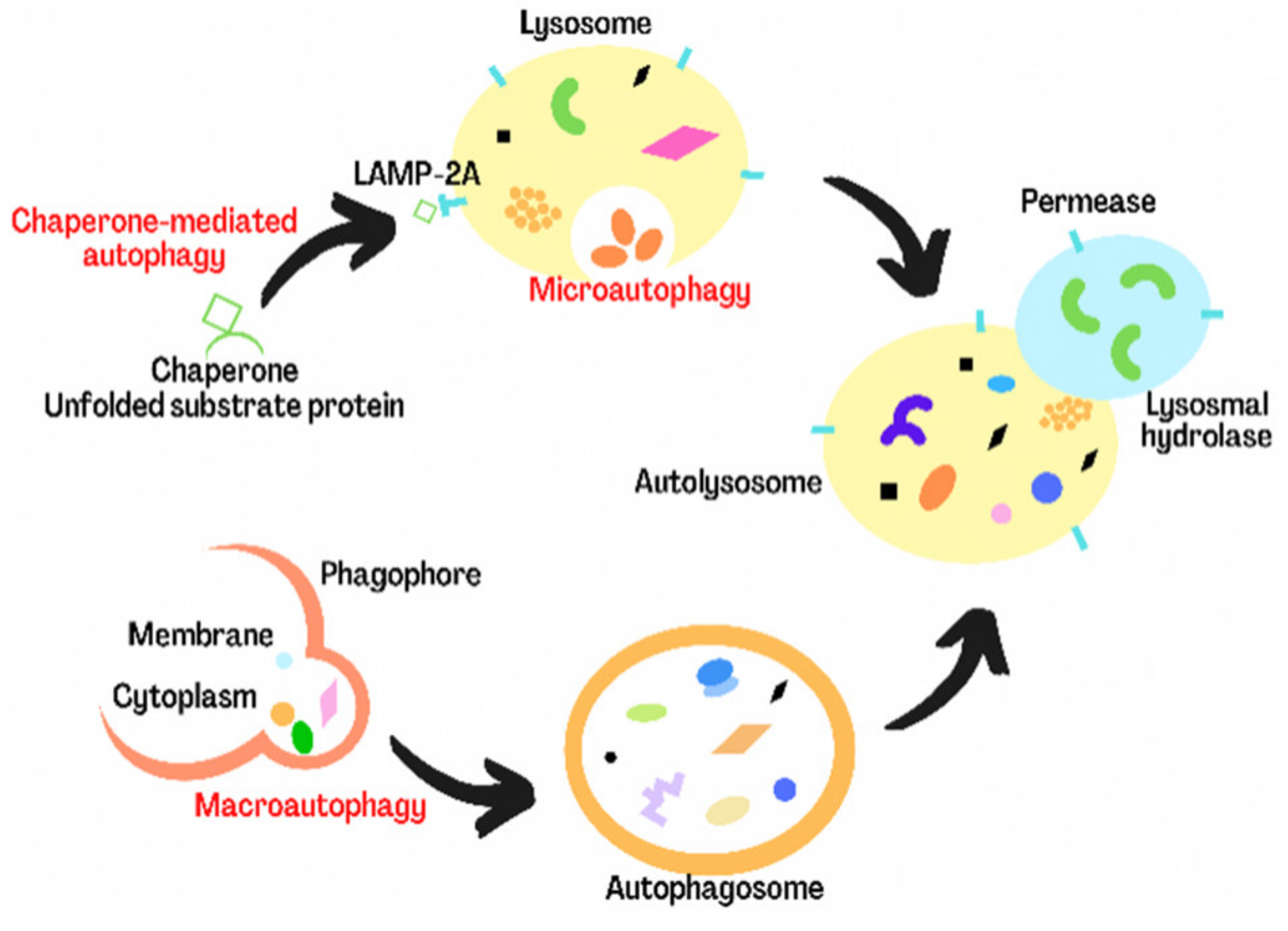
3. Overview of Autophagy
4. The Role of Pyroptosis and Autophagy in Ischemia/Reperfusion Injury of Nervous System
4.1. The Role of Pyroptosis and Autophagy in Cerebral Ischemia/Reperfusion Injury
4.2. The Role of Pyroptosis and Autophagy in Spinal Cord Ischemia Reperfusion Injury
4.3. The Role of Pyroptosis and Autophagy in Microglial with Oxygen-Glucose Deprivation/Reoxygenation
5. The Role of Pyroptosis and Autophagy in Myocardial Ischemia and Reperfusion Injury
6. The Role of Pyroptosis and Autophagy in Limb Ischemia/Reperfusion-Induced Muscular Injury
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Jiang, M.; Qi, L.; Wu, Y.; Song, D.; Gan, J.; Li, Y.; Bai, Y. Pyroptosis, a new bridge to tumor immunity. Cancer Sci. 2021, 112, 3979–3994. [Google Scholar] [CrossRef] [PubMed]
- Loveless, R.; Bloomquist, R.; Teng, Y. Pyroptosis at the forefront of anticancer immunity. J. Exp. Clin. Cancer Res. 2021, 40, 264. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.K.; Li, C.Y.; Lin, I.L.; Syue, W.J.; Chen, Y.F.; Cheng, K.C.; Teng, Y.N.; Lin, Y.H.; Yen, C.H.; Chiu, C.C. Inflammation-related pyroptosis, a novel programmed cell death pathway, and its crosstalk with immune therapy in cancer treatment. Theranostics 2021, 11, 8813–8835. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhao, Y.; Wang, K.; Shi, X.; Wang, Y.; Huang, H.; Zhuang, Y.; Cai, T.; Wang, F.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Shi, J.; Gao, W.; Shao, F. Pyroptosis: Gasdermin-Mediated Programmed Necrotic Cell Death. Trends Biochem. Sci. 2017, 42, 245–254. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Ruan, J.; Pan, Y.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Liu, H.; Yang, Y.; Wang, H. The Role of Autophagy and Pyroptosis in Liver Disorders. Int. J. Mol. Sci. 2022, 23, 6208. [Google Scholar] [CrossRef]
- Kitada, M.; Koya, D. Autophagy in metabolic disease and ageing. Nat. Rev. Endocrinol. 2021, 17, 647–661. [Google Scholar] [CrossRef]
- Ascenzi, F.; De Vitis, C.; Maugeri-Sacca, M.; Napoli, C.; Ciliberto, G.; Mancini, R. SCD1, autophagy and cancer: Implications for therapy. J. Exp. Clin. Cancer Res. 2021, 40, 265. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Petroni, G.; Amaravadi, R.K.; Baehrecke, E.H.; Ballabio, A.; Boya, P.; Bravo-San Pedro, J.M.; Cadwell, K.; Cecconi, F.; Choi, A.M.K.; et al. Autophagy in major human diseases. EMBO J. 2021, 40, e108863. [Google Scholar] [CrossRef]
- Lin, L.; Zhang, M.X.; Zhang, L.; Zhang, D.; Li, C.; Li, Y.L. Autophagy, Pyroptosis, and Ferroptosis: New Regulatory Mechanisms for Atherosclerosis. Front. Cell Dev. Biol. 2021, 9, 809955. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Wang, H.; Cui, N. Autophagy Regulation on Pyroptosis: Mechanism and Medical Implication in Sepsis. Mediat. Inflamm. 2021, 2021, 9925059. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.Y.; Peng, S.Y.; Ma, M.; Li, H.Y.; Wang, Z.; Chen, G. Protective effects of sevoflurane in cerebral ischemia reperfusion injury: A narrative review. Med. Gas. Res. 2021, 11, 152–154. [Google Scholar] [PubMed]
- Wu, M.Y.; Yiang, G.T.; Liao, W.T.; Tsai, A.P.; Cheng, Y.L.; Cheng, P.W.; Li, C.Y.; Li, C.J. Current Mechanistic Concepts in Ischemia and Reperfusion Injury. Cell Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.J.; Wu, Q.; Jiang, S.M.; Ding, L.; Liu, C.X.; Xu, M.; Wang, Y.; Zhou, Y.; Li, L. Pyroptosis: A New Frontier in Kidney Diseases. Oxid. Med. Cell Longev. 2021, 2021, 6686617. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, Y.; Li, R.; Zhu, L.; Fu, B.; Yan, T. Salidroside ameliorates Parkinson’s disease by inhibiting NLRP3-dependent pyroptosis. Aging 2020, 12, 9405–9426. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Zhou, B.; Sun, R.Y.; Ai, Y.L.; Cheng, K.; Li, F.N.; Wang, B.R.; Liu, F.J.; Jiang, Z.H.; Wang, W.J.; et al. The metabolite alpha-KG induces GSDMC-dependent pyroptosis through death receptor 6-activated caspase-8. Cell Res. 2021, 31, 980–997. [Google Scholar] [CrossRef]
