
Zinc Metalloprotease ProA from Legionella pneumophila Inhibits the Pro-Inflammatory Host Response by Degradation of Bacterial Flagellin

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation of Bacteria and Eukaryotic Cell Lines
2.2. Isolation and Purification of Zinc Metalloprotease ProA
2.3. Isolation of Native Flagellin
2.4. Flagellin Degradation Assay
2.5. Collection of Human Lung Tissue Explant Supernatant
2.6. HEK-Blue™ hTLR5 Detection Assay
2.7. Statistical Analyses
3. Results
3.1. Zinc Metalloprotease ProA Degrades Monomeric Flagellin
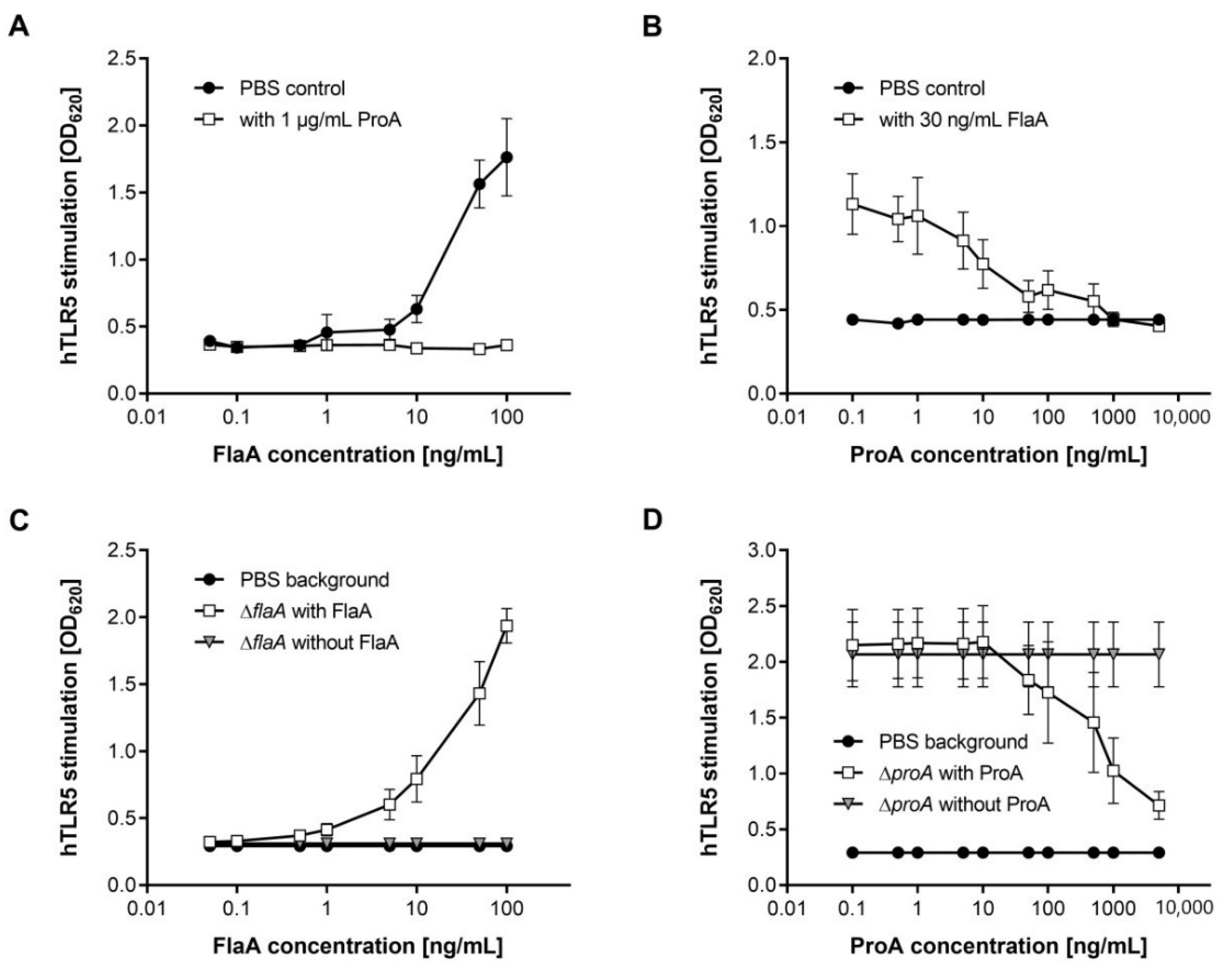
3.2. Flagellin-Mediated TLR5 Activation Is Prevented by ProA
3.3. Immune Recognition of the L. pneumophila WT via TLR5 Is Reduced by ProA
3.4. ProA Diminishes Extracellular FlaA Stimuli during Human Lung Tissue Infections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraser, D.W.; Tsai, T.R.; Orenstein, W.; Parkin, W.E.; Beecham, H.J.; Sharrar, R.G.; Harris, J.; Mallison, G.F.; Martin, S.M.; McDade, J.E.; et al. Legionnaires’ disease: Description of an epidemic of pneumonia. N. Engl. J. Med. 1977, 297, 1189–1197. [Google Scholar] [CrossRef]
- McDade, J.E.; Brenner, D.J.; Bozeman, F.M. Legionnaires’ disease bacterium isolated in 1947. Ann. Intern. Med. 1979, 90, 659–661. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Stamos, J.K.; Kamp, D.W. Infectivity of Legionella pneumophila Mip Mutant for Alveolar Epithelial Cells. Curr. Microbiol. 1995, 30, 247–250. [Google Scholar] [CrossRef]
- Steinert, M.; Hentschel, U.; Hacker, J. Legionella pneumophila: An Aquatic Microbe Goes Astray. FEMS Microbiol. Rev. 2002, 26, 149–162. [Google Scholar] [CrossRef]
- Swanson, M.S.; Hammer, B.K. Legionella pneumophila pathogenesis: A fateful journey from amoebae to macrophages. Annu. Rev. Microbiol. 2000, 54, 567–613. [Google Scholar] [CrossRef]
- Akira, S. Toll-like receptor signaling. J. Biol. Chem. 2003, 278, 38105–38108. [Google Scholar] [CrossRef] [Green Version]
- Janeway Jr, C.A.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef] [Green Version]
- Pedra, J.H.; Cassel, S.L.; Sutterwala, F.S. Sensing pathogens and danger signals by the inflammasome. Curr. Opin. Immunol. 2009, 21, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Vinzing, M.; Eitel, J.; Lippmann, J.; Hocke, A.C.; Zahlten, J.; Slevogt, H.; Dje N’Guessan, P.; Günther, S.; Schmeck, B.; Hippenstiel, S.; et al. NAIP and Ipaf control Legionella pneumophila replication in human cells. J. Immunol. 2008, 180, 6808–6815. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.D.; Andersen-Nissen, E.; Hayashi, F.; Strobe, K.; Bergman, M.A.; Barrett, S.L.R.; Cookson, B.T.; Aderem, A. Toll-like Receptor 5 Recognizes a Conserved Site on Flagellin Required for Protofilament Formation and Bacterial Motility. Nat. Immunol. 2003, 4, 1247–1253. [Google Scholar] [CrossRef]
