
Identification of Novel Plasma Biomarkers for Abdominal Aortic Aneurysm by Protein Array Analysis

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Sample Collection
2.2. Antibody Array and Data Processing
2.3. Bioinformatics Analysis
2.4. ELISA
2.5. Statistical Analysis
3. Results
3.1. Clinical Characteristics of the Participants and Study Design
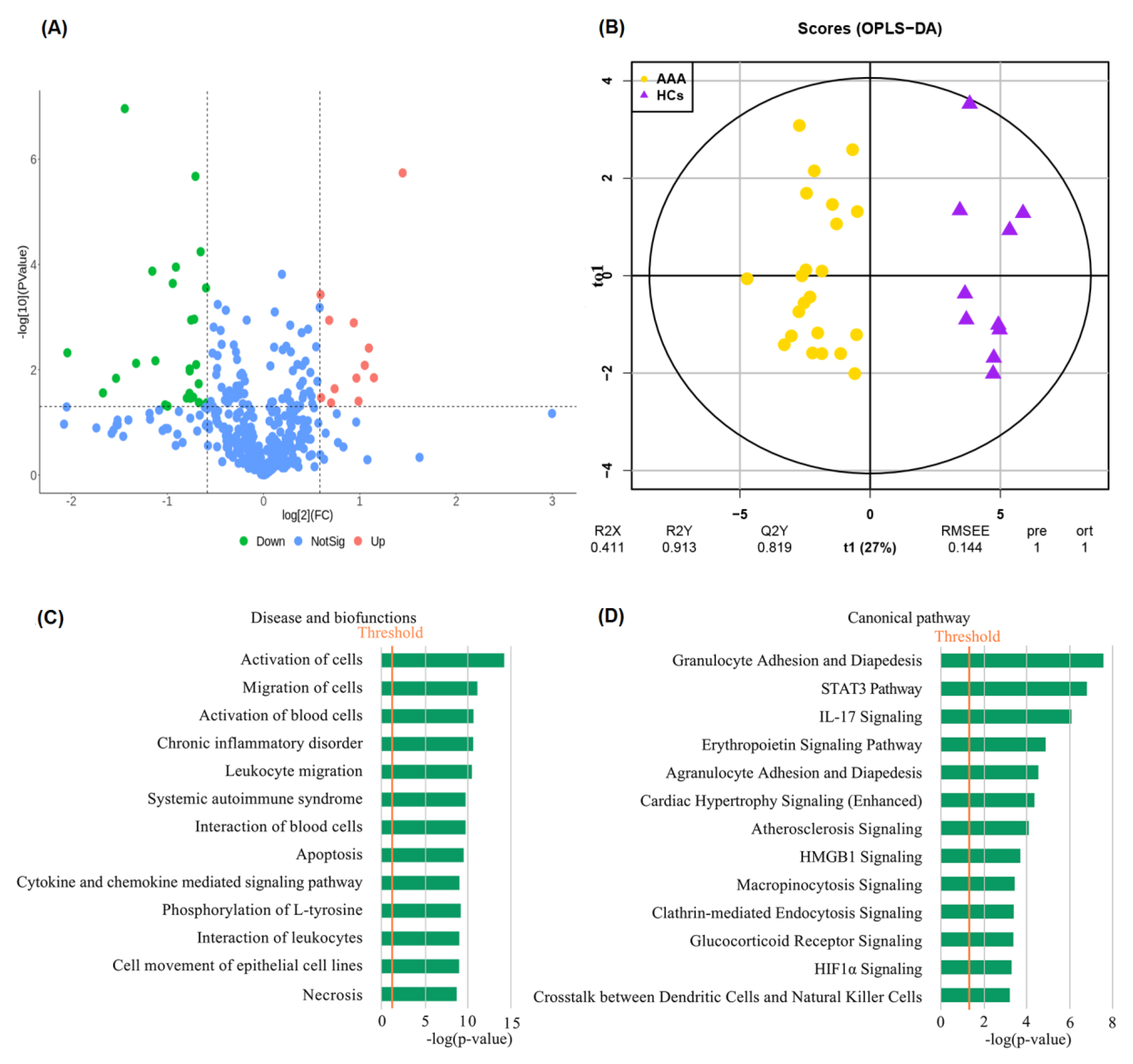
3.2. Identification of Differentially Expressed Proteins between AAA and HC
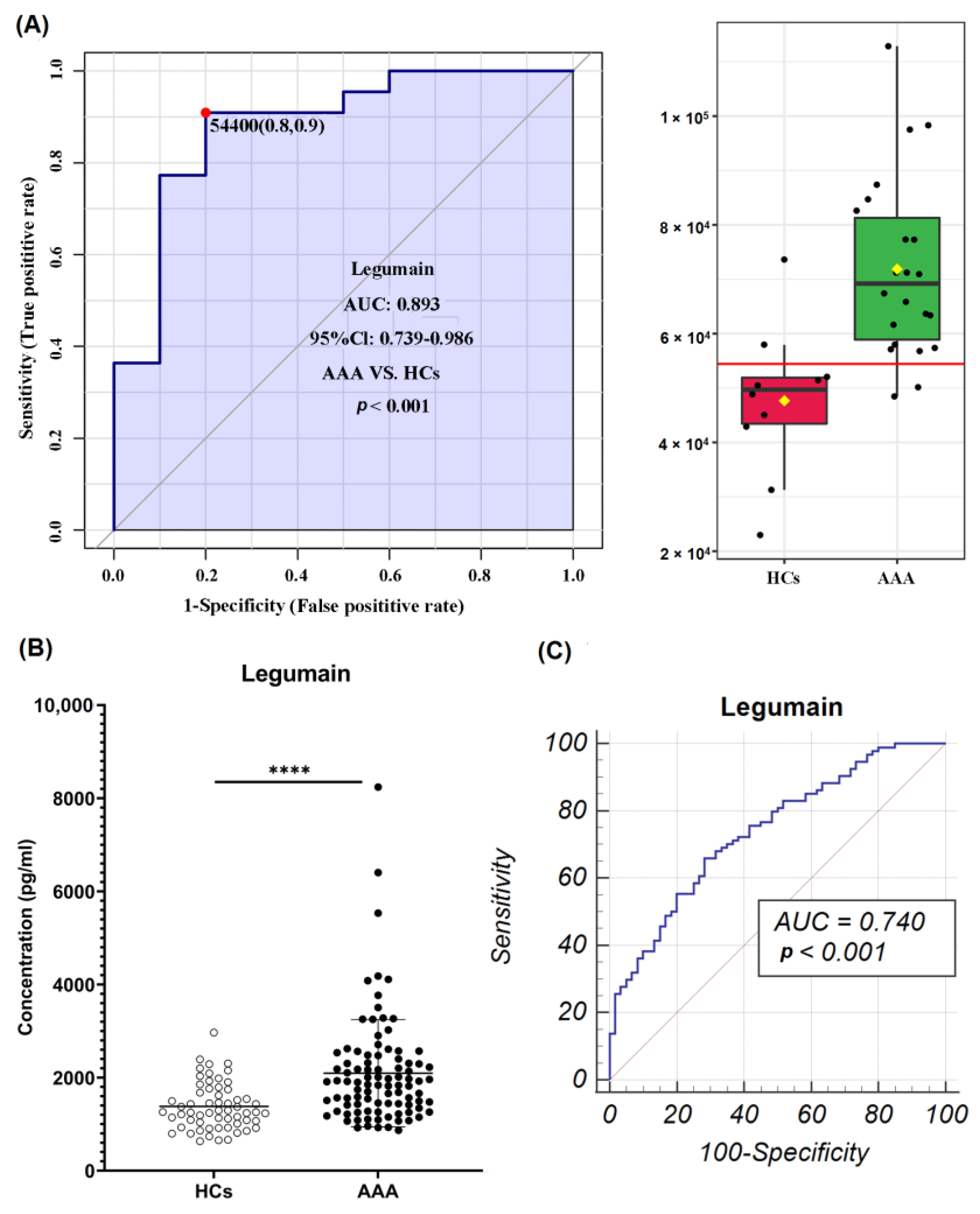
3.3. Identification of Potential Diagnostic Biomarkers of AAA
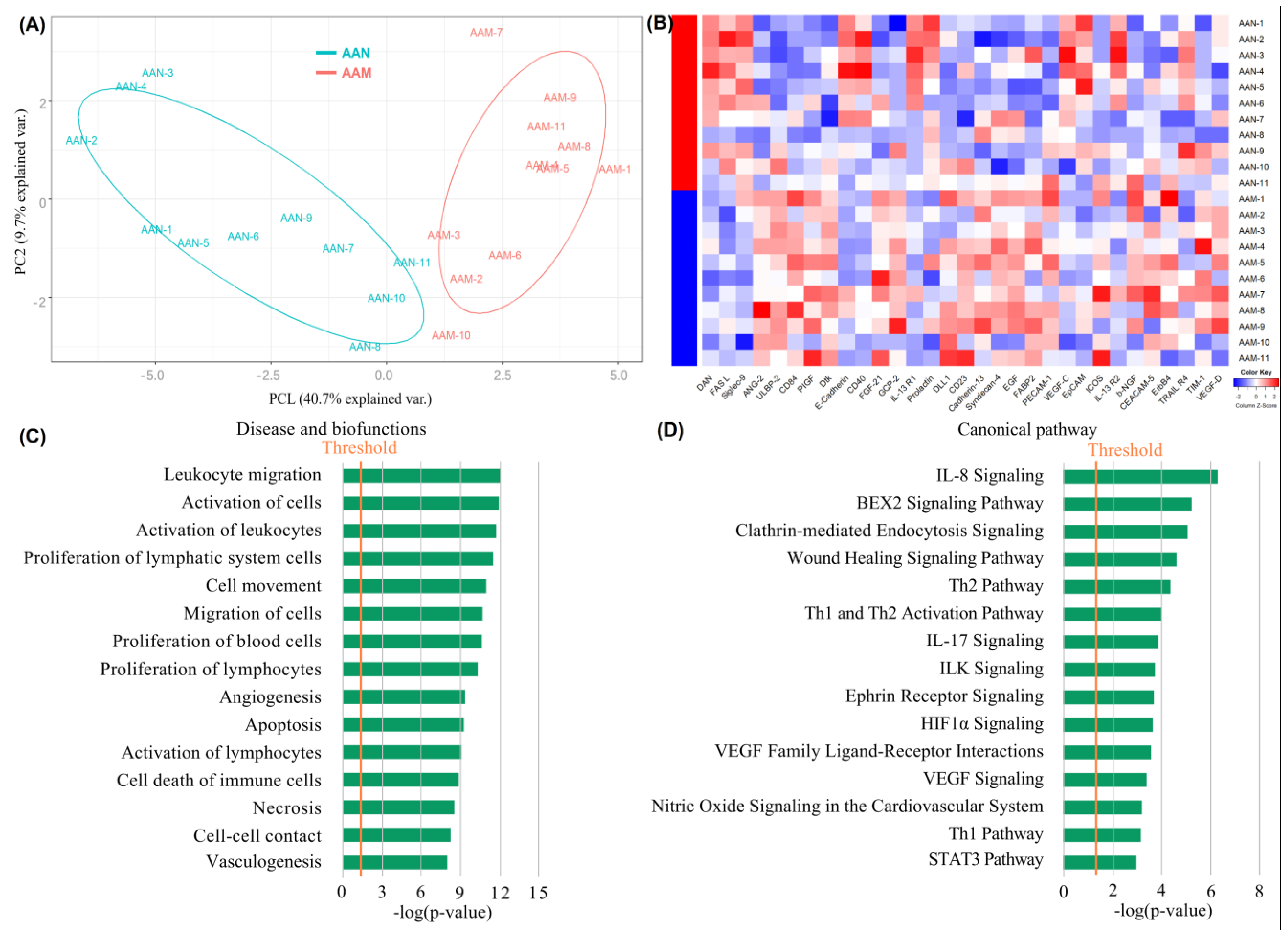
3.4. Exploration of Cytokine Expression Patterns in AAN and AAM Patients
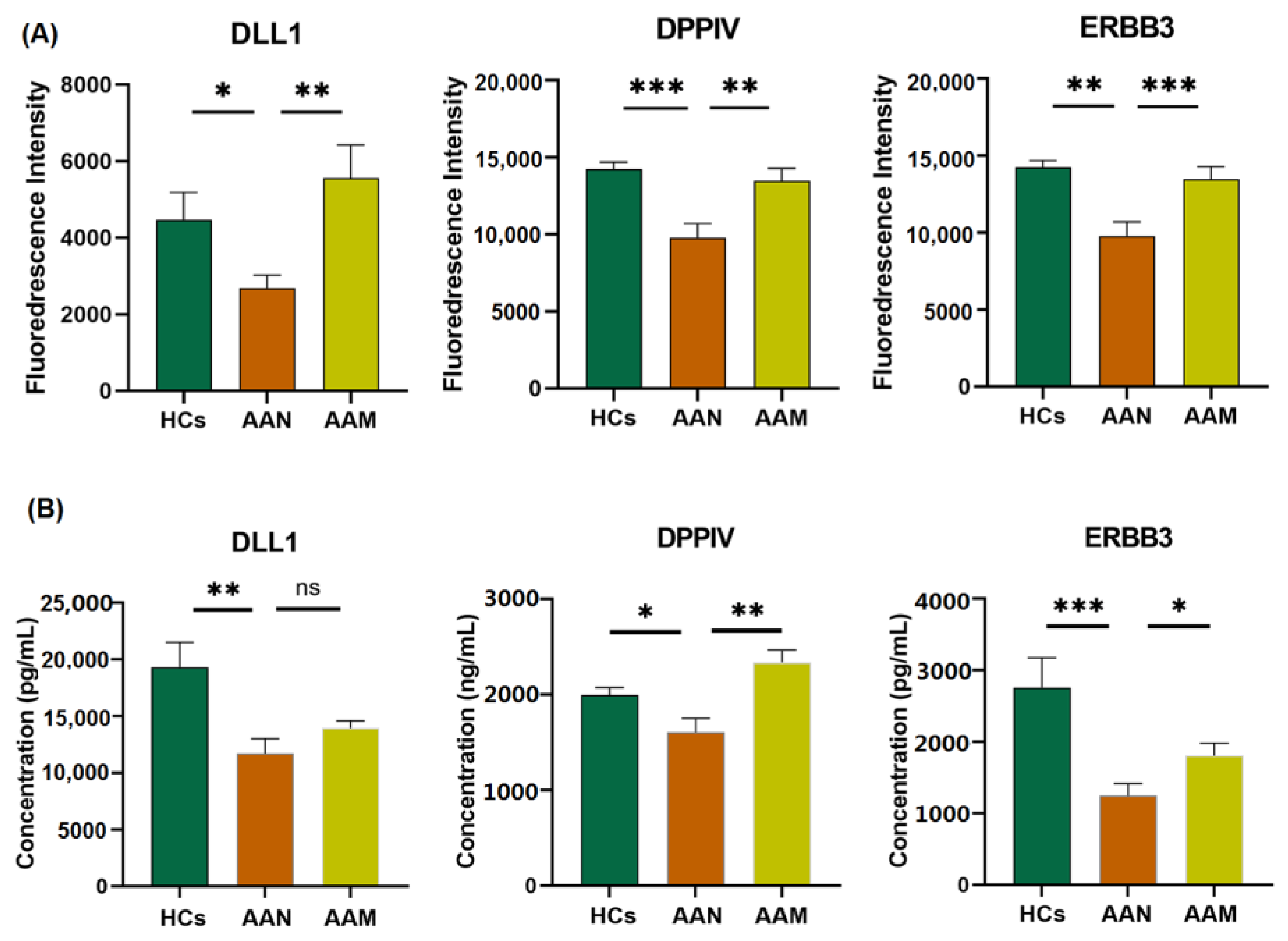
3.5. Identification of Differentially Expressed Proteins among AAM, AAN, and HC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schanzer, A.; Oderich, G.S. Management of Abdominal Aortic Aneurysms. N. Engl. J. Med. 2021, 385, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Oliver-Williams, C.; Sweeting, M.J.; Turton, G.; Parkin, D.; Cooper, D.; Rodd, C.; Thompson, S.G.; Earnshaw, J.J.; Gloucestershire and Swindon Abdominal Aortic Aneurysm Screening Programme. Lessons learned about prevalence and growth rates of abdominal aortic aneurysms from a 25-year ultrasound population screening programme. Br. J. Surg. 2018, 105, 68–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diseases, G.B.D.; Injuries, C. Global burden of 369 diseases and injuries in 204 countries and territories, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1204–1222. [Google Scholar]
- Lilja, F.; Wanhainen, A.; Mani, K. Changes in abdominal aortic aneurysm epidemiology. J. Cardiovasc. Surg. 2017, 58, 848–853. [Google Scholar] [CrossRef] [PubMed]
- Wanhainen, A.; Hultgren, R.; Linne, A.; Holst, J.; Gottsater, A.; Langenskiold, M.; Smidfelt, K.; Bjorck, M.; Svensjo, S.; Swedish Aneurysm Screening Study, G. Outcome of the Swedish Nationwide Abdominal Aortic Aneurysm Screening Program. Circulation 2016, 134, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Chaikof, E.L.; Dalman, R.L.; Eskandari, M.K.; Jackson, B.M.; Lee, W.A.; Mansour, M.A.; Mastracci, T.M.; Mell, M.; Murad, M.H.; Nguyen, L.L.; et al. The Society for Vascular Surgery practice guidelines on the care of patients with an abdominal aortic aneurysm. J. Vasc. Surg. 2018, 67, 2–77.