- Zheng, X.; Zhong, T.; Ma, Y.; Wan, X.; Qin, A.; Yao, B.; Zou, H.; Song, Y.; Yin, D. Bnip3 mediates doxorubicin-induced cardiomyocyte pyroptosis via caspase-3/GSDME. Life Sci. 2020, 242, 117186. [Google Scholar] [CrossRef]
- Ji, N.; Qi, Z.; Wang, Y.; Yang, X.; Yan, Z.; Li, M.; Ge, Q.; Zhang, J. Pyroptosis: A New Regulating Mechanism in Cardiovascular Disease. J. Inflamm. Res. 2021, 14, 2647–2666. [Google Scholar] [CrossRef]
- Li, Q.; Shi, N.; Cai, C.; Zhang, M.; He, J.; Tan, Y.; Fu, W. The Role of Mitochondria in Pyroptosis. Front. Cell Dev. Biol. 2020, 8, 630771. [Google Scholar] [CrossRef]
- Tan, Y.; Chen, Q.; Li, X.; Zeng, Z.; Xiong, W.; Li, G.; Li, X.; Yang, J.; Xiang, B.; Yi, M. Correction to: Pyroptosis: A new paradigm of cell death for fighting against cancer. J. Exp. Clin. Cancer Res. 2021, 40, 219. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Zhang, X.; Liu, N.; Tang, L.; Peng, C.; Chen, X. Pyroptosis: Mechanisms and diseases. Signal Transduct. Target. Ther. 2021, 6, 128. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yin, B.; Li, D.; Wang, G.; Han, X.; Sun, X. GSDME mediates caspase-3-dependent pyroptosis in gastric cancer. Biochem. Biophys. Res. Commun. 2018, 495, 1418–1425. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, H.; Li, D.; Zhou, X.; Qin, Q.; Zhang, Q. Caspase-3-mediated GSDME induced Pyroptosis in breast cancer cells through the ROS/JNK signalling pathway. J. Cell. Mol. Med. 2021, 25, 8159–8168. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.; Liu, H.; Zhu, R.; He, H.; Zhou, Y.; Zhang, Y.; Li, C.; Liang, D.; Zeng, Q.; et al. Bone marrow mesenchymal stem cell-derived exosomes attenuate cerebral ischemia-reperfusion injury-induced neuroinflammation and pyroptosis by modulating microglia M1/M2 phenotypes. Exp. Neurol. 2021, 341, 113700. [Google Scholar] [CrossRef]
- Shi, H.; Gao, Y.; Dong, Z.; Yang, J.; Gao, R.; Li, X.; Zhang, S.; Ma, L.; Sun, X.; Wang, Z.; et al. GSDMD-Mediated Cardiomyocyte Pyroptosis Promotes Myocardial I/R Injury. Circ. Res. 2021, 129, 383–396. [Google Scholar] [CrossRef]
- Ni, J.; Jiang, L.; Shen, G.; Xia, Z.; Zhang, L.; Xu, J.; Feng, Q.; Qu, H.; Xu, F.; Li, X. Hydrogen sulfide reduces pyroptosis and alleviates ischemia-reperfusion-induced acute kidney injury by inhibiting NLRP3 inflammasome. Life Sci. 2021, 284, 119466. [Google Scholar] [CrossRef]
- Lizama, B.N.; Chu, C.T. Neuronal autophagy and mitophagy in Parkinson’s disease. Mol. Aspects Med. 2021, 82, 100972. [Google Scholar] [CrossRef]
- Li, Y.J.; Lei, Y.H.; Yao, N.; Wang, C.R.; Hu, N.; Ye, W.C.; Zhang, D.M.; Chen, Z.S. Autophagy and multidrug resistance in cancer. Chin. J. Cancer 2017, 36, 52. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An overview of autophagy: Morphology, mechanism, and regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wu, D.; Wang, H. Hydrogen sulfide plays an important protective role by influencing autophagy in diseases. Physiol. Res. 2019, 68, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Antunes, F.; Erustes, A.G.; Costa, A.J.; Nascimento, A.C.; Bincoletto, C.; Ureshino, R.P.; Pereira, G.J.S.; Smaili, S.S. Autophagy and intermittent fasting: The connection for cancer therapy? Clinics 2018, 73 (Suppl. S1), e814s. