
Effects of Psychotropic Medication on Somatic Sterol Biosynthesis of Adult Mice
 ,
,  , and
, and 
Abstract
:1. Introduction
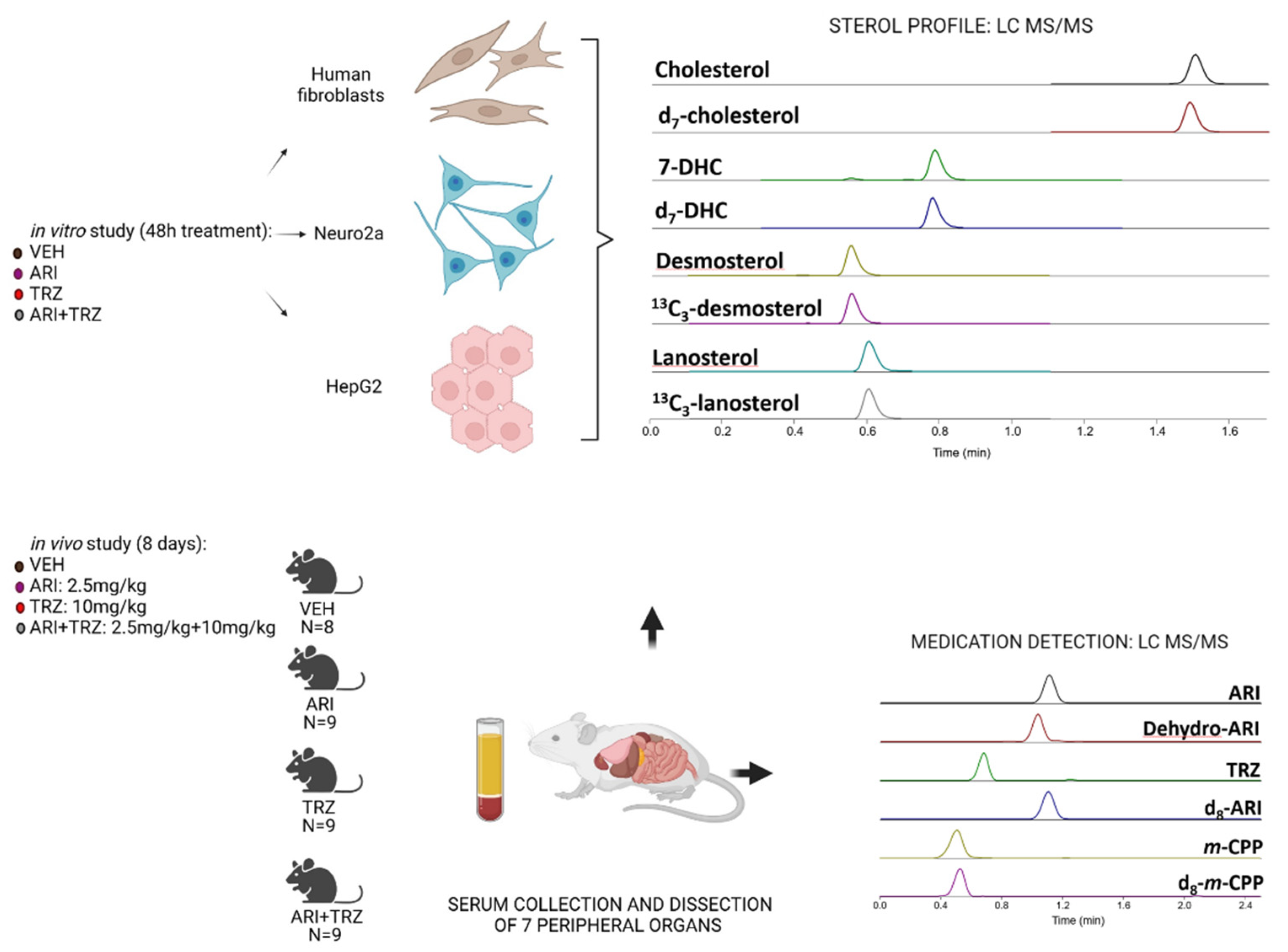
2. Materials and Methods
2.1. Reagents
2.2. Aripiprazole and Trazodone In Vivo Treatment
2.3. Tissue Dissection and Preparation for Sterol Analysis
2.4. LC-MS/MS Sterol Measurements
2.5. ARI and TRZ Measurements
2.6. Cell Cultures
2.7. Statistical Analyses
3. Results
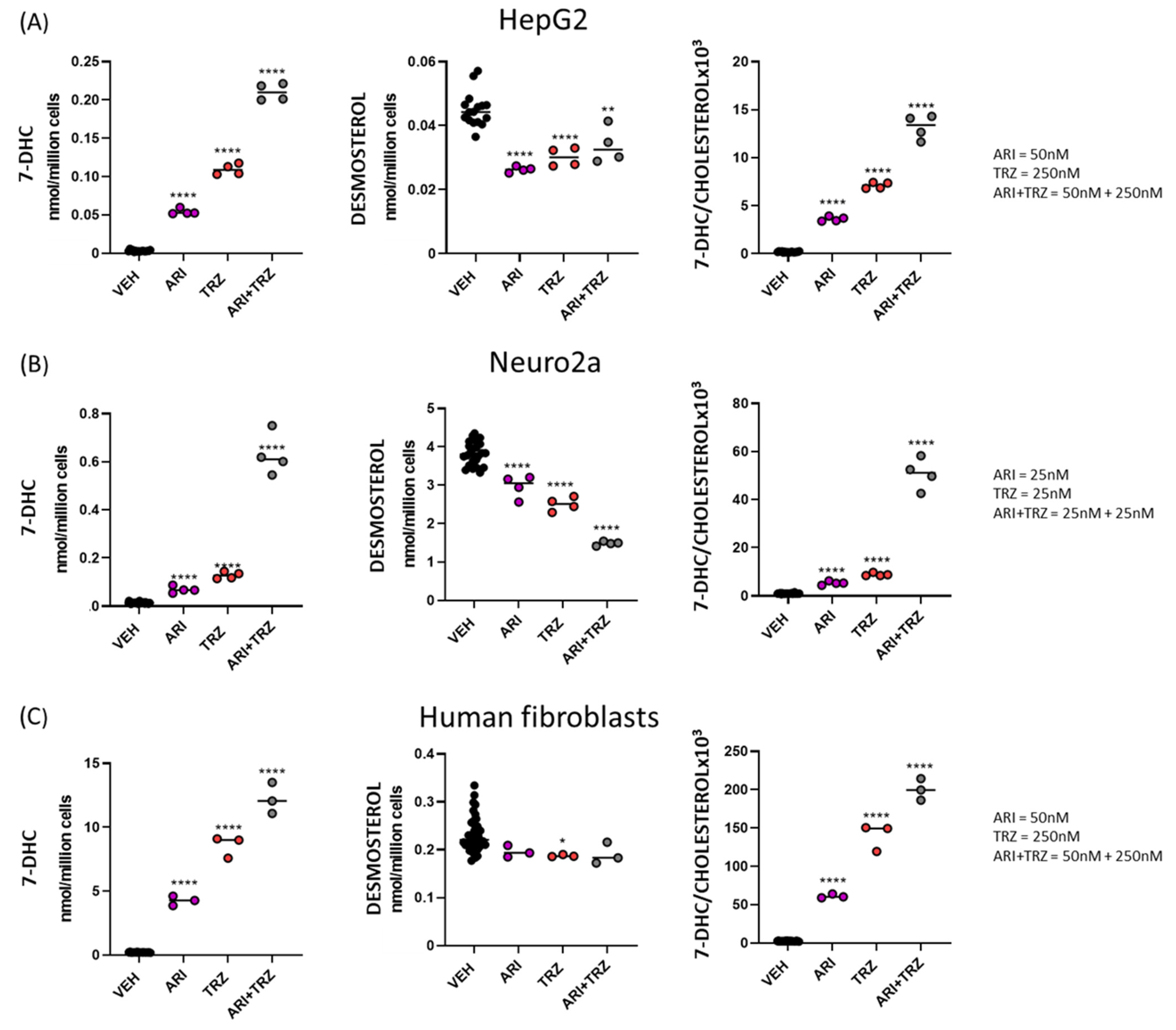
3.1. ARI + TRZ Polypharmacy Increases 7-DHC Levels in HepG2, Neuro2a and Human Fibroblast Cell Cultures Compared to Single Drug Treatment
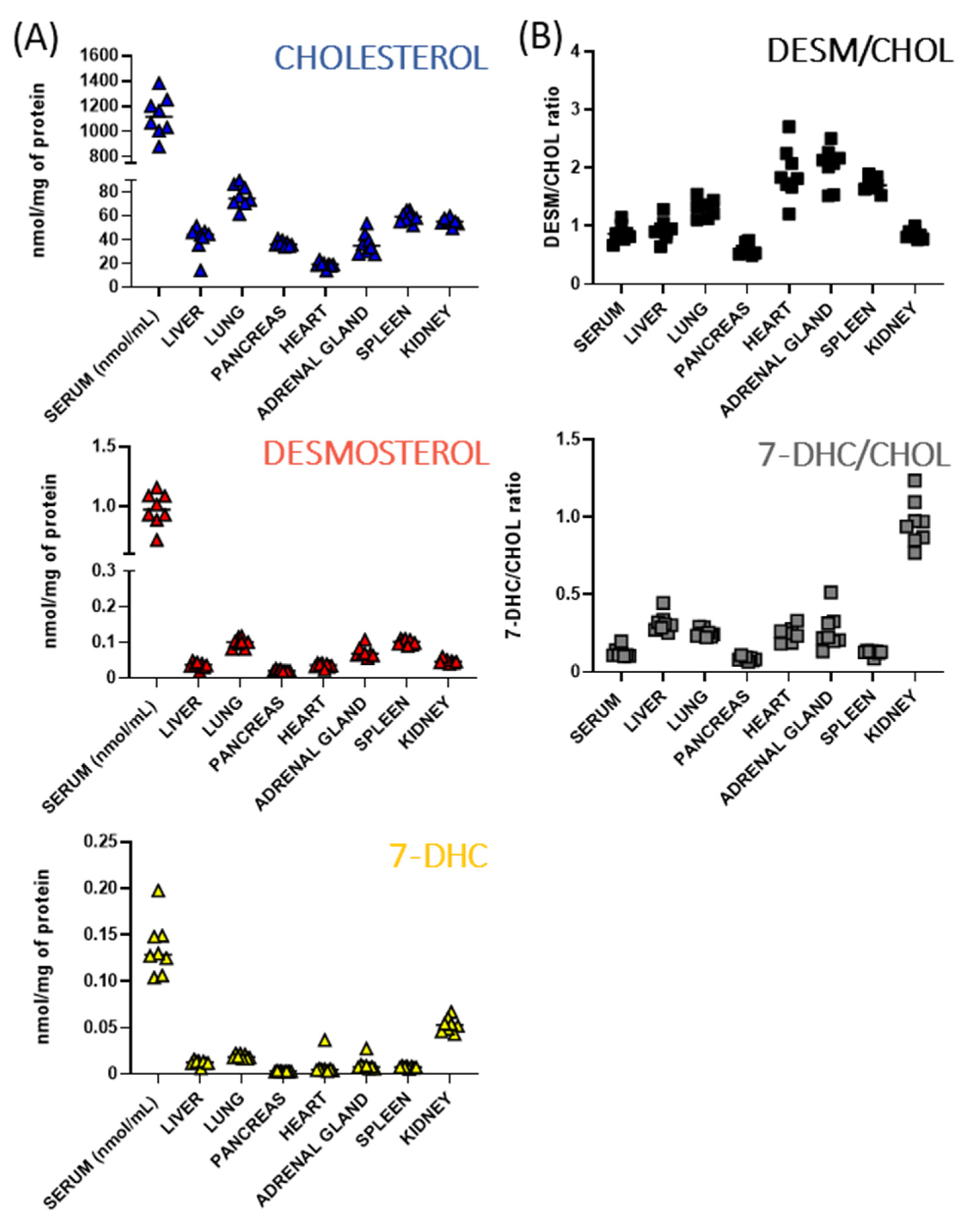