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.J.; Prabhakar, A.M.; Kwolek, C.J. Current status of the treatment of infrarenal abdominal aortic aneurysms. Cardiovasc. Diagn. Ther. 2018, 8, S191–S199. [Google Scholar] [CrossRef] [Green Version]
- Golledge, J.; Moxon, J.V.; Singh, T.P.; Bown, M.J.; Mani, K.; Wanhainen, A. Lack of an effective drug therapy for abdominal aortic aneurysm. J. Intern. Med. 2019, 288, 6–22. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Kong, W. Cellular signaling in Abdominal Aortic Aneurysm. Cell. Signal. 2020, 70, 109575. [Google Scholar] [CrossRef]
- Raffort, J.; Lareyre, F.; Clement, M.; Hassen-Khodja, R.; Chinetti, G.; Mallat, Z. Monocytes and macrophages in abdominal aortic aneurysm. Nat. Rev. Cardiol. 2017, 14, 457–471. [Google Scholar] [CrossRef]
- Ning, X.; Ding, N.; Ballew, S.H.; Hicks, C.W.; Coresh, J.; Selvin, E.; Pankow, J.; Tang, W.; Matsushita, K. Diabetes, its duration, and the long-term risk of abdominal aortic aneurysm: The Atherosclerosis Risk in Communities (ARIC) Study. Atherosclerosis 2020, 313, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Sweeting, M.J.; Thompson, S.G.; Brown, L.C.; Powell, J.T.; Collaborators, R. Meta-analysis of individual patient data to examine factors affecting growth and rupture of small abdominal aortic aneurysms. Br. J. Surg. 2012, 99, 655–665. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Umemoto, T.; Group, A. Negative association of diabetes with rupture of abdominal aortic aneurysm. Diab. Vasc. Dis. Res. 2016, 13, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Xiong, J.; Wu, Z.; Chen, C.; Wei, Y.; Guo, W. Association between diabetes and prevalence and growth rate of abdominal aortic aneurysms: A meta-analysis. Int. J. Cardiol. 