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Levine, B. Autosis and autophagic cell death: The dark side of autophagy. Cell Death Differ. 2015, 22, 367–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Huerta, P.; Troncoso-Escudero, P.; Jerez, C.; Hetz, C.; Vidal, R.L. The intersection between growth factors, autophagy and ER stress: A new target to treat neurodegenerative diseases? Brain Res. 2016, 1649, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Wang, Z.; Wang, J.; Wang, H. Exogenous Hydrogen Sulfide Plays an Important Role Through Regulating Autophagy in Ischemia/Reperfusion Injury. Front. Mol. Biosci. 2021, 8, 681676. [Google Scholar] [CrossRef]
- Rong, Y.; Fan, J.; Ji, C.; Wang, Z.; Ge, X.; Wang, J.; Ye, W.; Yin, G.; Cai, W.; Liu, W. USP11 regulates autophagy-dependent ferroptosis after spinal cord ischemia-reperfusion injury by deubiquitinating Beclin 1. Cell Death Differ. 2022, 29, 1164–1175. [Google Scholar] [CrossRef]
- Lv, S.; Liu, H.; Wang, H. The Interplay between Autophagy and NLRP3 Inflammasome in Ischemia/Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 8773. [Google Scholar] [CrossRef]
- Wicha, P.; Das, S.; Mahakkanukrauh, P. Blood-brain barrier dysfunction in ischemic stroke and diabetes: The underlying link, mechanisms and future possible therapeutic targets. Anat. Cell Biol. 2021, 54, 165–177. [Google Scholar] [CrossRef]
- Li, X.; Sun, B.; Wang, L.; Zhang, J.; Zhang, J.; Zhao, Z.; Wu, H.; Liu, X.; Zhou, Y.; Mossa-Basha, M.; et al. Association of Type 2 Diabetes Mellitus and Glycemic Control with Intracranial Plaque Characteristics in Patients With Acute Ischemic Stroke. J. Magn. Reson. Imaging 2021, 54, 655–666. [Google Scholar] [CrossRef]
- Wolf, V.; Abdul, Y.; Li, W.; Ergul, A. Impact of diabetes and ischemic stroke on the cerebrovasculature: A female perspective. Neurobiol. Dis. 2022, 167, 105667. [Google Scholar] [CrossRef]
- Muranyi, M.; Lacza, Z. Influence of Diabetes Mellitus on Cerebral Ischemia and Reperfusion Injury. Ph.D. Thesis, Semmelweis University, Budapest, Hungary, 2006. [Google Scholar]
- Liu, L.; Cao, Q.; Gao, W.; Li, B.Y.; Zeng, C.; Xia, Z.; Zhao, B. Melatonin ameliorates cerebral ischemia-reperfusion injury in diabetic mice by enhancing autophagy via the SIRT1-BMAL1 pathway. FASEB J. 2021, 35, e22040. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.; Chen, B.; Wang, X.; Gao, C.; Yu, H. Icariin enhance mild hypothermia-induced neuroprotection via inhibiting the activation of NF-kappaB in experimental ischemic stroke. Metab. Brain Dis. 2021, 36, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Omileke, D.; Azarpeykan, S.; Bothwell, S.W.; Pepperall, D.; Beard, D.J.; Coupland, K.; Patabendige, A.; Spratt, N.J. Short-duration hypothermia completed prior to reperfusion prevents intracranial pressure elevation following ischaemic stroke in rats. Sci. Rep. 2021, 11, 22354. [Google Scholar] [CrossRef]
- Tu, Y.; Guo, C.; Song, F.; Huo, Y.; Geng, Y.; Guo, M.; Bao, H.; Wu, X.; Fan, W. Mild hypothermia alleviates diabetes aggravated cerebral ischemic injury via activating autophagy and inhibiting pyroptosis. Brain Res. Bull. 2019, 150, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wen, Y.; Zhou, B.; Tian, Y.; Ning, Y.; Ding, H. Incomplete autophagy promotes the replication of Mycoplasma hyopneumoniae. J. Microbiol. 