
Selective Fatty Acid Retention and Turnover in the Freshwater Amphipod Pallaseopsis quadrispinosa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Amphipod Sampling
2.2. Phytoplankton Culturing
2.3. Experimental Setup
2.4. Lipid and Fatty Acid Analysis
2.5. Gas Chromatography Combustion Stable Isotope Ratio Mass Spectrometry (GC-C-IRMS)
2.6. Statistics
3. Results
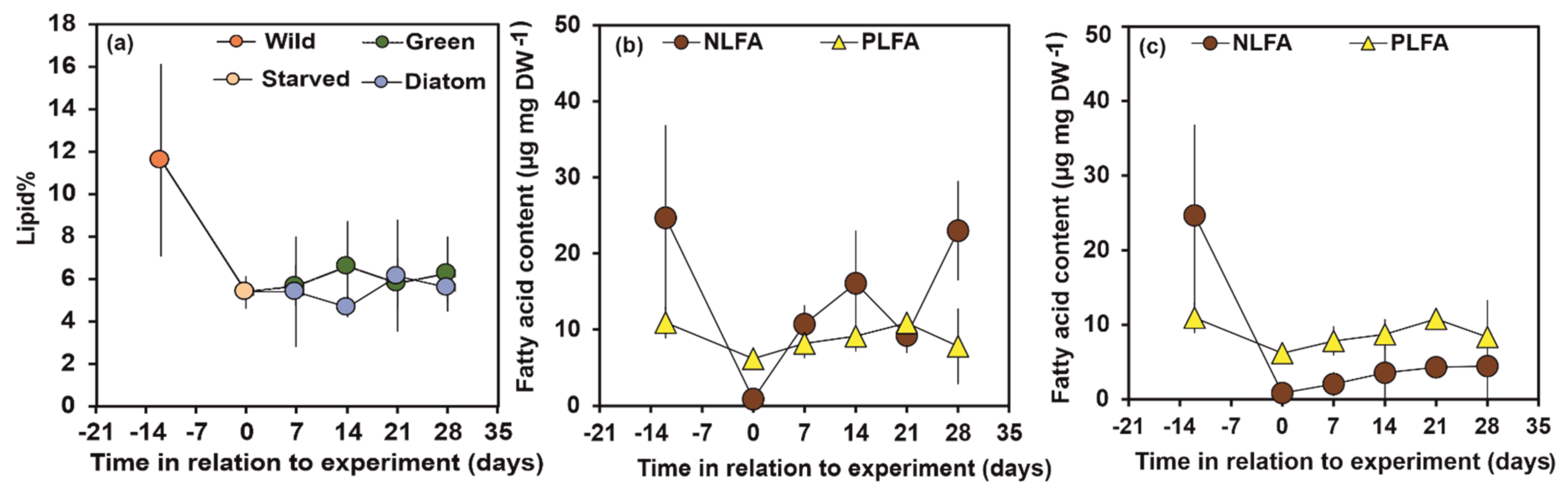
3.1. Total Lipid, NLFA and PLFA Content of Wild, Starved and Re-fed Amphipods
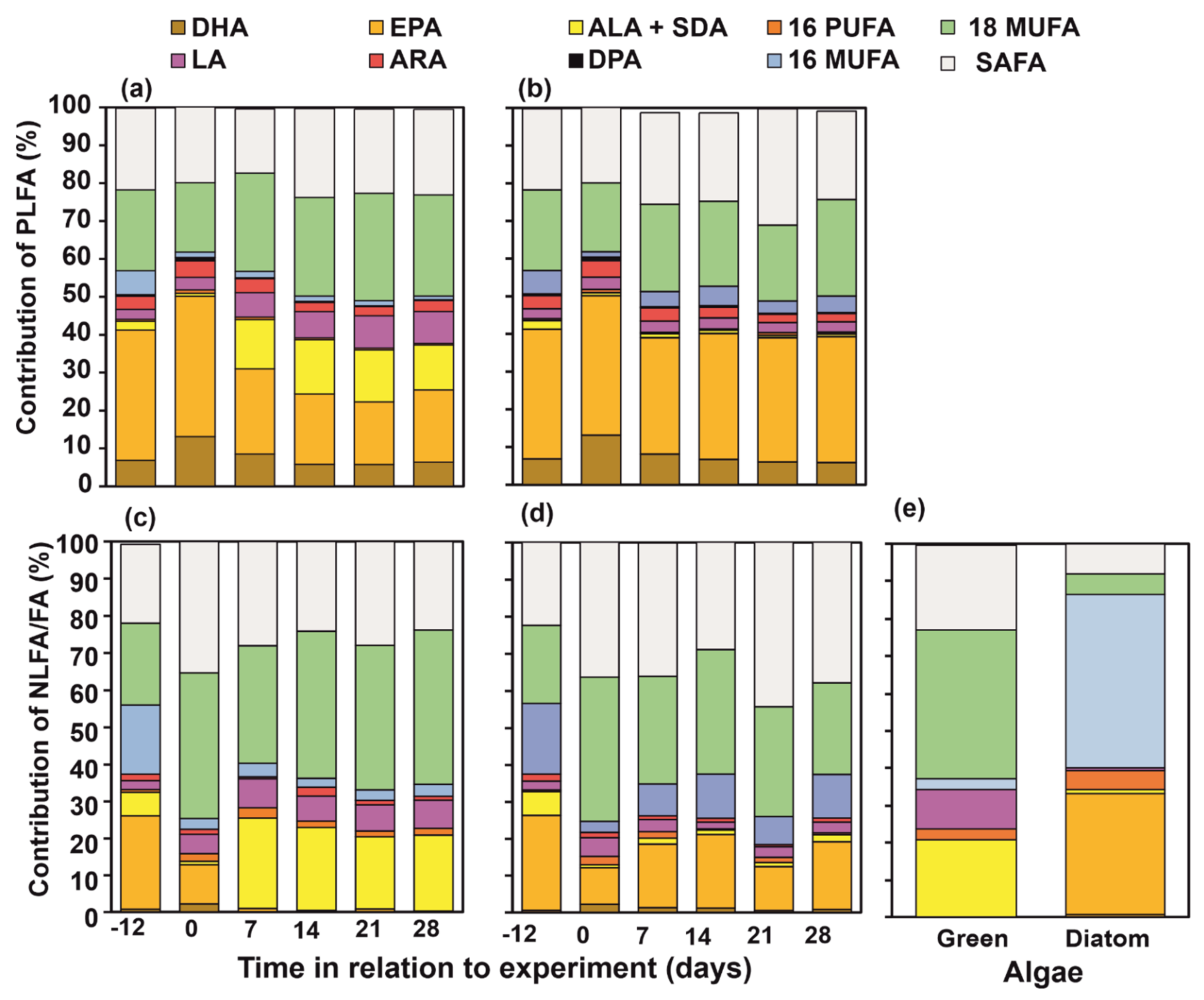
3.2. FA Profiles of Wild, Starved and Re-Fed Amphipods
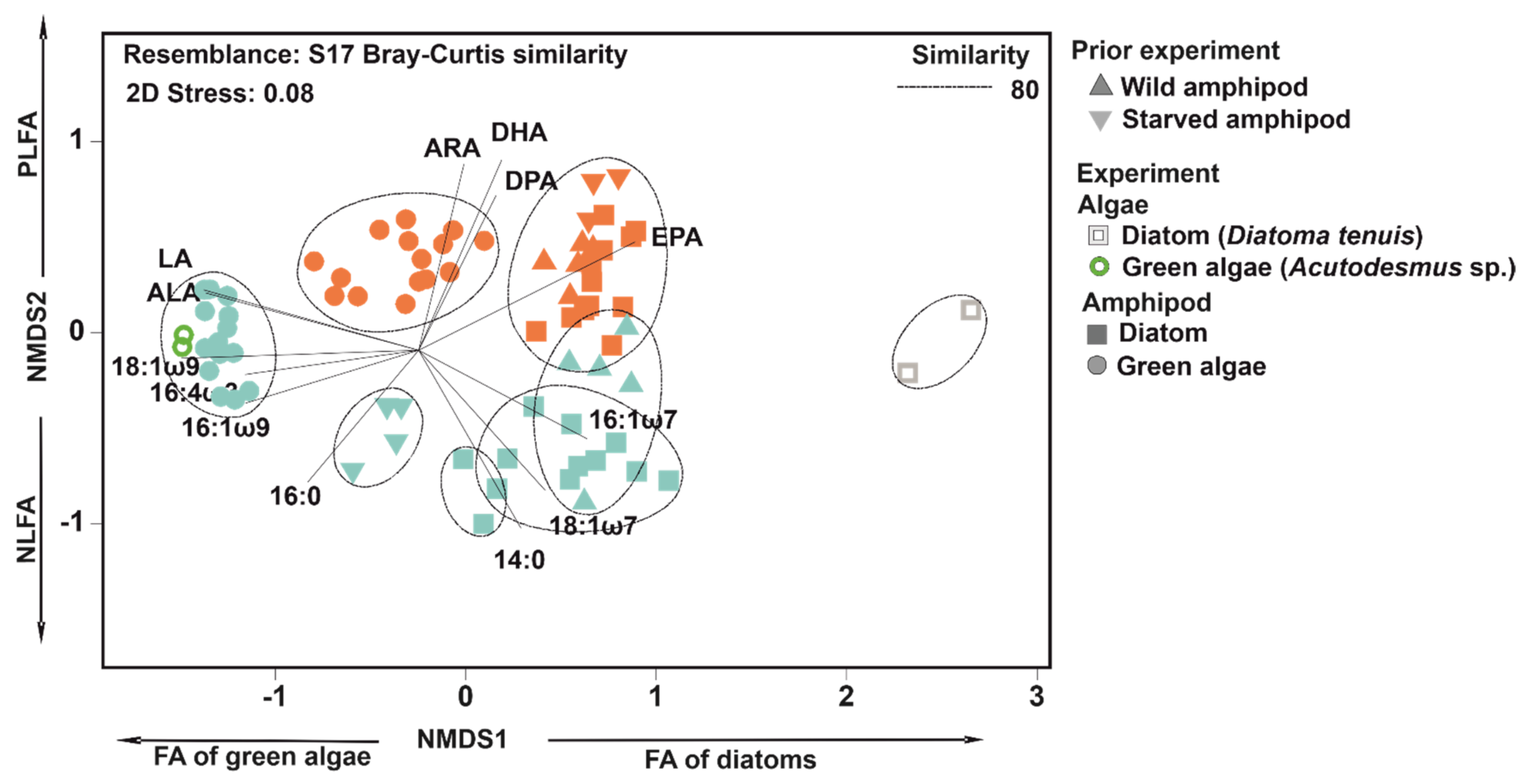
3.3. NLFA and PLFA Composition of Wild, Starved and Re-Fed Amphipods
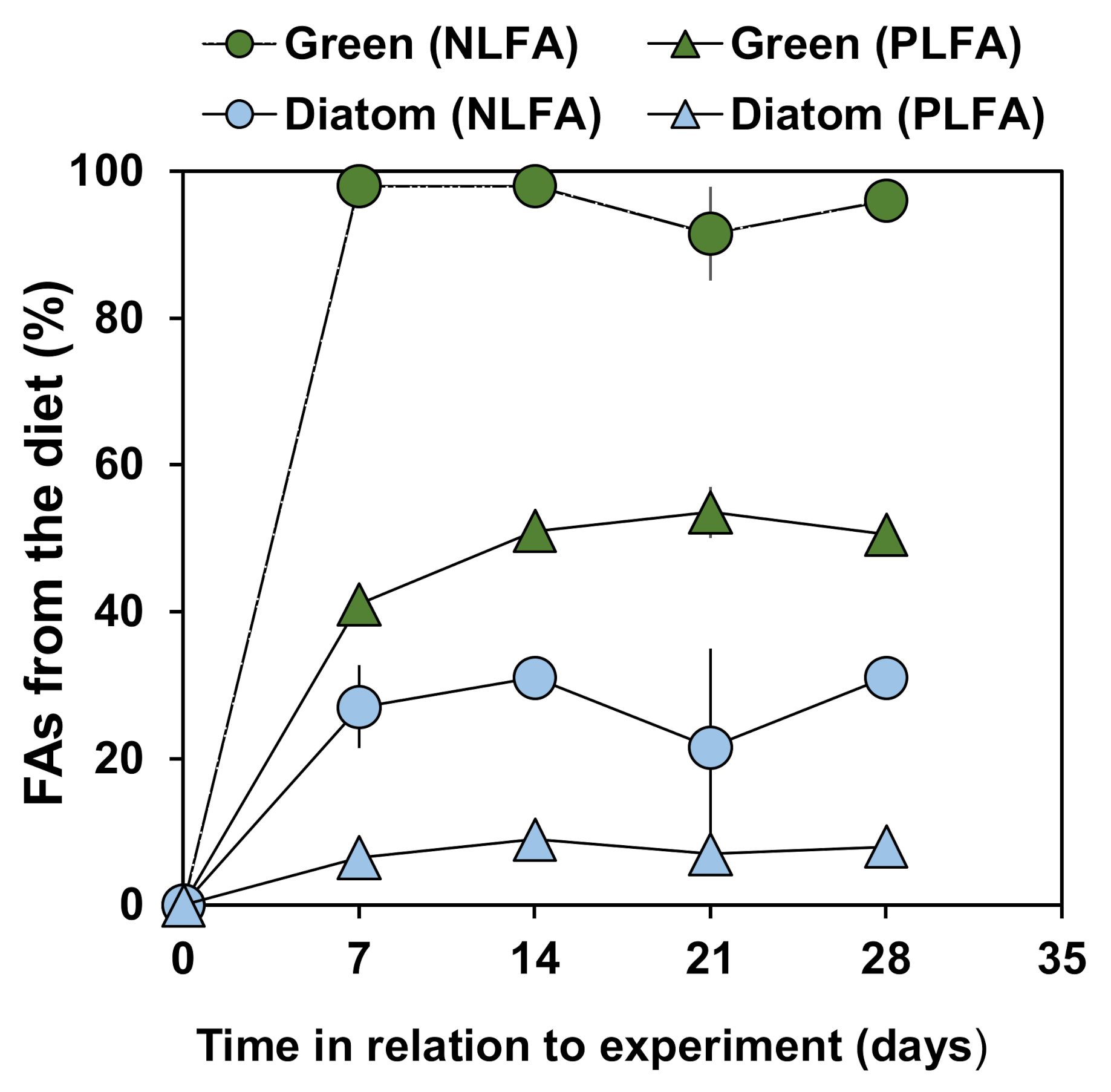
3.4. Overall Fatty Acid Turnover in Different Lipid Fraction of P. quadrispinosa
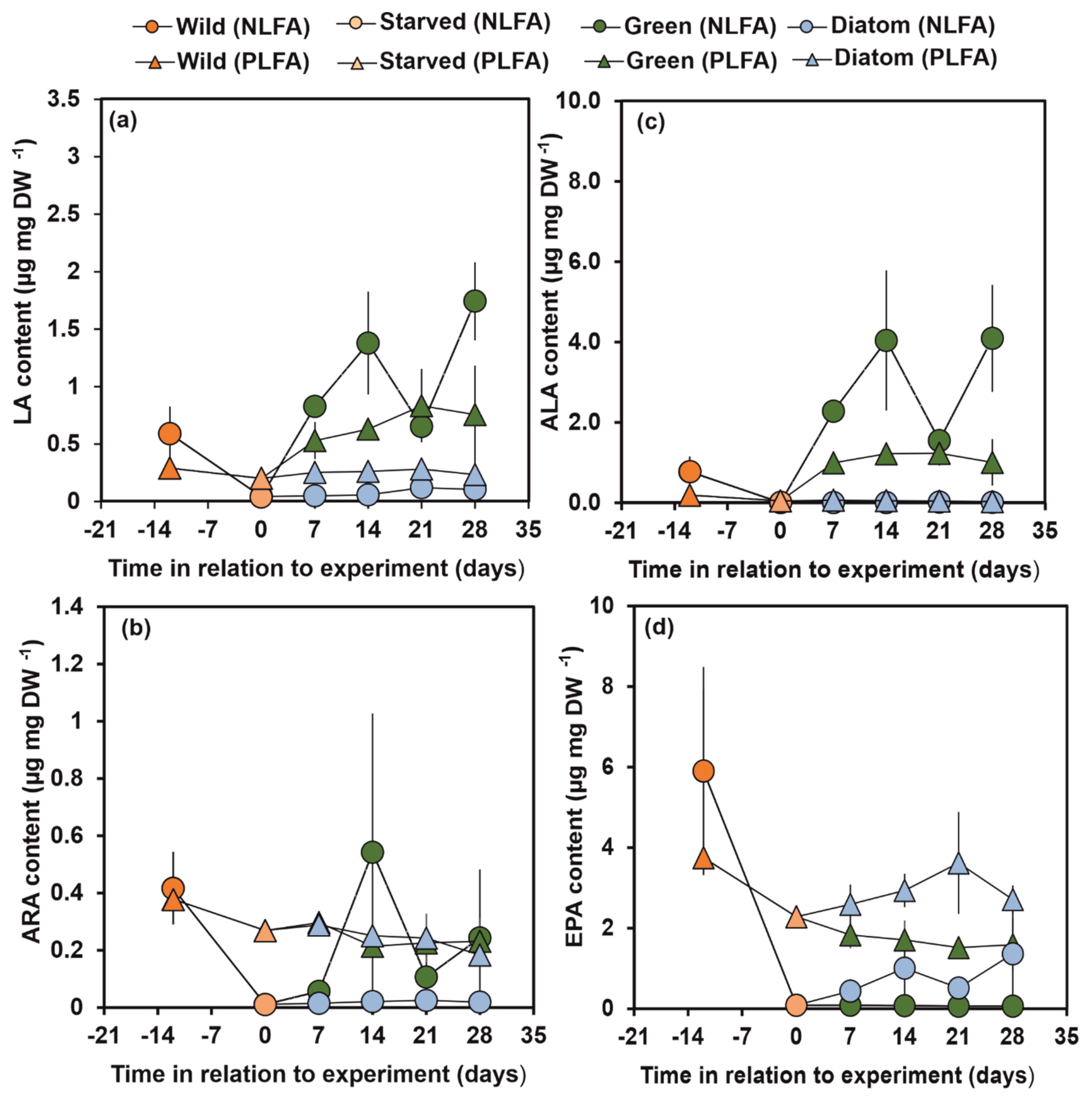
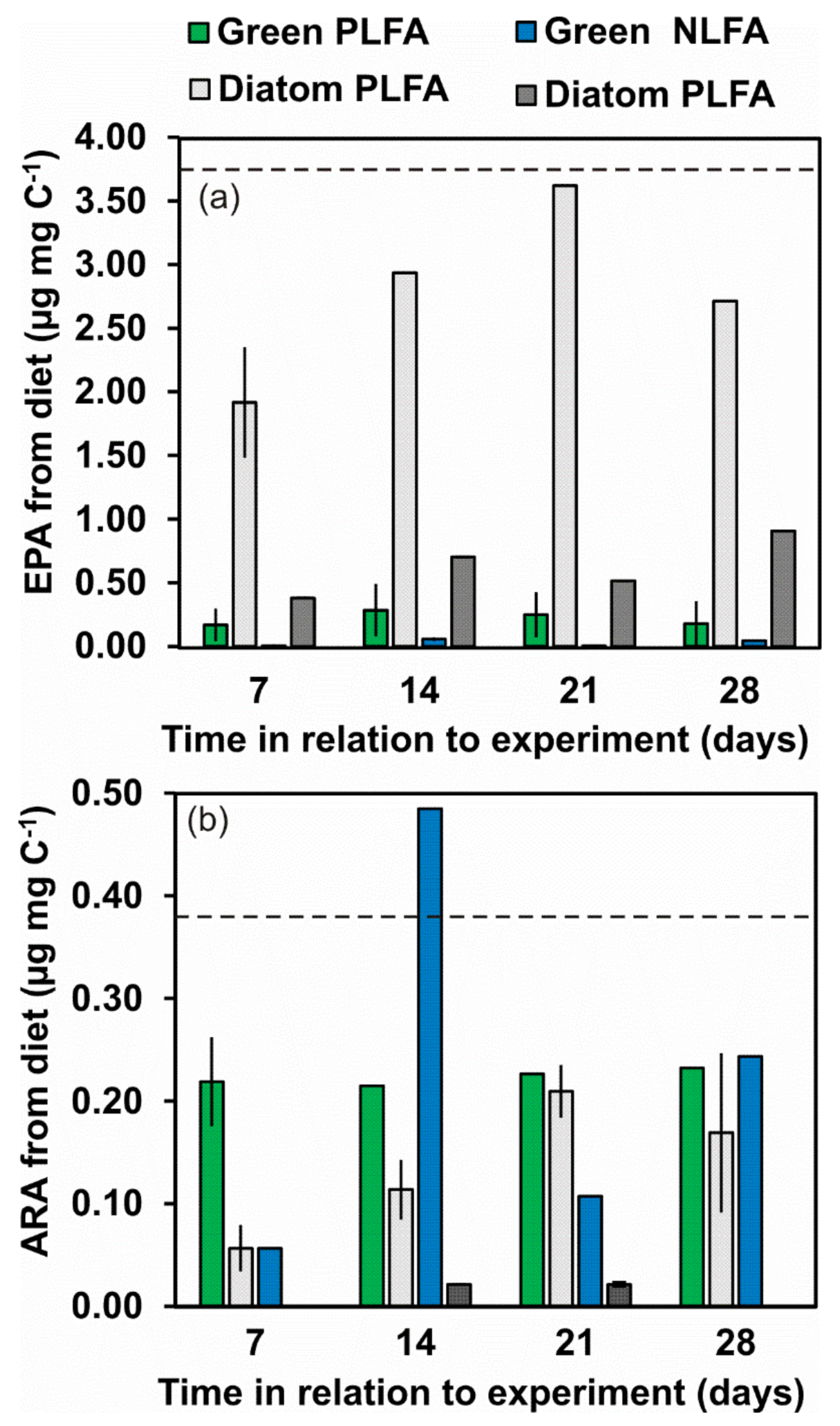
3.5. Turnover of Individual FA Based on δ13C of NLFA and PLFA
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Scott, C.L.; Falk-Petersen, S.; Gulliksen, B.; Lønne, O.; Sargent, J.R. Lipid indicators of the diet of the sympagic amphipod Gammarus wilkitzkii in the Marginal Ice Zone and in open waters of Svalbard (Arctic). Polar Biol. 2001, 24, 572–576. [Google Scholar] [CrossRef]
- Kraft, A.; Graeve, M.; Janssen, D.; Greenacre, M.; Falk-Petersen, S. Arctic pelagic amphipods: Lipid dynamics and life strategy. J. Plankton Res. 2015, 37, 790–807. [Google Scholar] [CrossRef] [Green Version]
- Dauby, P.; Nyssen, F.; De Broyer, C. Amphipods as food sources for higher trophic levels in the Southern Ocean: A synthesis. Antarctic Biology in a Global Context; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 129–134. [Google Scholar]
- Gerhardt, A.; Bloor, M.; Mills, C.L. Gammarus: Important Taxon in freshwater and marine changing environments. Int. J. Zool. 2011. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.W. Amphipoda: Chapter 39 of the light and smith manual: Intertidal invertebrates from central California to Oregon. In Completely Revised and Expanded; Oregon State University: Corvallis, OR, USA, 2007. [Google Scholar]