2016, 221, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Zagrapan, B.; Eilenberg, W.; Prausmueller, S.; Nawrozi, P.; Muench, K.; Hetzer, S.; Elleder, V.; Rajic, R.; Juster, F.; Martelanz, L.; et al. A Novel Diagnostic and Prognostic Score for Abdominal Aortic Aneurysms Based on D-Dimer and a Comprehensive Analysis of Myeloid Cell Parameters. Thromb. Haemost. 2019, 119, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Memon, A.; Zarrouk, M.; Ågren-Witteschus, S.; Sundquist, J.; Gottsäter, A.; Sundquist, K. Identification of novel diagnostic and prognostic biomarkers for abdominal aortic aneurysm. Eur. J. Prev. Cardiol. 2019, 27, 132–142. [Google Scholar] [CrossRef]
- Lee, R.; Cassimjee, I.; Huang, H.; Lapolla, P.; Ngetich, E.; Chandrashekar, A.; Charles, P.; Kessler, B.; Fischer, R.; Handa, A. Integrated Plasma and Tissue Proteomics Reveals Attractin Release by Intraluminal Thrombus of Abdominal Aortic Aneurysms and Improves Aneurysm Growth Prediction in Humans. Ann. Surg. 2022, 275, 1206–1211. [Google Scholar] [CrossRef]
- Wu, J.; Wang, W.; Chen, Z.; Xu, F.; Zheng, Y. Proteomics applications in biomarker discovery and pathogenesis for abdominal aortic aneurysm. Expert Rev. Proteom. 2021, 18, 305–314. [Google Scholar] [CrossRef]
- Wu, Q.Y.; Cheng, Z.; Zhou, Y.Z.; Zhao, Y.; Li, J.M.; Zhou, X.M.; Peng, H.L.; Zhang, G.S.; Liao, X.B.; Fu, X.M. A novel STAT3 inhibitor attenuates angiotensin II-induced abdominal aortic aneurysm progression in mice through modulating vascular inflammation and autophagy. Cell Death Dis. 2020, 11, 131. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.K.; Lu, G.; Jester, A.; Johnston, W.F.; Zhao, Y.; Hajzus, V.A.; Saadatzadeh, M.R.; Su, G.; Bhamidipati, C.M.; Mehta, G.S.; et al. Experimental abdominal aortic aneurysm formation is mediated by IL-17 and attenuated by mesenchymal stem cell treatment. Circulation 2012, 126, S38–S45. [Google Scholar] [CrossRef] [Green Version]
- Sharma, A.K.; Salmon, M.D.; Lu, G.; Su, G.; Pope, N.H.; Smith, J.R.; Weiss, M.L.; Upchurch, G.R., Jr. Mesenchymal Stem Cells Attenuate NADPH Oxidase-Dependent High Mobility Group Box 1 Production and Inhibit Abdominal Aortic Aneurysms. Arter. Thromb. Vasc. Biol. 2016, 36, 908–918. [Google Scholar]
