

Zebrafish Model of Severe Combined Immunodeficiency (SCID) Due to JAK3 Mutation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Genetic Manipulation and Analysis
2.3. Whole-Mount In Situ Hybridization (WISH)
2.4. RT-PCR and qRT2-PCR Analysis
2.5. Ex Vivo Analyses
2.6. Survival Analysis
3. Results
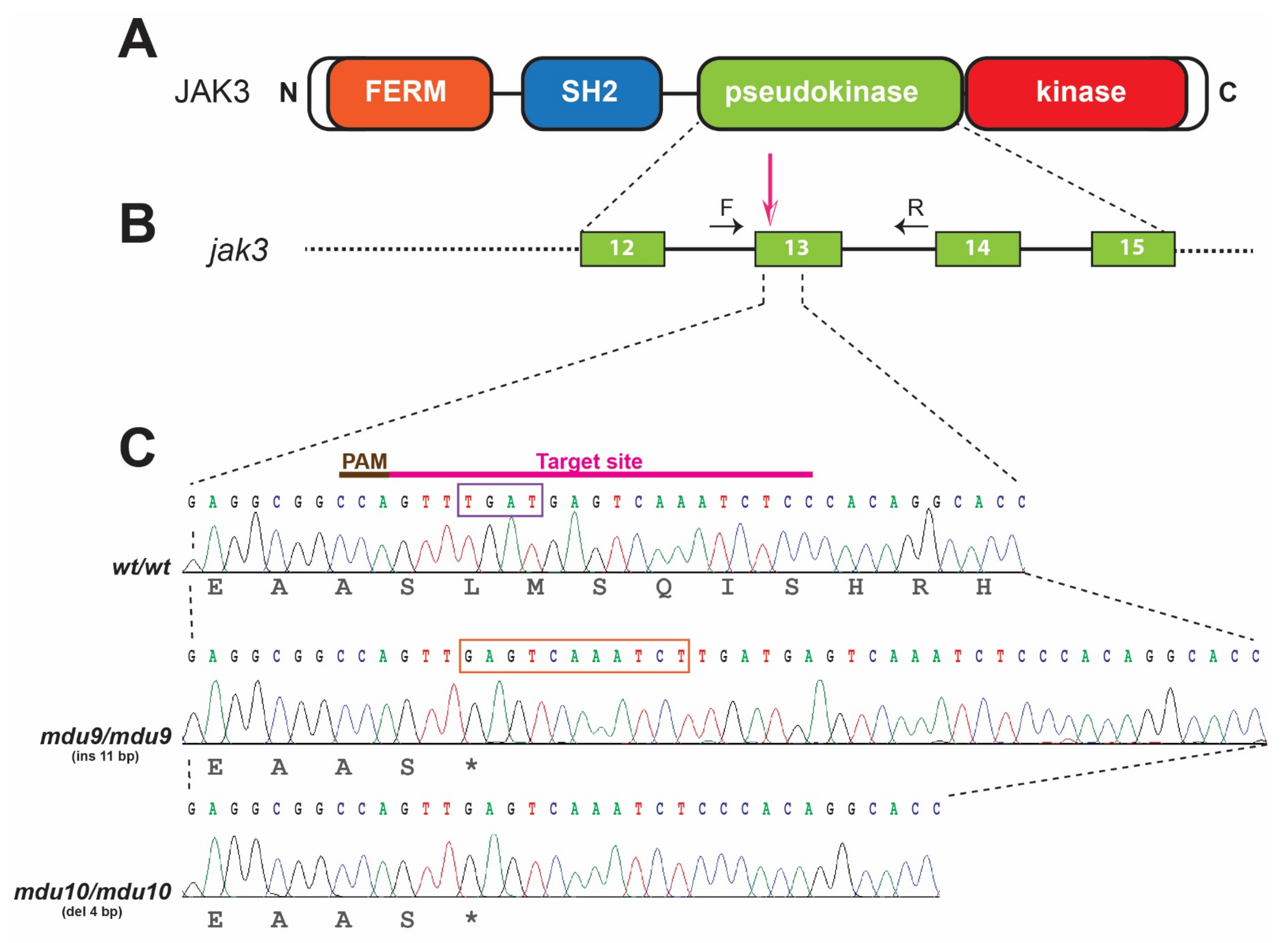
3.1. Generation of SCID-Related Jak3 Mutant Zebrafish
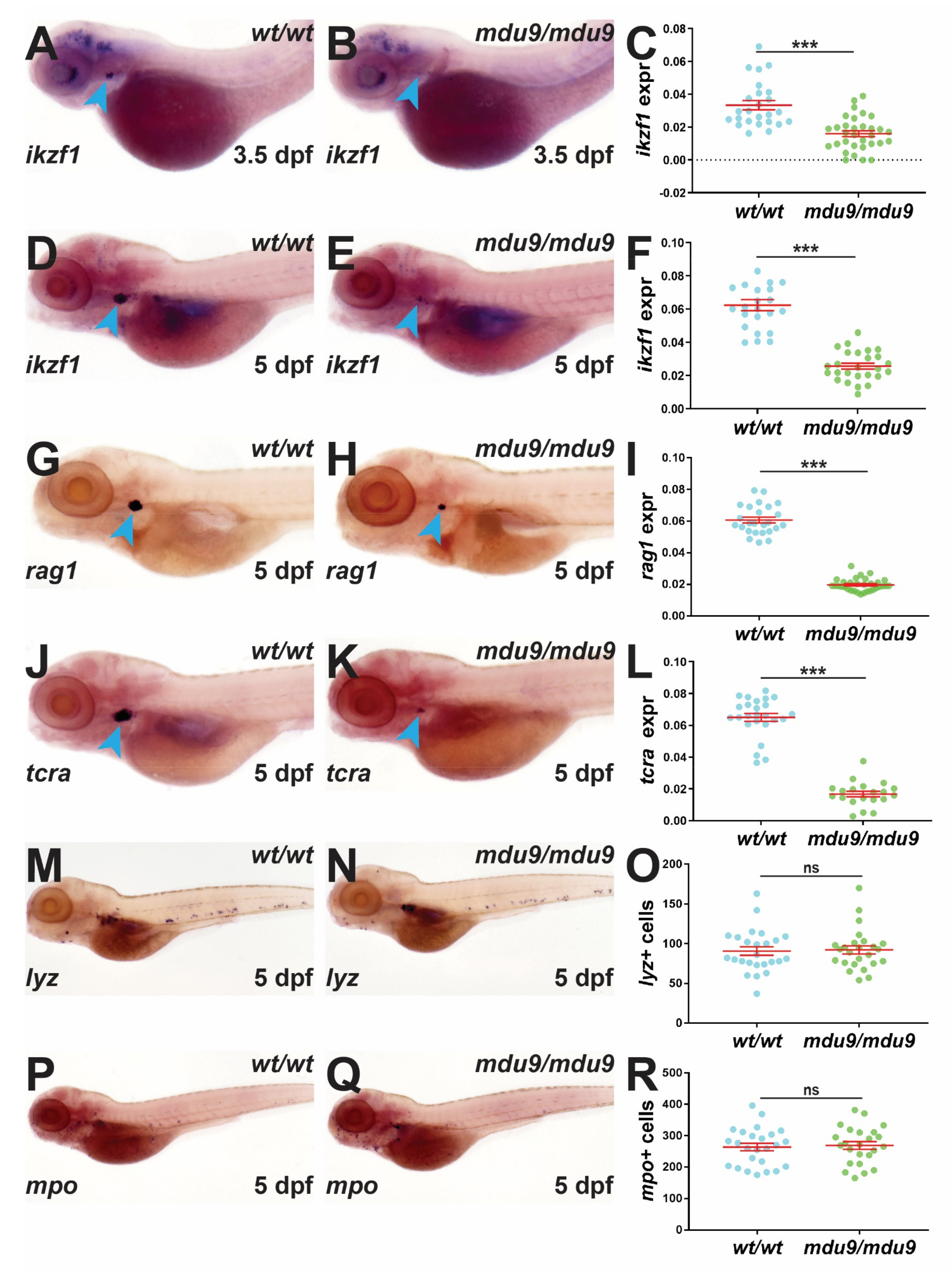
3.2. SCID-Related Jak3 Mutation Disrupts Embryonic Lymphopoiesis
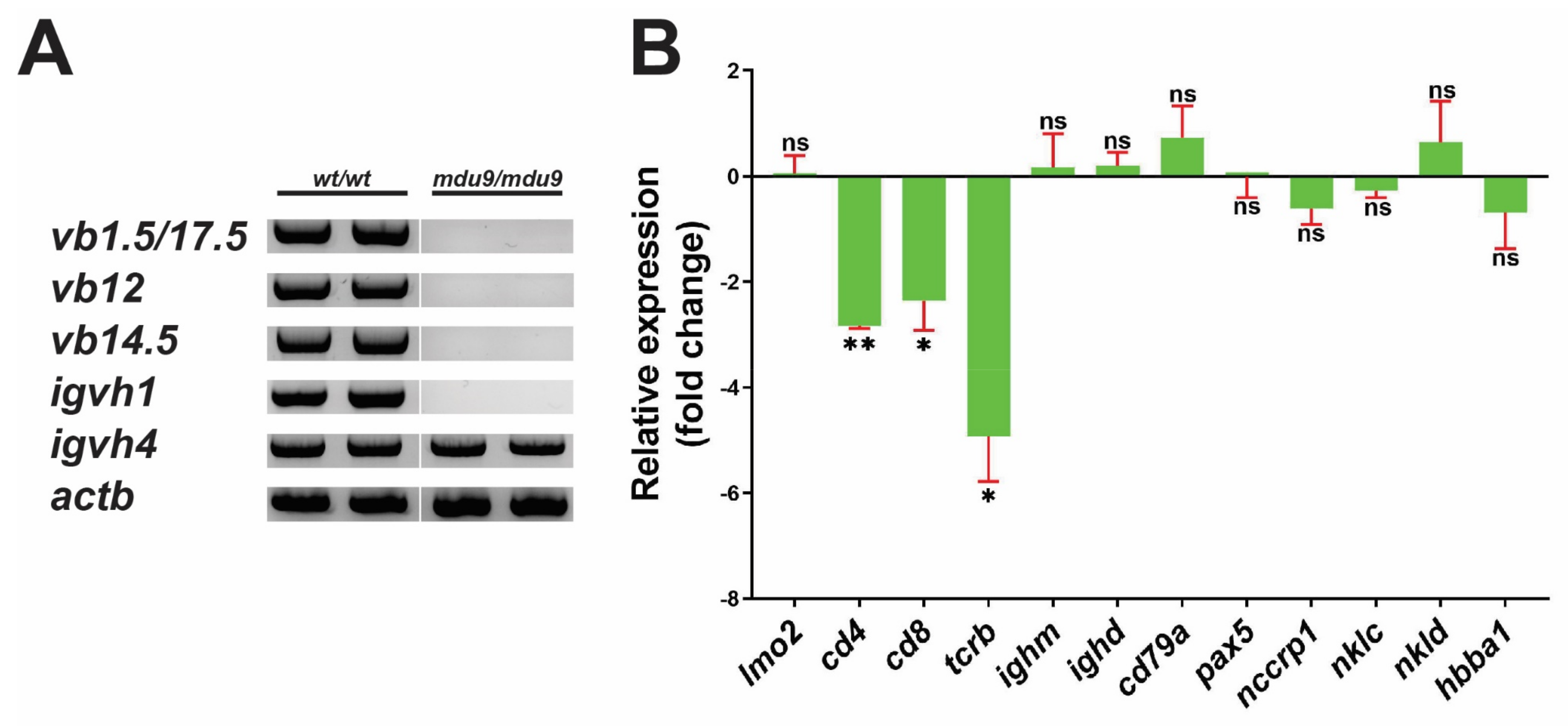
3.3. SCID-Related Jak3 Mutation Perturbs Larval Lymphopoiesis
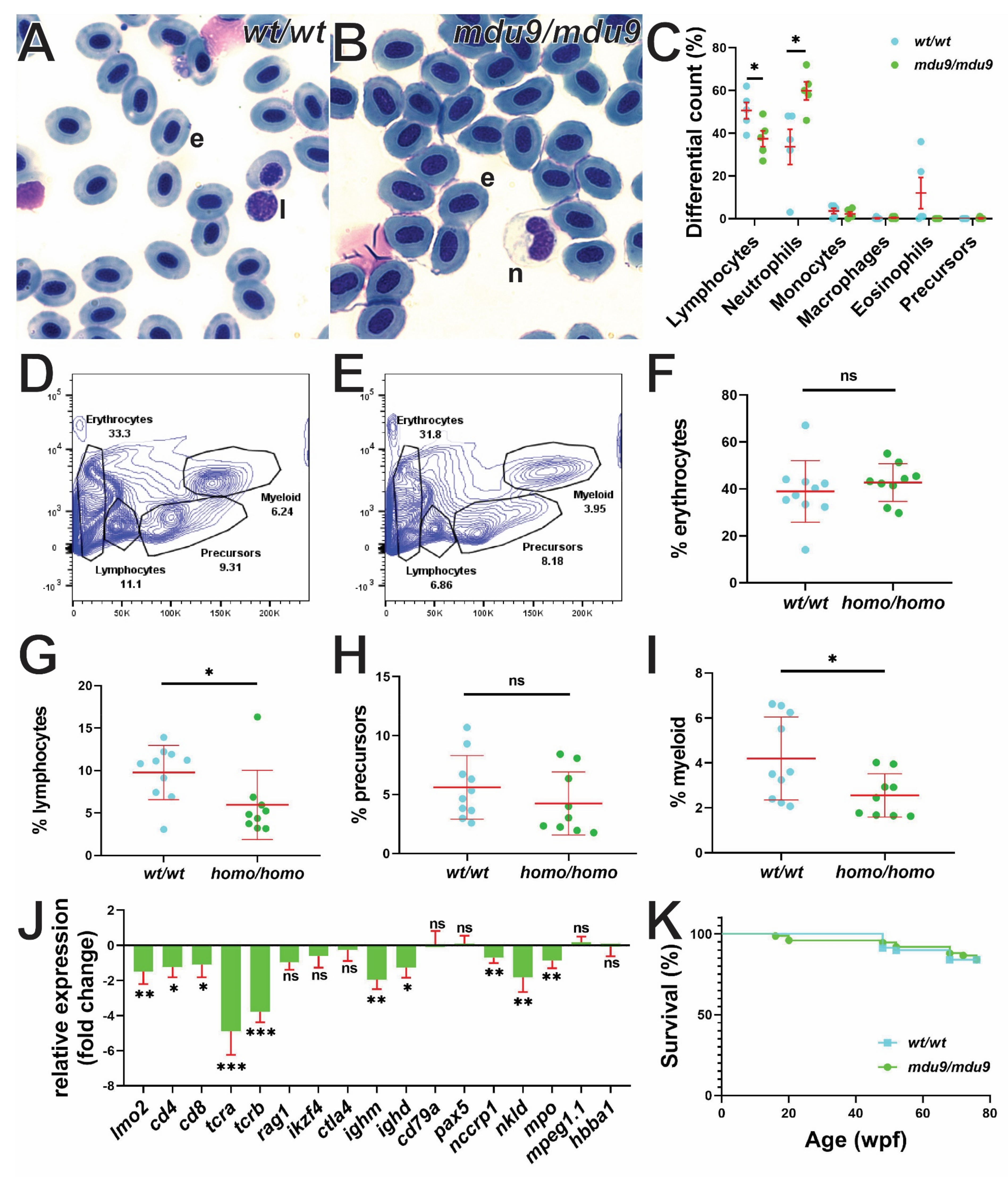
3.4. SCID-Related Jak3 Mutation Perturbs Adult Lymphopoiesis
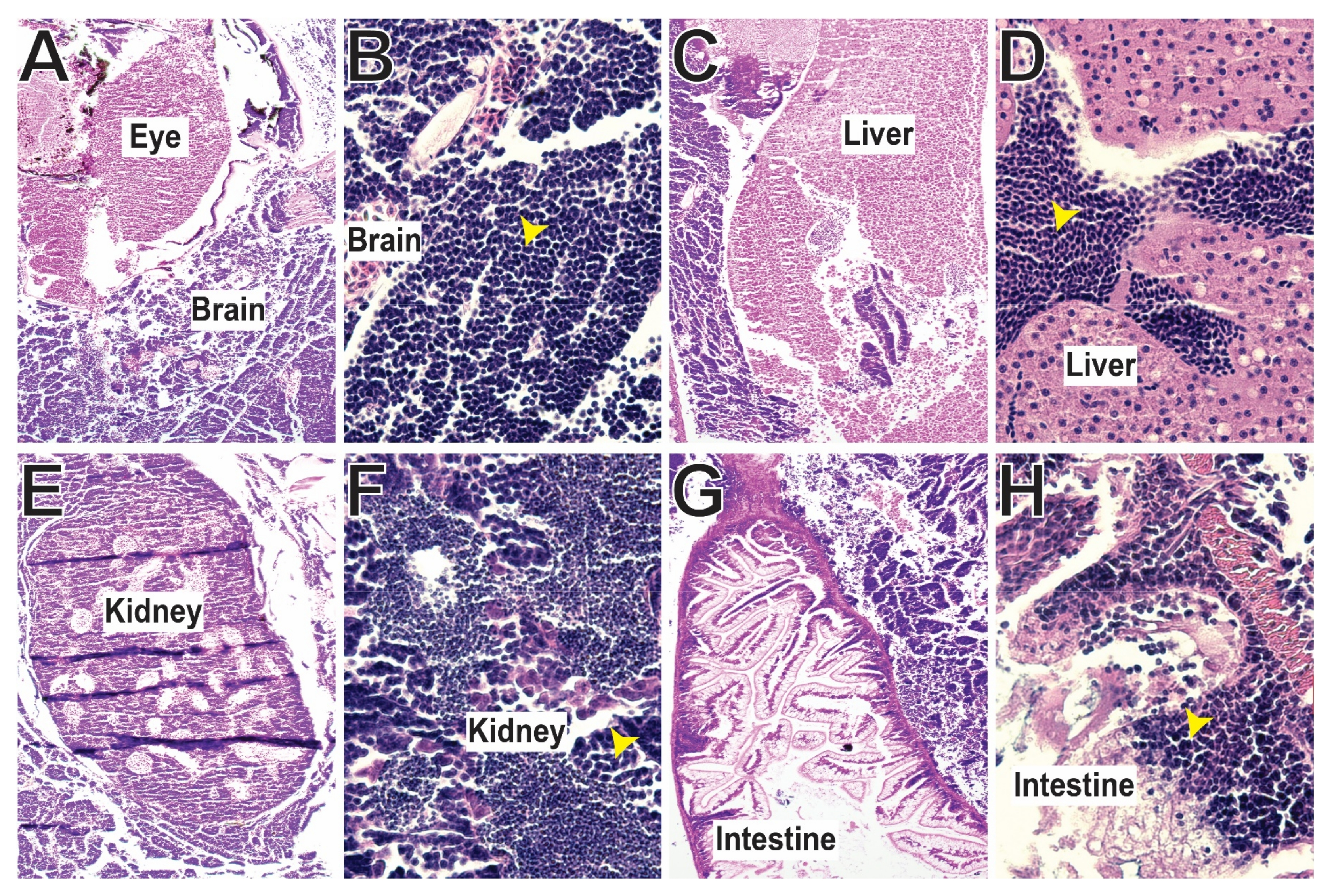
3.5. SCID-Related Jak3 Mutants Are Susceptible to Lymphocytic Leukemia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From clinical significance to quantification. Adv. Sci. 2021, 8, 2004433. [Google Scholar] [CrossRef] [PubMed]
- Gulati, K.; Guhathakurta, S.; Joshi, J.; Rai, N.; Ray, A. Cytokines and their role in health and disease: A brief overview. Mol. Immunol. 2016, 4, 121. [Google Scholar]
- Liongue, C.; O’Sullivan, L.A.; Trengove, M.C.; Ward, A.C. Evolution of JAK-STAT pathway components: Mechanisms and role in immune system development. PLoS ONE 2012, 7, e32777. [Google Scholar]
- Yamaoka, K.; Saharinen, P.; Pesu, M.; Holt, V.E.; Silvennoinen, O.; O’Shea, J.J. The Janus kinases (Jaks). Genome Biol. 2004, 5, 253. [Google Scholar] [CrossRef] [Green Version]
- Ghoreschi, K.; Laurence, A.; O’Shea, J.J. Janus kinases in immune cell signaling. Immunol. Rev. 2009, 228, 273–287. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.A.; Wang, L.-M.; Hanson, E.P.; Sun, X.-J.; White, M.F.; Oakes, S.A.; Pierce, J.H.; O’Shea, J.J. Interleukins 2, 4, 7, and 15 stimulate tyrosine phosphorylation of insulin receptor substrates 1 and 2 in T cells potential role of Jak kinases. J. Biol. Chem. 1995, 270, 28527–28530. [Google Scholar] [CrossRef] [Green Version]