- Segerstråle, S.G. The immigration of glacial relicts into Northern Europe in the light of recent geological research. Fenn. Int. J. Geogr. 1982, 160, 303–312. [Google Scholar]
- Hill, C. Life cycle and spatial distribution of the amphipod Pallasea quadrispinosa in a lake in northern Sweden. Ecography 1988, 11, 298–304. [Google Scholar] [CrossRef]
- Kołodziejczyk, A.; Niedomagała, W. Relict invertebrates in lacustrine habitats: Life history of Pallaseopsis quadrispinosa (Sars, 1867)(Amphipoda: Gammaridae) in a deep mesotrophic lake. Pol. J. Ecol. 2009, 57, 729–740. [Google Scholar]
- Hadley, N.F. Adaptive Role of Lipids in Biological Systems; Wiley and Sons: Hoboken, NJ, USA, 1985. [Google Scholar]
- Cavaletto, J.F.; Gardner, W.S. Seasonal dynamics of lipids in freshwater benthic invertebrates. In Lipids in Freshwater Ecosystems; Springer: Berlin/Heidelberg, Germany, 1999; pp. 109–131. [Google Scholar]
- Clarke, A.; Skadsheim, A.; Holmes, L.J. Lipid biochemistry and reproductive biology in two species of Gammaridae (Crustacea: Amphipoda). Mar. Biol. 1985, 88, 247–263. [Google Scholar] [CrossRef]
- Lee, R.F.; Hagen, W.; Kattner, G. Lipid storage in marine zooplankton. Mar. Ecol. Prog. Ser. 2006, 307, 273–306. [Google Scholar] [CrossRef]
- Hiltunen, M.; Strandberg, U.; Keinänen, M.; Taipale, S.; Kankaala, P. Distinctive lipid composition of the copepod Limnocalanus macrurus with a high abundance of polyunsaturated fatty acids. Lipids 2014, 49, 919–932. [Google Scholar] [CrossRef]
- Napolitano, G.E.; Ackman, R.G. Lipids and hydrocarbons in Corophium volutator from Minas Basin, Nova Scotia. Mar. Biol. 1989, 100, 333–338. [Google Scholar] [CrossRef]
- Graeve, M.; Dauby, P.; Scailteur, Y. Combined lipid, fatty acid and digestive tract content analyses: A penetrating approach to estimate feeding modes of Antarctic amphipods. Polar Biol. 2001, 24, 853–862. [Google Scholar]
