
Neuronal Firing and Glutamatergic Synapses in the Substantia Nigra Pars Reticulata of LRRK2-G2019S Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Brain Slice Electrophysiology
2.3. Chemicals and Drugs
2.4. Statistical Analysis
3. Results
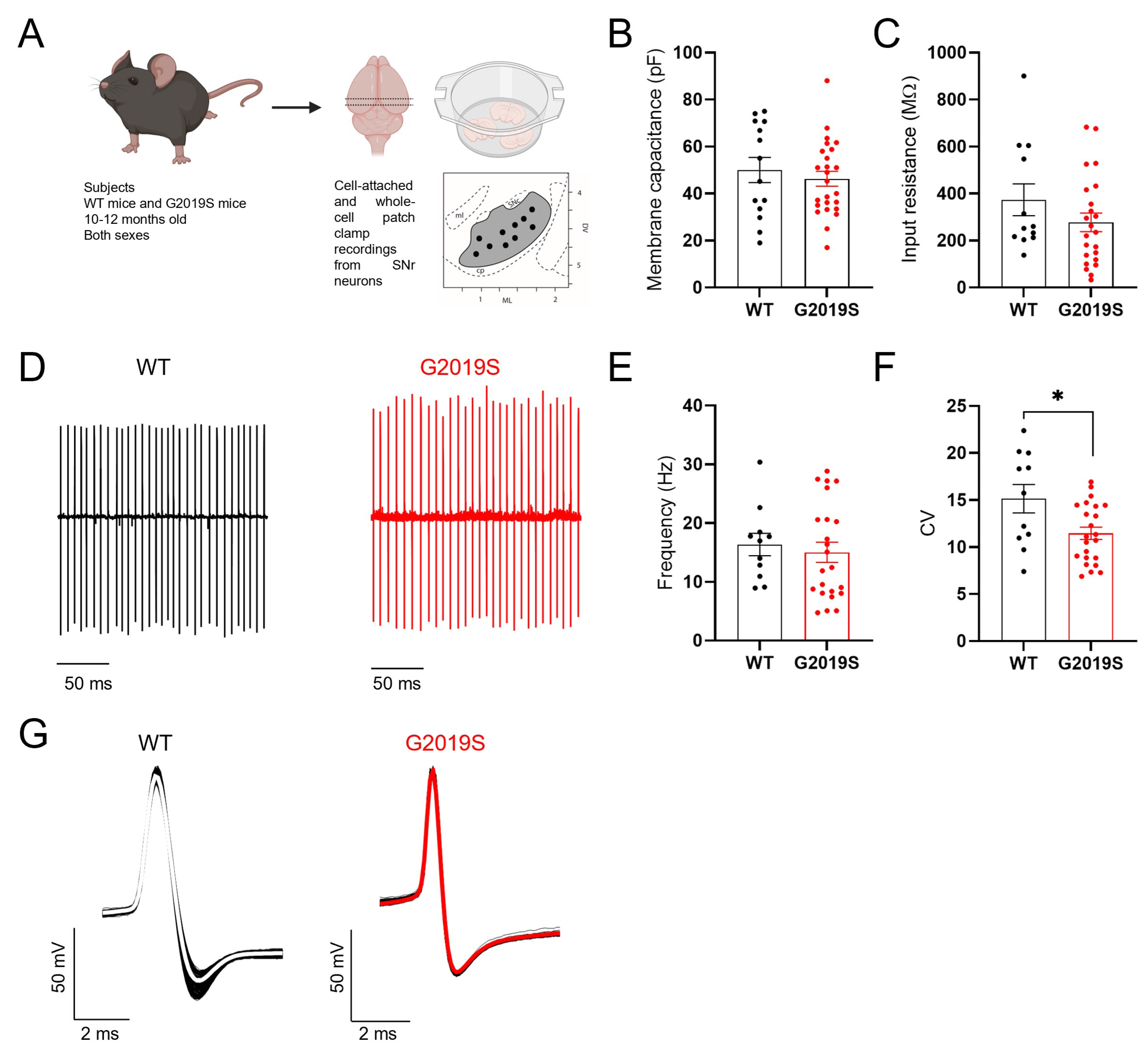
3.1. Intrinsic Properties of SNr GABAergic Neurons from Middle-Aged WT and G2019S Mice
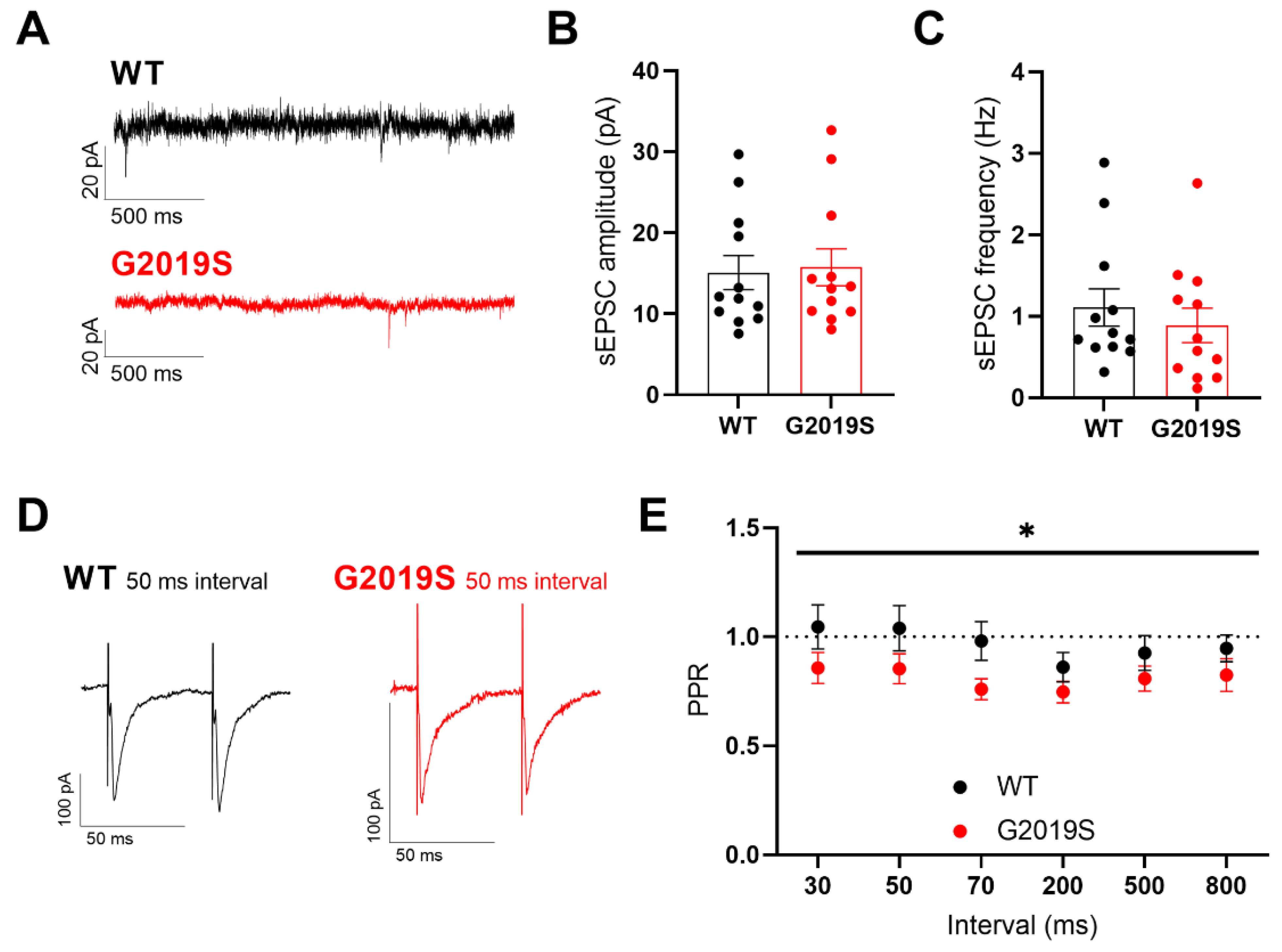
3.2. Glutamatergic Synaptic Transmission in SNr Neurons from Middle-Aged WT and G2019S Mice
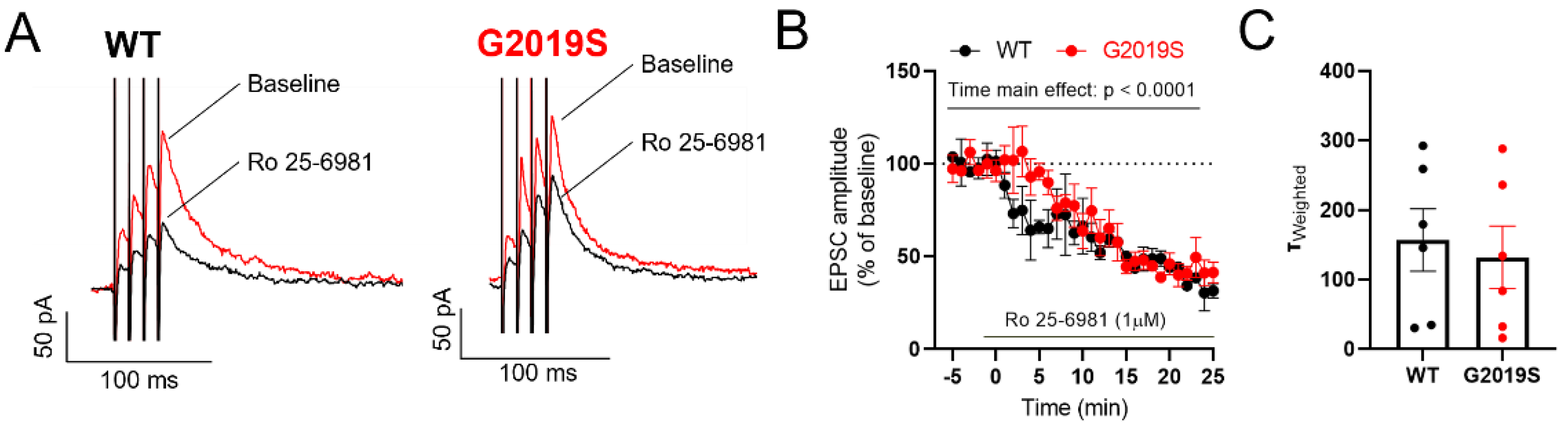
3.3. NMDARs in SNr Neurons from Middle-Aged G2019S and WT Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez-Rodriguez, P.; Zampese, E.; Surmeier, D.J. Selective neuronal vulnerability in Parkinson’s disease. Prog. Brain Res. 2020, 252, 61–89. [Google Scholar] [PubMed]
- Pirkevi, C.; Lesage, S.; Brice, A.; Basak, A.N. From genes to proteins in mendelian Parkinson’s disease: An overview. Anat. Rec. 2009, 292, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Yue, M.; Hinkle, K.M.; Davies, P.; Trushina, E.; Fiesel, F.C.; Christenson, T.A.; Schroeder, A.S.; Zhang, L.; Bowles, E.; Behrouz, B.; et al. Progressive dopaminergic alterations and mitochondrial abnormalities in LRRK2 G2019S knock-in mice. Neurobiol. Dis. 2015, 78, 172–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healy, D.G.; Falchi, M.; O’Sullivan, S.S.; Bonifati, V.; Durr, A.; Bressman, S.; Brice, A.; Aasly, J.; Zabetian, C.P.; Goldwurm, S.; et al. Phenotype, genotype, and worldwide genetic penetrance of LRRK2-associated Parkinson’s disease: A case-control study. Lancet Neurol. 2008, 7, 583–590. [Google Scholar] [CrossRef] [Green Version]
- Di Maio, R.; Hoffman, E.K.; Rocha, E.M.; Keeney, M.T.; Sanders, L.H.; De Miranda, B.R.; Zharikov, A.; Van Laar, A.; Stepan, A.F. LRRK2 activation in idiopathic Parkinson’s disease. Sci. Transl. Med. 2018, 10, eaar5429. [Google Scholar] [CrossRef] [Green Version]