- Johnston, J.A.; Kawamura, M.; Kirken, R.A.; Chen, Y.-Q.; Blake, T.B.; Shibuya, K.; Ortaldo, J.R.; McVicar, D.W.; O’Shea, J.J. Phosphorylation and activation of the Jak-3 Janus kinase in response to interleukin-2. Nature 1994, 370, 151–153. [Google Scholar] [CrossRef] [Green Version]
- Nelson, B.H.; McIntosh, B.C.; Rosencrans, L.L.; Greenberg, P.D. Requirement for an initial signal from the membrane-proximal region of the interleukin 2 receptor γc chain for Janus kinase activation leading to T cell proliferation. Proc. Natl. Acad. Sci. USA 1997, 94, 1878–1883. [Google Scholar] [CrossRef] [Green Version]
- Russell, S.M.; Johnston, J.A.; Noguchi, M.; Kawamura, M.; Bacon, C.M.; Friedmann, M.; Berg, M.; McVicar, D.W.; Witthuhn, B.A.; Silvennoinen, O. Interaction of IL-2Rβ and γc chains with Jak1 and Jak3: Implications for XSCID and XCID. Science 1994, 266, 1042–1045. [Google Scholar] [CrossRef] [Green Version]
- Bains, T.; Heinrich, M.; Loriaux, M.; Beadling, C.; Nelson, D.; Warrick, A.; Neff, T.; Tyner, J.; Dunlap, J.; Corless, C. Newly described activating JAK3 mutations in T-cell acute lymphoblastic leukemia. Leukemia 2012, 26, 2144–2146. [Google Scholar] [CrossRef]
- Kiyoi, H.; Yamaji, S.; Kojima, S.; Naoe, T. JAK3 mutations occur in acute megakaryoblastic leukemia both in Down syndrome children and non-Down syndrome adults. Leukemia 2007, 21, 574–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, G.C.; Tan, S.Y.; Tang, T.; Poon, S.L.; Allen, G.E.; Tan, L.; Chong, S.C.; Ong, W.S.; Tay, K.; Tao, M. Janus kinase 3–activating mutations identified in Natural killer/T-cell lymphoma. Cancer Discov. 2012, 2, 591–597. [Google Scholar] [CrossRef] [PubMed]
- Notarangelo, L.D.; Mella, P.; Jones, A.; de Saint Basile, G.; Savoldi, G.; Cranston, T.; Vihinen, M.; Schumacher, R.F. Mutations in severe combined immune deficiency (SCID) due to JAK3 deficiency. Hum. Mutat. 2001, 18, 255–263. [Google Scholar] [CrossRef]
