
Human Melanocortin-2 Receptor: Identifying a Role for Residues in the TM4, EC2, and TM5 Domains in Activation and Trafficking as a Result of Co-Expression with the Accessory Protein, Mrap1 in Chinese Hamster Ovary Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
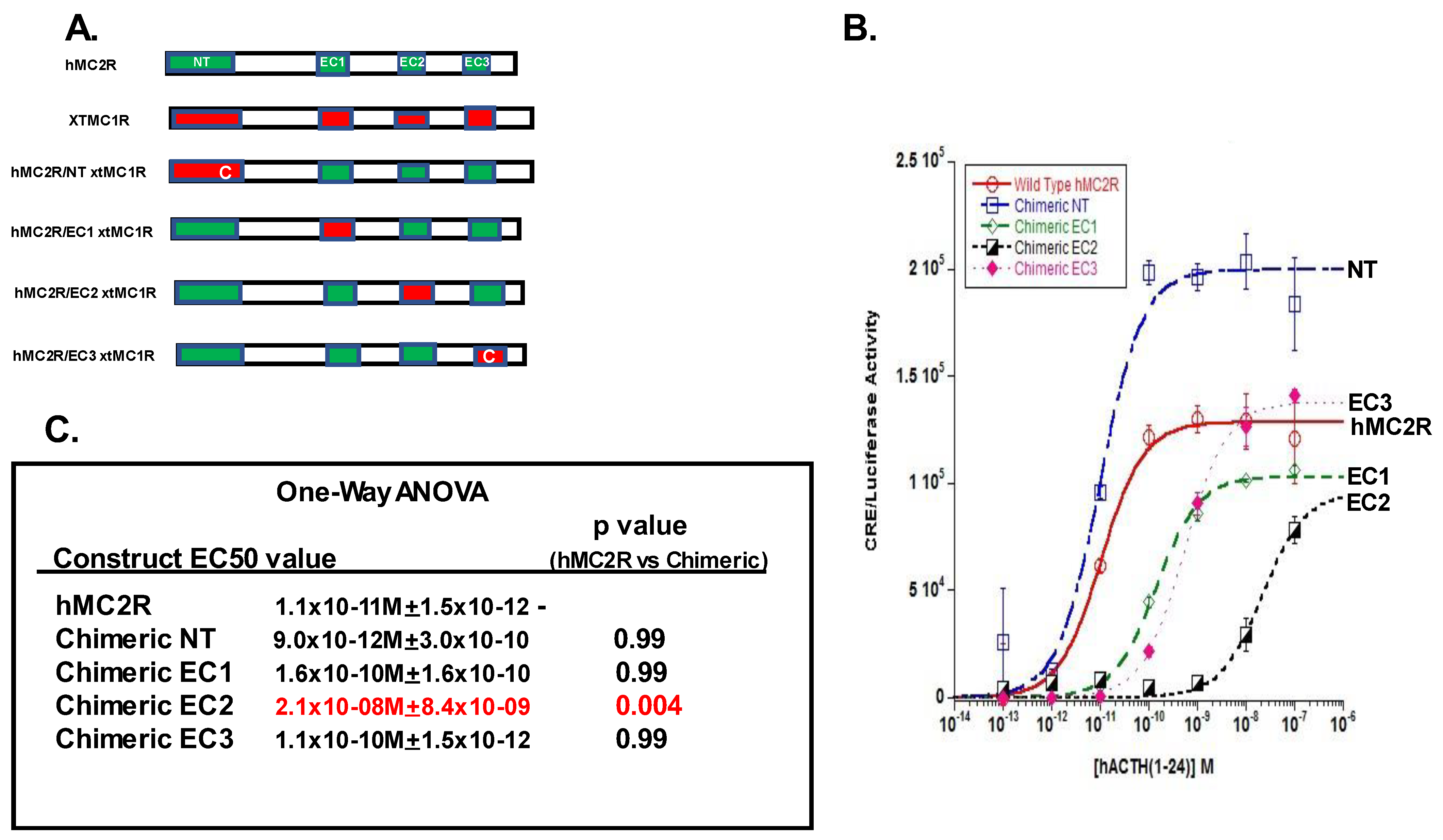
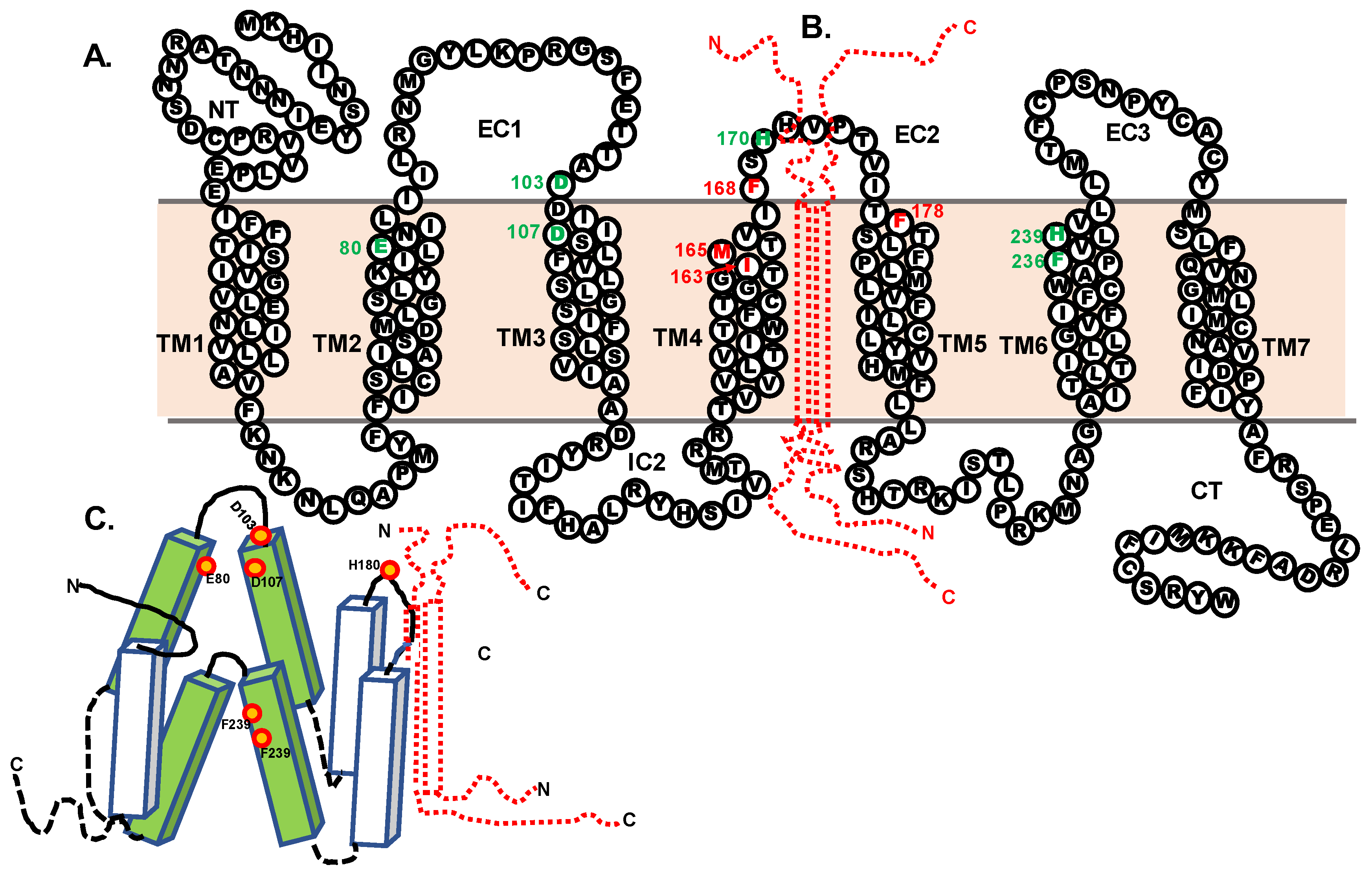
2.1. Chimer Receptor Paradigm
2.2. DNA Constructs
2.3. ACTH and α-MSH Peptides
2.4. Tissue Culture Procedure
2.5. cAMP Reporter Gene Assay (CRE-Luciferase Assay)
2.6. Cell Surface ELISA
3. Results
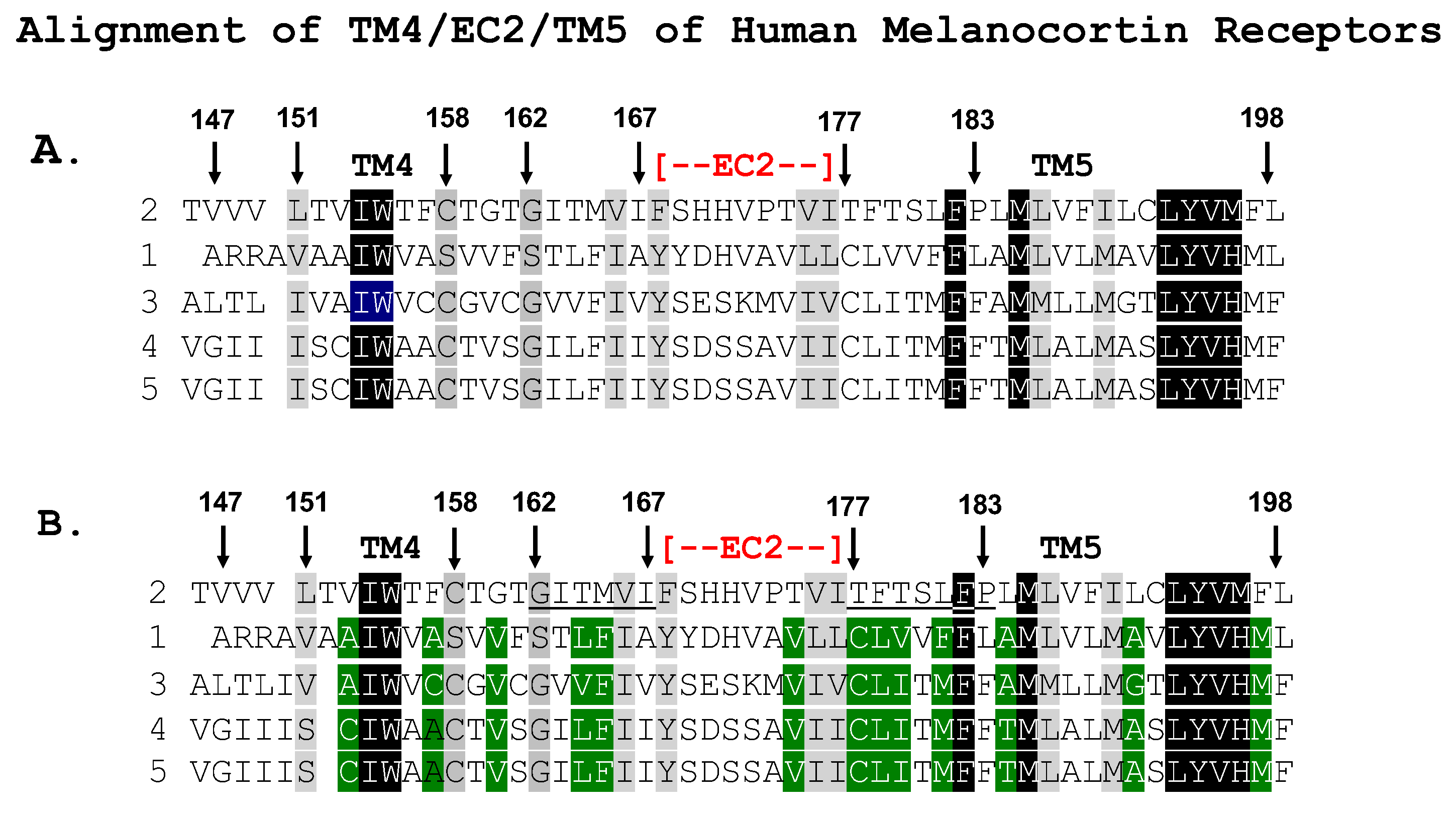
3.1. Chimeric Receptor Analysis
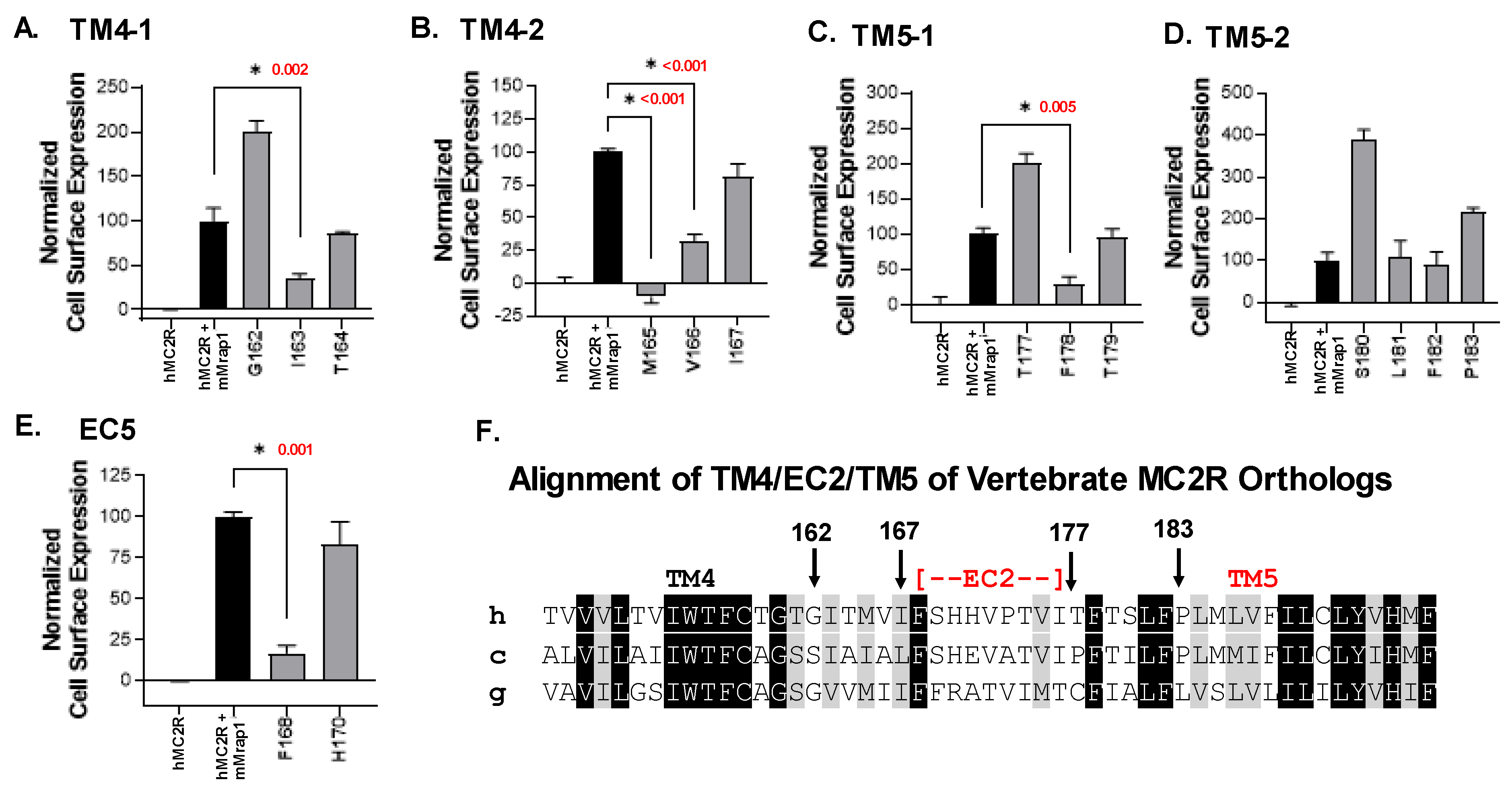
3.2. Single-Alanine Analysis of Residues in TM4, EC2, and TM5: Effects on Trafficking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cone, R.D. Studies on the physiological studies of the melanocortin system. Endocr. Rev. 2006, 27, 736–749. [Google Scholar] [CrossRef] [Green Version]
- Dores, R.M.; Liang, L.; Davis, P.; Thomas, A.L.; Petko, B. Melanocortin Receptors: Evolution of Ligand Selectivity for Melanocortin Peptides. J. Mol. Endocrinol. 2016, 56, T1–T15. [Google Scholar]
- Mountjoy, K.G.; Bird, I.M.; Rainey, W.E.; Cone, R.D. ACTH induces up-regulation of ACTH receptor mRNA in mouse and human adrenocortical cell lines. Mol. Cell. Endocrinol. 1994, 99, 17–20. [Google Scholar] [CrossRef]