3.2. Baseline Sterols Levels Differ across Peripheral Tissues of Adult Mice
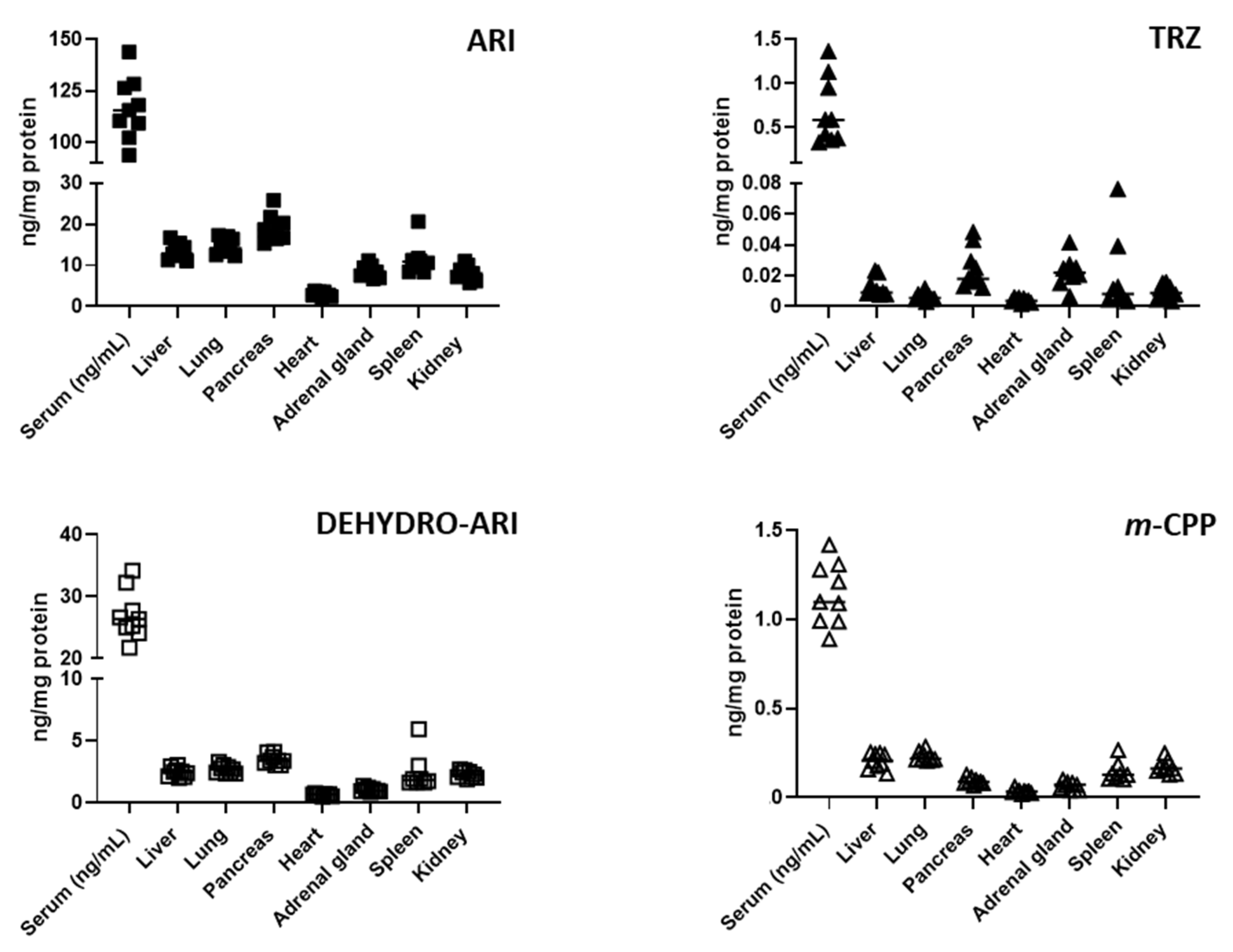
3.3. ARI, TRZ, and Their Metabolites Are Detectable in the Serum and Peripheral Organs of Treated Mice
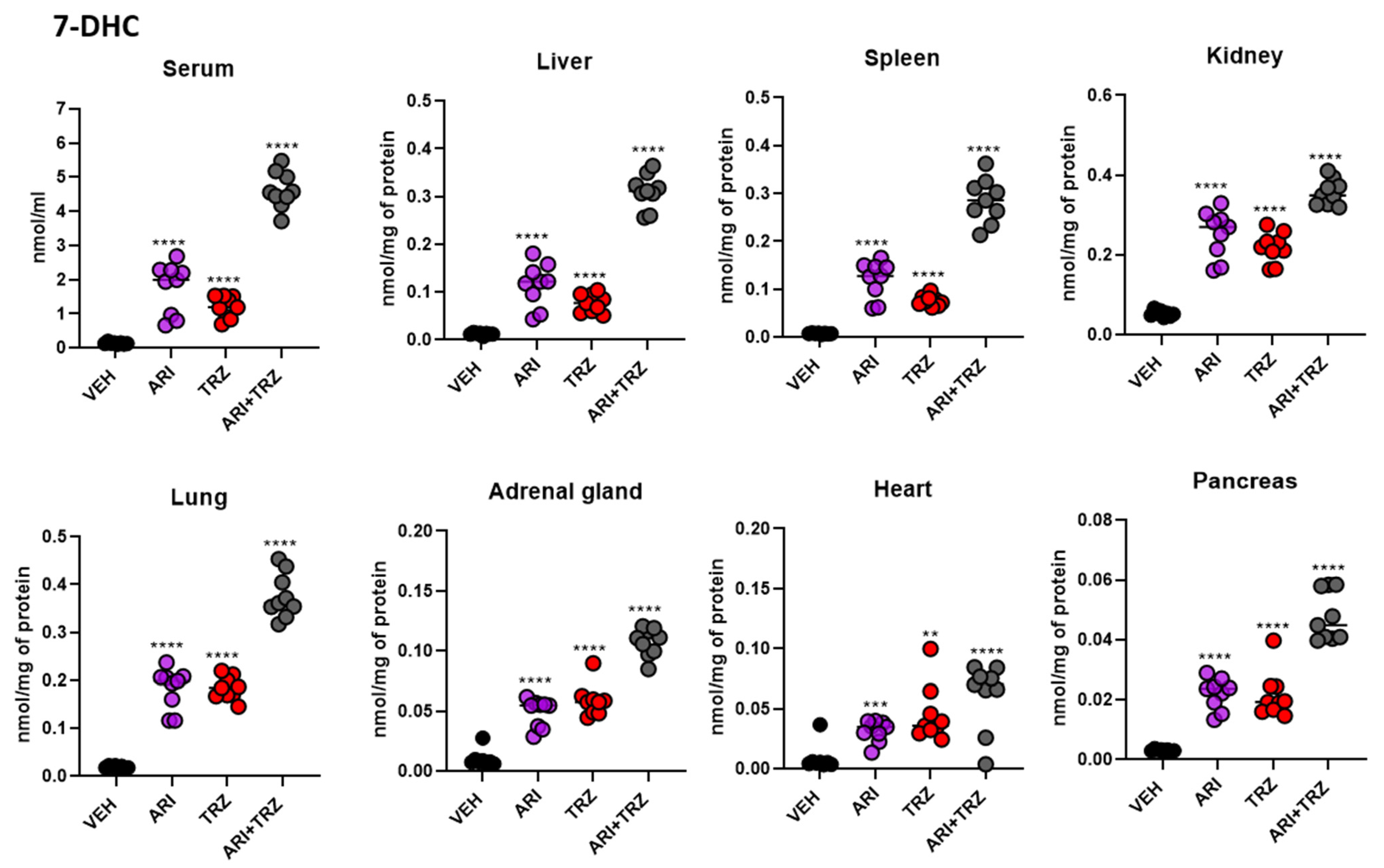
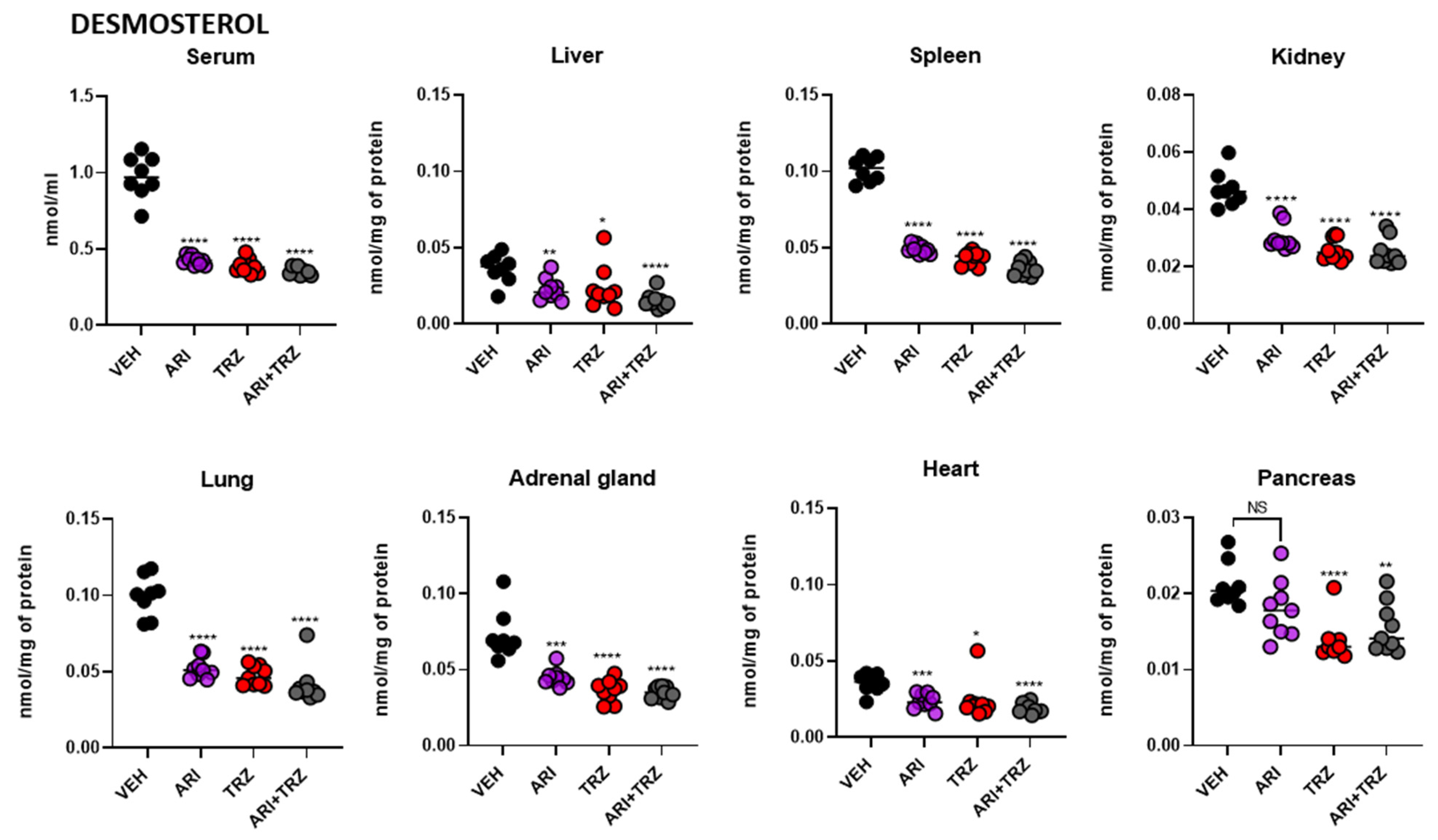
3.4. ARI and TRZ Increased 7-DHC in a Synergistic Fashion in Serum, Liver, and Spleen and Decreased DES in All Sample Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Correll, C.U.; Frederickson, A.M.; Kane, J.M.; Manu, P. Does antipsychotic polypharmacy increase the risk for metabolic syndrome? Schizophr. Res. 2007, 89, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Kukreja, S.; Kalra, G.; Shah, N.; Shrivastava, A. Polypharmacy in psychiatry: A review. Mens Sana Monogr. 2013, 11, 82–99. [Google Scholar]
- Carmona-Huerta, J.; Castiello-de Obeso, S.; Ramírez-Palomino, J.; Duran-Gutiérrez, R.; Cardona-Muller, D.; Grover-Paez, F.; Fernández-Dorantes, P.; Medina-Dávalos, R. Polypharmacy in a hospitalized psychiatric population: Risk estimation and damage quantification. BMC Psychiatry 2019, 19, 78. [Google Scholar] [CrossRef] [Green Version]
- Buckley, N.A.; Sanders, P. Cardiovascular adverse effects of antipsychotic drugs. Drug Saf. 2000, 23, 215–228. [Google Scholar] [CrossRef]
- Todorović Vukotić, N.; Đorđević, J.; Pejić, S.; Đorđević, N.; Pajović, S.B. Antidepressants- and antipsychotics-induced hepatotoxicity. Arch. Toxicol. 2021, 95, 767–789. [Google Scholar] [CrossRef]
- Castanheira, L.; Fernandes, E.; Levy, P.; Coentre, R. Aripiprazole-induced hepatitis: A case report. Clin. Psychopharmacol. Neurosci. Off. Sci. J. Korean Coll. Neuropsychopharmacol. 2019, 17, 551–555. [Google Scholar] [CrossRef]
- Livertox: Clinical and Research Information on Drug-Induced Liver Injury. Available online: https://www.ncbi.nlm.nih.gov/books/NBK548557/ (accessed on 5 August 2022).