- Macchi, P.; Villa, A.; Giliani, S.; Sacco, M.G.; Frattini, A.; Porta, F.; Ugazio, A.G.; Johnston, J.A.; Candotti, F.; O’Sheai, J.J. Mutations of Jak-3 gene in patients with autosomal severe combined immune deficiency (SCID). Nature 1995, 377, 65–68. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.M.; Tayebi, N.; Nakajima, H.; Riedy, M.C.; Roberts, J.L.; Aman, M.J.; Migone, T.-S.; Noguchi, M.; Markert, M.L.; Buckley, R.H. Mutation of Jak3 in a patient with SCID: Essential role of Jak3 in lymphoid development. Science 1995, 270, 797–800. [Google Scholar] [CrossRef] [Green Version]
- Rasighaemi, P.; Basheer, F.; Liongue, C.; Ward, A.C. Zebrafish as a model for leukemia and other hematopoietic disorders. J. Hematol. Oncol. 2015, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Liongue, C.; Ward, A.C. Evolution of class I cytokine receptors. BMC Evol. Biol. 2007, 7, 120. [Google Scholar] [CrossRef] [Green Version]
- Sertori, R.; Jones, R.; Basheer, F.; Rivera, L.; Dawson, S.; Loke, S.; Heidary, S.; Dhillon, A.; Liongue, C.; Ward, A.C. Generation and characterization of a zebrafish IL-2Rγc SCID model. Int. J. Mol. Sci. 2022, 23, 2385. [Google Scholar] [CrossRef]
- Moore, J.C.; Tang, Q.; Yordán, N.T.; Moore, F.E.; Garcia, E.G.; Lobbardi, R.; Ramakrishnan, A.; Marvin, D.L.; Anselmo, A.; Sadreyev, R.I. Single-cell imaging of normal and malignant cell engraftment into optically clear prkdc-null SCID zebrafish. J. Exp. Med. 2016, 213, 2575–2589. [Google Scholar] [CrossRef] [Green Version]
- Iwanami, N.; Mateos, F.; Hess, I.; Riffel, N.; Soza-Ried, C.; Schorpp, M.; Boehm, T. Genetic evidence for an evolutionarily conserved role of IL-7 signaling in T cell development of zebrafish. J. Immunol. 2011, 186, 7060–7066. [Google Scholar] [CrossRef] [Green Version]
- Basheer, F.; Bulleeraz, V.; Ngo, V.Q.; Liongue, C.; Ward, A.C. In vivo impact of JAK3 A573V mutation revealed using zebrafish. Cell. Mol. Life Sci. 2022, 79, 322. [Google Scholar] [CrossRef]
- Langenau, D.M.; Traver, D.; Ferrando, A.A.; Kutok, J.L.; Aster, J.C.; Kanki, J.P.; Lin, S.; Prochownik, E.; Trede, N.S.; Zon, L.I. Myc-induced T cell leukemia in transgenic zebrafish. Science 2003, 299, 887–890. [Google Scholar] [CrossRef]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Garritano, S.; Gemignani, F.; Voegele, C.; Nguyen-Dumont, T.; Calvez-Kelm, L.; De Silva, D.; Lesueur, F.; Landi, S.; Tavtigian, S.V. Determining the effectiveness of High Resolution Melting analysis for SNP genotyping and mutation scanning at the TP53 locus. BMC Genet. 