- Metherell, L.A.; Chapple, J.P.; Cooray, S.; David, A.; Becker, C.; Ruschendorf, F.; Naville, D.; Begeot, M.; Khoo, B.; Nurnberg, P.; et al. Mutations in MRAP, encoding a new interacting partner of the ACTH receptor, cause familial glucocorticoid deficiency type 2. Nat. Genet. 2005, 37, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Sebag, J.A.; Hinkle, P.M. Melanocortin-2 receptor accessory protein MRAP forms antiparallel homodimers. Proc. Natl. Acad. Sci. USA 2007, 2104, 20244–20249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinkle, P.M.; Sebag, J.A. Structure and function of the melanocortin2 receptor accessory protein (MRAP). Mol. Cell. Endocrinol. 2009, 300, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webb, R.R.; Clark, A.J.L. Minireview: The melanocortin 2 receptor accessory proteins. Mol. Endocrinol. 2010, 24, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Sebag, J.A.; Hinkle, P.M. Regions of melanocortin 2 (MC2) receptor accessory protein necessary for dual topology and MC2 receptor trafficking and signaling. J. Biol. Chem. 2009, 284, 610–618. [Google Scholar] [CrossRef] [Green Version]
- Chan, L.F.; Webb, T.R.; Chung, T.-T.; Meimaridou, E.; Cooray, S.N.; Guasti, L.; Chapple, J.P.; Egertova, M.; Elphick, M.R.; Cheetham, M.E.; et al. MRAP and MRAP2 are bidirectional regulators of the melanocortin receptor family. Proc. Natl. Acad. Sci. USA 2009, 106, 6146–6151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, L.; Sebag, J.A.; Eagelston, L.; Serasinghe, M.N.; Veo, K.; Reinick, C.; Angleson, J.; Hinkle, P.M.; Dores, R.M. Functional Expression of frog and rainbow trout melanocortin 2 receptors using heterologous Mrap1s. Gen. Comp. Endocrinol. 2011, 174, 5–14. [Google Scholar] [CrossRef]