2021, 59, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.D.; Sheng, R.; Zhang, L.S.; Han, R.; Zhang, X.; Zhang, X.D.; Han, F.; Fukunaga, K.; Qin, Z.H. Neuronal injury in rat model of permanent focal cerebral ischemia is associated with activation of autophagic and lysosomal pathways. Autophagy 2008, 4, 762–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xia, H.; Kim, M.; Xu, L.; Li, Y.; Zhang, L.; Cai, Y.; Norberg, H.V.; Zhang, T.; Furuya, T.; et al. Beclin1 controls the levels of p53 by regulating the deubiquitination activity of USP10 and USP13. Cell 2011, 147, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Jin, S.; Wu, Y.; Xian, H.; Tian, S.; Liu, D.A.; Guo, Z.; Cui, J. Auto-ubiquitination of NEDD4-1 Recruits USP13 to Facilitate Autophagy through Deubiquitinating VPS34. Cell Rep. 2020, 30, 2807–2819.e4. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Zhang, J.; Liang, L.; Liu, N.; Qi, M.; Zhao, S.; Su, J.; Liu, J.; Peng, C.; Chen, X.; et al. Potent USP10/13 antagonist spautin-1 suppresses melanoma growth via ROS-mediated DNA damage and exhibits synergy with cisplatin. J. Cell. Mol. Med. 2020, 24, 4324–4340. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, S.; Jiao, W.; Zhong, Z.; Yang, Y.; Claret, F.X.; Elkabets, M.; Wang, F.; Wang, R.; Zhong, Y.; et al. Hydroxychloroquine synergizes with the PI3K inhibitor BKM120 to exhibit antitumor efficacy independent of autophagy. J. Exp. Clin. Cancer Res. 2021, 40, 374. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, Z.; Wu, T.; Zhang, Q.; Lu, F.; Gu, J.; Jiang, T.; Xue, J. Inhibition of autophagy-dependent pyroptosis attenuates cerebral ischaemia/reperfusion injury. J. Cell. Mol. Med. 2021, 25, 5060–5069. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Jiang, L.; Li, Q.; Liu, X.; Zhang, T.; Dong, L.; Liu, T.; Liu, L.; Hu, G.; Sun, X.; et al. Acrolein induces NLRP3 inflammasome-mediated pyroptosis and suppresses migration via ROS-dependent autophagy in vascular endothelial cells. Toxicology 2018, 410, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Zhu, X.L.; Zhao, B.X.; Shi, L.; Wang, W.; Hu, W.; Qin, S.L.; Chen, B.H.; Zhou, P.H.; Qiu, B.; et al. Adrenomedullin alleviates the pyroptosis of Leydig cells by promoting autophagy via the ROS-AMPK-mTOR axis. Cell Death Dis. 2019, 10, 489. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Li, L.; Huang, Y.; Qian, D.; Liu, W.; Zhang, C.; Luo, Y.; Zhou, Z.; Kong, F.; Zhao, X.; et al. Salidroside Ameliorates Mitochondria-Dependent Neuronal Apoptosis after Spinal Cord Ischemia-Reperfusion Injury Partially through Inhibiting Oxidative Stress and Promoting Mitophagy. Oxid. Med. Cell Longev. 2020, 2020, 3549704. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, B.; Shi, E. Perillaldehyde Alleviates Spinal Cord Ischemia-Reperfusion Injury Via Activating the Nrf2 Pathway. J. Surg. Res. 2021, 268, 308–317. [Google Scholar] [CrossRef]
- Qian, X.; Wu, Y.H.; Che, Y.Y.; Zhao, W.; Shu, L.F.; Zhu, J.; Wang, Y.H.; Chen, T. IP3R-mediated activation of BK channels contributes to mGluR5-induced protection against spinal cord ischemia-reperfusion injury. Neurochem. Int. 2021, 150, 105191. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, K.; Eldeiry, M.; Aftab, M.; Ryan, T.J.; Roda, G.; Meng, X.; Weyant, M.J.; Cleveland, J.C., Jr.; Fullerton, D.A.; Reece, T.B. Pretreatment with Diazoxide Attenuates Spinal Cord Ischemia-Reperfusion Injury Through Signaling Transducer and Activator of Transcription 3 Pathway. Ann. Thorac. Surg. 2019, 107, 733–739. [Google Scholar] [CrossRef]
- Li, H.; Dong, X.; Yang, Y.; Jin, M.; Cheng, W. The Neuroprotective Mechanism of Spinal Cord Stimulation in Spinal Cord Ischemia/Reperfusion Injury. Neuromodulation 2021, 24, 43–48. [Google Scholar] [CrossRef]
