
Early Social Enrichment Modulates Tumor Progression and p53 Expression in Adult Mice

 ,
,  , , , and
, , , and 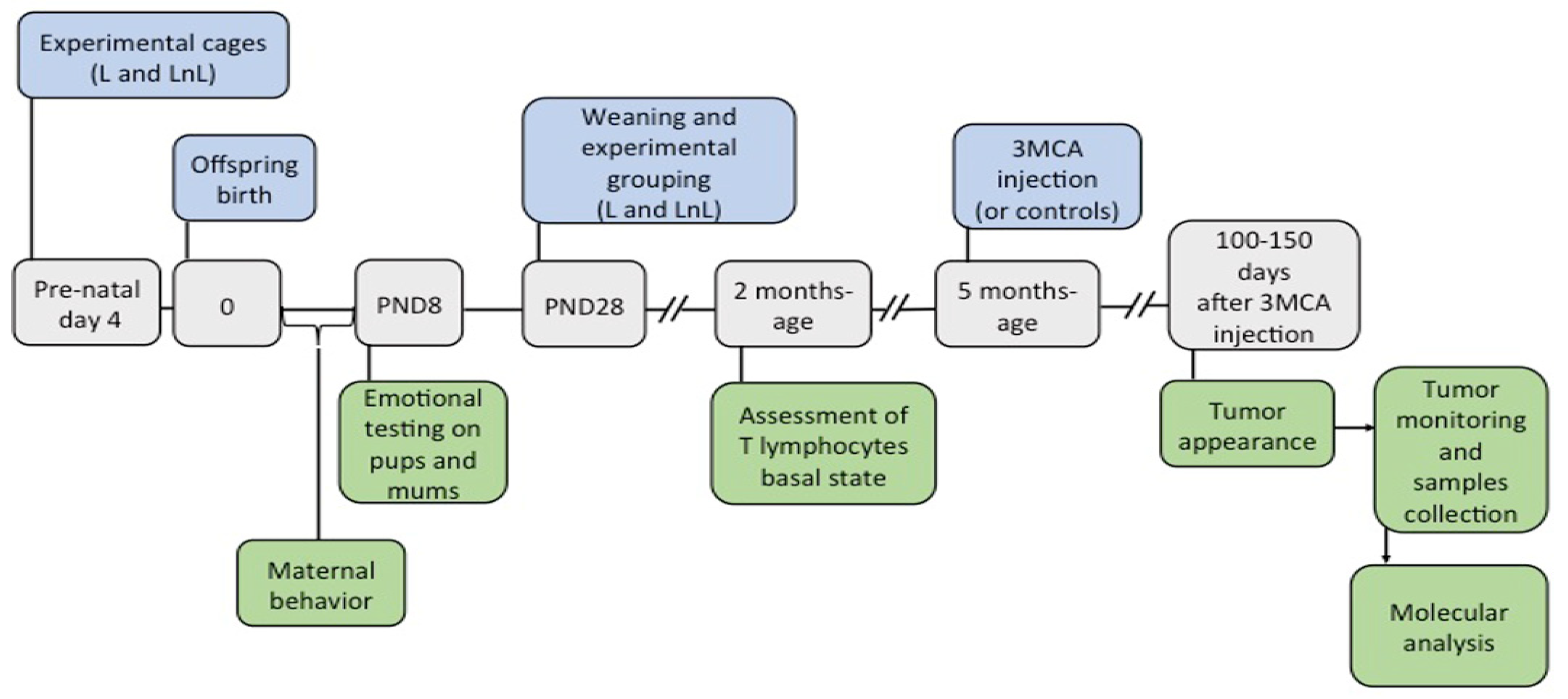{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Social Enrichment, and Experimental Groups
2.2. Behavioral Tests and Measurement of Body Temperature
2.3. Chemical Carcinogenesis and Tissue Collection
2.4. Protein Analysis
2.5. mRNA Analysis
2.5.1. RNA Extraction and cDNA Synthesis
2.5.2. Genetic Analysis of TP53 Mutation
2.5.3. RT-PCR
2.6. FACS Analysis
3. Results
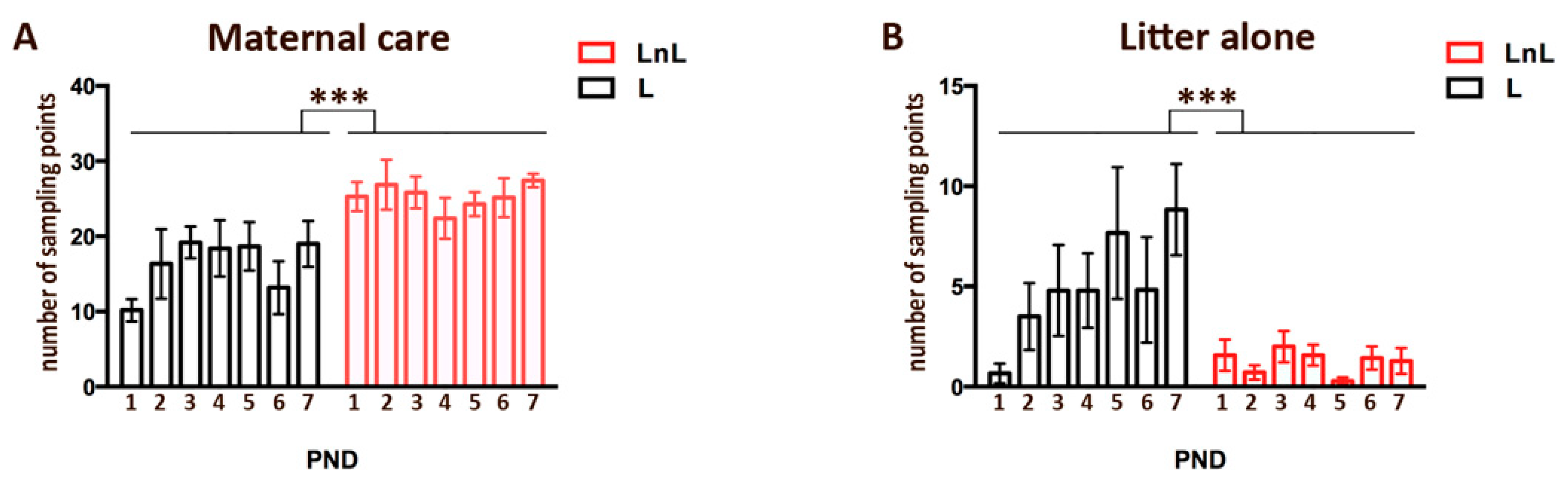
3.1. LnL Mice Receive More Maternal Care Than L Mice during the First Postnatal Week
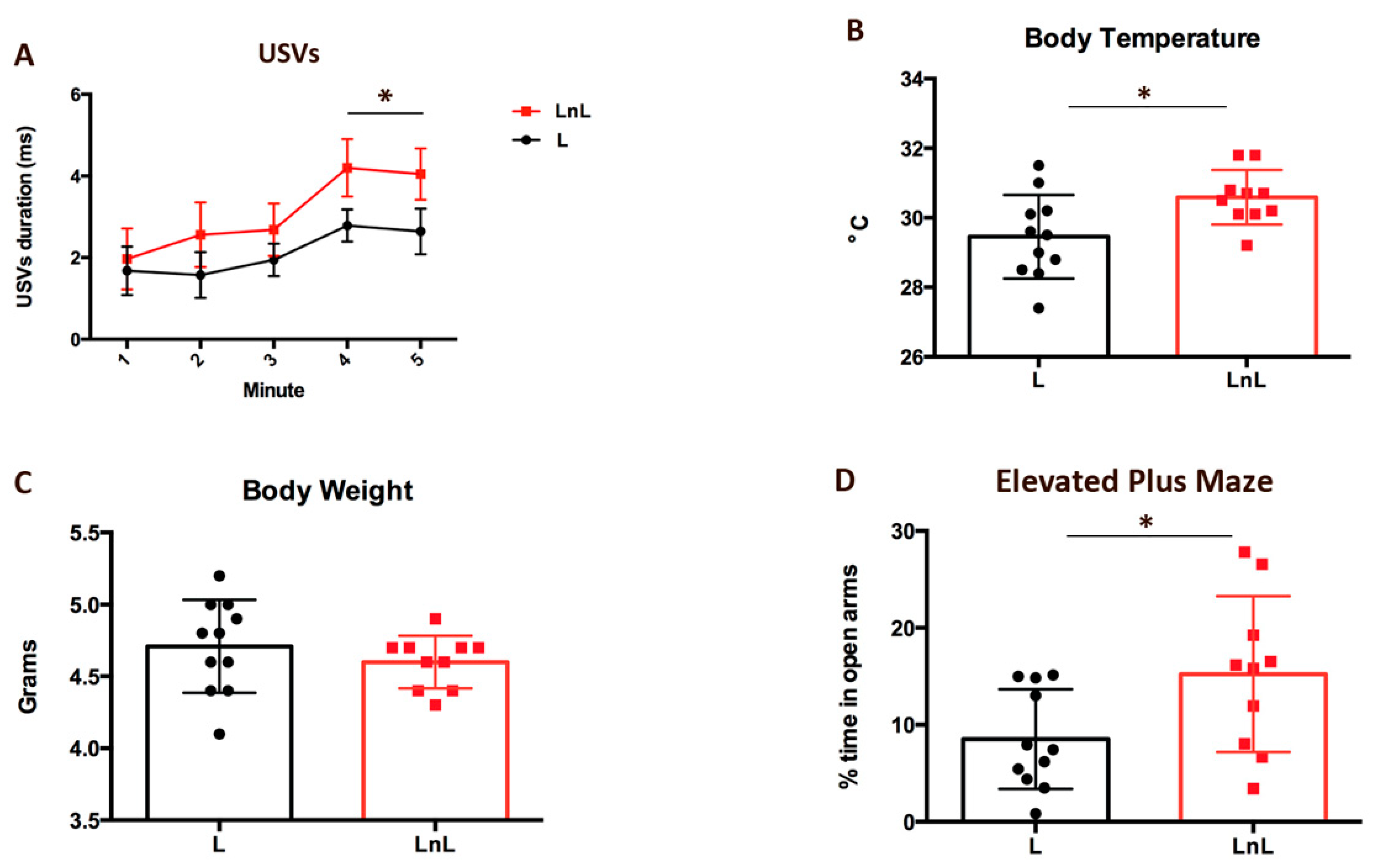
3.2. LnL Condition Favors Resilience to Separation in Pups and Reduces Anxiety in Mothers
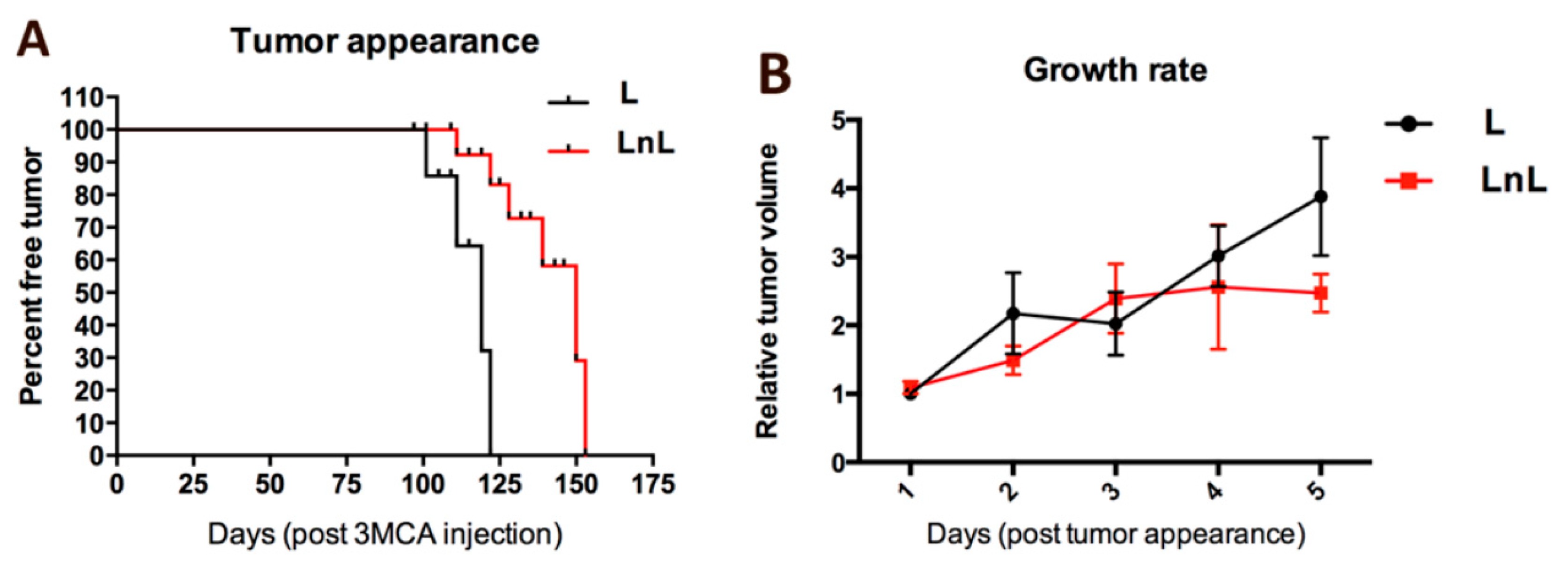
3.3. LnL Condition Delays the Onset of 3MCA-Induced Fibrosarcomas in Adult Mice
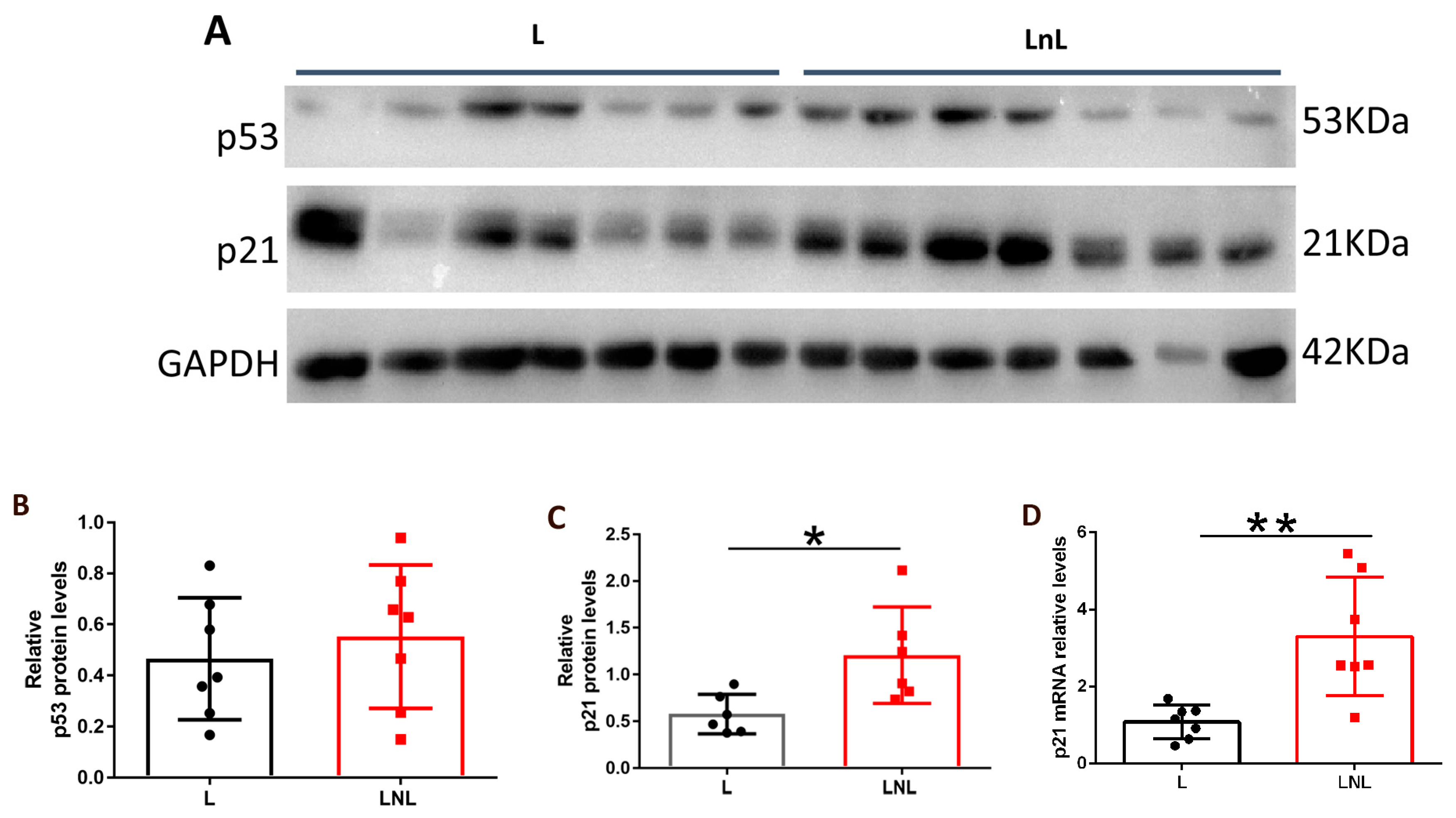
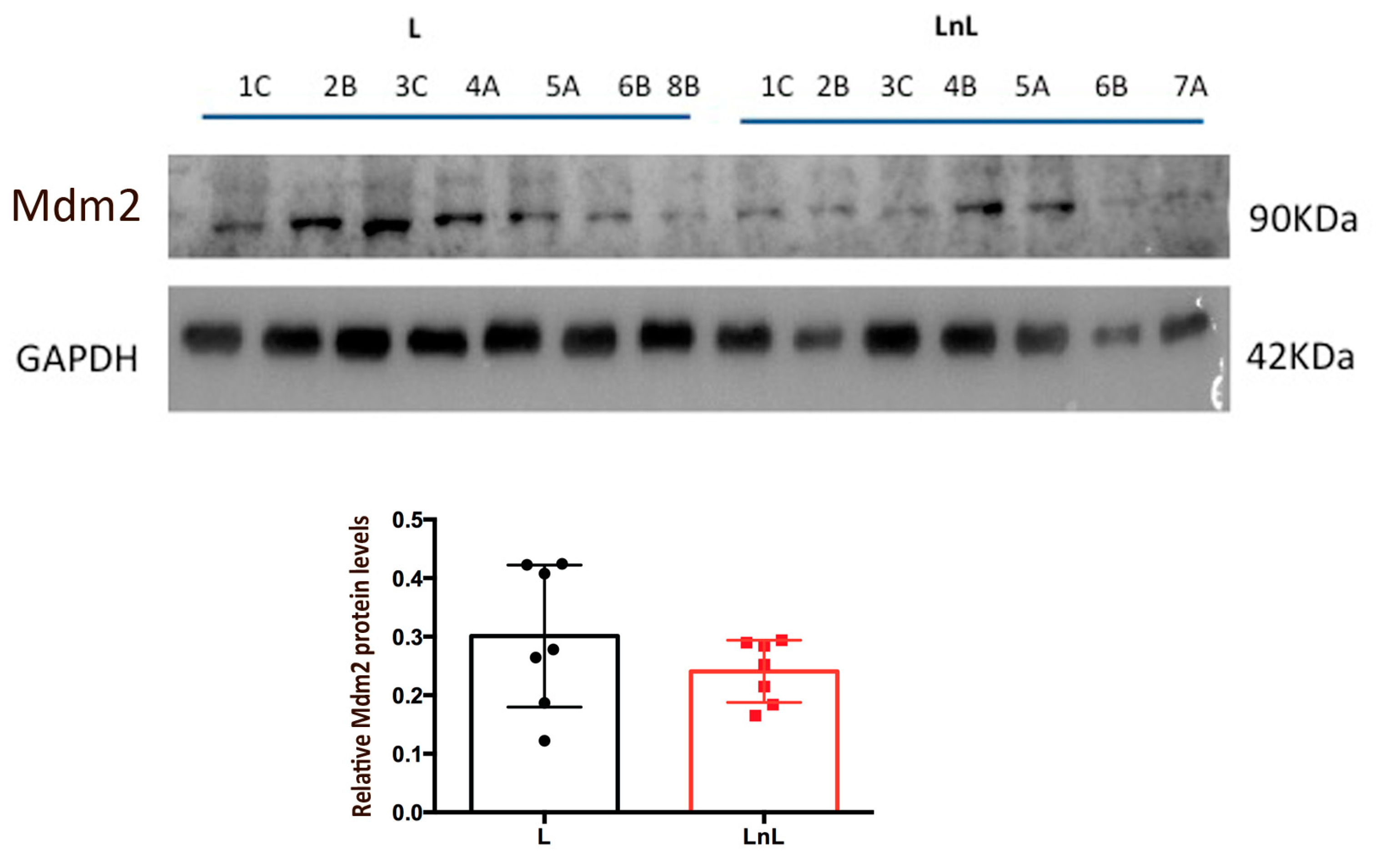
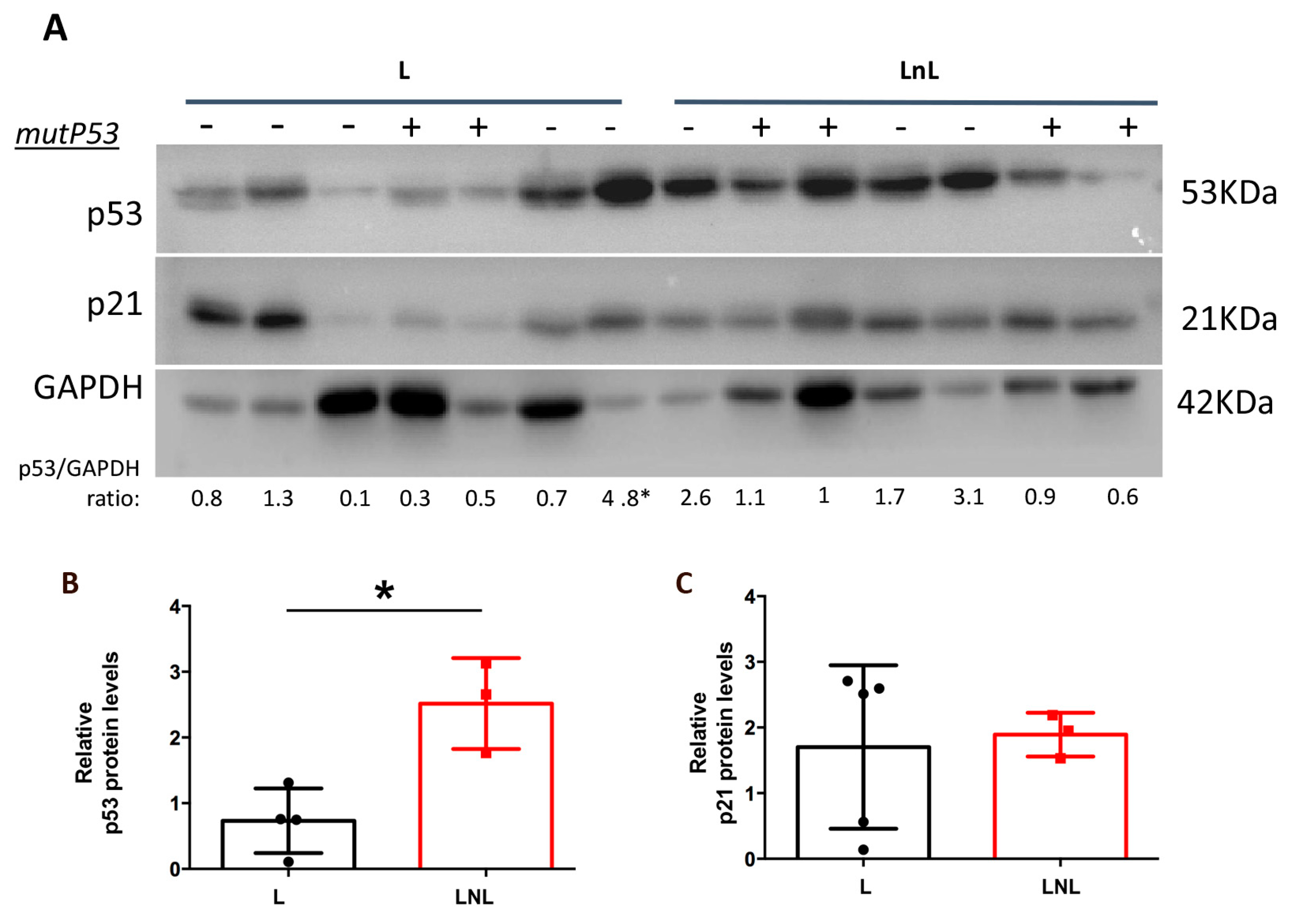