- Cooray, S.N.; Almiro Do Vale, I.; Leung, K.Y.; Webb, T.R.; Chapple, J.P.; Egertova, M.; Cheetham, M.E.; Elphick, M.R.; Clark, A.J. The melanocortin 2 receptor accessory protein exists as a homodimer and is essential for the function of the melanocortin 2 receptor in the mouse y1 cell line. Endocrinol. 2008, 149, 1935–1941. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.; Dolan, T.M.; Maben, Z.J.; Hinkle, P.M. Adrenocorticotropic hormone (ACTH) responses require actions of the melanocortin-2 receptor accessory protein on the extracellular surface of the plasma membrane. J. Biol. Chem. 2015, 290, 27972–27985. [Google Scholar] [CrossRef] [Green Version]
- Chepurny, O.G.; Holz, G.G. A novel cyclic adenosine monophosphateresponsiveluciferase reporter incorporating a nonpalindromic cyclic adenosine monophosphate response element provides optimal performance for use in G protein coupled receptor drug discovery efforts. J. Biomol. Screen. 2007, 12, 740–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noon, L.A.; Franklin, J.M.; King, P.J.; Goulding, N.J.; Hunyady, L.; Clark, A.J. Failed export of the adrenocorticotropin receptor from the endoplasmic reticulum in non-adrenal cells: Evidence in support of a requirement for a specific adrenal accessory factor. J. Endocrinol. 2002, 174, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Reinick, C.L.; Liang, L.; Angleson, J.K.; Dores, R.M. Identification of an Mrap-independent melanocortin-2 receptor: Functional expression of the cartilaginous fish, Callorhinchus milii, melanocortin-2 receptor in CHO cells. Endocrinol. 2012, 153, 4757–4765. [Google Scholar] [CrossRef] [Green Version]
