
Engineering of Thermal Stability in a Cold-Active Oligo-1,6-Glucosidase from Exiguobacterium sibiricum with Unusual Amino Acid Content

 , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
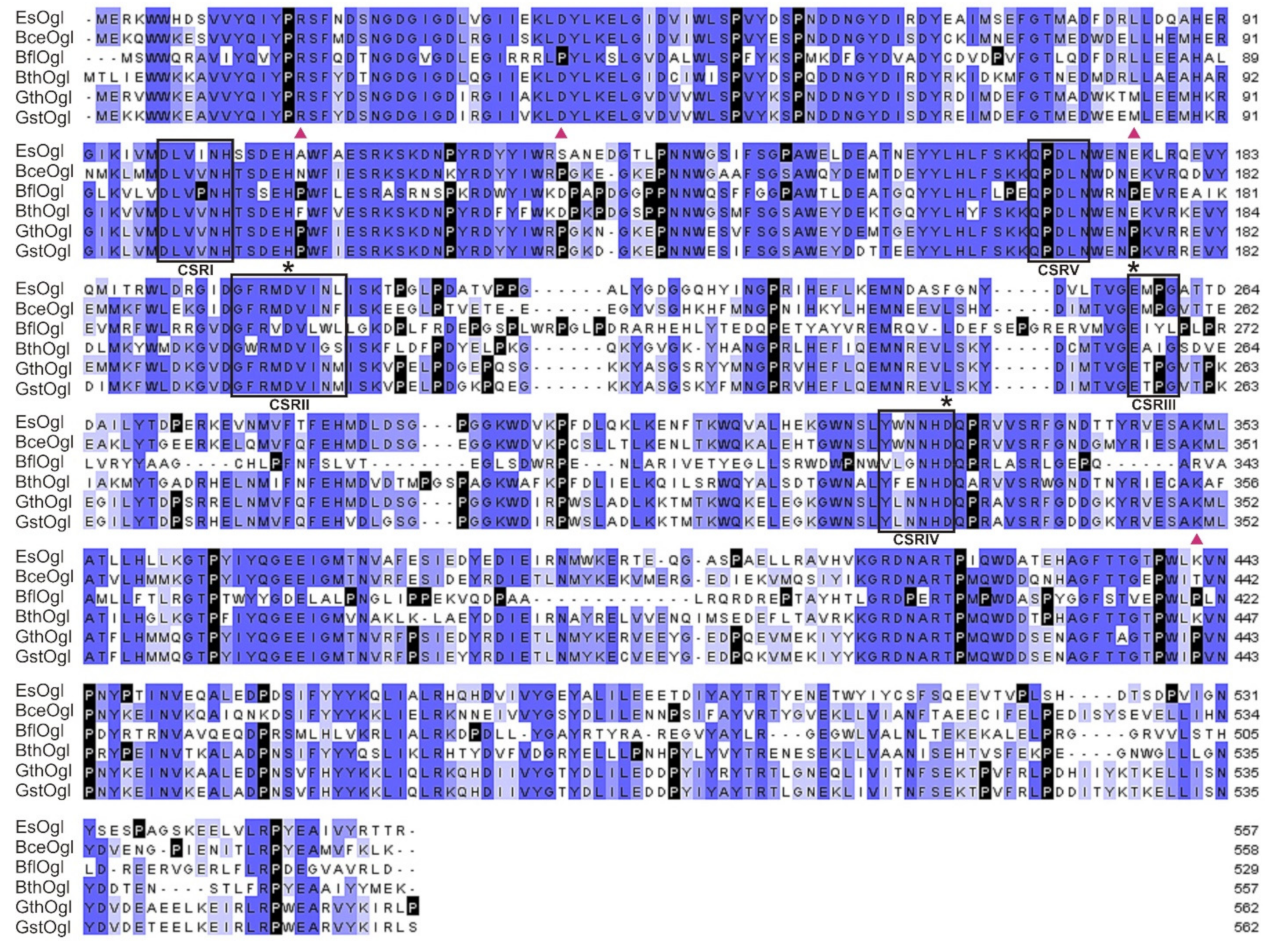
2.1. Bioinformatic Analysis
2.2. EsOgl Gene Cloning and Expression
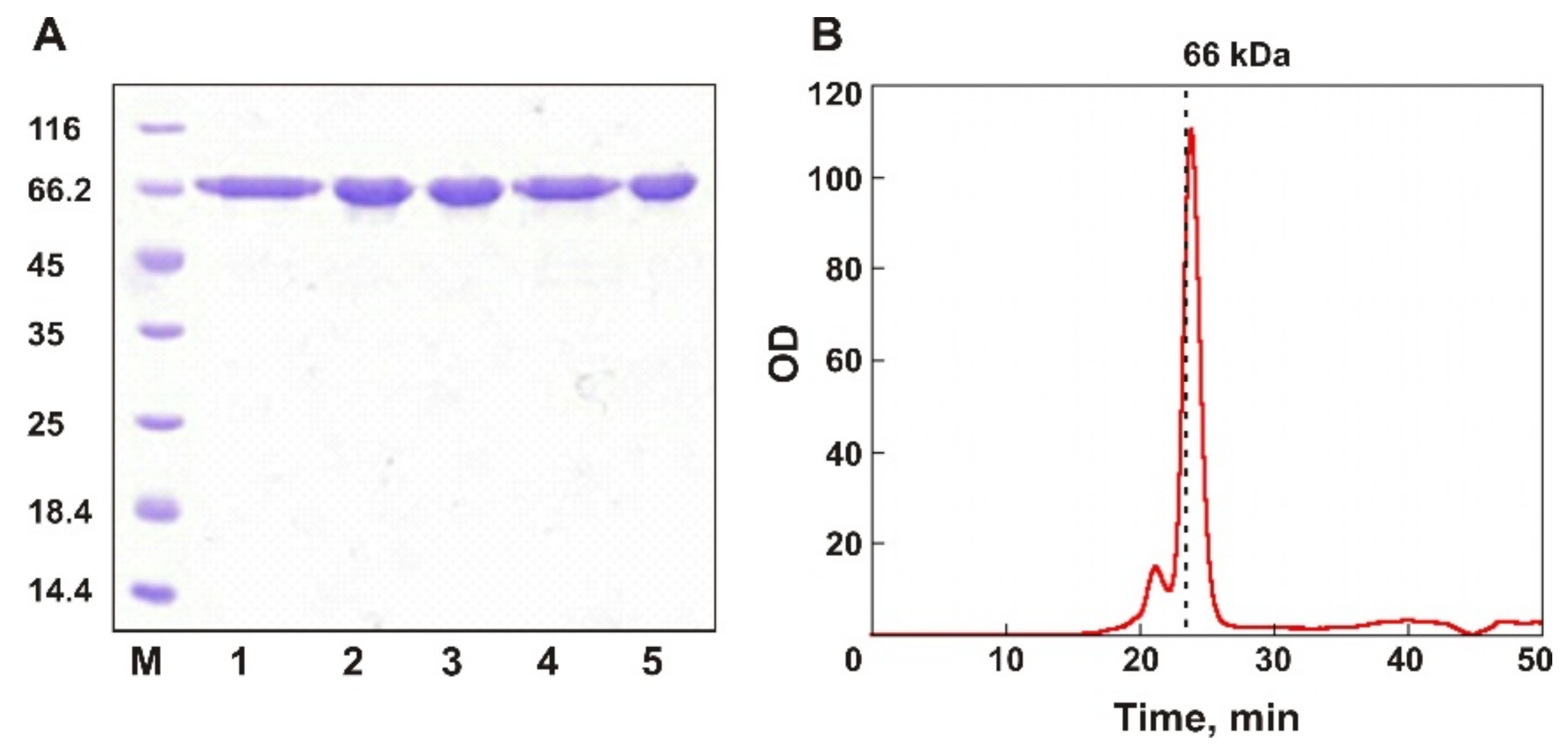
2.3. Protein Purification
2.4. Biochemical Characterization of the Enzyme
2.5. Construction and Characterization of the Mutant Variants
2.6. SEC Analysis
2.7. Circular Dichroism Spectra
3. Results
3.1. Sequence Analysis and Gene Cloning of EsOgl
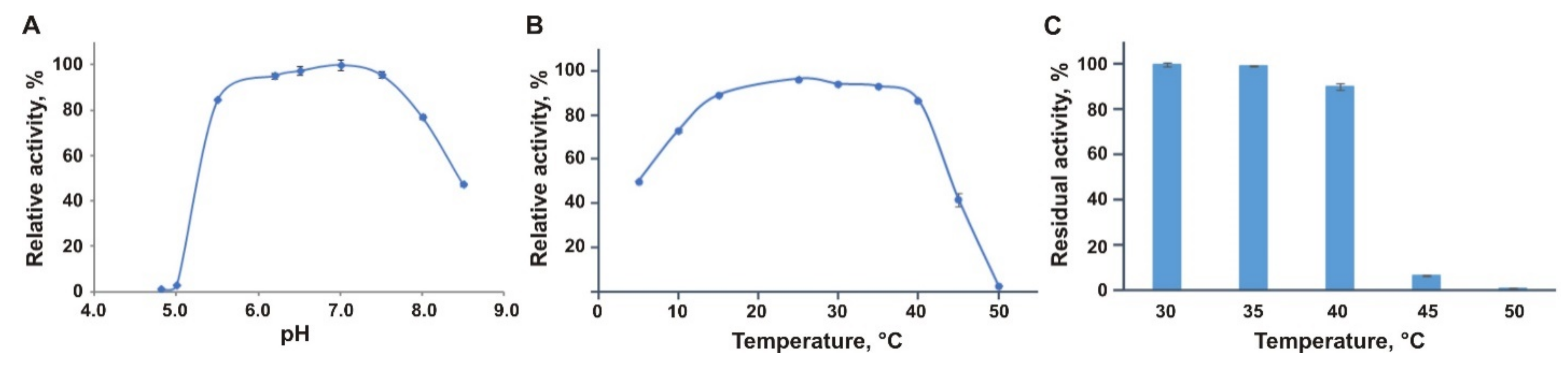
3.2. Biochemical Characterization of EsOgl
3.3. Thermal Stability of EsOgl
3.4. Kinetic Characteristics of the Enzyme
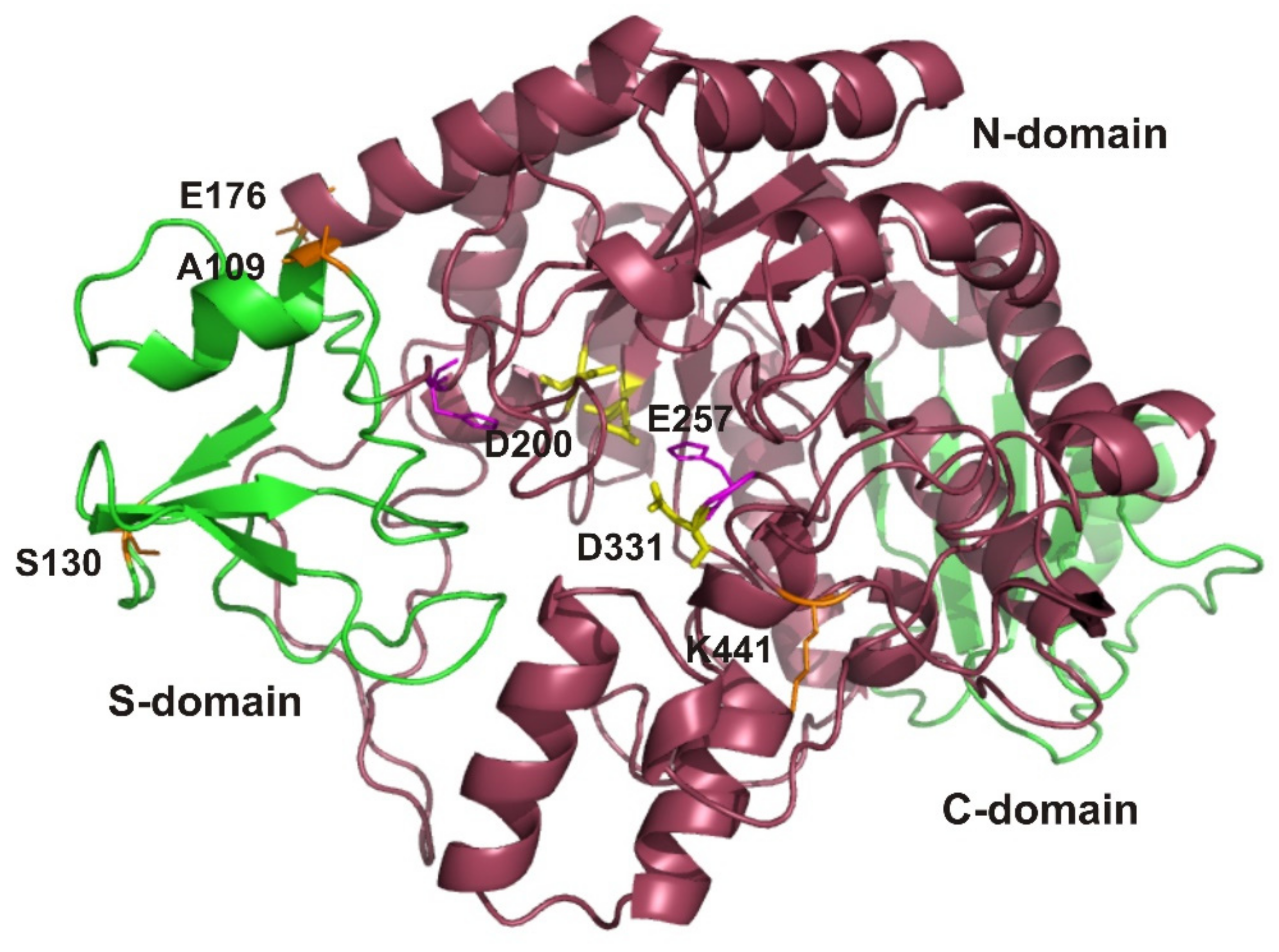
3.5. EsOgl Structure Modeling and Site-Directed Mutagenesis
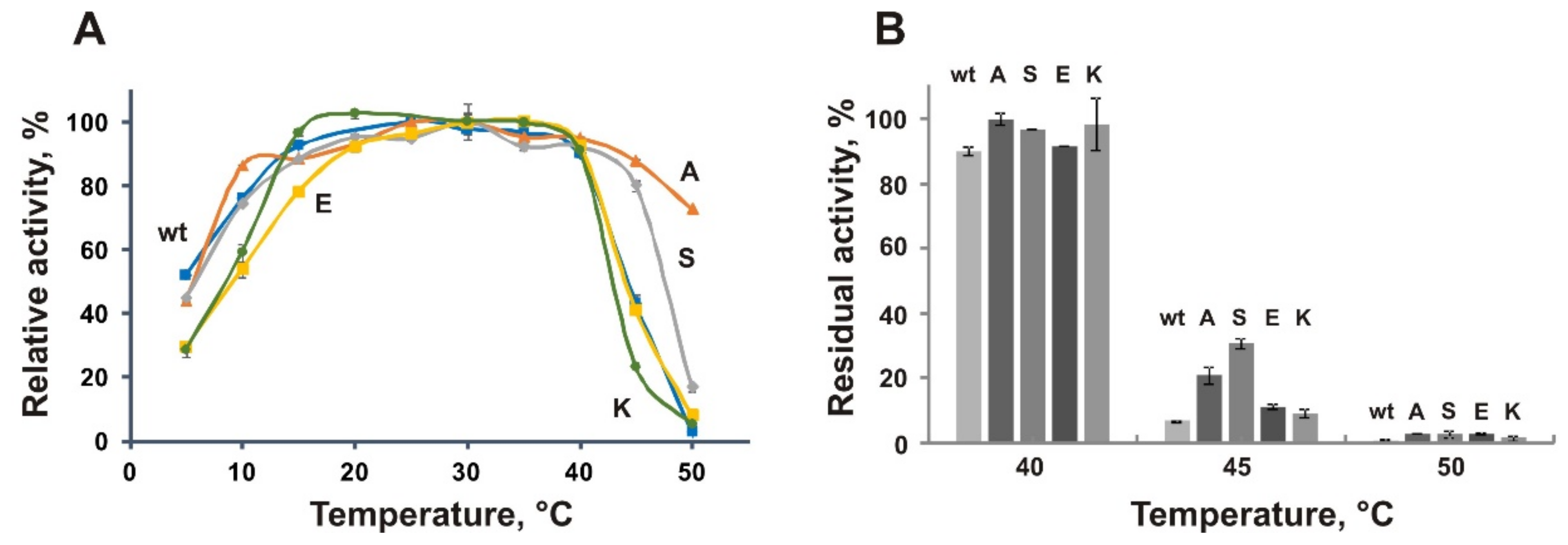
3.6. Biochemical Characterization of the Mutant Proteins A109P, S130P, E176P, and K441P
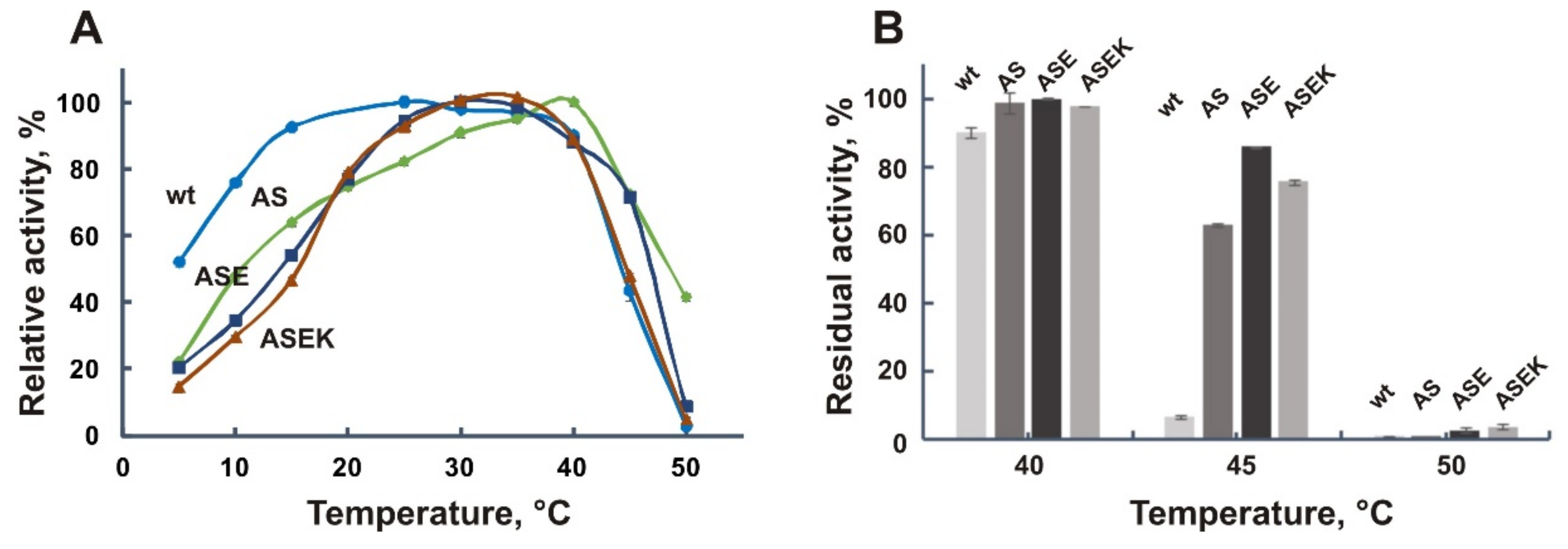
3.7. Effect of the Combined Mutations
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hua, X.; Yang, R. Enzymes in starch processing. In Enzymes in Food and Beverage Processing; Chandrasekaran, M., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 139–170. [Google Scholar] [CrossRef]
