
Statins Alleviate Tumor Hypoxia in Prostate Cancer Models by Decreasing Oxygen Consumption: An Opportunity for Radiosensitization?
 ,
,  ,
,  and
and 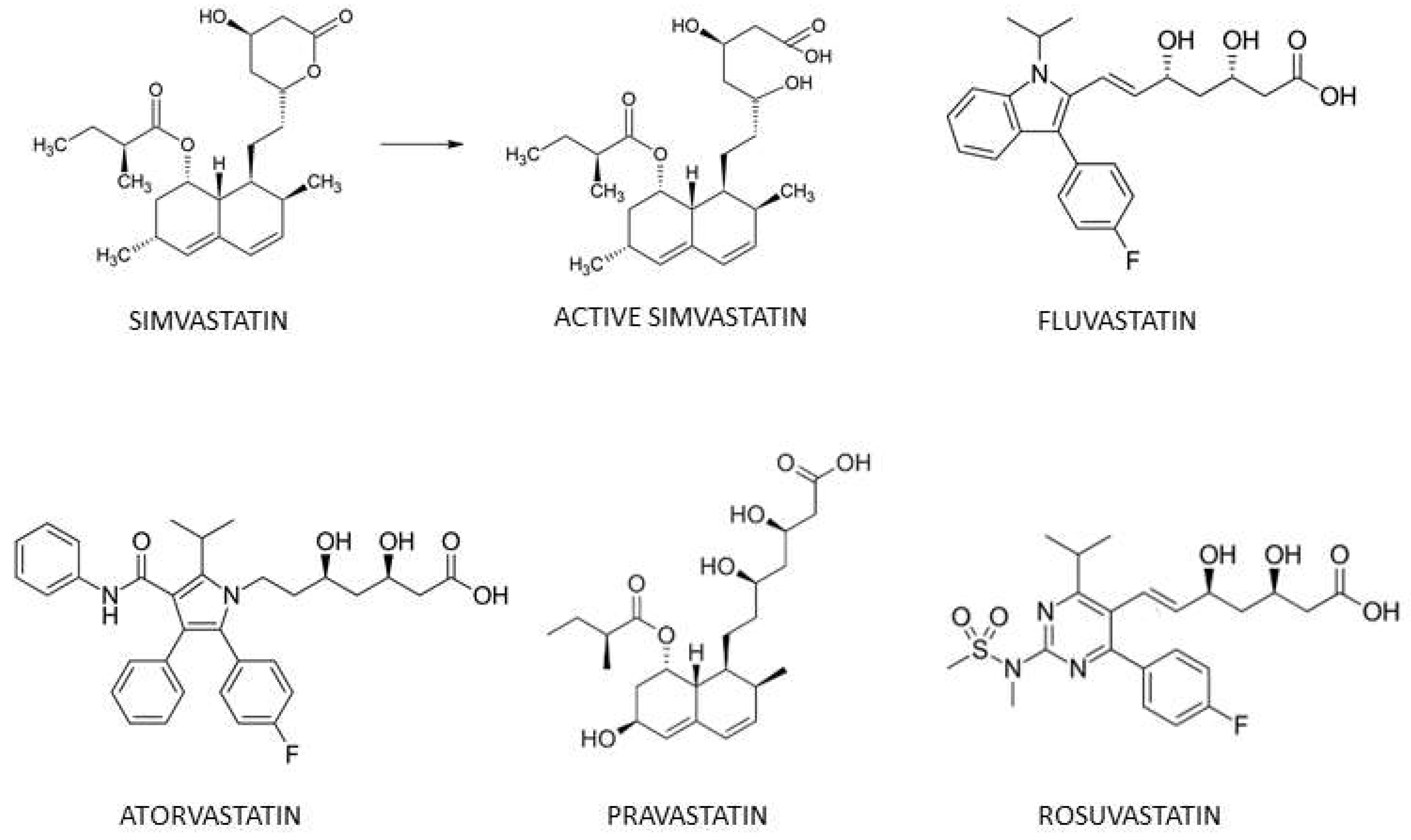{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Line and Culture
2.3. OCR Measurements by the Seahorse XF96 Technology
2.4. OCR Measurements by EPR Respirometry
2.5. Mitochondrial Superoxide Assessment by EPR Spectrometry
2.6. Tumor Models In Vivo and Treatments
2.7. In Vivo EPR Oximetry
2.8. Irradiation Dose Assessment
2.9. Tumor Growth-Delay
2.10. Statistics
3. Results
3.1. OCR in Prostate Cancer Cells Is Decreased by Simvastatin and Fluvastatin, But Is Unchanged after Pravastatin, Rosuvastatin or Atorvastatin Exposure