- Mallet, N.; Delgado, L.; Chazalon, M.; Miguelez, C.; Baufreton, J. Cellular and Synaptic Dysfunctions in Parkinson’s Disease: Stepping out of the Striatum. Cells 2019, 8, 1005. [Google Scholar] [CrossRef] [Green Version]
- McGregor, M.M.; Nelson, A.B. Circuit Mechanisms of Parkinson’s Disease. Neuron 2019, 101, 1042–1056. [Google Scholar] [CrossRef] [Green Version]
- McElvain, L.E.; Chen, Y.; Moore, J.D.; Brigidi, G.S.; Bloodgood, B.L.; Lim, B.K.; Costa, R.M.; Kleinfeld, D. Specific populations of basal ganglia output neurons target distinct brain stem areas while collateralizing throughout the diencephalon. Neuron 2021, 109, 1721–1738.e4. [Google Scholar] [CrossRef]
- Adeosun, S.O.; Hou, X.; Zheng, B.; Melrose, H.L.; Mosley, T.; Wang, J.M. Human LRRK2 G2019S mutation represses post-synaptic protein PSD95 and causes cognitive impairment in transgenic mice. Neurobiol. Learn Mem. 2017, 142, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Melrose, H.L.; Dachsel, J.C.; Behrouz, B.; Lincoln, S.J.; Yue, M.; Hinkle, K.M.; Kent, C.B.; Korvatska, E.; Taylor, J.P.; Witten, L.; et al. Impaired dopaminergic neurotransmission and microtubule-associated protein tau alterations in human LRRK2 transgenic mice. Neurobiol. Dis. 2010, 40, 503–517. [Google Scholar] [CrossRef]
- Winner, B.; Melrose, H.L.; Zhao, C.; Hinkle, K.M.; Yue, M.; Kent, C.; Braithwaite, A.T.; Ogholikhan, S.; Aigner, R.; Winkler, J.; et al. Adult neurogenesis and neurite outgrowth are impaired in LRRK2 G2019S mice. Neurobiol. Dis. 2011, 41, 706–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matikainen-Ankney, B.A.; Kezunovic, N.; Mesias, R.E.; Tian, Y.; Williams, F.M.; Huntley, G.W.; Benson, D.L. Altered Development of Synapse Structure and Function in Striatum Caused by Parkinson's Disease-Linked LRRK2-G2019S Mutation. J. Neurosci 2016, 36, 7128–7141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skiteva, O.; Yao, N.; Sitzia, G.; Chergui, K. LRRK2-G2019S mice display alterations in glutamatergic synaptic transmission in midbrain dopamine neurons. J. Neurochem. 