3.4. LnL Condition Is Associated to Increased p53 Activity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Antoni, M.H.; Lutgendorf, S.K.; Cole, S.W.; Dhabhar, F.S.; Sephton, S.; McDonald, P.G.; Stefanek, M.; Sood, A.K. The influence of bio-behavioural factors on tumour biology: Pathways and mechanisms. Nat. Cancer 2006, 6, 240–248. [Google Scholar] [CrossRef]
- Allison, P.; Guichard, C.; Fung, K.; Gilain, L. Dispositional Optimism Predicts Survival Status 1 Year after Diagnosis in Head and Neck Cancer Patients. J. Clin. Oncol. 2003, 21, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Eckerling, A.; Ricon-Becker, I.; Sorski, L.; Sandbank, E.; Ben-Eliyahu, S. Stress and cancer: Mechanisms, significance and future directions. Nat. Cancer 2021, 21, 767–785. [Google Scholar] [CrossRef] [PubMed]
- Mravec, B.; Tibensky, M.; Horvathova, L. Stress and cancer. Part I: Mechanisms mediating the effect of stressors on cancer. J. Neuroimmunol. 2020, 346, 577311. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.L.; Farrar, W.B.; Golden-Kreutz, D.M.; Glaser, R.; Emery, C.F.; Crespin, T.R.; Shapiro, C.L.; Carson, W.E. Psychological, Behavioral, and Immune Changes after a Psychological Intervention: A Clinical Trial. J. Clin. Oncol. 2004, 22, 3570–3580. [Google Scholar] [CrossRef]