- Fernandes, N.F.; Martin, R.R.; Schenker, S. Trazodone-induced hepatotoxicity: A case report with comments on drug-induced hepatotoxicity. Am. J. Gastroenterol. 2000, 95, 532–535. [Google Scholar] [CrossRef]
- Huhn, M.; Nikolakopoulou, A.; Schneider-Thoma, J.; Krause, M.; Samara, M.; Peter, N.; Arndt, T.; Bäckers, L.; Rothe, P.; Cipriani, A.; et al. Comparative efficacy and tolerability of 32 oral antipsychotics for the acute treatment of adults with multi-episode schizophrenia: A systematic review and network meta-analysis. Lancet 2019, 394, 939–951. [Google Scholar] [CrossRef] [Green Version]
- Citrome, L. A review of aripiprazole in the treatment of patients with schizophrenia or bipolar i disorder. Neuropsychiatr. Dis. Treat. 2006, 2, 427–443. [Google Scholar] [CrossRef] [Green Version]
- Citrome, L.; Kalsekar, I.; Baker, R.A.; Hebden, T. A review of real-world data on the effects of aripiprazole on weight and metabolic outcomes in adults. Curr. Med. Res. Opin. 2014, 30, 1629–1641. [Google Scholar] [CrossRef]
- Fagiolini, A.; Comandini, A.; Catena Dell’Osso, M.; Kasper, S. Rediscovering trazodone for the treatment of major depressive disorder. CNS Drugs 2012, 26, 1033–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.J.; Saadabadi, A. Trazodone. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470560/ (accessed on 5 August 2022).
- Berardis, D.; Fornaro, M.; Ventriglio, A.; Valchera, A.; Vellante, F.; Pettorruso, M.; Martinotti, G.; Fraticelli, S.; Giannantonio, M.D. Trazodone add-on in COVID-19-related selective serotonin reuptake inhibitor-resistant post-traumatic stress disorder in healthcare workers: Two case reports. Clin. Psychopharmacol. Neurosci. Off. Sci. J. Korean Coll. Neuropsychopharmacol. 2021, 19, 780–785. [Google Scholar] [CrossRef] [PubMed]
- Gettu, N.; Saadabadi, A. Aripiprazole. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK547739/ (accessed on 5 August 2022).
- Korade, Z.; Kim, H.Y.; Tallman, K.A.; Liu, W.; Koczok, K.; Balogh, I.; Xu, L.; Mirnics, K.; Porter, N.A. The effect of small molecules on sterol homeostasis: Measuring 7-dehydrocholesterol in dhcr7-deficient neuro2a cells and human fibroblasts. J. Med. Chem. 2016, 59, 1102–1115. [Google Scholar] [CrossRef] [Green Version]
- Hall, P.; Michels, V.; Gavrilov, D.; Matern, D.; Oglesbee, D.; Raymond, K.; Rinaldo, P.; Tortorelli, S. Aripiprazole and trazodone cause elevations of 7-dehydrocholesterol in the absence of smith-lemli-opitz syndrome. Mol. Genet. Metab. 2013, 110, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Korade, Ž.; Liu, W.; Warren, E.B.; Armstrong, K.; Porter, N.A.; Konradi, C. Effect of psychotropic drug treatment on sterol metabolism. Schizophr. Res. 2017, 187, 74–81. [Google Scholar] [CrossRef]
- Korade, Z.; Allen, L.B.; Anderson, A.; Tallman, K.A.; Genaro-Mattos, T.C.; Porter, N.A.; Mirnics, K. Trazodone effects on developing brain. Transl. Psychiatry 2021, 11, 85. [Google Scholar] [CrossRef]
- Genaro-Mattos, T.C.; Allen, L.B.; Anderson, A.; Tallman, K.A.; Porter, N.A.; Korade, Z.; Mirnics, K. Maternal aripiprazole exposure interacts with 7-dehydrocholesterol reductase mutations and alters embryonic neurodevelopment. Mol. Psychiatry 2019, 24, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Kelley, R.I. Diagnosis of smith-lemli-opitz syndrome by gas chromatography/mass spectrometry of 7-dehydrocholesterol in plasma, amniotic fluid and cultured skin fibroblasts. Clin. Chim. Acta Int. J. Clin. Chem. 1995, 236, 45–58. [Google Scholar] [CrossRef]
- Lamberson, C.R.; Muchalski, H.; McDuffee, K.B.; Tallman, K.A.; Xu, L.; Porter, N.A. Propagation rate constants for the peroxidation of sterols on the biosynthetic pathway to cholesterol. Chem. Phys. Lipids 2017, 207, 51–58. [Google Scholar] [CrossRef]
- Xu, L.; Davis, T.A.; Porter, N.A. Rate constants for peroxidation of polyunsaturated fatty acids and sterols in solution and in liposomes. J. Am. Chem. Soc. 2009, 131, 13037–13044. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Korade, Z.; Rosado, J.D.A.; Liu, W.; Lamberson, C.R.; Porter, N.A. An oxysterol biomarker for 7-dehydrocholesterol oxidation in cell/mouse models for smith-lemli-opitz syndrome. J. Lipid Res. 2011, 52, 1222–1233. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Korade, Z.; Porter, N.A. Oxysterols from free radical chain oxidation of 7-dehydrocholesterol: Product and mechanistic studies. J. Am. Chem. Soc. 2010, 132, 2222–2232. [Google Scholar] [CrossRef] [Green Version]
- Korade, Z.; Xu, L.; Shelton, R.; Porter, N.A. Biological activities of 7-dehydrocholesterol-derived oxysterols: Implications for smith-lemli-opitz syndrome. J. Lipid Res. 2010, 51, 3259–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeffer, B.A.; Xu, L.; Porter, N.A.; Rao, S.R.; Fliesler, S.J. Differential cytotoxic effects of 7-dehydrocholesterol-derived oxysterols on cultured retina-derived cells: Dependence on sterol structure, cell type, and density. Exp. Eye Res. 2016, 145, 297–316. [Google Scholar] [CrossRef] [Green Version]