3.2. Modulation of Mitochondrial Superoxide Production by Simvastatin and Fluvastatin
3.3. Simvastatin and Fluvastatin Alleviate Tumor Hypoxia in Prostate Cancer Models
3.4. Does the Combination Statin/Irradiation Lead to an Improved Response to Radiotherapy?
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Bello, E.; Zwergel, C.; Mai, A.; Valente, S. The Innovative Potential of Statins in Cancer: New Targets for New Therapies. Front. Chem. 2020, 8, 516. [Google Scholar] [CrossRef] [PubMed]
- Duarte, J.A.; de Barros, A.L.B.; Leite, E.A. The potential use of simvastatin for cancer treatment: A review. Biomed. Pharmacother. 2021, 141, 111858. [Google Scholar] [CrossRef] [PubMed]
- Müller, C.; Bockhorn, A.G.; Klusmeier, S.; Kiehl, M.; Roeder, C.; Kalthoff, H.; Koch, O.M. Lovastatin inhibits proliferation of pancreatic cancer cell lines with mutant as well as with wild-type K-ras oncogene but has different effects on protein phosphorylation and induction of apoptosis. Int. J. Oncol. 1998, 12, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Katarzyna, K.; Kaminski, R.; Legat, M.; Kopec, M.; Nowis, D.; Skierski, J.S.; Koronkiewicz, M.; Jakóbisiak, M.; Golab, J. Cerivastatin demonstrates enhanced antitumor activity against human breast cancer cell lines when used in combination with doxorubicin or cisplatin. Int. J. Oncol. 2004, 24, 1149–1157. [Google Scholar]
- Khanzada, U.K.; Pardo, O.E.; Meier, C.; Downward, J.; Seckl, M.J.; Arcaro, A. Potent inhibition of small-cell lung cancer cell growth by simvastatin reveals selective functions of Ras isoforms in growth factor signalling. Oncogene 2006, 25, 877–887. [Google Scholar] [CrossRef] [Green Version]
- Afshari, A.R.; Mollazadeh, H.; Henney, N.C.; Jamialahmad, T.; Sahebkar, A. Effects of statins on brain tumors: A review. Semin. Cancer Biol. 2021, 73, 116–133. [Google Scholar] [CrossRef]
- Voorneveld, P.W.; Reimers, M.S.; Bastiaannet, E.; Jacobs, R.J.; van Eijk, R.; Zanders, M.M.; Herings, R.M.; van Herk-Sukel, M.P.; Kodach, L.; van Wezel, T.; et al. Statin Use after Diagnosis of Colon Cancer and Patient Survival. Gastroenterology 2017, 153, 470–479. [Google Scholar] [CrossRef]
- Li, X.; Liu, L.; Hu, Y. Statin use and the prognosis of patients with hepatocellular carcinoma: A meta-analysis. Biosci. Rep. 2020, 40, BSR20200232. [Google Scholar] [CrossRef] [Green Version]
- Kotti, A.; Holmqvist, A.; Albertsson, M.; Sun, X.-F. Survival benefit of statins in older patients with rectal cancer: A Swedish population-based cohort study. J. Geriatr. Oncol. 2019, 10, 690–697. [Google Scholar] [CrossRef]
- Gupta, A.; Stokes, W.; Eguchi, M.; Hararah, M.; Amini, A.; Mueller, A.; Morgan, R.; Bradley, C.; Raben, D.; McDermott, J.; et al. Statin use associated with improved overall and cancer specific survival in patients with head and neck cancer. Oral Oncol. 2019, 90, 54–66. [Google Scholar] [CrossRef]
- Xia, D.K.; Hu, Z.G.; Tian, Y.F.; Zeng, F.J. Statin use and prognosis of lung cancer: A systematic review and meta-analysis of observational studies and randomized controlled trials. Drug Des. Dev. Ther. 2019, 13, 405–422. [Google Scholar] [CrossRef]