- Gewirtz, A.T.; Navas, T.A.; Lyons, S.; Godowski, P.J.; Madara, J.L. Cutting Edge: Bacterial Flagellin Activates Basolaterally Expressed TLR5 to Induce Epithelial Proinflammatory Gene Expression. J. Immunol. 2001, 167, 1882–1885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevance, F.F.; Hughes, K.T. Coordinating assembly of a bacterial macromolecular machine. Nat. Rev. Microbiol. 2008, 6, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samatey, F.A.; Imada, K.; Nagashima, S.; Vonderviszt, F.; Kumasaka, T.; Yamamoto, M.; Namba, K. Structure of the bacterial flagellar protofilament and implications for a switch for supercoiling. Nature 2001, 410, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Yonekura, K.; Maki-Yonekura, S.; Namba, K. Complete atomic model of the bacterial flagellar filament by electron cryomicroscopy. Nature 2003, 424, 643–650. [Google Scholar] [CrossRef]
- Donnelly, M.A.; Steiner, T.S. Two Nonadjacent Regions in Enteroaggregative Escherichia coli Flagellin Are Required for Activation of Toll-like Receptor 5. J. Biol. Chem. 2002, 277, 40456–40461. [Google Scholar] [CrossRef] [Green Version]
- Forstnerič, V.; Ivičak-Kocjan, K.; Plaper, T.; Jerala, R.; Benčina, M. The role of the C-terminal D0 domain of flagellin in activation of Toll like receptor 5. PLoS Pathog. 2017, 13, e1006574. [Google Scholar]
- Song, W.S.; Jeon, Y.J.; Namgung, B.; Hong, M.; Yoon, S. A conserved TLR5 binding and activation hot spot on flagellin. Sci. Rep. 2017, 7, 40878. [Google Scholar] [CrossRef] [Green Version]
- Gohda, J.; Matsumura, T.; Inoue, J.I. Cutting Edge: TNFR-Associated Factor (TRAF) 6 Is Essential for MyD88-Dependent Pathway but Not Toll/IL-1 Receptor Domain-Containing Adaptor-Inducing IFN-Beta (TRIF)-Dependent Pathway in TLR Signaling. J. Immunol. 2004, 173, 2913–2917. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, C.; Heuner, K.; Brand, B.C.; Hacker, J.; Steinert, M. Flagellum of Legionella pneumophila positively affects the early phase of infection of eukaryotic host cells. Infect. Immun. 2001, 69, 2116–2122. [Google Scholar] [CrossRef] [Green Version]
- Hammer, B.K.; Tateda, E.S.; Swanson, M.S. A two-component regulator induces the transmission phenotype of stationary-phase Legionella pneumophila. Mol. Microbiol. 2002, 44, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Heuner, K.; Steinert, M. The Flagellum of Legionella pneumophila and Its Link to the Expression of the Virulent Phenotype. Int. J. Med. Microbiol. 2003, 293, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Bickel, M. The Role of Interleukin-8 in Inflammation and Mechanisms of Regulation. J. Periodontol. 1993, 64, 456–460. [Google Scholar]