- Brown, A.J.; Coates, H.W.; Sharpe, L.J. Chapter 10 - cholesterol synthesis. In Biochemistry of Lipids, Lipoproteins and Membranes, 7th ed.; Ridgway, N.D., McLeod, R.S., Eds.; Elsevier: Sydney, NSW, Australia, 2021; pp. 317–355. [Google Scholar]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Fleischhacker, W.W. Aripiprazole. Expert Opin. Pharmacother. 2005, 6, 2091–2101. [Google Scholar] [CrossRef]
- Liu, W.; Xu, L.; Lamberson, C.; Haas, D.; Korade, Z.; Porter, N.A. A highly sensitive method for analysis of 7-dehydrocholesterol for the study of smith-lemli-opitz syndrome. J. Lipid Res. 2014, 55, 329–337. [Google Scholar] [CrossRef] [Green Version]
- Allen, L.B.; Genaro-Mattos, T.C.; Anderson, A.; Porter, N.A.; Mirnics, K.; Korade, Z. Amiodarone alters cholesterol biosynthesis through tissue-dependent inhibition of emopamil binding protein and dehydrocholesterol reductase 24. ACS Chem. Neurosci. 2020, 11, 1413–1423. [Google Scholar] [CrossRef]
- Jones, P.J.; Leitch, C.A.; Li, Z.C.; Connor, W.E. Human cholesterol synthesis measurement using deuterated water. Theoretical and procedural considerations. Arterioscler. Thromb. A J. Vasc. Biol. 1993, 13, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Nestel, P.J. Cholesterol turnover in man. In Advances in Lipid Research; Paoletti, R., Kritchevsky, D., Eds.; Elsevier: Canberra, ACT, Australia, 1970; Volume 8, pp. 1–39. [Google Scholar]
- Röschlau, P.; Bernt, E.; Gruber, W. Cholesterol and esterified cholesterol. In Methods of Enzymatic Analysis, 2nd ed.; Bergmeyer, H.U., Ed.; Academic Press: Cambridge, MA, USA, 1974; pp. 1890–1893. [Google Scholar]
- Heller, F.R. Cholesterol esterifying capacity of various organs in cholesterol-fed guinea pigs. Lipids 1983, 18, 18–24. [Google Scholar] [CrossRef]
- Mojtabai, R.; Olfson, M. National trends in psychotropic medication polypharmacy in office-based psychiatry. Arch. Gen. Psychiatry 2010, 67, 26–36. [Google Scholar] [CrossRef]
- Gallego, J.A.; Nielsen, J.; De Hert, M.; Kane, J.M.; Correll, C.U. Safety and tolerability of antipsychotic polypharmacy. Expert Opin. Drug Saf. 2012, 11, 527–542. [Google Scholar] [CrossRef] [Green Version]
- Stroup, T.S.; Gray, N. Management of common adverse effects of antipsychotic medications. World Psychiatry Off. J. World Psychiatr. Assoc. 2018, 17, 341–356. [Google Scholar] [CrossRef]
- Xu, L.; Mirnics, K.; Bowman, A.B.; Liu, W.; Da, J.; Porter, N.A.; Korade, Z. Dhceo accumulation is a critical mediator of pathophysiology in a smith-lemli-opitz syndrome model. Neurobiol. Dis. 2012, 45, 923–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korade, Z.; Xu, L.; Mirnics, K.; Porter, N.A. Lipid biomarkers of oxidative stress in a genetic mouse model of smith-lemli-opitz syndrome. J. Inherit. Metab. Dis. 2013, 36, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.A.; Chattopadhyay, A. Membrane cholesterol regulates endocytosis and trafficking of the serotonin(1a) receptor: Insights from acute cholesterol depletion. Biochimica et biophysica acta. Mol. Cell Biol. Lipids 2021, 1866, 158882. [Google Scholar] [CrossRef] [PubMed]
- Di Sciascio, G.; Riva, M.A. Aripiprazole: From pharmacological profile to clinical use. Neuropsychiatr. Dis. Treat. 2015, 11, 2635–2647. [Google Scholar]
- Mandrioli, R.; Protti, M.; Mercolini, L. New-generation, non-ssri antidepressants: Therapeutic drug monitoring and pharmacological interactions. Part 1: Snris, smss, saris. Curr. Med. Chem. 2018, 25, 772–792. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Liu, S.; Sun, Q.; Zhu, J.-X. Dopamine receptors in the gastrointestinal tract. In Dopamine in the Gut; Springer: Singapore, 2021; pp. 53–85. [Google Scholar]
- Bornstein, J.C. Serotonin in the gut: What does it do? Front. Neurosci. 2012, 6, 16. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.Y.; Korade, Z.; Tallman, K.A.; Liu, W.; Weaver, C.D.; Mirnics, K.; Porter, N.A. Inhibitors of 7-dehydrocholesterol reductase: Screening of a collection of pharmacologically active compounds in neuro2a cells. Chem. Res. Toxicol. 2016, 29, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Wages, P.A.; Kim, H.H.; Korade, Z.; Porter, N.A. Identification and characterization of prescription drugs that change levels of 7-dehydrocholesterol and desmosterol. J. Lipid Res. 2018, 59, 1916–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallman, K.A.; Allen, L.B.; Klingelsmith, K.B.; Anderson, A.; Genaro-Mattos, T.C.; Mirnics, K.; Porter, N.A.; Korade, Z. Prescription medications alter neuronal and glial cholesterol synthesis. ACS Chem. Neurosci. 2021, 12, 735–745. [Google Scholar] [CrossRef] [PubMed]