2009, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.L.; Lengi, A.; Brown, S.M.; Chen, M.; Zhou, Y.-J.; O’Shea, J.J.; Buckley, R.H. Janus kinase 3 (JAK3) deficiency: Clinical, immunologic, and molecular analyses of 10 patients and outcomes of stem cell transplantation. Blood 2004, 103, 2009–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willett, C.E.; Kawasaki, H.; Amemiya, C.T.; Lin, S.; Steiner, L.A. Ikaros expression as a marker for lymphoid progenitors during zebrafish development. Dev. Dyn. 2001, 222, 694–698. [Google Scholar] [CrossRef]
- Danilova, N.; Hohman, V.S.; Sacher, F.; Ota, T.; Willett, C.E.; Steiner, L.A. T cells and the thymus in developing zebrafish. Dev. Comp. Immunol. 2004, 28, 755–767. [Google Scholar] [CrossRef]
- Langenau, D.M.; Ferrando, A.A.; Traver, D.; Kutok, J.L.; Hezel, J.-P.D.; Kanki, J.P.; Zon, L.I.; Look, A.T.; Trede, N.S. In vivo tracking of T cell development, ablation, and engraftment in transgenic zebrafish. Proc. Natl. Acad. Sci. USA 2004, 101, 7369–7374. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Wen, Z. Cloning and expression pattern of the lysozyme C gene in zebrafish. Mech. Dev. 2002, 113, 69–72. [Google Scholar] [CrossRef]
- Lieschke, G.J.; Oates, A.C.; Crowhurst, M.O.; Ward, A.C.; Layton, J.E. Morphologic and functional characterization of granulocytes and macrophages in embryonic and adult zebrafish. Blood 2001, 98, 3087–3096. [Google Scholar] [CrossRef] [PubMed]
- Page, D.M.; Wittamer, V.; Bertrand, J.Y.; Lewis, K.L.; Pratt, D.N.; Delgado, N.; Schale, S.E.; McGue, C.; Jacobsen, B.H.; Doty, A. An evolutionarily conserved program of B-cell development and activation in zebrafish. Blood 2013, 122, e1–e11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrie-Hanson, L.; Hohn, C.; Hanson, L. Characterization of rag 1 mutant zebrafish leukocytes. BMC Immunol. 2009, 10, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorpp, M.; Bialecki, M.; Diekhoff, D.; Walderich, B.; Odenthal, J.; Maischein, H.-M.; Zapata, A.G.; Boehm, T. Conserved functions of Ikaros in vertebrate lymphocyte development: Genetic evidence for distinct larval and adult phases of T cell development and two lineages of B cells in zebrafish. J. Immunol. 2006, 177, 2463–2476. [Google Scholar] [CrossRef] [Green Version]