- Yu, M.; Qi, B.; Xiaoxiang, W.; Xu, J.; Liu, X. Baicalein increases cisplatin sensitivity of A549 lung adenocarcinoma cells via PI3K/Akt/NF-kappaB pathway. Biomed. Pharmacother. 2017, 90, 677–685. [Google Scholar] [CrossRef]
- Yan, W.; Ma, X.; Zhao, X.; Zhang, S. Baicalein induces apoptosis and autophagy of breast cancer cells via inhibiting PI3K/AKT pathway in vivo and vitro. Drug Des. Devel. Ther. 2018, 12, 3961–3972. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Tan, Z.; Huang, H.; Zeng, Y.; Chen, S.; Wei, J.; Huang, G.; Qian, C.; Yuan, G.; He, S. Baicalein pre-treatment alleviates hepatic ischemia/reperfusion injury in mice by regulating the Nrf2/ARE pathway. Exp. Ther. Med. 2021, 22, 1380. [Google Scholar] [CrossRef]
- Yang, S.; Wang, H.; Yang, Y.; Wang, R.; Wang, Y.; Wu, C.; Du, G. Baicalein administered in the subacute phase ameliorates ischemia-reperfusion-induced brain injury by reducing neuroinflammation and neuronal damage. Biomed. Pharmacother. 2019, 117, 109102. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Xu, H.; Li, J.; Hu, X.; Wang, X.; Huang, Y.; Li, Y.; Sheng, S.; Wang, Y.; Xu, H.; et al. Baicalein Attenuates Pyroptosis and Endoplasmic Reticulum Stress Following Spinal Cord Ischemia-Reperfusion Injury via Autophagy Enhancement. Front. Pharmacol. 2020, 11, 1076. [Google Scholar] [CrossRef]
- Zhang, Q.; He, X.; Yu, Q.; Liu, Y.; Zhu, Y.; Yang, L.; Shang, P.; Zhang, J.; Liu, H.; Feng, F. Endoplasmic reticulum stress regulates pyroptosis in BPDE-induced BEAS-2B cells. Environ. Toxicol. 2022, 37, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Hu, Z.; Li, Q.; Li, H.; Chen, W.; Huo, H.; Han, Q.; Zhang, H.; Guo, J.; Hu, L.; et al. Endoplasmic Reticulum Stress Contributes to Copper-Induced Pyroptosis via Regulating the IRE1alpha-XBP1 Pathway in Pig Jejunal Epithelial Cells. J. Agric. Food Chem. 2022, 70, 1293–1303. [Google Scholar] [CrossRef]
- Simard, J.C.; Vallieres, F.; de Liz, R.; Lavastre, V.; Girard, D. Silver nanoparticles induce degradation of the endoplasmic reticulum stress sensor activating transcription factor-6 leading to activation of the NLRP-3 inflammasome. J. Biol. Chem. 2015, 290, 5926–5939. [Google Scholar] [CrossRef] [Green Version]
- Ezquerro, S.; Mocha, F.; Fruhbeck, G.; Guzman-Ruiz, R.; Valenti, V.; Mugueta, C.; Becerril, S.; Catalan, V.; Gomez-Ambrosi, J.; Silva, C.; et al. Ghrelin Reduces TNF-alpha-Induced Human Hepatocyte Apoptosis, Autophagy, and Pyroptosis: Role in Obesity-Associated NAFLD. J. Clin. Endocrinol. Metab. 2019, 104, 21–37. [Google Scholar]
- Meng, Z.; Liu, H.; Zhang, J.; Zheng, Z.; Wang, Z.; Zhang, L.; Jia, Z.; Sui, Y. Sesamin promotes apoptosis and pyroptosis via autophagy to enhance antitumour effects on murine T-cell lymphoma. J. Pharmacol. Sci. 2021, 147, 260–270. [Google Scholar] [CrossRef]
- Tetorou, K.; Sisa, C.; Iqbal, A.; Dhillon, K.; Hristova, M. Current Therapies for Neonatal Hypoxic-Ischaemic and Infection-Sensitised Hypoxic-Ischaemic Brain Damage. Front. Synaptic Neurosci. 2021, 13, 709301. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, Z.; Wang, H. Downregulation of lncRNA GAS5 prevents mitochondrial apoptosis and hypoxic-ischemic brain damage in neonatal rats through the microRNA-128-3p/Bax/Akt/GSK-3beta axis. Neuroreport 2021, 32, 1395–1402. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, T.; Wang, P.; Shen, Y.; Huang, T. Overexpression of Long Noncoding RNA H19 Inhibits Cardiomyocyte Apoptosis in Neonatal Rats with Hypoxic-Ischemic Brain Damage Through the miR-149-5p/LIF/PI3K/Akt Axis. Biopreserv. Biobank. 2021, 19, 376–385. [Google Scholar] [CrossRef]