- Van der Maarel, M.J.; van der Veen, B.; Uitdehaag, J.C.; Leemhuis, H.; Dijkhuizen, L. Properties and applications of starch-converting enzymes of the α-amylase family. J. Biotechnol. 2002, 94, 137–155. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Tang, C.; Lu, Y.; Yao, L.; Kan, Y. Microbial Oligo-α-1,6-Glucosidase: Current Developments and Future Perspectives. Starch Stärke 2020, 72, 1900172. [Google Scholar] [CrossRef]
- Watanabe, K.; Miyake, K.; Suzuki, Y. Identification of Catalytic and Substrate-binding Site Residues in Bacillus cereus ATCC7064 Oligo-1,6-glucosidase. Biosci. Biotechnol. Biochem. 2001, 65, 2058–2064. [Google Scholar] [CrossRef]
- Watanabe, K.; Kitamura, K.; Suzuki, Y. Analysis of the critical sites for protein thermostabilization by proline substitution in oligo-1,6-glucosidase from Bacillus coagulans ATCC 7050 and the evolutionary consideration of proline residues. Appl. Envrion. Microbiol. 1996, 62, 2066–2073. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Iha, H.; Ohashi, A.; Suzuki, Y. Cloning and expression in Escherichia coli of an extremely thermostable oligo-1,6-glucosidase gene from Bacillus thermoglucosidasius. J. Bacteriol. 1989, 171, 1219–1222. [Google Scholar] [CrossRef] [Green Version]
- Kashiwabara, S.-I.; Matsuki, Y.; Kishimoto, T.; Suzuki, Y. Clustered Proline Residues around the Active-Site Cleft in Thermostable Oligo-1,6-glucosidase of Bacillus flavocaldarius KP1228. Biosci. Biotechnol. Biochem. 1998, 62, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, K.S.; Cavicchioli, R. Cold-Adapted Enzymes. Annu. Rev. Biochem. 2006, 75, 403–433. [Google Scholar] [CrossRef] [Green Version]