- A McGregor, B.; Antoni, M.H.; Boyers, A.; Alferi, S.M.; Blomberg, B.B.; Carver, C. Cognitive–behavioral stress management increases benefit finding and immune function among women with early-stage breast cancer. J. Psychosom. Res. 2004, 56, 1–8. [Google Scholar] [CrossRef]
- Coyne, J.C.; Tennen, H. Positive Psychology in Cancer Care: Bad Science, Exaggerated Claims, and Unproven Medicine. Ann. Behav. Med. 2010, 39, 16–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutgendorf, S.K.; Sood, A.K.; Antoni, M.H. Host Factors and Cancer Progression: Biobehavioral Signaling Pathways and Interventions. J. Clin. Oncol. 2010, 28, 4094–4099. [Google Scholar] [CrossRef] [PubMed]
- Vousden, K.H.; Prives, C. Blinded by the Light: The Growing Complexity of p53. Cell 2009, 137, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, G.; Pan, C.; Bei, J.-X.; Li, B.; Liang, C.; Xu, Y.; Fu, X. Mutant p53 in Cancer Progression and Targeted Therapies. Front. Oncol. 2020, 10, 595187. [Google Scholar] [CrossRef] [PubMed]
- Donehower, L.A.; Harvey, M.; Slagle, B.L.; McArthur, M.J.; Montgomery, C.A., Jr.; Butel, J.S.; Bradley, A. Mice deficient for p53 are developmentally normal but susceptible to spontaneous tumours. Nature 1992, 356, 215–221. [Google Scholar] [CrossRef]
- García-Cao, I.; García-Cao, M.; Martín-Caballero, J.; Criado, L.M.; Klatt, P.; Flores, J.M.; Weill, J.-C.; Blasco, M.A.; Serrano, M. ’Super p53’ mice exhibit enhanced DNA damage response, are tumor resistant and age normally. EMBO J. 2002, 21, 6225–6235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Fontela, C.; Mandinova, A.; Aaronson, S.A.; Lee, A.M.S.W. Emerging roles of p53 and other tumour-suppressor genes in immune regulation. Nat. Rev. Immunol. 2016, 16, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Liu, S.; Liang, Y.; Zhou, Z.; Bian, W.; Liu, X. Stress Hormone Cortisol Enhances Bcl2 Like-12 Expression to Inhibit p53 in Hepatocellular Carcinoma Cells. Am. J. Dig. Dis. 2017, 62, 3495–3500. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Liu, L.; Zhang, C.; Zheng, T.; Wang, J.; Lin, M.; Zhao, Y.; Wang, X.; Levine, A.J.; Hu, W. Chronic restraint stress attenuates p53 function and promotes tumorigenesis. Proc. Natl. Acad. Sci. USA 2012, 109, 