- Zhang, M.; Sui, W.; Cheng, C.; Xue, F.; Tian, Z.; Cheng, J.; Zhang, J.; Zhang, T.; Zhang, J.; Wang, W.; et al. Erythropoietin promotes abdominal aortic aneurysms in mice through angiogenesis and inflammatory infiltration. Sci. Transl. Med. 2021, 13, eaaz4959. [Google Scholar] [CrossRef] [PubMed]
- Li, D.Y.; Busch, A.; Jin, H.; Chernogubova, E.; Pelisek, J.; Karlsson, J.; Sennblad, B.; Liu, S.; Lao, S.; Hofmann, P.; et al. H19 Induces Abdominal Aortic Aneurysm Development and Progression. Circulation 2018, 138, 1551–1568. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Lin, Y. The Mechanism of Asparagine Endopeptidase in the Progression of Malignant Tumors: A Review. Cells 2021, 10, 1153. [Google Scholar] [CrossRef]
- Davis, F.M.; Daugherty, A.; Lu, H.S. Updates of Recent Aortic Aneurysm Research. Arter. Thromb. Vasc. Biol. 2019, 39, e83–e90. [Google Scholar] [CrossRef] [PubMed]
- Solberg, R.; Smith, R.; Almlof, M.; Tewolde, E.; Nilsen, H.; Johansen, H.T. Legumain expression, activity and secretion are increased during monocyte-to-macrophage differentiation and inhibited by atorvastatin. Biol. Chem. 2015, 396, 71–80. [Google Scholar] [CrossRef]
- Pan, L.; Bai, P.; Weng, X.; Liu, J.; Chen, Y.; Chen, S.; Ma, X.; Hu, K.; Sun, A.; Ge, J. Legumain Is an Endogenous Modulator of Integrin alphavbeta3 Triggering Vascular Degeneration, Dissection, and Rupture. Circulation 2022, 145, 659–674. [Google Scholar] [CrossRef]
- Dattani, N.; Sayers, R.D.; Bown, M.J. Diabetes mellitus and abdominal aortic aneurysms: A review of the mechanisms underlying the negative relationship. Diab. Vasc. Dis. Res. 2018, 15, 367–374. [Google Scholar] [CrossRef] [Green Version]
- Raffort, J.; Lareyre, F.; Clement, M.; Hassen-Khodja, R.; Chinetti, G.; Mallat, Z. Diabetes and aortic aneurysm: Current state of the art. Cardiovasc. Res. 2018, 114, 1702–1713. [Google Scholar] [CrossRef] [Green Version]
- Kessler, V.; Klopf, J.; Eilenberg, W.; Neumayer, C.; Brostjan, C. AAA Revisited: A Comprehensive Review of Risk Factors, Management, and Hallmarks of Pathogenesis. Biomedicines 2022, 10, 94. [Google Scholar] [CrossRef]