- Cardwell, C.R.; Spence, A.D.; Hughes, C.M.; Murray, L.J. Statin use after esophageal cancer diagnosis and survival: A population based cohort study. Cancer Epidemiol. 2017, 48, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Gray, R.T.; Loughrey, M.B.; Bankhead, P.; Cardwell, C.; McQuaid, S.; O’Neill, R.F.; Arthur, K.; Bingham, V.; Mcgready, C.; Gavin, A.T.; et al. Statin use, candidate mevalonate pathway biomarkers, and colon cancer survival in a population-based cohort study. Br. J. Cancer 2017, 116, 1652–1659. [Google Scholar] [CrossRef] [Green Version]
- Seckl, M.J.; Ottensmeier, C.; Cullen, M.; Schmid, P.; Ngai, Y.; Muthukumar, D.; Thompson, J.; Harden, S.V.; Middleton, G.; Fife, K.M.; et al. Multicenter, Phase III, Randomized, Double-Blind, Placebo-Controlled Trial of Pravastatin Added to First-Line Standard Chemotherapy in Small-Cell Lung Cancer (LUNGSTAR). J. Clin. Oncol. 2017, 35, 1506–1514. [Google Scholar] [CrossRef]
- Kollmeier, M.A.; Katz, M.S.; Mak, K.; Yamada, Y.; Feder, D.J.; Zhang, Z.; Jia, X.; Shi, W.; Zelefsky, M.J. Improved biochemical outcomes with statin use in patients with high-risk localized prostate cancer treated with radiotherapy. Int. J. Radiat. Oncol. Biol. Phys. 2011, 79, 713–718. [Google Scholar] [CrossRef]
- Moyad, M.A.; Merrick, G.S.; Butler, W.M.; Wallner, K.E.; Galbreath, R.W.; Butler, E.G.; Allen, Z.A.; Adamovich, E. Statins, especially atorvastatin, may improve survival following brachytherapy for clinically localized prostate cancer. Urol. Nurs. 2006, 26, 298–303. [Google Scholar]
- Oh, D.S.; Koontz, B.; Freedland, S.J.; Gerber, L.; Patel, P.; Lewis, S.; Yoo, D.S.; Oleson, J.; Salama, J.K. Statin use is associated with decreased prostate cancer recurrence in men treated with brachytherapy. World J. Urol. 2015, 33, 93–97. [Google Scholar] [CrossRef]
- Park, H.S.; Schoenfeld, J.D.; Mailhot, R.B.; Shive, M.; Hartman, R.I.; Ogembo, R.; Mucci, L.A. Statins and prostate cancer recurrence following radical prostatectomy or radiotherapy: A systematic review and meta-analysis. Ann. Oncol. 2013, 24, 1427–1434. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, M.; Xing, D.; Feng, Y. Atorvastatin enhances radiosensitivity in hypoxia-induced prostate cancer cells related with HIF-1α inhibition. Biosci. Rep. 2017, 37, BSR20170340. [Google Scholar] [CrossRef] [Green Version]
- Dhani, N.; Fyles, A.; Hedley, D.; Milosevic, M. The clinical significance of hypoxia in human cancers. Semin. Nucl. Med. 2015, 45, 110–121. [Google Scholar] [CrossRef]
- Walsh, J.C.; Lebedev, A.; Aten, E.; Madsen, K.; Marciano, L.; Kolb, H.C. The clinical importance of assessing tumor hypoxia: Relationship of tumor hypoxia to prognosis and therapeutic opportunities. Antioxid. Redox Signal. 2014, 21, 1516–1554. [Google Scholar] [CrossRef] [PubMed]