- Moore, F.E.; Garcia, E.G.; Lobbardi, R.; Jain, E.; Tang, Q.; Moore, J.C.; Cortes, M.; Molodtsov, A.; Kasheta, M.; Luo, C.C. Single-cell transcriptional analysis of normal, aberrant, and malignant hematopoiesis in zebrafish. J. Exp. Med. 2016, 213, 979–992. [Google Scholar] [CrossRef] [Green Version]
- Chan, F.-Y.; Robinson, J.; Brownlie, A.; Shivdasani, R.A.; Donovan, A.; Brugnara, C.; Kim, J.; Lau, B.-C.; Witkowska, H.E.; Zon, L.I. Characterization of adult α-and β-globin genes in the zebrafish. Blood 1997, 89, 688–700. [Google Scholar] [CrossRef]
- Zakrzewska, A.; Cui, C.; Stockhammer, O.W.; Benard, E.L.; Spaink, H.P.; Meijer, A.H. Macrophage-specific gene functions in Spi1-directed innate immunity. Blood 2010, 116, e1–e11. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Lin, J.-X.; Leonard, W.J. Interleukin-2 at the crossroads of effector responses, tolerance, and immunotherapy. Immunity 2013, 38, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Malek, T.R. The biology of interleukin-2. Annu. Rev. Immunol. 2008, 26, 453–479. [Google Scholar] [CrossRef]
- Meazza, R.; Azzarone, B.; Orengo, A.M.; Ferrini, S. Role of common-gamma chain cytokines in NK cell development and function: Perspectives for immunotherapy. J. Biomed. Biotechnol. 2011, 2011, 861920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochman, Y.; Spolski, R.; Leonard, W.J. New insights into the regulation of T cells by γc family cytokines. Nat. Rev. Immunol. 2009, 9, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Cornejo, M.G.; Boggon, T.J.; Mercher, T. JAK3: A two-faced player in hematological disorders. Int. J. Biochem. Cell Biol. 2009, 41, 2376–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nosaka, T.; Van Deursen, J.M.; Tripp, R.A.; Thierfelder, W.E.; Witthuhn, B.A.; McMickle, A.P.; Doherty, P.C.; Grosveld, G.C.; Ihle, J.N. Defective lymphoid development in mice lacking Jak3. Science 1995, 270, 800–802. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Saijo, K.; Takahashi, T.; Osawa, M.; Areas, H.; Hirayama, N.; Miyake, K.; Nakauchi, H.; Shirasawa, T.; Saito, T. Developmental defects of lymphoid cells in Jak3 kinase-deficient mice. Immunity 1995, 3, 771–782. [Google Scholar] [CrossRef] [Green Version]
- Thomis, D.C.; Berg, L.J. The role of Jak3 in lymphoid development, activation, and signaling. Curr. Opin. Immunol. 1997, 9, 541–547. [Google Scholar] [CrossRef]
- Gurniak, C.B.; Berg, L.J. Murine JAK3 is preferentially expressed in hematopoietic tissues and lymphocyte precursor cells. Blood 1996, 87, 3151–3160. [Google Scholar] [CrossRef] [Green Version]
- Lai, K.S.; Jin, Y.; Graham, D.K.; Witthuhn, B.A.; Ihle, J.N.; Liu, E.T. A kinase-deficient splice variant of the human JAK3 is expressed in hematopoietic and epithelial cancer cells. J. Biol. Chem. 1995, 270, 25028–25036. [Google Scholar] [CrossRef] [Green Version]
- Cacalano, N.A.; Migone, T.-S.; Bazan, F.; Hanson, E.P.; Chen, M.; Candotti, F.; O’Shea, J.J.; Johnston, J.A. Autosomal SCID caused by a point mutation in the N-terminus of Jak3: Mapping of the Jak3–receptor interaction domain. EMBO J. 1999, 18, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Candotti, F.; Oakes, S.A.; Johnston, J.A.; Giliani, S.; Schumacher, R.F.; Mella, P.; Fiorini, M.; Ugazio, A.G.; Badolato, R.; Notarangelo, L.D. Structural and functional basis for JAK3-deficient severe combined immunodeficiency. Blood 1997, 90, 3996–4003. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, M.; Yi, H.; Rosenblatt, H.M.; Filipovich, A.H.; Adelstein, S.; Modi, W.S.; McBride, O.W.; Leonard, W.J. Interleukin-2 receptor γ chain mutation results in X-linked severe combined immunodeficiency in humans. Cell 1993, 73, 147–157. [Google Scholar] [CrossRef]