- Kattner, G.; Hagen, W.; Lee, R.F.; Campbell, R.; Deibel, D.; Falk-Petersen, S.; Graeve, M.; Hansen, B.W.; Hirche, H.; Jónasdóttir, S.H. Perspectives on marine zooplankton lipids. Can. J. Fish Aquat. Sci. 2007, 64, 1628–1639. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, N.L. Adaptations to temperature in the cellular membranes of crustacea: Membrane structure and metabolism. J. Biol. 1990, 15, 1–8. [Google Scholar] [CrossRef]
- Brett, M.T.; Müller-Navarra, D.C.; Persson, J. Crustacean zooplankton fatty acid composition. In Lipids in Aquatic Ecosystems; Springer: Berlin/Heidelberg, Germany, 2009; pp. 115–146. [Google Scholar]
- Taipale, S.J.; Kahilainen, K.K.; Holtgrieve, G.W.; Peltomaa, E.T. Simulated eutrophication and browning alters zooplankton nutritional quality and determines juvenile fish growth and survival. Ecol. Evol. 2018, 8, 2671–2687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, J.G.; Sargent, J.R. Arachidonic acid in aquaculture feeds: Current status and future opportunities. Aquaculture 2003, 218, 491–499. [Google Scholar] [CrossRef]
- Kainz, M.; Arts, M.T.; Mazumder, A. Essential fatty acids in the planktonic food web and their ecological role for higher trophic levels. Limnol. Oceanogr. 2004, 49, 1784–1793. [Google Scholar] [CrossRef]
- Strandberg, U.; Hiltunen, M.; Jelkänen, E.; Taipale, S.J.; Kainz, M.J.; Brett, M.T.; Kankaala, P. Selective transfer of polyunsaturated fatty acids from phytoplankton to planktivorous fish in large boreal lakes. Sci. Total Environ. 2015, 536, 858–865. [Google Scholar] [CrossRef]
- Arts, M.T.; Brett, M.T.; Kainz, M. Lipids in Aquatic Ecosystems; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Goedkoop, W.; Sonesten, L.; Ahlgren, G.; Boberg, M. Fatty acids in profundal benthic invertebrates and their major food resources in Lake Erken, Sweden: Seasonal variation and trophic indications. Can. J. Fish Aquat. Sci. 2000, 57, 2267–2279. [Google Scholar] [CrossRef]
- Lau, D.C.; Vrede, T.; Pickova, J.; Goedkoop, W. Fatty acid composition of consumers in boreal lakes–variation across species, space and time. Freshw. Biol. 2012, 57, 24–38. [Google Scholar] [CrossRef]
- Maazouzi, C.; Masson, G.; Izquierdo, M.S.; Pihan, J. Fatty acid composition of the amphipod Dikerogammarus villosus: Feeding strategies and trophic links. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 868–875. [Google Scholar] [CrossRef]