- Feller, G. Psychrophilic Enzymes: From Folding to Function and Biotechnology. Science 2013, 2013, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Littlechild, J.A. Enzymes from Extreme Environments and Their Industrial Applications. Front. Bioeng. Biotechnol. 2015, 3, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pucci, F.; Rooman, M. Physical and molecular bases of protein thermal stability and cold adaptation. Curr. Opin. Struct. Biol. 2017, 42, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Bakermans, C. Psychrophiles: Life in the cold. In Extremophiles: Microbiology and Biotechnology; Anitoris, R.P., Ed.; Caister Academic Press: Poole, UK, 2012; Volume 3, pp. 53–76. [Google Scholar]
- De Maayer, P.; Anderson, D.; Cary, S.; Cowan, D.A. Some like it cold: Understanding the survival strategies of psychrophiles. EMBO Rep. 2014, 15, 508–517. [Google Scholar] [CrossRef]
- Collins, T.; Margesin, R. Psychrophilic lifestyles: Mechanisms of adaptation and biotechnological tools. Appl. Microbiol. Biotechnol. 2019, 103, 2857–2871. [Google Scholar] [CrossRef] [PubMed]
- Rivkina, E.; Laurinavichius, K.; McGrath, J.; Tiedje, J.; Shcherbakova, V.; Gilichinsky, D. Microbial life in permafrost. Adv. Space Res. 2004, 33, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Vishnivetskaya, T.A.; Petrova, M.; Urbance, J.; Ponder, M.; Moyer, C.L.; Gilichinsky, D.A.; Tiedje, J.M. Bacterial