2022, 161, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Skelton, P.D.; Tokars, V.; Parisiadou, L. LRRK2 at Striatal Synapses: Cell-Type Specificity and Mechanistic Insights. Cells 2022, 11, 169. [Google Scholar] [CrossRef]
- Ibañez-Sandoval, O.; Hernández, A.; Florán, B.; Galarraga, E.; Tapia, D.; Valdiosera, R.; Erlij, D.; Aceves, J.; Bargas, J. Control of the subthalamic innervation of substantia nigra pars reticulata by D1 and D2 dopamine receptors. J. Neurophysiol. 2006, 95, 1800–1811. [Google Scholar] [CrossRef] [Green Version]
- Qin, Q.; Zhi, L.T.; Li, X.T.; Yue, Z.Y.; Li, G.Z.; Zhang, H. Effects of LRRK2 Inhibitors on Nigrostriatal Dopaminergic Neurotransmission. CNS Neurosci. Ther. 2017, 23, 162–173. [Google Scholar] [CrossRef]
- Sitzia, G.; Mantas, I.; Zhang, X.; Svenningsson, P.; Chergui, K. NMDA receptors are altered in the substantia nigra pars reticulata and their blockade ameliorates motor deficits in experimental parkinsonism. Neuropharmacology 2020, 174, 108136. [Google Scholar] [CrossRef]
- Yao, N.; Skiteva, O.; Zhang, X.; Svenningsson, P.; Chergui, K. Ketamine and its metabolite (2R,6R)-hydroxynorketamine induce lasting alterations in glutamatergic synaptic plasticity in the mesolimbic circuit. Mol. Psychiatry 2018, 23, 2066–2077. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Pinto, M.J.; Kanodia, H.; Falasconi, A.; Sigrist, M.; Esposito, M.S.; Arber, S. Functional diversity for body actions in the mesencephalic locomotor region. Cell 2021, 184, 4564–4578.e18. [Google Scholar] [CrossRef]
- Zhou, F.M.; Lee, C.R. Intrinsic and integrative properties of substantia nigra pars reticulata neurons. Neuroscience 2011, 198, 69–94. [Google Scholar] [CrossRef]