7013–7018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harshaw, C.; Culligan, J.J.; Alberts, J.R. Sex Differences in Thermogenesis Structure Behavior and Contact within Huddles of Infant Mice. PLoS ONE 2014, 9, e87405. [Google Scholar] [CrossRef]
- D’Amato, F.R.; Zanettini, C.; Sgobio, C.; Sarli, C.; Carone, V.; Moles, A.; Ammassari-Teule, M. Intensification of maternal care by double-mothering boosts cognitive function and hippocampal morphology in the adult offspring. Hippocampus 2011, 21, 298–308. [Google Scholar] [CrossRef] [PubMed]
- Oddi, D.; Subashi, E.; Middei, S.; Bellocchio, L.; Lemaire-Mayo, V.; Guzmán, M.; Crusio, W.; D’Amato, F.; Pietropaolo, S. Early Social Enrichment Rescues Adult Behavioral and Brain Abnormalities in a Mouse Model of Fragile X Syndrome. Neuropsychopharmacology 2015, 40, 1113–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbugino, L.; Centofante, E.; D’Amato, F.R. Early Social Enrichment Improves Social Motivation and Skills in a Monogenic Mouse Model of Autism, theOprm1−/−Mouse. Neural Plast. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.-H.; Pan, Y.; Zhu, L.; Landa, L.; Yoo, J.; Spencer, C.; Lorenzo, I.; Brilliant, M.; Noebels, J.; Beaudet, A.L. Altered Ultrasonic Vocalization and Impaired Learning and Memory in Angelman Syndrome Mouse Model with a Large Maternal Deletion from Ube3a to Gabrb3. PLoS ONE 2010, 5, e12278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucà, R.; Di Blasio, G.; Gallo, D.; Monteleone, V.; Manni, I.; Fici, L.; Buttarelli, M.; Ciolli, G.; Pellegrino, M.; Teveroni, E.; et al. Estrogens Counteract Platinum-Chemosensitivity by Modifying the Subcellular Localization of MDM4. Cancers 2019, 11, 1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lillberg, K.; Verkasalo, P.K.; Kaprio, J.; Teppo, L.; Helenius, H.; Koskenvuo, M. Stressful Life Events and Risk of Breast Cancer in 10,808 Women: A Cohort Study. Am. J. Epidemiol. 2003, 157, 415–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, P.; Kaplan, G.A. Social Connections and Risk for Cancer: Prospective Evidence from the Alameda County Study. Behav. Med. 1990, 16, 101–110. [Google Scholar] [CrossRef]
- Cao, L.; Liu, X.; Lin, E.