Community in Ancient Siberian Permafrost as Characterized by Culture and Culture-Independent Methods. Astrobiology 2006, 6, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Hinsa-Leasure, S.; Bakermans, C. Diversity of Bacteria in Permafrost. In Cold-Adapted Microorganisms; Yumoto, I., Ed.; Caister Academic Press: Poole, UK, 2013; Volume 1, p. 1. [Google Scholar]
- Jansson, J.; Taş, N. The microbial ecology of permafrost. Nat. Rev. Genet. 2014, 12, 414–425. [Google Scholar] [CrossRef]
- Margesin, R.; Collins, T. Microbial ecology of the cryosphere (glacial and permafrost habitats): Current knowledge. Appl. Microbiol. Biotechnol. 2019, 103, 2537–2549. [Google Scholar] [CrossRef] [Green Version]
- Petrovskaya, L.E.; Novototskaya-Vlasova, K.A.; Spirina, E.V.; Khokhlova, G.V.; Rivkina, E.M.; Gilichinsky, D.A.; Dolgikh, D.A.; Kirpichnikov, M. Lipolytic enzymes of microorganisms from permafrost cryopegs. Dokl. Biol. Sci. 2012, 445, 279–282. [Google Scholar] [CrossRef]
- Novototskaya-Vlasova, K.A.; Petrovskaya, L.E.; Rivkina, E.; Dolgikh, D.; Kirpichnikov, M. Characterization of a cold-active lipase from Psychrobacter cryohalolentis K5T and its deletion mutants. Biochemistry 2013, 78, 385–394. [Google Scholar] [CrossRef]