- Masnoon, N.; Shakib, S.; Kalisch-Ellett, L.; Caughey, G.E. What is polypharmacy? A systematic review of definitions. BMC Geriatr. 2017, 17, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpe, M.; Chin, D.; Paneni, F. The challenge of polypharmacy in cardiovascular medicine. Fundam. Clin. Pharmacol. 2010, 24, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Korade, Z.; Genaro-Mattos, T.C.; Tallman, K.A.; Liu, W.; Garbett, K.A.; Koczok, K.; Balogh, I.; Mirnics, K.; Porter, N.A. Vulnerability of dhcr7(+/-) mutation carriers to aripiprazole and trazodone exposure. J. Lipid Res. 2017, 58, 2139–2146. [Google Scholar] [CrossRef]
- Allen, L.B.; Mirnics, K. Metoprolol inhibits developmental brain sterol biosynthesis in mice. Biomolecules 2022, 12, 1211. [Google Scholar] [CrossRef] [PubMed]
- Genaro-Mattos, T.C.; Klingelsmith, K.B.; Allen, L.B.; Anderson, A.; Tallman, K.A.; Porter, N.A.; Korade, Z.; Mirnics, K. Sterol biosynthesis inhibition in pregnant women taking prescription medications. ACS Pharmacol. Transl. Sci. 2021, 4, 848–857. [Google Scholar] [CrossRef] [PubMed]
- Siwek, M.; Woroń, J.; Gorostowicz, A.; Wordliczek, J. Adverse effects of interactions between antipsychotics and medications used in the treatment of cardiovascular disorders. Pharmacol. Rep. PR 2020, 72, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Correll, C.U.; Solmi, M.; Veronese, N.; Bortolato, B.; Rosson, S.; Santonastaso, P.; Thapa-Chhetri, N.; Fornaro, M.; Gallicchio, D.; Collantoni, E.; et al. Prevalence, incidence and mortality from cardiovascular disease in patients with pooled and specific severe mental illness: A large-scale meta-analysis of 3,211,768 patients and 113,383,368 controls. World Psychiatry Off. J. World Psychiatr. Assoc. 2017, 16, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Kilbourne, A.M.; Brar, J.S.; Drayer, R.A.; Xu, X.; Post, E.P. Cardiovascular disease and metabolic risk factors in male patients with schizophrenia, schizoaffective disorder, and bipolar disorder. Psychosomatics 2007, 48, 412–417. [Google Scholar] [CrossRef]
- Goldstein, B.I.; Schaffer, A.; Wang, S.; Blanco, C. Excessive and premature new-onset cardiovascular disease among adults with bipolar disorder in the us nesarc cohort. J. Clin. Psychiatry 2015, 76, 163–169. [Google Scholar] [CrossRef]
- Goldstein, B.I.; Fagiolini, A.; Houck, P.; Kupfer, D.J. Cardiovascular disease and hypertension among adults with bipolar i disorder in the united states. Bipolar Disord. 2009, 11, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, S.; Blanca, B.; Davies, S.; Savović, J.; Richards, A.; Sullivan, S.; Moran, P. Antipsychotic polypharmacy and metabolic syndrome in schizophrenia: A review of systematic reviews. FOCUS 2020, 18, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Misawa, F.; Shimizu, K.; Fujii, Y.; Miyata, R.; Koshiishi, F.; Kobayashi, M.; Shida, H.; Oguchi, Y.; Okumura, Y.; Ito, H.; et al. Is antipsychotic polypharmacy associated with metabolic syndrome even after adjustment for lifestyle effects?: A cross-sectional study. BMC Psychiatry 2011, 11, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunakaran, U.; Park, K.G. A systematic review of oxidative stress and safety of antioxidants in diabetes: Focus on islets and their defense. Diabetes Metab. J. 2013, 37, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez-Vicente, A.; Hong, N.; Garvin, J.L. Effects of reactive oxygen species on renal tubular transport. Am. J. Physiol. Ren. Physiol. 2019, 317, F444–F455. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Liu, J.; Hou, F.; Liu, Z.; Cao, X.; Seo, H.; Gao, B. Cholesterol induces pancreatic β cell apoptosis through oxidative stress pathway. Cell Stress Chaperones 2011, 16, 539–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zager, R.A.; Johnson, A.C.; Hanson, S.Y.; Shah, V.O. Acute tubular injury causes dysregulation of cellular cholesterol transport proteins. Am. J. Pathol. 2003, 163, 313–320. [Google Scholar] [CrossRef] [Green Version]
- Rajamäki, K.; Lappalainen, J.; Öörni, K.; Välimäki, E.; Matikainen, S.; Kovanen, P.T.; Eklund, K.K. Cholesterol crystals activate the nlrp3 inflammasome in human macrophages: A novel link between cholesterol metabolism and inflammation. PLoS ONE 2010, 5, e11765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parhofer, K.G. Interaction between glucose and lipid metabolism: More than diabetic dyslipidemia. Diabetes Metab. J. 2015, 39, 353–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balog, M.; Anderson, A.; Genaro-Mattos, T.C.; Korade, Z.; Mirnics, K. Individual and simultaneous treatment with antipsychotic aripiprazole and antidepressant trazodone inhibit sterol biosynthesis in the adult brain. J. Lipid Res. 2022, 63, 10249. [Google Scholar] [CrossRef] [PubMed]