- Dupuis, J.P.; Feyder, M.; Miguelez, C.; Garcia, L.; Morin, S.; Choquet, D.; Hosy, E.; Bezard, E.; Fisone, G.; Bioulac, B.H.; et al. Dopamine-Dependent Long-Term Depression at Subthalamo-Nigral Synapses Is Lost in Experimental Parkinsonism. J. Neurosci. 2013, 33, 14331–14341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, S.; Wei, W.; Zhou, F.-M. Molecular and functional differences in voltage-activated sodium currents between GABA projection neurons and dopamine neurons in the substantia nigra. J. Neurophysiol. 2011, 106, 3019–3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.R.; Tepper, J.M. Morphological and physiological properties of parvalbumin- and calretinin-containing gamma-aminobutyric acidergic neurons in the substantia nigra. J. Comp. Neurol. 2007, 500, 958–972. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, G.; Tan, K.R. Synergistic Nigral Output Pathways Shape Movement. Cell Rep. 2019, 27, 2184–2198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramonet, D.; Daher, J.P.; Lin, B.M.; Stafa, K.; Kim, J.; Banerjee, R.; Westerlund, M.; Pletnikova, O.; Glauser, L.; Yang, L.; et al. Dopaminergic neuronal loss, reduced neurite complexity and autophagic abnormalities in transgenic mice expressing G2019S mutant LRRK2. PLoS ONE 2011, 6, e18568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, J.; Bang, Y.; Choi, J.H.; Han, A.; Kwon, M.S.; Liu, K.H.; Choi, H.J. LRRK2 G2019S Induces Anxiety/Depression-like Behavior before the Onset of Motor Dysfunction with 5-HT(1A) Receptor Upregulation in Mice. J. Neurosci. 2018, 38, 1611–1621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matikainen-Ankney, B.A.; Kezunovic, N.; Menard, C.; Flanigan, M.E.; Zhong, Y.; Russo, S.J.; Benson, D.L.; Huntley, G.W. Parkinson’s Disease-Linked LRRK2-G2019S Mutation Alters Synaptic Plasticity and Promotes Resilience to Chronic Social Stress in Young Adulthood. J. Neurosci. 