- Boyko, K.M.; Kryukova, M.V.; Petrovskaya, L.E.; Nikolaeva, A.Y.; Korzhenevsky, D.A.; Novototskaya-Vlasova, K.A.; Rivkina, E.M.; Dolgikh, D.A.; Kirpichnikov, M.; Popov, V. Crystal structure of PMGL2 esterase from the hormone-sensitive lipase family with GCSAG motif around the catalytic serine. PLoS ONE 2020, 15, e0226838. [Google Scholar] [CrossRef]
- Bezsudnova, E.Y.; Stekhanova, T.N.; Popinako, A.V.; Rakitina, T.V.; Nikolaeva, A.Y.; Boyko, K.M.; Popov, V. Diaminopelargonic acid transaminase from Psychrobacter cryohalolentis is active toward (S)-(-)-1-phenylethylamine, aldehydes and α-diketones. Appl. Microbiol. Biotechnol. 2018, 102, 9621–9633. [Google Scholar] [CrossRef]
- Löwe, J.; Ingram, A.A.; Gröger, H. Enantioselective synthesis of amines via reductive amination with a dehydrogenase mutant from Exigobacterium sibiricum: Substrate scope, co-solvent tolerance and biocatalyst immobilization. Bioorg. Med. Chem. 2018, 26, 1387–1392. [Google Scholar] [CrossRef]
- Santiago, M.; Ramírez, C.; Zamora, R.; Parra, L.P. Discovery, Molecular Mechanisms, and Industrial Applications of Cold-Active Enzymes. Front. Microbiol. 2016, 7, 1408. [Google Scholar] [CrossRef] [PubMed]
- Barroca, M.; Santos, G.D.A.; Gerday, C.; Collins, T. Biotechnological Aspects of Cold-Active Enzymes. In Psychrophiles: From Biodiversity to Biotechnology; Springer Science and Business Media LLC: Berlin, Germany, 2017; pp. 461–475. [Google Scholar]
- Al-Ghanayem, A.A.; Joseph, B. Current prospective in using cold-active enzymes as eco-friendly detergent additive. Appl. Microbiol. Biotechnol. 2020, 104, 2871–2882. [Google Scholar] [CrossRef]