- Barney, E.; Dores, M.R.; McAvoy, D.; Davis, P.; Racareanu, R.-C.; Iki, A.; Hyodo, S.; Dores, R.M. Elephant shark melanocortin receptors: Novel interactions with MRAP1 and implication for the HPI axis. Gen. Comp. Endocrinol. 2019, 272, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Dores, R.M.; Londraville, R.L.; Prokop, J.; Davis, P.; Dewey, N.; Lesinski, L. The evolution of melanocortin receptors: The relationship between pharmacology and physiology. J. Mol. Endocrinol. 52 2014, T29–T42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Chen, M.; Allen, R.; Kesterson, J.; Harmon, C. Structural insights into the role of the ACTH receptor cysteine residues on receptor function. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R1120–R1126. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Aprahamian, C.J.; Kesterson, R.A.; Harmon, C.M.; Yang, Y. Molecular identification of the human melanocortin-2 receptor responsible for ligand binding and signaling. Biochemistry 2007, 46, 11389–11397. [Google Scholar] [CrossRef] [Green Version]
- Chung, T.T.; Webb, T.R.; Chan, L.F.; Cooray, S.N.; Metherell, L.A.; King, P.J.; Chapple, J.P.; Clark, A.J.L. The majority of ACTH receptor (MC2R) mutations found in familial glucocorticoid deficiency type 1 lead to defective trafficking of the receptor to the cell surface. J. Clinical Endocrinol. Metabol. 2008, 93, 4948–4954. [Google Scholar] [CrossRef] [PubMed]