- Li, H.; Qi, J.; Wei, J.; Xu, B.; Min, S.; Wang, L.; Si, Y.; Qiu, H. Long non-coding RNA ANRIL mitigates neonatal hypoxic-ischemic brain damage via targeting the miR-378b/ATG3 axis. Am. J. Transl. Res. 2021, 13, 11585–11596. [Google Scholar] [PubMed]
- Shu, J.; Jiang, L.; Wang, M.; Wang, R.; Wang, X.; Gao, C.; Xia, Z. Human bone marrow mesenchymal stem cells-derived exosomes protect against nerve injury via regulating immune microenvironment in neonatal hypoxic-ischemic brain damage model. Immunobiology 2022, 227, 152178. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yuan, Y.; Zhang, X.; Lu, Y.; Dong, N.; Jiang, X.; Xu, J.; Zheng, D. Human Umbilical Cord Mesenchymal Stem Cell-Derived Exosomes Attenuate Oxygen-Glucose Deprivation/Reperfusion-Induced Microglial Pyroptosis by Promoting FOXO3a-Dependent Mitophagy. Oxid. Med. Cell. Longev. 2021, 2021, 6219715. [Google Scholar] [CrossRef]
- Sun, K.; Jing, X.; Guo, J.; Yao, X.; Guo, F. Mitophagy in degenerative joint diseases. Autophagy 2021, 17, 2082–2092. [Google Scholar] [CrossRef] [PubMed]
- Duffner, P.K.; Cohen, M.E.; Parker, M.S. Prospective intellectual testing in children with brain tumors. Ann. Neurol. 1988, 23, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Mitrovsky, G.; Vasanthi, H.R.; Das, D.K. Antiaging properties of a grape-derived antioxidant are regulated by mitochondrial balance of fusion and fission leading to mitophagy triggered by a signaling network of Sirt1-Sirt3-Foxo3-PINK1-PARKIN. Oxid. Med. Cell. Longev. 2014, 2014, 345105. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wu, B.; Zhong, B.; Lin, L.; Ding, Y.; Jin, X.; Huang, Z.; Lin, M.; Wu, H.; Xu, D. Naringenin alleviates myocardial ischemia/reperfusion injury by regulating the nuclear factor-erythroid factor 2-related factor 2 (Nrf2) /System xc-/ glutathione peroxidase 4 (GPX4) axis to inhibit ferroptosis. Bioengineered 2021, 12, 10924–10934. [Google Scholar] [CrossRef]
- Casin, K.M.; Calvert, J.W. Dynamic Regulation of Cysteine Oxidation and Phosphorylation in Myocardial Ischemia-Reperfusion Injury. Cells 2021, 10, 2388. [Google Scholar] [CrossRef]
- Du, A.; Xie, Y.; Ouyang, H.; Lu, B.; Jia, W.; Xu, H.; Ji, L. Si-Miao-Yong-An Decoction for Diabetic Retinopathy: A Combined Network Pharmacological and In Vivo Approach. Front. Pharmacol. 2021, 12, 763163. [Google Scholar] [CrossRef]
- Ren, Y.; Chen, X.; Li, P.; Zhang, H.; Su, C.; Zeng, Z.; Wu, Y.; Xie, X.; Wang, Q.; Han, J.; et al. Si-Miao-Yong-An decoction ameliorates cardiac function through restoring the equilibrium of SOD and NOX2 in heart failure mice. Pharmacol. Res. 2019, 146, 104318. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Xin, S.; Zhu, L.; Wang, M.; Hao, Y.; Zhao, Y.; Li, Y.; Hou, Y. Si-Miao-Yong-An Decoction Maintains the Cardiac Function and Protects Cardiomyocytes from Myocardial Ischemia and Reperfusion Injury. Evid. Based Complement. Alternat. Med. 2021, 2021, 8968464. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhong, P.; Sun, L. Exogenous hydrogen sulfide mitigates NLRP3 inflammasome-mediated inflammation through promoting autophagy via the AMPK-mTOR pathway. Biol. Open 2019, 8, bio043653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, L.; Wang, X.; Pan, J.; Zhang, M.; Liu, D.; Liu, M.; Li, L.; An, F. The co-treatment of rosuvastatin with dapagliflozin synergistically inhibited apoptosis via activating the PI3K/AKt/mTOR signaling pathway in myocardial ischemia/reperfusion injury rats. Open Med. 2021, 15, 47–57. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, H.; Liu, D.H.; Wang, G.N. Effects of dexmedetomidine on myocardial ischemia-reperfusion injury through PI3K-Akt-mTOR signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6736–6743. [Google Scholar]