- Nelson, M.M.; Mooney, B.D.; Nichols, P.D.; Phleger, C.F. Lipids of Antarctic Ocean amphipods: Food chain interactions and the occurrence of novel biomarkers. Mar. Chem. 2001, 73, 53–64. [Google Scholar] [CrossRef]
- Auel, H.; Harjes, M.; Da Rocha, R.; Stübing, D.; Hagen, W. Lipid biomarkers indicate different ecological niches and trophic relationships of the Arctic hyperiid amphipods Themisto abyssorum and T. libellula. Polar Biol. 2002, 25, 374–383. [Google Scholar] [CrossRef]
- Baeza-Rojano, E.; Hachero-Cruzado, I.; Guerra-García, J.M. Nutritional analysis of freshwater and marine amphipods from the Strait of Gibraltar and potential aquaculture applications. J. Sea Res. 2014, 85, 29–36. [Google Scholar] [CrossRef]
- Koussoroplis, A.; Lemarchand, C.; Bec, A.; Desvilettes, C.; Amblard, C.; Fournier, C.; Berny, P.; Bourdier, G. From aquatic to terrestrial food webs: Decrease of the docosahexaenoic acid/linoleic acid ratio. Lipids 2008, 43, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Hixson, S.M.; Sharma, B.; Kainz, M.J.; Wacker, A.; Arts, M.T. Production, distribution, and abundance of long-chain omega-3 polyunsaturated fatty acids: A fundamental dichotomy between freshwater and terrestrial ecosystems. Environ. Rev. 2015, 23, 414–424. [Google Scholar] [CrossRef]
- Del Rio, C.M.; McWilliams, S.R. How essential fats affect bird performance and link aquatic ecosystems and terrestrial consumers. Proc. Natl. Acad. Sci. USA 2016, 113, 11988–11990. [Google Scholar] [CrossRef] [Green Version]
- Twining, C.W.; Brenna, J.T.; Hairston, N.G., Jr.; Flecker, A.S. Highly unsaturated fatty acids in nature: What we know and what we need to learn. Oikos 2016, 125, 749–760. [Google Scholar] [CrossRef]
- Twining, C.W.; Brenna, J.T.; Lawrence, P.; Shipley, J.R.; Tollefson, T.N.; Winkler, D.W. Omega-3 long-chain polyunsaturated fatty acids support aerial insectivore performance more than food quantity. Proc. Natl. Acad. Sci. USA 2016, 113, 10920–10925. [Google Scholar] [CrossRef] [Green Version]