- Hawn, T.R.; Verbon, A.; Lettinga, K.D.; Zhao, L.P.; Li, S.S.; Laws, R.J.; Skerrett, S.J.; Beutler, B.; Schroeder, L.; Nachman, A.; et al. A Common Dominant TLR5 Stop Codon Polymorphism Abolishes Flagellin Signaling and Is Associated with Susceptibility to Legionnaires’ Disease. J. Exp. Med. 2003, 198, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Hawn, T.R.; Berrington, W.R.; Smith, I.A.; Uematsu, S.; Akira, S.; Aderem, A.; Smith, K.D.; Skerrett, S.J. Altered inflammatory responses in TLR5-deficient mice infected with Legionella pneumophila. J. Immunol. 2007, 179, 6981–6987. [Google Scholar] [CrossRef] [Green Version]
- Mukaida, N.; Harada, A.; Matsushima, K. Interleukin-8 (IL-8) and Monocyte Chemotactic and Activating Factor (MCAF/MCP-1), Chemokines Essentially Involved in Inflammatory and Immune Reactions. Cytokine Growth Factor Rev. 1998, 9, 9–23. [Google Scholar] [CrossRef]
- Takamatsu, R.; Teruya, H.; Takeshima, E.; Ishikawa, C.; Matsumoto, K.; Mukaida, N.; Li, J.-D.; Heuner, K.; Higa, F.; Fujita, J.; et al. Retraction: Molecular characterization of Legionella pneumophila-induced interleukin-8 expression in T cells. BMC Microbiol. 2011, 10, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Alhede, M.; Bjarnsholt, T.; Givskov, M.; Alhede, M. Pseudomonas aeruginosa biofilms: Mechanisms of immune evasion. Adv. Appl. Microbiol. 2014, 86, 1–40. [Google Scholar] [CrossRef]
- Mejías-Luque, R.; Gerhard, M. Immune Evasion Strategies and Persistence of Helicobacter Pylori. Curr. Top. Microbiol. Immunol. 2017, 400, 53–71. [Google Scholar] [CrossRef]
- Zecconi, A.; Scali, F. Staphylococcus aureus Virulence Factors in Evasion from Innate Immune Defenses in Human and Animal Diseases. Immunol. Lett. 2013, 150, 12–22. [Google Scholar] [CrossRef]
- Andersen-Nissen, E.; Smith, K.D.; Strobe, K.L.; Barrett, S.L.R.; Cookson, B.T.; Logan, S.M.; Aderem, A. Evasion of Toll-like receptor 5 by flagellated bacteria. Proc. Natl. Acad. Sci. USA 2005, 102, 9247–9252. [Google Scholar] [CrossRef] [Green Version]
- Bardoel, B.W.; van der Ent, S.; Pel, M.J.C.; Tommassen, J.; Pieterse, C.M.J.; van Kessel, K.P.M.; van Strijp, J.A.G. Pseudomonas evades immune recognition of flagellin in both mammals and plants. PLoS Pathog. 2011, 7, e1002206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casilag, F.; Lorenz, A.; Krueger, J.; Klawonn, F.; Weiss, S.; Häussler, S. The LasB Elastase of Pseudomonas aeruginosa Acts in Concert with Alkaline Protease AprA To Prevent Flagellin-Mediated Immune Recognition. Infect. Immun. 2015, 84, 162–171. [Google Scholar] [CrossRef] [Green Version]
- Dreyfus, L.A.; Iglewski, B.H. Purification and Characterization of an Extracellular Protease of Legionella pneumophila. Infect. Immun. 1986, 51, 736–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hales, L.M.; Shuman, H.A. Legionella pneumophila contains a type II general secretion pathway required for growth in amoebae as well as for secretion of the Msp protease. Infect. Immun. 1999, 67, 3662–3666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.R.; Miller, R.D.; Iglewski, B.H. In vitro production of an extra-cellular protease by Legionella pneumophila. Infect. Immun. 