- Sun, W.; Lu, H.; Dong, S.; Li, R.; Chu, Y.; Wang, N.; Zhao, Y.; Zhang, Y.; Wang, L.; Sun, L.; et al. Beclin1 controls caspase-4 inflammsome activation and pyroptosis in mouse myocardial reperfusion-induced microvascular injury. Cell Commun. Signal 2021, 19, 107. [Google Scholar] [CrossRef]
- Apichartpiyakul, P.; Shinlapawittayatorn, K.; Rerkasem, K.; Chattipakorn, S.C.; Chattipakorn, N. Mechanisms and Interventions on Acute Lower Limb Ischemia/Reperfusion Injury: A Review and Insights from Cell to Clinical Investigations. Ann. Vasc. Surg. 2022. [Google Scholar] [CrossRef]
- Eliason, J.L.; Wakefield, T.W. Metabolic consequences of acute limb ischemia and their clinical implications. Semin. Vasc. Surg. 2009, 22, 29–33. [Google Scholar] [CrossRef]
- Liu, L.; Yang, C.; Qiu, T.; Shen, X.; Liu, B.; Qi, X.; Song, G. Hydrogen alleviates acute lung injury induced by limb ischaemia/reperfusion in mice. Life Sci. 2021, 279, 119659. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, W.; Chen, Y.; Du, Z.; Zhu, F.; Wang, T.; Jiang, B. Ischemic postconditioning ameliorates acute kidney injury induced by limb ischemia/reperfusion via transforming TLR4 and NF-kappaB signaling in rats. J. Orthop. Surg. Res. 2021, 16, 416. [Google Scholar] [CrossRef]
- Kobayashi, J.; Murata, I. Nitrite as a pharmacological intervention for the successful treatment of crush syndrome. Physiol. Rep. 2018, 6, e13633. [Google Scholar] [CrossRef] [PubMed]
- Mangla, B.; Javed, S.; Sultan, M.H.; Kumar, P.; Kohli, K.; Najmi, A.; Alhazmi, H.A.; Al Bratty, M.; Ahsan, W. Sulforaphane: A review of its therapeutic potentials, advances in its nanodelivery, recent patents, and clinical trials. Phytother. Res. 2021, 35, 5440–5458. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Hwang, E.S.; Choi, G.Y.; Kim, H.B.; Park, K.S.; Sul, J.Y.; Hwang, Y.; Choi, G.W.; Kim, B.I.; Park, H.; et al. Sulforaphane enhances long-term potentiation and ameliorate scopolamine-induced memory impairment. Physiol. Behav. 2021, 238, 113467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Wu, Q. Sulforaphane protects intestinal epithelial cells against lipopolysaccharide-induced injury by activating the AMPK/SIRT1/PGC-1a pathway. Bioengineered 2021, 12, 4349–4360. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Y.; Wang, L.; Yan, C.; Liu, H.; Zhang, W.; Zhao, H.; Cheng, C.; Chen, Z.; Xu, T.; et al. Oridonin attenuates hind limb ischemia-reperfusion injury by modulating Nrf2-mediated oxidative stress and NLRP3-mediated inflammation. J. Ethnopharmacol. 2022, 292, 115206. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Zhou, W.; Men, H.; Bao, T.; Sun, Y.; Wang, Q.; Tan, Y.; Keller, B.B.; Tong, Q.; et al. Ferroptosis is essential for diabetic cardiomyopathy and is prevented by sulforaphane via AMPK/NRF2 pathways. Acta Pharm. Sin. B 2022, 12, 708–722. [Google Scholar] [CrossRef]
- Hua, D.; Luo, A.; Wu, Z.; Huang, C.; Li, S.; Xu, X.; Xu, J.; Yang, C.; Wang, D.; Liu, C. Sulforaphane improves cognitive dysfunction after surgery and anesthesia in mice: The role of Keap1-Nrf2 signaling. Brain Res. Bull. 2022, 181, 1–11. [Google Scholar] [CrossRef]
- Sandouka, S.; Shekh-Ahmad, T. Induction of the Nrf2 Pathway by Sulforaphane Is Neuroprotective in a Rat Temporal Lobe Epilepsy Model. Antioxidants 2021, 10, 1702. [Google Scholar] [CrossRef]