- Pogozheva, I.D.; Chai, B.X.; Lomize, A.L.; Fong, T.M.; Weinberg, D.H.; Nargund, R.P.; Mulholland, M.W.; Grants, I.; Mosberg, H.I. Interactions of human melanocortin 4 receptor with nonpeptide and peptide agonists. Biochemistry 2005, 44, 11329–11341. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Gimenez, L.E.; Hernandez, C.C.; Wu, Y.; Wein, A.H.; Han, G.W.; McClary, K.; Mittal, S.R.; Burdsall, K.; Stauch, B.; et al. Determination of the melanocortin-4 receptor structure identifies Ca2+ as a cofactor for ligand binding. Science 2020, 368, 428–433. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.; Veo, K.; Angleson, J.; Dores, R.M. Modeling the Evolution of the MC2R and MC5R genes: Studies on the Cartilaginous Fish, Heterondotus francisci. Gen. Comp. Endocrinol. 2009, 161, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Dores, R.M. ACTH, the MSHs and the Melanocortin Receptors: Revisiting the work of Robert Schwyzer—A 30 Year Retrospective. Ann. N. Y. Acad. Sci. 2009, 1163, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Schwyzer, R. ACTH: A short introductory review. Ann. N. Y. Acad. Sci. 1977, 297, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Fridmanis, D.; Petrovska, R.; Pjanova, D.; Schioth, H.B.; Klovins, J. Replacement of short segments within transmembrane domains of MC2R disrupts retention signal. J. Mol. Endocrinol. 2014, 53, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Buckley, D.I.; Ramachandran, J. Characterization of corticotropin receptors on adrenocortical cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7431–7435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dores, R.M.; Scuba-Gray, M.; McNally, B.; Davis, P.; Takahashi, A. Evaluating the interactions between red stingray (Dasyatis