- Monroig, Ó.; Kabeya, N. Desaturases and elongases involved in polyunsaturated fatty acid biosynthesis in aquatic invertebrates: A comprehensive review. Fish. Sci. 2018, 84, 911–928. [Google Scholar] [CrossRef] [Green Version]
- Taipale, S.J.; Vuorio, K.; Strandberg, U.; Kahilainen, K.K.; Järvinen, M.; Hiltunen, M.; Peltomaa, E.; Kankaala, P. Lake eutrophication and brownification downgrade availability and transfer of essential fatty acids for human consumption. Environ. Int. 2016, 96, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Taipale, S.J.; Hiltunen, M.; Vuorio, K.; Peltomaa, E. Suitability of phytosterols alongside fatty acids as chemotaxonomic biomarkers for phytoplankton. Front. Plant Sci. 2016, 7, 212. [Google Scholar] [CrossRef]
- Peltomaa, E.T.; Aalto, S.L.; Vuorio, K.M.; Taipale, S.J. The importance of phytoplankton biomolecule availability for secondary production. Front. Ecol. Evol. 2017, 5, 128. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Quigley, M.A.; Cavaletto, J.F.; Gordon, W. Seasonal changes in lipid content and composition in the benthic amphipods Monoporeia affinis and Pontoporeia femorata. Limnol. Oceanogr. 1992, 37, 1280–1289. [Google Scholar] [CrossRef]
- Kainz, M.J.; Johannsson, O.E.; Arts, M.T. Diet effects on lipid composition, somatic growth potential, and survival of the benthic amphipod Diporeia spp. J. Great Lakes Res. 2010, 36, 351–356. [Google Scholar] [CrossRef]
- Twining, C.W.; Taipale, S.J.; Ruess, L.; Bec, A.; Martin-Creuzburg, D.; Kainz, M.J. Stable isotopes of fatty acids: Current and future perspectives for advancing trophic ecology. Philos. Trans. R. Soc. B 2020, 375, 20190641. [Google Scholar] [CrossRef] [PubMed]
- Bell, M.V.; Dick, J.R.; Anderson, T.R.; Pond, D.W. Application of liposome and stable isotope tracer techniques to study polyunsaturated fatty acid biosynthesis in marine zooplankton. J. Plankton Res. 2007, 29, 417–422. [Google Scholar] [CrossRef]