2018, 38, 9700–9711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guevara, C.A.; Matikainen-Ankney, B.A.; Kezunovic, N.; LeClair, K.; Conway, A.P.; Menard, C.; Flanigan, M.E.; Pfau, M.; Russo, S.J.; Benson, D.L.; et al. LRRK2 mutation alters behavioral, synaptic, and nonsynaptic adaptations to acute social stress. J. Neurophysiol. 2020, 123, 2382–2389. [Google Scholar] [CrossRef]
- Foster, N.N.; Barry, J.; Korobkova, L.; Garcia, L.; Gao, L.; Becerra, M.; Sherafat, Y.; Peng, B.; Li, X.; Choi, J.H.; et al. The mouse cortico-basal ganglia-thalamic network. Nature 2021, 598, 188–194. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Measurement | WT Mice n = 11 Neurons | G2019S Mice n = 15 Neurons | Test | p Value |
|---|---|---|---|---|
| Interspike membrane potential (mV) | −51.92 ± 1.39 | −50.56 ± 0.79 | Unpaired t-test | 0.3711 |
| AP amplitude (mV) | 62.71 ± 2.33 | 57.80 ± 1.48 | Mann-Whitney | 0.2171 |
| AP half width (ms) | 0.53 ± 0.04 | 0.50 ± 0.03 | Unpaired t-test | 0.592 |
| AP threshold (mV) | −38.80 ± 0.95 | −38.19 ± 1.18 | Unpaired t-test | 0.7088 |
| AHP * amplitude (mV) | −47.89 ± 1.95 | −46.92 ± 1.13 | Unpaired t-test | 0.6521 |
| AHP * duration (ms) | 1.90 ± 0.26 | 2.20 ± 0.26 | Mann-Whitney | 0.4648 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sitzia, G.; Skiteva, O.; Chergui, K. Neuronal Firing and Glutamatergic Synapses in the Substantia Nigra Pars Reticulata of LRRK2-G2019S Mice. Biomolecules 2022, 12, 1635. https://doi.org/10.3390/biom12111635
Sitzia G, Skiteva O, Chergui K. Neuronal Firing and Glutamatergic Synapses in the Substantia Nigra Pars Reticulata of LRRK2-G2019S Mice. Biomolecules. 2022; 12(11):1635. https://doi.org/10.3390/biom12111635
Chicago/Turabian StyleSitzia, Giacomo, Olga Skiteva, and Karima Chergui. 2022. "Neuronal Firing and Glutamatergic Synapses in the Substantia Nigra Pars Reticulata of LRRK2-G2019S Mice" Biomolecules 12, no. 11: 1635. https://doi.org/10.3390/biom12111635