- Xiu, M.X.; Liu, Y.M.; Kuang, B.H. The Role of DLLs in Cancer: A Novel Therapeutic Target. OncoTargets Ther. 2020, 13, 3881–3901. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Ren, P.; Nguyen, M.; Coselli, J.S.; Shen, Y.H.; LeMaire, S.A. Notch signaling in descending thoracic aortic aneurysm and dissection. PLoS ONE 2012, 7, e52833. [Google Scholar] [CrossRef] [PubMed]
- Biros, E.; Walker, P.J.; Nataatmadja, M.; West, M.; Golledge, J. Downregulation of transforming growth factor, beta receptor 2 and Notch signaling pathway in human abdominal aortic aneurysm. Atherosclerosis 2012, 221, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.J.; Redmond, E.M.; Gillespie, D.L.; Knight, P.A.; Cullen, J.P.; Cahill, P.A.; Morrow, D.J. Differential expression of Hedgehog/Notch and transforming growth factor-beta in human abdominal aortic aneurysms. J. Vasc. Surg. 2015, 62, 464–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubey, M.; Chhabra, N.F.; Gradinger, D.; Sanz-Moreno, A.; Lickert, H.; Przemeck, G.K.H.; Hrabe de Angelis, M. DLL1- and DLL4-Mediated Notch Signaling Is Essential for Adult Pancreatic Islet Homeostasis. Diabetes 2020, 69, 915–926. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, D.M.; Zhou, S.; Rebalka, I.A.; MacDonald, B.; Moradi, J.; Krause, M.P.; Al-Sajee, D.; Punthakee, Z.; Tarnopolsky, M.A.; Hawke, T.J. Decreased Satellite Cell Number and Function in Humans and Mice with Type 1 Diabetes Is the Result of Altered Notch Signaling. Diabetes 2016, 65, 3053–3061. [Google Scholar] [CrossRef] [Green Version]
- Min, X.H.; Yu, T.; Qing, Q.; Yuan, Y.H.; Zhong, W.; Chen, G.C.; Zhao, L.N.; Deng, N.; Zhang, L.F.; Chen, Q.K. Abnormal differentiation of intestinal epithelium and intestinal barrier dysfunction in diabetic mice associated with depressed Notch/NICD transduction in Notch/Hes1 signal pathway. Cell Biol. Int. 2014, 38, 1194–1204. [Google Scholar] [CrossRef]
- Mentlein, R. Dipeptidyl-peptidase IV (CD26)--role in the inactivation of regulatory peptides. Regul. Pept. 1999, 85, 9–24. [Google Scholar] [CrossRef]
- Lindquist Liljeqvist, M.; Eriksson, L.; Villard, C.; Lengquist, M.; Kronqvist, M.; Hultgren, R.; Roy, J. Dipeptidyl peptidase-4 is increased in the abdominal aortic aneurysm vessel wall and is associated with aneurysm disease processes. PLoS ONE 2020, 15, e0227889. [Google Scholar] [CrossRef] [Green Version]