- Colliez, F.; Gallez, B.; Jordan, B.F. Assessing Tumor Oxygenation for Predicting Outcome in Radiation Oncology: A Review of Studies Correlating Tumor Hypoxic Status and Outcome in the Preclinical and Clinical Settings. Front. Oncol. 2017, 7, 10. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Kelleher, D.K.; Hockel, M.H. Oxygen status of malignant tumors: Pathogenesis of hypoxia and significance for tumor therapy. Semin. Oncol. 2001, 28, 29–35. [Google Scholar] [CrossRef]
- Vaupel, P. Tumor microenvironmental physiology and its implications for radiation oncology. Semin. Radiat. Oncol. 2004, 14, 198–206. [Google Scholar] [CrossRef]
- Horsman, M.R.; Wouters, B.G.; Joiner, M.C.; Overgaard, J. The oxygen effect and fractionated radiotherapy. In Basic Clinical Radiobiology, 4th ed.; Joiner, M., van der Kogel, A., Eds.; Hodder Arnold: London, UK, 2009; pp. 207–216. [Google Scholar]
- Secomb, T.; Hsu, R.; Ong, E.T.; Gross, J.F.; Dewhirst, M.W. Analysis of the effects of oxygen supply and demand on hypoxic fraction in tumors. Acta Oncol. 1995, 34, 313–316. [Google Scholar] [CrossRef]
- Diepart, C.; Karroum, O.; Magat, J.; Feron, O.; Verrax, J.; Calderon, P.B.; Grégoire, V.; Leveque, P.; Stockis, J.; Dauguet, N.; et al. Arsenic trioxide treatment decreases the oxygen consumption rate of tumor cells and radiosensitizes solid tumors. Cancer Res. 2012, 72, 482–490. [Google Scholar] [CrossRef] [Green Version]
- Jordan, B.F.; Grégoire, V.; Demeure, R.J.; Sonveaux, P.; Feron, O.; O’Hara, J.; VanHulle, V.P.; Delzenne, N.; Gallez, B. Insulin increases the sensitivity of tumors to irradiation: Involvement of an increase in tumor oxygenation mediated by a nitric oxide-dependent decrease of the tumor cells oxygen consumption. Cancer Res. 2002, 62, 3555–3561. [Google Scholar]
- Crokart, N.; Jordan, B.F.; Baudelet, C.; Cron, G.O.; Hotton, J.; Radermacher, K.; Grégoire, V.; Beghein, N.; Martinive, P.; Bouzin, C.; et al. Glucocorticoids modulate tumor radiation response through a decrease in tumor oxygen consumption. Clin. Cancer Res. 2007, 13, 630–635. [Google Scholar] [CrossRef] [Green Version]
- Ashton, T.M.; Fokas, E.; Kunz-Schughart, L.A.; Folkes, L.K.; Anbalagan, S.; Huether, M.; Kelly, C.J.; Pirovano, G.; Buffa, F.M.; Hammond, E.M.; et al. The anti-malarial atovaquone increases radiosensitivity by alleviating tumour hypoxia. Nat. Commun. 2016, 7, 12308. [Google Scholar] [CrossRef] [Green Version]
- Zannella, V.E.; Dal Pra, A.; Muaddi, H.; McKee, T.D.; Stapleton, S.; Sykes, J.; Glicksman, R.; Chaib, S.; Zamiara, P.; Milosevic, M.; et al. Reprogramming metabolism with metformin improves tumor oxygenation and radiotherapy response. Clin. Cancer Res. 2013, 19, 6741–6750. [Google Scholar] [CrossRef] [Green Version]
- De Preter, G.; Deriemaeker, C.; Danhier, P.; Brisson, L.; Pham, T.T.C.; Grégoire, V.; Jordan, B.F.; Sonveaux, P.; Gallez, B. A Fast Hydrogen Sulfide-Releasing Donor Increases the Tumor Response to Radiotherapy. Mol. Cancer Ther. 2016, 15, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Gallez, B.; Neveu, M.-A.; Danhier, P.; Jordan, B.F. Manipulation of tumor oxygenation and radiosensitivity through modification of cell respiration. A critical review of approaches and imaging biomarkers for therapeutic guidance. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 700–711. [Google Scholar] [CrossRef]