- Thomis, D.C.; Gurniak, C.B.; Tivol, E.; Sharpe, A.H.; Berg, L.J. Defects in B lymphocyte maturation and T lymphocyte activation in mice lacking Jak3. Science 1995, 270, 794–797. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, P.; Xin, Z.; Shi, C.; Bai, Y.; Sun, X.; Zhao, Y.; Wang, X.; Liu, L.; Zhao, X. Biological characteristics of severe combined immunodeficient mice produced by CRISPR/Cas9-mediated Rag2 and IL2rg mutation. Front. Genet. 2019, 10, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byambaa, S.; Uosaki, H.; Hara, H.; Nagao, Y.; Abe, T.; Shibata, H.; Nureki, O.; Ohmori, T.; Hanazono, Y. Generation of novel Il2rg-knockout mice with clustered regularly interspaced short palindromic repeats (CRISPR) and Cas9. Exp. Anim. 2020, 69, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sertori, R.; Liongue, C.; Basheer, F.; Lewis, K.L.; Rasighaemi, P.; de Coninck, D.; Traver, D.; Ward, A.C. Conserved IL-2Rγc signaling mediates lymphopoiesis in zebrafish. J. Immunol. 2016, 196, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puel, A.; Ziegler, S.F.; Buckley, R.H. Defective IL7R expression in T-B+ NK+ severe combined immunodeficiency. Nat. Genet. 1998, 20, 394–397. [Google Scholar] [CrossRef] [PubMed]
- Roifman, C.M.; Zhang, J.; Chitayat, D.; Sharfe, N. A partial deficiency of interleukin-7Rα is sufficient to abrogate T-cell development and cause severe combined immunodeficiency. Blood 2000, 96, 2803–2807. [Google Scholar] [CrossRef] [PubMed]
- Peschon, J.J.; Morrissey, P.J.; Grabstein, K.H.; Ramsdell, F.J.; Maraskovsky, E.; Gliniak, B.C.; Park, L.S.; Ziegler, S.F.; Williams, D.E.; Ware, C.B. Early lymphocyte expansion is severely impaired in interleukin 7 receptor-deficient mice. J. Exp. Med. 1994, 180, 1955–1960. [Google Scholar] [CrossRef] [Green Version]
- von Freeden-Jeffry, U.; Vieira, P.; Lucian, L.A.; McNeil, T.; Burdach, S.; Murray, R. Lymphopenia in interleukin (IL)-7 gene-deleted mice identifies IL-7 as a nonredundant cytokine. J. Exp. Med. 1995, 181, 1519–1526. [Google Scholar] [CrossRef] [Green Version]
- He, Y.-W.; Malek, T.R. Interleukin-7 receptor alpha is essential for the development of gamma delta+ T cells, but not Natural killer cells. J. Exp. Med. 1996, 184, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Grossman, W.J.; Verbsky, J.W.; Yang, L.; Berg, L.J.; Fields, L.E.; Chaplin, D.D.; Ratner, L. Dysregulated myelopoiesis in mice lacking Jak3. Blood 1999, 94, 932–939. [Google Scholar] [CrossRef] [PubMed]