- De Troch, M.; Boeckx, P.; Cnudde, C.; Van Gansbeke, D.; Vanreusel, A.; Vincx, M.; Caramujo, M.J. Bioconversion of fatty acids at the basis of marine food webs: Insights from a compound-specific stable isotope analysis. Mar. Ecol. Prog. Ser. 2012, 465, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Taipale, S.J.; Kainz, M.J.; Brett, M.T. Diet-switching experiments show rapid accumulation and preferential retention of highly unsaturated fatty acids in Daphnia. Oikos 2011, 120, 1674–1682. [Google Scholar] [CrossRef]
- Taipale, S.J.; Peltomaa, E.; Hiltunen, M.; Jones, R.I.; Hahn, M.W.; Biasi, C.; Brett, M.T. Inferring phytoplankton, terrestrial plant and bacteria bulk δ¹³C values from compound specific analyses of lipids and fatty acids. PLoS ONE 2015, 10, e0133974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galloway, A.W.; Budge, S.M. The Critical Importance of Experimentation in Biomarker-based Trophic Ecology; Royal Society Publishing: London, UK, 2020; Volume 375. [Google Scholar]
- Guillard, R.R.; Lorenzen, C.J. Yellow-green algae with chlorophyllide C 1, 2. J. Phycol. 1972, 8, 10–14. [Google Scholar] [CrossRef]
- Moore, D.W.; Farrar, J.D. Effect of growth on reproduction in the freshwater amphipod, Hyalella azteca (Saussure). Hydrobiologia 1996, 328, 127–134. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Taipale, S.; Kankaala, P.; Hämäläinen, H.; Jones, R.I. Seasonal shifts in the diet of lake zooplankton revealed by phospholipid fatty acid analysis. Freshw. Biol. 2009, 54, 90–104. [Google Scholar] [CrossRef]
- Bec, A.; Perga, M.; Koussoroplis, A.; Bardoux, G.; Desvilettes, C.; Bourdier, G.; Mariotti, A. Assessing the reliability of fatty acid–specific stable isotope analysis for trophic studies. Methods Ecol. Evol. 2011, 2, 651–659. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting started with PRIMER v7. Primer-E Plymouth Plymouth Mar. Lab. 2015, 20. [Google Scholar]
- Clarke, K.R.; Warwick, R.M.; Brown, B.E. An index showing breakdown of seriation, related to disturbance, in a coral-reef assemblage. Mar. Ecol. Prog. Ser. 1993, 102, 153–160. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics 2006, 62, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.L.; Gregg, J.W. Uncertainty in source partitioning using stable isotopes. Oecologia 2001, 127, 171–179. [Google Scholar] [CrossRef]