- Bonifacio, A.; Mullen, P.J.; Mityko, I.S.; Navegantes, L.C.; Bouitbir, J.; Krähenbühl, S. Simvastatin induces mitochondrial dysfunction and increased atrogin-1 expression in H9c2 cardiomyocytes and mice in vivo. Arch. Toxicol. 2016, 90, 203–215. [Google Scholar] [CrossRef]
- Kwak, H.-B.; Thalacker-Mercer, A.; Anderson, E.J.; Lin, C.-T.; Kane, D.A.; Lee, N.-S.; Cortright, R.N.; Bamman, M.M.; Neufer, P.D. Simvastatin impairs ADP-stimulated respiration and increases mitochondrial oxidative stress in primary human skeletal myotubes. Free Radic. Biol. Med. 2012, 52, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Satoh, K.; Nakai, T.; Ichihara, K. Influence of 3-hydroxy-3-methylglutaryl coenzyme A reductase inhibitors on mitochondrial respiration in rat liver during ischemia. Eur. J. Pharmacol. 1994, 270, 365–369. [Google Scholar] [CrossRef]
- Thews, O.; Kelleher, D.K.; Vaupel, P. In vivo oxygen consumption rate of DS sarcoma cells on inhibition of DNA synthesis. Cancer Res. 1996, 56, 2009–2012. [Google Scholar]
- Ferrick, D.A.; Neilson, A.; Beeson, C. Advances in measuring cellular bioenergetics using extracellular flux. Drug Discov. Today 2008, 13, 268–274. [Google Scholar] [CrossRef]
- Matsuura, M.; Suzuki, T.; Suzuki, M.; Tanaka, R.; Ito, E.; Saito, T. Statin-mediated reduction of osteopontin expression induces apoptosis and cell growth arrest in ovarian clear cell carcinoma. Oncol. Rep. 2011, 25, 41–47. [Google Scholar] [CrossRef]
- Goc, A.; Kochuparambil, S.T.; Al-Husein, B.; Al-Azayzih, A.; Mohammad, S.; Somanath, P.R. Simultaneous modulation of the intrinsic and extrinsic pathways by simvastatin in mediating prostate cancer cell apoptosis. BMC Cancer 2012, 12, 409. [Google Scholar] [CrossRef] [Green Version]
- Zanfardino, M.; Spampanato, C.; De Cicco, R.; Buommino, E.; De Filippis, A.; Baiano, S.; Barra, A.; Morelli, F. Simvastatin reduces melanoma progression in a murine model. Int. J. Oncol. 2013, 43, 1763–1770. [Google Scholar] [CrossRef] [Green Version]
- Bocci, G.; Fioravanti, A.; Orlandi, P.; Bernardini, N.; Collecchi, P.; Del Tacca, M.; Danesi, R. Fluvastatin synergistically enhances the antiproliferative effect of gemcitabine in human pancreatic cancer MIAPaCa-2 cells. Br. J. Cancer 2005, 93, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Diepart, C.; Verrax, J.; Calderon, P.B.; Feron, O.; Jordan, B.F.; Gallez, B. Comparison of methods for measuring oxygen consumption in tumor cells in vitro. Anal. Biochem. 2010, 396, 250–256. [Google Scholar] [CrossRef]
- D’Hose, D.; Danhier, P.; Northshield, H.; Isenborghs, P.; Jordan, B.F.; Gallez, B. A versatile EPR toolbox for the simultaneous measurement of oxygen consumption and superoxide production. Redox Biol. 2021, 40, 101852. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Harrison, D.G. Methods for Detection of Mitochondrial and Cellular Reactive Oxygen Species. Antioxid. Redox Signal. 2014, 20, 372–382. [Google Scholar] [CrossRef]