1981, 34, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Tiaden, A.; Spirig, T.; Weber, S.S.; Brüggemann, H.; Bosshard, R.; Buchrieser, C.; Hilbi, H. The Legionella pneumophila response regulator LqsR promotes host cell interactions as an element of the virulence regulatory network controlled by RpoS and LetA. Cell. Microbiol. 2007, 9, 2903–2920. [Google Scholar] [CrossRef]
- Keen, M.G.; Hoffman, P.S. Characterization of a Legionella pneumophila Extracellular Protease Exhibiting Hemolytic and Cytotoxic Activities. Infect. Immun. 1989, 57, 732–738. [Google Scholar] [CrossRef] [Green Version]
- Müller, H.E. Proteolytic Action of Legionella pneumophila on Human Serum Proteins. Infect. Immun. 1980, 27, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Quinn, F.D.; Tompkins, L.S. Analysis of a cloned sequence of Legionella pneumophila encoding a 38 kD metalloprotease possessing haemolytic and cytotoxic activities. Mol. Microbiol. 1989, 3, 797–805. [Google Scholar] [CrossRef]
- Scheithauer, L.; Thiem, S.; Schmelz, S.; Dellmann, A.; Büssow, K.; Brouwer, R.M.H.J.; Ünal, C.M.; Blankenfeldt, W.; Steinert, M. Zinc metalloprotease ProA of Legionella pneumophila increases alveolar septal thickness in human lung tissue explants by collagen IV degradation. Cell. Microbiol. 2021, 23, e13313. [Google Scholar] [CrossRef]
- Conlan, J.W.; Baskerville, A.; Ashworth, L.A. Separation of Legionella pneumophila proteases and purification of a protease which produces lesions like those of Legionnaires’ disease in guinea pig lung. J. Gen. Microbiol. 1986, 132, 1565–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechnitzer, C.; Diamant, M.; Pedersen, B.K. Inhibition of Human Natural Killer Cell Activity by Legionella pneumophila Protease. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 1989, 8, 989–992. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.; Baskerville, A.; Dowsett, A.B.; Conlan, J.W. Immunocytochemical demonstration of the association between Legionella pneumophila, its tissue-destructive protease, and pulmonary lesions in experimental Legionnaires’ disease. J. Pathol. 1987, 153, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Hoppe, J.; Ünal, C.M.; Thiem, S.; Grimpe, L.; Goldmann, T.; Gaßler, N.; Richter, M.; Shevchuk, O.; Steinert, M. PilY1 Promotes Legionella pneumophila Infection of Human Lung Tissue Explants and Contributes to Bacterial Adhesion, Host Cell Invasion, and Twitching Motility. Front. Cell. Infect. Microbiol. 2017, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Jäger, J.; Marwitz, S.; Tiefenau, J.; Rasch, J.; Shevchuk, O.; Kugler, C.; Goldmann, T.; Steinert, M. Human lung tissue explants reveal novel interactions during Legionella pneumophila infections. Infect. Immun. 2014, 82, 275–285. [Google Scholar] [CrossRef] [Green Version]