- Rane, S.G.; Mangan, J.K.; Amanullah, A.; Wong, B.C.; Vora, R.K.; Liebermann, D.A.; Hoffman, B.; Grana, X.; Reddy, E.P. Activation of the Jak3 pathway is associated with granulocytic differentiation of myeloid precursor cells. Blood 2002, 100, 2753–2762. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, J.J.; Husa, M.; Li, D.; Hofmann, S.R.; Watford, W.; Roberts, J.L.; Buckley, R.H.; Changelian, P.; Candotti, F. Jak3 and the pathogenesis of severe combined immunodeficiency. Mol. Immunol. 2004, 41, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.H.; Chua, H.L.; Gong, Z.; Lam, T.J.; Sin, Y.M. Development and maturation of the immune system in zebrafish, Danio rerio: A gene expression profiling, in situ hybridization and immunological study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef]
- Sunyer, J.O. Fishing for mammalian paradigms in the teleost immune system. Nat. Immunol. 2013, 14, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Mayor, P.C.; Eng, K.H.; Singel, K.L.; Abrams, S.I.; Odunsi, K.; Moysich, K.B.; Fuleihan, R.; Garabedian, E.; Lugar, P.; Ochs, H.D. Cancer in primary immunodeficiency diseases: Cancer incidence in the United States Immune Deficiency Network Registry. J. Allergy Clin. Immunol. 2018, 141, 1028–1035. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basheer, F.; Lee, E.; Liongue, C.; Ward, A.C. Zebrafish Model of Severe Combined Immunodeficiency (SCID) Due to JAK3 Mutation. Biomolecules 2022, 12, 1521. https://doi.org/10.3390/biom12101521
Basheer F, Lee E, Liongue C, Ward AC. Zebrafish Model of Severe Combined Immunodeficiency (SCID) Due to JAK3 Mutation. Biomolecules. 2022; 12(10):1521. https://doi.org/10.3390/biom12101521
Chicago/Turabian StyleBasheer, Faiza, Effie Lee, Clifford Liongue, and Alister C. Ward. 2022. "Zebrafish Model of Severe Combined Immunodeficiency (SCID) Due to JAK3 Mutation" Biomolecules 12, no. 10: 1521. https://doi.org/10.3390/biom12101521