- Dikalov, S.I.; Kirilyuk, I.; Voynov, M.; Grigor’Ev, I. EPR detection of cellular and mitochondrial superoxide using cyclic hydroxylamines. Free Radic. Res. 2010, 45, 417–430. [Google Scholar] [CrossRef]
- Scheinok, S.; Capeloa, T.; Porporato, P.E.; Sonveaux, P.; Gallez, B. An EPR Study Using Cyclic Hydroxylamines to Assess the Level of Mitochondrial ROS in Superinvasive Cancer Cells. Cell Biochem. Biophys. 2020, 78, 249–254. [Google Scholar] [CrossRef]
- Beckman, J.S.; Minor, R.L., Jr.; White, C.W.; Repine, J.E.; Rosen, G.M.; Freeman, B.A. Superoxide dismutase and catalase conjugated to polyethylene glycol increases endothelial enzyme activity and oxidant resistance. J. Biol. Chem. 1988, 263, 6884–6892. [Google Scholar] [CrossRef]
- Baudelet, C.; Gallez, B. Effect of anesthesia on the signal intensity in tumors using BOLD-MRI: Comparison with flow measurements by Laser Doppler flowmetry and oxygen measurements by luminescence-based probes. Magn. Reson. Imaging 2004, 22, 905–912. [Google Scholar] [CrossRef]
- Gallez, B.; Jordan, B.F.; Baudelet, C.; Misson, P.D. Pharmacological modifications of the partial pressure of oxygen in murine tumors: Evaluation using in vivo EPR oximetry. Magn. Reson. Med. 1999, 42, 627–630. [Google Scholar] [CrossRef]
- Gallez, B.; Baudelet, C.; Jordan, B.F. Assessment of tumor oxygenation by electron paramagnetic resonance: Principles and applications. NMR Biomed. 2004, 17, 240–262. [Google Scholar] [CrossRef]
- Swartz, H.M.; Flood, A.B.; Schaner, P.E.; Halpern, H.; Williams, B.B.; Pogue, B.W.; Gallez, B.; Vaupel, P. How best to interpret measures of levels of oxygen in tissues to make them effective clinical tools for care of patients with cancer and other oxygen-dependent pathologies. Physiol. Rep. 2020, 8, e14541. [Google Scholar] [CrossRef] [PubMed]
- Gallez, B. Oxygenation Status in Normal Tissues, Pathological Tissues and Malignant Tumors: A pO2 Database Based on Electron Paramagnetic Resonance (EPR) Oximetry Measurements. Appl. Magn. Reson. 2021, 52, 1395–1450. [Google Scholar] [CrossRef]
- Jordan, B.F.; Baudelet, C.; Gallez, B. Carbon-centered radicals as oxygen sensors for in vivo electron paramagnetic resonance: Screening for an optimal probe among commercially available charcoals. Magn. Reson. Mater. Phys. Biol. Med. 1998, 7, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Begg, A.C. Principles and practice of the tumor growth delay assay. In Rodent Tumor Models in Experimental Cancer Therapy; Kallmann, R.F., Ed.; Pergamon Press: New York, NY, USA, 1987; pp. 114–121. [Google Scholar]
- Zips, D.; Thames, H.D.; Baumann, M. New anticancer agents: In vitro and in vivo evaluation. In Vivo 2005, 19, 1–7. [Google Scholar]
- Jezek, J.; Cooper, K.F.; Strich, R. Reactive Oxygen Species and Mitochondrial Dynamics: The Yin and Yang of Mitochondrial Dysfunction and Cancer Progression. Antioxidants 2018, 7, 13. [Google Scholar] [CrossRef]
- Kalyanaraman, B.; Cheng, G.; Hardy, M.; Ouari, O.; Bennett, B.; Zielonka, J. Teaching the basics of reactive oxygen species and their relevance to cancer biology: Mitochondrial reactive oxygen species detection, redox signaling, and targeted therapies. Redox Biol. 2018, 15, 347–362. [Google Scholar] [CrossRef]