- Scheithauer, L.; Steinert, M. Infection of Human Lung Tissue Explants (HLTEs) with Legionella pneumophila. In Legionella, 2nd ed.; Buchrieser, C., Hilbi, H., Eds.; Humana Press: New York, NY, USA, 2019; pp. 323–331. ISBN 978-1-4939-9047-4. [Google Scholar]
- Lang, C.; Rastew, E.; Hermes, B.; Siegbrecht, E.; Ahrends, R.; Banerji, S.; Flieger, A. Zinc metalloproteinase ProA directly activates Legionella pneumophila PlaC glycerophospholipid:cholesterol acyltransferase. J. Biol. Chem. 2012, 287, 23464–23478. [Google Scholar] [CrossRef] [Green Version]
- Wintermeyer, E.; Ludwig, B.; Steinert, M.; Schmidt, B.; Fischer, G.; Hacker, J. Influence of site specifically altered Mip proteins on intracellular survival of Legionella pneumophila in eukaryotic cells. Infect. Immun. 1995, 63, 4576–4583. [Google Scholar] [CrossRef] [Green Version]
- Jacobi, S.; Schade, R.; Heuner, K. Characterization of the Alternative Sigma Factor sigma54 and the Transcriptional Regulator FleQ of Legionella pneumophila, Which Are Both Involved in the Regulation Cascade of Flagellar Gene Expression. J. Bacteriol. 2004, 186, 2540–2547. [Google Scholar] [CrossRef] [Green Version]
- Rossier, O.; Starkenburg, S.R.; Cianciotto, N.P. Legionella pneumophila type II protein secretion promotes virulence in the A/J mouse model of Legionnaires’ disease pneumonia. Infect. Immun. 2004, 72, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Secades, P.; Guijarro, J.A. Purification and Characterization of an Extracellular Protease from the Fish Pathogen Yersinia ruckeri and Effect of Culture Conditions on Production. Appl. Environ. Microbiol. 1999, 65, 3969–3975. [Google Scholar] [CrossRef] [Green Version]
- Montie, T.C.; Craven, R.C.; Holder, I.A. Flagellar Preparations from Pseudomonas aeruginosa: Isolation and Characterization. Infect. Immun. 1982, 35, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heuner, K.; Bender-Beck, L.; Brand, B.C.; Lück, P.C.; Mann, K.H.; Marre, R.; Ott, M.; Hacker, J. Cloning and Genetic Characterization of the Flagellum Subunit Gene (flaA) of Legionella pneumophila Serogroup 1. Infect. Immun. 1995, 63, 2499–2507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, S.R. Digital image processing and analysis with ImageJ. Curr. Protoc. Essent. Lab. Tech. 2010, 3, 3A–3C. [Google Scholar]
- Black, W.J.; Quinn, F.D.; Tompkins, L.S. Legionella pneumophila zinc metalloprotease is structurally and functionally homologous to Pseudomonas aeruginosa elastase. J. Bacteriol. 1990, 172, 2608–2613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobb, L.M.; Mychaleckyj, J.C.; Wozniak, D.J.; López-Boado, Y.S. Pseudomonas aeruginosa Flagellin and Alginate Elicit Very Distinct Gene Expression Patterns in Airway Epithelial Cells: Implications for Cystic Fibrosis Disease. J. Immunol. 2004, 173, 5659–5670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honko, A.N.; Mizel, S.B. Effects of flagellin on innate and adaptive immunity. Immunol. Res. 2005, 33, 83–101. [Google Scholar] [CrossRef]