- Valentine, R.C.; Valentine, D.L. Omega-3 Fatty Acids and the DHA Principle; CRC press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Kolanowski, W.; Stolyhwo, A.; Grabowski, M. Fatty acid composition of selected fresh water gammarids (Amphipoda, Crustacea): A potentially innovative source of omega-3 LC PUFA. J. Am. Oil Chem. Soc. 2007, 84, 827–833. [Google Scholar] [CrossRef]
- Salonen, J.K.; Hiltunen, M.; Figueiredo, K.; Paavilainen, P.; Sinisalo, T.; Strandberg, U.; Kankaala, P.; Taskinen, J. Population structure, life cycle, and trophic niche of the glacial relict amphipod, Gammaracanthus lacustris, in a large boreal lake. Freshw. Biol. 2019, 64, 2176–2188. [Google Scholar] [CrossRef]
- Morris, R.J. The endemic faunae of Lake Baikal: Their general biochemistry and detailed lipid composition. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1984, 222, 51–78. [Google Scholar]
- Makhutova, O.N.; Sushchik, N.N.; Gladyshev, M.I.; Ageev, A.V.; Pryanichnikova, E.G.; Kalachova, G.S. Is the fatty acid composition of freshwater zoobenthic invertebrates controlled by phylogenetic or trophic factors? Lipids 2011, 46, 709–721. [Google Scholar] [CrossRef]
- Vesterinen, J.; Keva, O.; Kahilainen, K.K.; Strandberg, U.; Hiltunen, M.; Kankaala, P.; Taipale, S.J. Nutritional quality of littoral macroinvertebrates and pelagic zooplankton in subarctic lakes. Limnol. Oceanogr. 2020. [Google Scholar] [CrossRef]
- Koussoroplis, A.; Nussbaumer, J.; Arts, M.T.; Guschina, I.A.; Kainz, M.J. Famine and feast in a common freshwater calanoid: Effects of diet and temperature on fatty acid dynamics of Eudiaptomus gracilis. Limnol. Oceanogr. 2014, 59, 947–958. [Google Scholar] [CrossRef]
- Taipale, S.J.; Galloway, A.W.; Aalto, S.L.; Kahilainen, K.K.; Strandberg, U.; Kankaala, P. Terrestrial carbohydrates support freshwater zooplankton during phytoplankton deficiency. Sci. Rep. 2016, 6, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrand, M.; Lerch, S.J.; Shrestha, R.P. Understanding diatom cell wall silicification—Moving forward. Front. Mar. Sci. 2018, 5, 125. [Google Scholar] [CrossRef] [Green Version]