akajei) melanocortin receptors and elephant shark (Callorhinchus milii) Mrap1 and Mrap2 following stimulation with either stingray ACTH(1-24) or stingray Des-Acetyl-αMSH: A pharmacological study in Chinese Hamster Ovary Cells. Gen. Comp. Endocrinol. 2018, 265, 133–140. [Google Scholar]
- Hoglin, B.E.; Miner, M.; Dores, R.M. Pharmacological Properties of Whale Shark (Rhincodon typus) Melanocortin-2 Receptor and Melancortin-5 Receptor: Interaction with MRAP1 and MRAP2. Gen. Comp. Endocrinol. 2020, 293, 113463. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, P.V.; Shaughnessy, C.A.; Dores, R.M. Human Melanocortin-2 Receptor: Identifying a Role for Residues in the TM4, EC2, and TM5 Domains in Activation and Trafficking as a Result of Co-Expression with the Accessory Protein, Mrap1 in Chinese Hamster Ovary Cells. Biomolecules 2022, 12, 1422. https://doi.org/10.3390/biom12101422
Davis PV, Shaughnessy CA, Dores RM. Human Melanocortin-2 Receptor: Identifying a Role for Residues in the TM4, EC2, and TM5 Domains in Activation and Trafficking as a Result of Co-Expression with the Accessory Protein, Mrap1 in Chinese Hamster Ovary Cells. Biomolecules. 2022; 12(10):1422. https://doi.org/10.3390/biom12101422
Chicago/Turabian StyleDavis, Perry V., Ciaran A. Shaughnessy, and Robert M. Dores. 2022. "Human Melanocortin-2 Receptor: Identifying a Role for Residues in the TM4, EC2, and TM5 Domains in Activation and Trafficking as a Result of Co-Expression with the Accessory Protein, Mrap1 in Chinese Hamster Ovary Cells" Biomolecules 12, no. 10: 1422. https://doi.org/10.3390/biom12101422