- Shikhagaie, M.; Bergqvist, A.; Andersson, C.; Mori, M.; Krohn, K.; Dahl, R.; Hoffman, H.J.; Bjermer, L.; Erjefält, J. Altered distribution patterns of TLR5 and TLR7 in the central airways of severe asthmatics (P4427). J. Immunol. 2013, 190, 206.2. [Google Scholar]
- Kawamoto, Y.; Morinaga, Y.; Kimura, Y.; Kaku, N.; Kosai, K.; Uno, N.; Hasegawa, H.; Yanagihara, K. TNF-α inhibits the growth of Legionella pneumophila in airway epithelial cells by inducing apoptosis. J. Infect. Chemother. 2017, 23, 51–55. [Google Scholar] [CrossRef]
- McCoy-Simandle, K.; Stewart, C.R.; Dao, J.; DebRoy, S.; Rossier, O.; Bryce, P.J.; Cianciotto, N.P. Legionella pneumophila type II secretion dampens the cytokine response of infected macrophages and epithelia. Infect. Immun. 2011, 79, 1984–1997. [Google Scholar] [CrossRef] [Green Version]
- Hell, W.; Essig, A.; Bohnet, S.; Gatermann, S.; Marre, R. Cleavage of tumor necrosis factor-alpha by Legionella exoprotease. APMIS 1993, 101, 120–126. [Google Scholar] [CrossRef]
- Mintz, C.S.; Miller, R.D.; Gutgsell, N.S.; Malek, T. Legionella pneumophila protease inactivates interleukin-2 and cleaves CD4 on human T cells. Infect. Immun. 1993, 61, 3416–3421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, F.; Cornicelli, M.D.; Kovach, M.A.; Newstead, M.W.; Zeng, X.; Kumar, A.; Gao, N.; Yoon, S.G.; Gallo, R.L.; Standiford, T.J. Flagellin stimulates protective lung mucosal immunity: Role of cathelicidin-related antimicrobial peptide. J. Immunol. 2010, 185, 1142–1149. [Google Scholar] [CrossRef] [PubMed]
- Im, J.; Jeon, J.H.; Cho, M.K.; Woo, S.S.; Kang, S.S.; Yun, C.H.; Lee, K.; Chung, D.K.; Han, S.H. Induction of IL-8 expression by bacterial flagellin is mediated through lipid raft formation and intracellular TLR5 activation in A549 cells. Mol. Immunol. 2009, 47, 614–622. [Google Scholar] [CrossRef] [PubMed]
- Schmeck, B.; N’Guessan, P.D.; Ollomang, M.; Lorenz, J.; Zahlten, J.; Opitz, B.; Flieger, A.; Suttorp, N.; Hippenstiel, S. Legionella pneumophila-induced NF-κB- and MAPK-dependent cytokine release by lung epithelial cells. Eur. Respir. J. 2007, 29, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Zeng, H.; Lyons, S.; Carlson, A.; Merlin, D.; Neish, A.S.; Gewirtz, A.T. TLR5-mediated activation of p38 MAPK regulates epithelial IL-8 expression via posttranscriptional mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, G282–G290. [Google Scholar] [CrossRef]
- Kunkel, S.L.; Standiford, T.; Kasahara, K.; Strieter, R.M. Interleukin-8 (IL-8): The major neutrophil chemotactic factor in the lung. Exp. Lung Res. 1991, 17, 17–23. [Google Scholar] [CrossRef]
- Akhter, A.; Gavrilin, M.A.; Frantz, L.; Washington, S.; Ditty, C.; Limoli, D.; Day, C.; Sarkar, A.; Newland, C.; Butchar, J.; et al. Caspase-7 activation by the Nlrc4/Ipaf inflammasome restricts Legionella pneumophila infection. PLoS Pathog. 2009, 5, e1000361. [Google Scholar] [CrossRef] [Green Version]