- Yang, H.; Liu, L.; Li, J.; Chen, J.; Du, G. Rational Design to Improve Protein Thermostability: Recent Advances and Prospects. Chembioeng. Rev. 2015, 2, 87–94. [Google Scholar] [CrossRef]
- Denard, C.A.; Ren, H.; Zhao, H. Improving and repurposing biocatalysts via directed evolution. Curr. Opin. Chem. Biol. 2015, 25, 55–64. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. Protein Engineering as a Tool for the Development of Novel Bioproduction Systems. Blue Biotechnol. 2013, 137, 25–40. [Google Scholar] [CrossRef]
- Qu, G.; Li, A.; Acevedo-Rocha, C.G.; Sun, Z.; Reetz, M.T. The Crucial Role of Methodology Development in Directed Evolution of Selective Enzymes. Angew. Chem. Int. Ed. 2020, 59, 13204–13231. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.F.; Ivanova, N.; He, Z.; Huebner, M.; Zhou, J.; Tiedje, J.M. Architecture of thermal adaptation in an Exiguobacterium sibiricum strain isolated from 3 million year old permafrost: A genome and transcriptome approach. BMC Genom. 2008, 9, 547. [Google Scholar] [CrossRef] [Green Version]
- ClustalOmega. Available online: https://www.ebi.ac.uk/Tools/msa/clustalo/ (accessed on 9 March 2021).
- ProtParam. Available online: http://web.expasy.org/protparam/ (accessed on 18 March 2021).
- SWISS-MODEL. Available online: https://swissmodel.expasy.org (accessed on 24 March 2021).
- DichroWeb. Available online: http://pcddb.cryst.bbk.ac.uk (accessed on 12 May 2021).