- Cenik, B.; Palka, J.M.; Thompson, B.M.; McDonald, J.G.; Tamminga, C.A.; Cenik, C.; Brown, E.S. Desmosterol and 7-dehydrocholesterol concentrations in post mortem brains of depressed people: The role of trazodone. Transl. Psychiatry 2022, 12, 139. [Google Scholar] [CrossRef] [PubMed]
- Porter, F.D. Smith–lemli–opitz syndrome: Pathogenesis, diagnosis and management. Eur. J. Hum. Genet. 2008, 16, 535–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sever, N.; Mann, R.K.; Xu, L.; Snell, W.J.; Hernandez-Lara, C.I.; Porter, N.A.; Beachy, P.A. Endogenous b-ring oxysterols inhibit the hedgehog component smoothened in a manner distinct from cyclopamine or side-chain oxysterols. Proc. Natl. Acad. Sci. USA 2016, 113, 5904–5909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boland, M.R.; Tatonetti, N.P. Investigation of 7-dehydrocholesterol reductase pathway to elucidate off-target prenatal effects of pharmaceuticals: A systematic review. Pharm. J. 2016, 16, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.S.; Goldstein, J.L. Familial hypercholesterolemia: Defective binding of lipoproteins to cultured fibroblasts associated with impaired regulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase activity. Proc. Natl. Acad. Sci. USA 1974, 71, 788–792. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.S.; Dana, S.E.; Goldstein, J.L. Regulation of 3-hydroxy-3-methylglutaryl coenzyme a reductase activity in human fibroblasts by lipoproteins. Proc. Natl. Acad. Sci. USA 1973, 70, 2162–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radhakrishnan, A.; Ikeda, Y.; Kwon, H.J.; Brown, M.S.; Goldstein, J.L. Sterol-regulated transport of srebps from endoplasmic reticulum to golgi: Oxysterols block transport by binding to insig. Proc. Natl. Acad. Sci. USA 2007, 104, 6511–6518. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.-P.; Seemann, J.; Goldstein, J.L.; Brown, M.S. Sterol-regulated transport of srebps from endoplasmic reticulum to golgi: Insig renders sorting signal in scap inaccessible to copii proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 6519–6526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korade, Z.; Heffer, M.; Mirnics, K. Medication effects on developmental sterol biosynthesis. Mol. Psychiatry 2022, 27, 490–501. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Treatment | 7-DHC ± SEM | DES ± SEM |
|---|---|---|---|
| Serum (nmol/mL) | VEH | 0.136 ± 0.011 | 0.975 ± 0.050 |
| ARI | 1.754 ± 0.263 | 0.425 ± 0.010 | |
| TRZ | 1.194 ± 0.107 | 0.388 ± 0.016 | |
| ARI + TRZ | 4.616 ± 0.189 | 0.353 ± 0.008 | |
| Liver (nmol/mg) | VEH | 0.012 ± 0.001 | 0.036 ± 0.003 |
| ARI | 0.115 ± 0.015 | 0.023 ± 0.002 | |
| TRZ | 0.077 ± 0.006 | 0.024 ± 0.005 | |
| ARI + TRZ | 0.311 ± 0.012 | 0.015 ± 0.002 | |
| Spleen (nmol/mg) | VEH | 0.007 ± 0.0004 | 0.101 ± 0.003 |
| ARI | 0.120 ± 0.013 | 0.049 ± 0.001 | |
| TRZ | 0.076 ± 0.004 | 0.043 ± 0.001 | |
| ARI + TRZ | 0.285 ± 0.015 | 0.036 ± 0.002 | |
| Kidney (nmol/mg) | VEH | 0.053 ± 0.003 | 0.047 ± 0.002 |
| ARI | 0.252 ± 0.020 | 0.030 ± 0.002 | |
| TRZ | 0.219 ± 0.013 | 0.026 ± 0.001 | |
| ARI + TRZ | 0.357 ± 0.011 | 0.025 ± 0.002 | |
| Lung (nmol/mg) | VEH | 0.019 ± 0.001 | 0.100 ± 0.005 |
| ARI | 0.183 ± 0.014 | 0.052 ± 0.002 | |
| TRZ | 0.184 ± 0.008 | 0.047 ± 0.002 | |
| ARI + TRZ | 0.376 ± 0.015 | 0.041 ± 0.004 | |
| Adrenal gl. (nmol/mg) | VEH | 0.010 ± 0.003 | 0.074 ± 0.006 |
| ARI | 0.049 ± 0.004 | 0.045 ± 0.002 | |
| TRZ | 0.059 ± 0.004 | 0.036 ± 0.002 | |
| ARI + TRZ | 0.107 ± 0.004 | 0.035 ± 0.001 | |
| Heart (nmol/mg) | VEH | 0.009 ± 0.0003 | 0.036 ± 0.001 |
| ARI | 0.032 ± 0.003 | 0.024 ± 0.002 | |
| TRZ | 0.045 ± 0.008 | 0.024 ± 0.004 | |
| ARI + TRZ | 0.061 ± 0.009 | 0.019 ± 0.001 | |
| Pancreas (nmol/mg) | VEH | 0.003 ± 0.0001 | 0.021 ± 0.001 |
| ARI | 0.022 ± 0.002 | 0.018 ± 0.001 | |
| TRZ | 0.022 ± 0.003 | 0.014 ± 0.001 | |
| ARI + TRZ | 0.048 ± 0.003 | 0.016 ± 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balog, M.; Anderson, A.C.; Heffer, M.; Korade, Z.; Mirnics, K. Effects of Psychotropic Medication on Somatic Sterol Biosynthesis of Adult Mice. Biomolecules 2022, 12, 1535. https://doi.org/10.3390/biom12101535
Balog M, Anderson AC, Heffer M, Korade Z, Mirnics K. Effects of Psychotropic Medication on Somatic Sterol Biosynthesis of Adult Mice. Biomolecules. 2022; 12(10):1535. https://doi.org/10.3390/biom12101535
Chicago/Turabian StyleBalog, Marta, Allison C Anderson, Marija Heffer, Zeljka Korade, and Karoly Mirnics. 2022. "Effects of Psychotropic Medication on Somatic Sterol Biosynthesis of Adult Mice" Biomolecules 12, no. 10: 1535. https://doi.org/10.3390/biom12101535