- Kohashi, K.; Hiromura, M.; Mori, Y.; Terasaki, M.; Watanabe, T.; Kushima, H.; Shinmura, K.; Tomoyasu, M.; Nagashima, M.; Hirano, T. A Dipeptidyl Peptidase-4 Inhibitor but not Incretins Suppresses Abdominal Aortic Aneurysms in Angiotensin II-Infused Apolipoprotein E-Null Mice. J. Atheroscler. Thromb. 2016, 23, 441–454. [Google Scholar] [CrossRef] [Green Version]
- Stoian, A.P.; Sachinidis, A.; Stoica, R.A.; Nikolic, D.; Patti, A.M.; Rizvi, A.A. The efficacy and safety of dipeptidyl peptidase-4 inhibitors compared to other oral glucose-lowering medications in the treatment of type 2 diabetes. Metabolism 2020, 109, 154295. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, J.; Nargis, T.; Tantia, O.; Ghosh, S.; Chakrabarti, P. Increased Plasma Dipeptidyl Peptidase-4 (DPP4) Activity Is an Obesity-Independent Parameter for Glycemic Deregulation in Type 2 Diabetes Patients. Front. Endocrinol. 2019, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Gao, Y.; Baskota, A.; Chen, T.; Ran, X.; Tian, H. Increased plasma DPP4 activity is predictive of prediabetes and type 2 diabetes onset in Chinese over a four-year period: Result from the China National Diabetes and Metabolic Disorders Study. J. Clin. Endocrinol. Metab. 2014, 99, E2330–E2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hafeez, U.; Parslow, A.C.; Gan, H.K.; Scott, A.M. New insights into ErbB3 function and therapeutic targeting in cancer. Expert Rev. Anticancer. Ther. 2020, 20, 1057–1074. [Google Scholar] [CrossRef]
- Nguyen, D.Q.; Hoang, D.H.; Nguyen Vo, T.T.; Huynh, V.; Ghoda, L.; Marcucci, G.; Nguyen, L.X.T. The role of ErbB3 binding protein 1 in cancer: Friend or foe? J. Cell. Physiol. 2018, 233, 9110–9120. [Google Scholar] [CrossRef]
- Li, B.; Wang, Z.; Chen, R.; Hong, J.; Wu, Q.; Hu, J.; Hu, Z.; Zhang, M. Up regulation of isoleucyl-tRNA synthetase promotes vascular smooth muscle cells dysfunction via p38 MAPK/PI3K signaling pathways. Life Sci. 2019, 224, 51–57. [Google Scholar] [CrossRef]