- Miao, E.A.; Andersen-Nissen, E.; Warren, S.E.; Aderem, A. TLR5 and Ipaf: Dual sensors of bacterial flagellin in the innate immune system. Semin. Immunopathol. 2007, 29, 275–288. [Google Scholar] [CrossRef]
- Molofsky, A.B.; Byrne, B.G.; Whitfield, N.N.; Madigan, C.A.; Fuse, E.T.; Tateda, K.; Swanson, M.S. Cytosolic recognition of flagellin by mouse macrophages restricts Legionella pneumophila infection. J. Exp. Med. 2006, 203, 1093–1104. [Google Scholar] [CrossRef]
- Truchan, H.K.; Christman, H.D.; White, R.C.; Rutledge, N.S.; Cianciotto, N.P. Type II Secretion Substrates of Legionella pneumophila Translocate Out of the Pathogen-Occupied Vacuole via a Semipermeable Membrane. mBio 2017, 8, e00870-17. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Description | References |
| HEK-BlueTM hTLR5 | HEK 293 cell line expressing human TLR5, SEAP reporter | InvivoGen, Toulouse, France |
| Strain | Description | References |
| L. pneumophila Corby WT | WT reference strain | [48] |
| L. pneumophila Corby ΔproA | proA::nptI, proA deletion mutant, Kmr | [40] |
| L. pneumophila Corby ΔproA proA | proA deletion mutant with pMMB2002-proA, Kmr, Cmlr | [40] |
| L. pneumophila Corby ΔflaA | flaA::nptI, flaA deletion mutant, Kmr | [49] |
| L. pneumophila Corby ΔflaA flaA | flaA deletion mutant with pMMB2002-flaA, Kmr, Cmlr | This study |
| P. aeruginosa PAO1 | WT strain | ATCC 15692 |
| Plasmid | Description | References |
| pMMB2002 | pMMB207-derived expression plasmid for L. pneumophila, deleted mobA gene, Cmlr | [50] |
| pMMB2002-proA | pMMB2002-derived vector expressing the proA gene, Cmlr | [40] |
| pMMB2002-flaA | pMMB2002-derived vector expressing the flaA gene, Cmlr | This study |
| Primer | 5′-3′-Sequence | References |
| FlaA-Pr_fw | CATGAGCTCTCGACTTGATAACCCGAACC | This study |
| FlaA+_rv | AAGGTACCCTATCGACCTAACAATGATAATAC | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scheithauer, L.; Thiem, S.; Ünal, C.M.; Dellmann, A.; Steinert, M. Zinc Metalloprotease ProA from Legionella pneumophila Inhibits the Pro-Inflammatory Host Response by Degradation of Bacterial Flagellin. Biomolecules 2022, 12, 624. https://doi.org/10.3390/biom12050624
Scheithauer L, Thiem S, Ünal CM, Dellmann A, Steinert M. Zinc Metalloprotease ProA from Legionella pneumophila Inhibits the Pro-Inflammatory Host Response by Degradation of Bacterial Flagellin. Biomolecules. 2022; 12(5):624. https://doi.org/10.3390/biom12050624
Chicago/Turabian StyleScheithauer, Lina, Stefanie Thiem, Can M. Ünal, Ansgar Dellmann, and Michael Steinert. 2022. "Zinc Metalloprotease ProA from Legionella pneumophila Inhibits the Pro-Inflammatory Host Response by Degradation of Bacterial Flagellin" Biomolecules 12, no. 5: 624. https://doi.org/10.3390/biom12050624