- Oslancová, A.; Janeček, Š. Oligo-1,6-glucosidase and neopullulanase enzyme subfamilies from the α-amylase family defined by the fifth conserved sequence region. Cell. Mol. Life Sci. 2002, 59, 1945–1959. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Kitamura, K.; Iha, H.; Suzuki, Y. Primary structure of the oligo-1,6-glucosidase of Bacillus cereus ATCC7064 deduced from the nucleotide sequence of the cloned gene. JBIC J. Biol. Inorg. Chem. 1990, 192, 609–620. [Google Scholar] [CrossRef]
- Tsujimoto, Y.; Tanaka, H.; Takemura, R.; Yokogawa, T.; Shimonaka, A.; Matsui, H.; Kashiwabara, S.-I.; Watanabe, K.; Suzuki, Y. Molecular determinants of substrate recognition in thermostable α-glucosidases belonging to glycoside hydrolase family 13. J. Biochem. 2007, 142, 87–93. [Google Scholar] [CrossRef]
- Watanabe, K.; Masuda, T.; Ohashi, H.; Mihara, H.; Suzuki, Y. Multiple Proline Substitutions Cumulatively Thermostabilize Bacillus Cereus ATCC7064 Oligo-1,6-Glucosidase. Irrefragable Proof Supporting the Proline Rule. JBIC J. Biol. Inorg. Chem. 1994, 226, 277–283. [Google Scholar] [CrossRef]
- Land, H.; Campillo-Brocal, J.C.; Humble, M.S.; Berglund, P. B-factor Guided Proline Substitutions in Chromobacterium violaceum Amine Transaminase: Evaluation of the Proline Rule as a Method for Enzyme Stabilization. ChemBioChem 2019, 20, 1297–1304. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Hata, Y.; Kizaki, H.; Katsube, Y.; Suzuki, Y. The refined crystal structure of Bacillus cereus oligo-1,6-glucosidase at 2.0 å resolution: Structural characterization of proline-substitution sites for protein thermostabilization. J. Mol. Biol. 1997, 269, 142–153. [Google Scholar] [CrossRef]
- Rodrigues, D.F.; Goris, J.; Vishnivetskaya, T.A.; Gilichinsky, D.; Thomashow, M.F.; Tiedje, J.M. Characterization of Exiguobacterium isolates from the Siberian permafrost. Description of Exiguobacterium sibiricum sp. nov. Extremophiles 2006, 10, 285–294. [Google Scholar] [CrossRef]
- Gianese, G.; Argos, P.; Pascarella, S. Structural adaptation of enzymes to low temperatures. Protein Eng. Des. Sel. 2001, 14, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Delgado, S.; Flórez, A.B.; Guadamuro, L.; Mayo, B. Genetic and biochemical characterization of an oligo-α-1,6-glucosidase from Lactobacillus plantarum. Int. J. Food Microbiol. 2017, 246, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Leiros, H.-K.S.; Willassen, N.P.; Smalås, A.O. Residue determinants and sequence analysis of cold-adapted trypsins. Extremophiles 1999, 3, 205–219. [Google Scholar] [CrossRef]
- Georlette, D.; Jónsson, Z.O.; Van Petegem, F.; Chessa, J.-P.; Van Beeumen, J.; Hübscher, U.; Gerday, C. A DNA ligase from the psychrophile Pseudoalteromonas haloplanktis gives insights into the adaptation of proteins to low temperatures. JBIC J. Biol. Inorg. Chem. 2000, 267, 3502–3512. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J. Amino acid preferences for specific locations at the ends of alpha helices. Science 1988, 240, 1648–1652. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Chishiro, K.; Kitamura, K.; Suzuki, Y. Proline residues responsible for thermostability occur with high frequency in the loop regions of an extremely thermostable oligo-1,6-glucosidase from Bacillus thermoglucosidasius KP1006. J. Biol. Chem. 1991, 266, 24287–24294. [Google Scholar] [CrossRef]
- Tian, J.; Wang, P.; Gao, S.; Chu, X.; Wu, N.; Fan, Y. Enhanced thermostability of methyl parathion hydrolase from Ochrobactrum sp. M231 by rational engineering of a glycine to proline mutation. FEBS J. 2010, 277, 4901–4908. [Google Scholar] [CrossRef]
- Watanabe, K.; Suzuki, Y. Protein thermostabilization by proline substitutions. J. Mol. Catal. B Enzym. 1998, 4, 167–180. [Google Scholar] [CrossRef]