-J.D.; Wang, C.; Choi, E.Y.; Riban, V.; Lin, B.; During, M.J. Environmental and Genetic Activation of a Brain-Adipocyte BDNF/Leptin Axis Causes Cancer Remission and Inhibition. Cell 2010, 142, 52–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nachat-Kappes, R.; Pinel, A.; Combe, K.; Lamas, B.; Farges, M.-C.; Rossary, A.; Goncalves-Mendes, N.; Caldefie-Chezet, F.; Vasson, M.-P.; Basu, S. Effects of Enriched Environment on COX-2, Leptin and Eicosanoids in a Mouse Model of Breast Cancer. PLoS ONE 2012, 7, e51525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benaroya-Milshtein, N.; Apter, A.; Yaniv, I.; Kukulansky, T.; Raz, N.; Haberman, Y.; Halpert, H.; Pick, C.G.; Hollander, N. Environmental Enrichment Augments the Efficacy of Idiotype Vaccination for B-cell Lymphoma. J. Immunother. 2007, 30, 517–522. [Google Scholar] [CrossRef] [Green Version]
- Westwood, J.A.; Darcy, P.K.; Kershaw, M.H. Environmental enrichment does not impact on tumor growth in mice. F1000Research 2013, 2, 140. [Google Scholar] [CrossRef] [PubMed]
- Sivaraman, L.; Conneely, O.M.; Medina, D.; O’Malley, B.W. p53 is a potential mediator of pregnancy and hormone-induced resistance to mammary carcinogenesis. Proc. Natl. Acad. Sci. USA 2001, 98, 12379–12384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginger, M.R.; Gonzalez-Rimbau, M.F.; Gay, J.P.; Rosen, J.M. Persistent