- Ni, X.Q.; Zhang, Y.R.; Jia, L.X.; Lu, W.W.; Zhu, Q.; Ren, J.L.; Chen, Y.; Zhang, L.S.; Liu, X.; Yu, Y.R.; et al. Inhibition of Notch1-mediated inflammation by intermedin protects against abdominal aortic aneurysm via PI3K/Akt signaling pathway. Aging 2021, 13, 5164–5184. [Google Scholar] [CrossRef]
- Wang, D.; Pan, G. The Association between rs2292239 Polymorphism in ERBB3 Gene and Type 1 Diabetes: A Meta-Analysis. Biomed. Res. Int. 2019, 2019, 7689642. [Google Scholar] [CrossRef] [Green Version]
- Hemi, R.; Paz, K.; Wertheim, N.; Karasik, A.; Zick, Y.; Kanety, H. Transactivation of ErbB2 and ErbB3 by tumor necrosis factor-alpha and anisomycin leads to impaired insulin signaling through serine/threonine phosphorylation of IRS proteins. J. Biol. Chem. 2002, 277, 8961–8969. [Google Scholar] [CrossRef] [Green Version]
- Hemi, R.; Yochananov, Y.; Barhod, E.; Kasher-Meron, M.; Karasik, A.; Tirosh, A.; Kanety, H. p38 mitogen-activated protein kinase-dependent transactivation of ErbB receptor family: A novel common mechanism for stress-induced IRS-1 serine phosphorylation and insulin resistance. Diabetes 2011, 60, 1134–1145. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Discovery Cohort | Validation Cohort 1 | Validation Cohort 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| HC | AAN | AAM | p-Value | HC | AAA | p-Value | HC | AAN | AAM | p-Value | |
| Number | 10 | 11 | 11 | — | 60 | 94 | — | 10 | 11 | 11 | — |
| Sex (male/female) | 8/2 | 10/1 | 10/1 | 0.688 | 51/9 | 81/13 | 0.819 | 8/2 | 10/1 | 10/1 | 0.688 |
| Age (years) | 68 ± 9 | 69 ± 8 | 73 ± 8 | 0.309 | 68 ± 7 | 69 ± 10 | 0.438 | 67 ± 3 | 69 ± 7 | 70 ± 7 | 0.443 |
| Diameter (mm) | — | 60 ± 16 | 47 ± 10 | 0.019 | — | 53 ± 13 | — | — | 51 ± 8 | 41 ± 7 | 0.003 |
| Hypertension | 4 | 7 | 7 | 0.458 | 39 | 73 | 0.085 | 7 | 8 | 7 | 0.895 |
| Type 2 Diabetes | 0 | 0 | 11 | — | 11 | 17 | 0.969 | 0 | 0 | 11 | — |
| Dyslipidemia | 3 | 8 | 7 | 0.119 | 28 | 58 | 0.071 | 4 | 8 | 7 | 0.293 |
| Gene Symbol | UniPort ID | Ratio (AAA/HCs) | p-Value |
|---|---|---|---|
| MMP-7 | P09237 | 2.72 | 1.82 × 10−6 |
| Clusterin | P10909 | 0.37 | 1.09 × 10−7 |
| RGM-B | Q6NW40 | 0.61 | 2.11 × 10−6 |
| Legumain | Q99538 | 1.51 | 0.0003 |