- Kulakova, L.; Galkin, A.; Nakayama, T.; Nishino, T.; Esaki, N. Cold-active esterase from Psychrobacter sp. Ant300: Gene cloning, characterization, and the effects of Gly→Pro substitution near the active site on its catalytic activity and stability. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2004, 1696, 59–65. [Google Scholar] [CrossRef]
- Cipolla, A.; D’Amico, S.; Barumandzadeh, R.; Matagne, A.; Feller, G. Stepwise Adaptations to Low Temperature as Revealed by Multiple Mutants of Psychrophilic α-Amylase from Antarctic Bacterium. J. Biol. Chem. 2011, 286, 38348–38355. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of the Enzyme, Reference | Optimum/Assay Temperature, °C | T °C at Which t1/2 = 10 min | Km, mM | kcat, s−1 | Pro Number |
|---|---|---|---|---|---|
| E. sibiricum (this work) | 20–35 */25 | ~45 ** | 1.06 | 260 | 28 |
| B. cereus [4] | 41/35 | 44 | 0.80 | 483 | 19 |
| B. coagulans [5] | 62/50 | 61 | 0.17 | 253 | 24 |
| B. thermoglucosidasius [6,39] | 70–75/60 | 71 | 0.24 | 233 | 32 |
| Additive | Concentration | Relative Activity, % |
|---|---|---|
| CaCl2 | 1 mM | 99 ± 2 |
| MgCl2 | 103 ± 2 | |
| NiCl2 | 105 ± 3 | |
| MnCl2 | 98 ± 5 | |
| CuCl2 | 94 ± 5 | |
| CoCl2 | 94 ± 1 | |
| KCl | 102 ± 1 | |
| β-MЭ | 101 ± 2 | |
| EDTA | 92 ± 1 | |
| PMSF | 93 ± 4 | |
| Triton X-100 | 0.05% | 90 ± 1 |
| Tween-20 | 89 ± 3 | |
| SDS | 86 ± 3 | |
| Triton X-100 | 0.5% | 92 ± 1 |
| Tween-20 | 96 ± 1 | |
| Ethanol | 10% | 85 ± 1 |
| Methanol | 95 ± 2 | |
| iPrOH | 67 ± 1 | |
| Acetonitrile | 48 ± 3 | |
| Acetone | 56 ± 2 | |
| DMSO | 79 ± 1 | |
| Ethanol | 20% | 41 ± 1 |
| Methanol | 57 ± 1 | |
| iPrOH | 9 ± 2 | |
| Acetonitrile | 0.7 ± 0.1 | |
| Acetone | 9 ± 2 | |
| DMSO | 35 ± 1 |
| Enzyme | Topt °C * | Residual Activity after 45 °C, % | t1/2 at 45 °C, min | kin, min−1 | Km, mM | kcat, min−1 × 104 | kcat/KM, min−1 × mM−1 × 104 |
|---|---|---|---|---|---|---|---|
| wt EsOgl | 20–35 | 7 | 11 | 0.059 ± 0.002 | 1.06 ± 0.02 | 1.5 ± 0.2 | 1.4 |
| A109P | 23–35 | 28 | 20 | 0.033 ± 0.002 | 1.43 ± 0.09 | 1.5 ± 0.2 | 1.0 |
| S130P | 20–32 | 42 | 29 | 0.023 ± 0.001 | 1.66 ± 0.06 | 2.4 ± 0.3 | 1.5 |
| E176P | 25–37 | 15 | 16 | 0.042 ± 0.002 | 1.97 ± 0.08 | 2.0 ± 0.3 | 1.0 |
| K441P | 15–37 | 14 | 12 | 0.055 ± 0.005 | 0.93 ± 0.08 | 1.0 ± 0.1 | 1.1 |
| AS | 35–41 | 63 | 61 | 0.0109 ± 0.0002 | 1.37 ± 0.04 | 1.3 ± 0.2 | 1.0 |
| ASE | 25–36 | 86 | 129 | 0.0052 ± 0.0001 | 0.92 ± 0.07 | 1.2 ± 0.2 | 1.3 |
| ASEK | 27–37 | 76 | 88 | 0.0076 ± 0.0002 | 0.85 ± 0.02 | 0.9 ± 0.1 | 1.0 |
| Protein | α-Helix, % | ß-Sheet, % | β-Turns, % | Unordered, % |
|---|---|---|---|---|
| wt EsOgl | 14.2 | 34.2 | 19.6 | 21.8 |
| ASE | 16.4 | 31.6 | 17.4 | 34.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Berlina, Y.Y.; Petrovskaya, L.E.; Kryukova, E.A.; Shingarova, L.N.; Gapizov, S.S.; Kryukova, M.V.; Rivkina, E.M.; Kirpichnikov, M.P.; Dolgikh, D.A. Engineering of Thermal Stability in a Cold-Active Oligo-1,6-Glucosidase from Exiguobacterium sibiricum with Unusual Amino Acid Content. Biomolecules 2021, 11, 1229. https://doi.org/10.3390/biom11081229
Berlina YY, Petrovskaya LE, Kryukova EA, Shingarova LN, Gapizov SS, Kryukova MV, Rivkina EM, Kirpichnikov MP, Dolgikh DA. Engineering of Thermal Stability in a Cold-Active Oligo-1,6-Glucosidase from Exiguobacterium sibiricum with Unusual Amino Acid Content. Biomolecules. 2021; 11(8):1229. https://doi.org/10.3390/biom11081229
Chicago/Turabian StyleBerlina, Yana Y., Lada E. Petrovskaya, Elena A. Kryukova, Lyudmila N. Shingarova, Sultan Sh. Gapizov, Mariya V. Kryukova, Elizaveta M. Rivkina, Mikhail P. Kirpichnikov, and Dmitry A. Dolgikh. 2021. "Engineering of Thermal Stability in a Cold-Active Oligo-1,6-Glucosidase from Exiguobacterium sibiricum with Unusual Amino Acid Content" Biomolecules 11, no. 8: 1229. https://doi.org/10.3390/biom11081229