Changes in Gene Expression Induced by Estrogen and Progesterone in the Rat Mammary Gland. Mol. Endocrinol. 2001, 15, 1993–2009. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Middei, S.; Giorgini, L.; Vacca, V.; Storri, F.; Putti, S.; Strimpakos, G.; Raspa, M.; Scavizzi, F.; Moretti, F.; D’Amato, F.R. Early Social Enrichment Modulates Tumor Progression and p53 Expression in Adult Mice. Biomolecules 2022, 12, 532. https://doi.org/10.3390/biom12040532
Middei S, Giorgini L, Vacca V, Storri F, Putti S, Strimpakos G, Raspa M, Scavizzi F, Moretti F, D’Amato FR. Early Social Enrichment Modulates Tumor Progression and p53 Expression in Adult Mice. Biomolecules. 2022; 12(4):532. https://doi.org/10.3390/biom12040532
Chicago/Turabian StyleMiddei, Silvia, Ludovica Giorgini, Valentina Vacca, Francesca Storri, Sabrina Putti, Georgios Strimpakos, Marcello Raspa, Ferdinando Scavizzi, Fabiola Moretti, and Francesca R. D’Amato. 2022. "Early Social Enrichment Modulates Tumor Progression and p53 Expression in Adult Mice" Biomolecules 12, no. 4: 532. https://doi.org/10.3390/biom12040532