| Epo R | P19235 | 0.64 | 5.72 × 10−5 |
| JAM-A | Q9Y624 | 1.92 | 0.0013 |
| NRG1-b1 | Q02297 | 0.52 | 0.0002 |
| Cathepsin S | P25774 | 0.66 | 0.0003 |
| DAN | P41271 | 0.61 | 0.0011 |
| OSM | P13725 | 0.24 | 0.0046 |
| IL-13 R1 | P78552 | 0.59 | 0.0011 |
| ACE-2 | Q9BYF1 | 0.53 | 0.0001 |
| IL-1 RII | P27930 | 0.45 | 0.0001 |
| IL-13 R2 | Q14627 | 0.40 | 0.0075 |
| RANTES | P13501 | 1.60 | 0.0011 |
| FAS L | P48023 | 0.62 | 0.0080 |
| MICB | Q29980 | 0.59 | 0.0096 |
| TARC | Q92583 | 2.14 | 0.0039 |
| TPO | P40225 | 0.59 | 0.0348 |
| Galectin-7 | P47929 | 0.31 | 0.0276 |
| Syndecan-3 | O75056 | 0.35 | 0.0145 |
| ULBP-2 | Q9BZM5 | 2.07 | 0.0082 |
| E-Cadherin | P12830 | 0.66 | 0.0425 |
| GH 1 | P01241 | 0.59 | 0.0105 |
| TRAIL R4 | Q9UBN6 | 0.59 | 0.0332 |
| GITR L | Q9UNG2 | 0.49 | 0.0465 |
| B7-1 | P33681 | 0.63 | 0.0185 |
| LAP (TGFb1) | P01137 | 1.95 | 0.0144 |
| ADAMTS13 | Q76LX8 | 0.57 | 0.0345 |
| PECAM-1 | P16284 | 1.67 | 0.0230 |
| GROa | P09341 | 0.46 | 0.0068 |
| FABP2 | P12104 | 1.63 | 0.0427 |
| ANG-4 | Q9Y264 | 0.60 | 0.0332 |
| Granulysin | P22749 | 0.59 | 0.0280 |
| PDGF-AB | Q08CD2 | 2.21 | 0.0142 |
| WISP-1 | Q96FT7 | 0.63 | 0.0420 |
| EGF | P01133 | 1.98 | 0.0396 |
| PDGF-BB | P01127 | 1.51 | 0.0339 |
| BAFF | Q9Y275 | 0.50 | 0.0491 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Wang, W.; Xie, T.; Chen, Z.; Zhou, L.; Song, X.; Kan, H.; Lv, Y.; Wu, L.; Li, F.; et al. Identification of Novel Plasma Biomarkers for Abdominal Aortic Aneurysm by Protein Array Analysis. Biomolecules 2022, 12, 1853. https://doi.org/10.3390/biom12121853
Wu J, Wang W, Xie T, Chen Z, Zhou L, Song X, Kan H, Lv Y, Wu L, Li F, et al. Identification of Novel Plasma Biomarkers for Abdominal Aortic Aneurysm by Protein Array Analysis. Biomolecules. 2022; 12(12):1853. https://doi.org/10.3390/biom12121853
Chicago/Turabian StyleWu, Jianqiang, Wei Wang, Ting Xie, Zhaoran Chen, Lei Zhou, Xiaohong Song, Haoxuan Kan, Yanze Lv, Lianglin Wu, Fangda Li, and et al. 2022. "Identification of Novel Plasma Biomarkers for Abdominal Aortic Aneurysm by Protein Array Analysis" Biomolecules 12, no. 12: 1853. https://doi.org/10.3390/biom12121853