- Manda, K.; Juerß, D.; Fischer, P.; Schröder, A.; Koenen, A.; Hildebrandt, G. Simvastatin treatment varies the radiation response of human breast cells in 2D or 3D culture. Investig. New Drugs 2021, 39, 658–669. [Google Scholar] [CrossRef]
- Chen, Y.-A.; Shih, H.-W.; Lin, Y.-C.; Hsu, H.-Y.; Wu, T.-F.; Tsai, C.-H.; Wu, C.-L.; Wu, H.-Y.; Hsieh, J.-T.; Tang, C.-H.; et al. Simvastatin Sensitizes Radioresistant Prostate Cancer Cells by Compromising DNA Double-Strand Break Repair. Front. Pharmacol. 2018, 9, 600. [Google Scholar] [CrossRef] [Green Version]
- Karagkounis, G.; DeVecchio, J.; Ferrandon, S.; Kalady, M.F. Simvastatin enhances radiation sensitivity of colorectal cancer cells. Surg. Endosc. 2018, 32, 1533–1539. [Google Scholar] [CrossRef]
- McKenney, J.M. Pharmacologic characteristics of statins. Clin. Cardiol. 2003, 26 (Suppl. 3), III32–III38. [Google Scholar] [CrossRef]
- Bouitbir, J.; Sanvee, G.M.; Panajatovic, M.V.; Singh, F.; Krähenbühl, S. Mechanisms of statin-associated skeletal muscle-associated symptoms. Pharmacol. Res. 2020, 154, 104201. [Google Scholar] [CrossRef]
- Danhier, P.; Copetti, T.; De Preter, G.; Leveque, P.; Feron, O.; Jordan, B.F.; Sonveaux, P.; Gallez, B. Influence of cell detachment on the respiration rate of tumor and endothelial cells. PLoS ONE 2013, 8, e53324. [Google Scholar] [CrossRef] [Green Version]
- Whillans, D.W.; Hunt, J.W. A rapid-mixing comparison of the mechanisms of radiosensitization by oxygen and misonidazole in CHO cells. Radiat. Res. 1982, 90, 126–141. [Google Scholar] [CrossRef]
- Koch, C.J.; Stobbe, C.C.; Bump, E.A. The effect on the Km for radiosensitization at 0 degree C of thiol depletion by diethylmaleate pretreatment: Quantitative differences found using the radiation sensitizing agent misonidazole or oxygen. Radiat. Res. 1984, 98, 141–153. [Google Scholar] [CrossRef]
- Björkhem-Bergman, L.; Lindh, J.D.; Bergman, P. What is a relevant statin concentration in cell experiments claiming pleiotropic effects? Br. J. Clin. Pharmacol. 2011, 72, 164–165. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
d’Hose, D.; Mignion, L.; Hamelin, L.; Sonveaux, P.; Jordan, B.F.; Gallez, B. Statins Alleviate Tumor Hypoxia in Prostate Cancer Models by Decreasing Oxygen Consumption: An Opportunity for Radiosensitization? Biomolecules 2022, 12, 1418. https://doi.org/10.3390/biom12101418
d’Hose D, Mignion L, Hamelin L, Sonveaux P, Jordan BF, Gallez B. Statins Alleviate Tumor Hypoxia in Prostate Cancer Models by Decreasing Oxygen Consumption: An Opportunity for Radiosensitization? Biomolecules. 2022; 12(10):1418. https://doi.org/10.3390/biom12101418
Chicago/Turabian Styled’Hose, Donatienne, Lionel Mignion, Loïc Hamelin, Pierre Sonveaux, Bénédicte F. Jordan, and Bernard Gallez. 2022. "Statins Alleviate Tumor Hypoxia in Prostate Cancer Models by Decreasing Oxygen Consumption: An Opportunity for Radiosensitization?" Biomolecules 12, no. 10: 1418. https://doi.org/10.3390/biom12101418