- Sun, H.; Wang, J.; Bi, W.; Zhang, F.; Chi, K.; Shi, L.; Yuan, T.; Ma, K.; Gao, X. Sulforaphane Ameliorates Limb Ischemia/Reperfusion-Induced Muscular Injury in Mice by Inhibiting Pyroptosis and Autophagy via the Nrf2-ARE Pathway. Evid. Based Complement. Alternat. Med. 2022, 2022, 4653864. [Google Scholar] [CrossRef]
- Zou, Y.; Luo, X.; Feng, Y.; Fang, S.; Tian, J.; Yu, B.; Li, J. Luteolin prevents THP-1 macrophage pyroptosis by suppressing ROS production via Nrf2 activation. Chem. Biol. Interact. 2021, 345, 109573. [Google Scholar] [CrossRef]
- Zhou, T.; Li, Z.; Chen, H. Melatonin alleviates lipopolysaccharide (LPS)/adenosine triphosphate (ATP)-induced pyroptosis in rat alveolar Type II cells (RLE-6TN) through nuclear factor erythroid 2-related factor 2 (Nrf2)-driven reactive oxygen species (ROS) downregulation. Bioengineered 2022, 13, 1880–1892. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, W.; Li, S.; Yang, S.; Zhang, Y.; Hu, X.; Qiu, H.; Wu, J.; Xu, S.; Chu, T. Nox4 Promotes RANKL-Induced Autophagy and Osteoclastogenesis via Activating ROS/PERK/eIF-2alpha/ATF4 Pathway. Front. Pharmacol. 2021, 12, 751845. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Gao, J.; Li, P.; Wang, Y.; Qi, Q.; Liu, X.; Li, J.; Wang, C.; Du, L. Pyroptosis, metabolism, and tumor immune microenvironment. Clin. Transl. Med. 2021, 11, e492. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| The Type of Ischemia Reperfusion (I/R) Injury | The Role of Autophagy and Pyroptosis | Experimental Model | Reference |
|---|---|---|---|
| cerebral I/R injury aggravated by diabetes | mild hypothermia improved cerebral I/R injury aggravated by diabetes by inhibiting pyroptosis via promoting autophagy | type 2 diabetic rats/PC-12 cells with cerebral I/R injury | [45] |
| cerebral I/R injury | spautin-1 improved cerebral I/R injury by inhibiting autophagy and NLRP3 inflammasome-mediated pyroptosis | rats/PC-12 cells with cerebral I/R injury | [52] |
| spinal cord I/R injury (SCIRI) | baicalein suppressed pyroptosis and ER stress-mediated apoptosis to ameliorate SCIR by promoting autophagy | mice with SCIRI | [64] |
| microglial with oxygen-glucose deprivation/reoxygenation | MSC-exos attenuated I/R-induced pyroptosis of microglia by increasing FOXO3a expression to enhance mitophagy | oxygen-glucose deprivation/reperfusion-induced microglial | [75] |
| myocardial I/R injury | SMYAD improves myocardial I/R injury by promoting autophagy via mTOR pathway and inhibiting pyroptosis | mice/H9C2 cells with myocardial I/R injury | [83] |
| myocardial I/R injury | beclin1 overexpression ameliorated myocardial I/R injury by suppressing caspase-4-mediated pyroptosis by promoting autophagy | mice with myocardial I/R injury | [87] |
| limb I/R injury | SFN may inhibit autophagy and pyroptosis by activating Nrf2 pathway to mitigate muscle injury caused by limb I/R injury | mice with limb I/R injury | [101] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.; Yang, Y.; Si, X.; Liu, H.; Wang, H. The Role of Pyroptosis and Autophagy in Ischemia Reperfusion Injury. Biomolecules 2022, 12, 1010. https://doi.org/10.3390/biom12071010
Zhao H, Yang Y, Si X, Liu H, Wang H. The Role of Pyroptosis and Autophagy in Ischemia Reperfusion Injury. Biomolecules. 2022; 12(7):1010. https://doi.org/10.3390/biom12071010
Chicago/Turabian StyleZhao, Huijie, Yihan Yang, Xinya Si, Huiyang Liu, and Honggang Wang. 2022. "The Role of Pyroptosis and Autophagy in Ischemia Reperfusion Injury" Biomolecules 12, no. 7: 1010. https://doi.org/10.3390/biom12071010