- Strandberg, U.; Taipale, S.J.; Kainz, M.J.; Brett, M.T. Retroconversion of docosapentaenoic acid (n-6): An alternative pathway for biosynthesis of arachidonic acid in Daphnia magna. Lipids 2014, 49, 591–595. [Google Scholar] [CrossRef]
- Nielsen, B.L.H.; Gøtterup, L.; Jørgensen, T.S.; Hansen, B.W.; Hansen, L.H.; Mortensen, J.; Jepsen, P.M. n-3 PUFA biosynthesis by the copepod Apocyclops royi documented using fatty acid profile analysis and gene expression analysis. Biol. Open 2019, 8. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Sim. (PLFA, NLFA, %) | Main FA (PLFA/NLFA) |
|---|---|
| Wild (92.1, 85.4) | EPA, 18:1ω9, 16:0, DHA / EPA, 16:1 ω7, 18:1ω9, 16:0 |
| Starved (89.7, 91.3) | EPA, 18:1ω9, DHA, 16:0 / 18:1ω9, 16:0, 18:0, EPA |
| Green (87.1, 87.3) | 18:1ω9, EPA, 16:0, ALA / 18:1ω9, 16:0, EPA |
| Diatom (86.6, 81.0) | EPA, 16:0, 18:1ω9, 18:1ω7 / 16:0, EPA, 18:1ω9, 18:1ω7 |
| Comparison | Average Dis. (%) | Main FA | Cum. % |
|---|---|---|---|
| NLFA | |||
| Wild vs. starved | 37.7 | 16:1ω7, 18:1ω9, EPA, 16:0 | 69.7 |
| Starved vs. green algae | 31.0 | ALA, EPA, 18:0, 18:1ω9 | 63.1 |
| Starved vs. diatom | 31.2 | 18:1ω9, 16:1ω7, EPA, 18:1ω7 | 59.9 |
| Green algae vs. diatom | 50.0 | ALA, 18:1ω9, EPA, 18:1ω7 | 64.4 |
| Green algae vs. Acutodesmus sp. | 12.6 | 18:1ω9, ALA, 16:0, LA | 59.9 |
| Diatom vs. Diatoma tenuis | 56.4 | 16:1ω7, 16:0, 18:1ω9 | 62.9 |
| PLFA | |||
| Wild vs. starved | 14.8 | DHA, 16:1ω7, 16:0, EPA | 63.8 |
| Starved vs. green algae | 32.1 | EPA, ALA, 18:1ω9 | 61.8 |
| Starved vs. diatom | 18.0 | DHA, EPA, 16:0, 18:1ω7 | 63.6 |
| Green algae vs. diatom | 31.5 | EPA, ALA, 18:1ω9, 18:1ω7 | 67.9 |
| Green algae vs. Acutodesmus sp. | 32.9 | EPA, 18:1ω9, DHA, ALA | 66.6 |
| Diatom vs. Diatoma tenuis | 49.9 | 16:1ω7, 16:0, 18:1ω9 | 70.3 |
| SAFA | MUFA | ω-6 PUFA | ω-3 PUFA | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | Day(s) | 14:0 | 16:0 | 18:0 | 16:1ω7 | 18:1ω7 | 18:1ω9 | LA | ARA | ALA | SDA | EPA | DHA |
| Green, PLFA | 7 | 51.2 ± 13.4 | 49.2 ± 6.5 | 4.6 ± 1.0 | 30.4 ± 3.0 | 82 ± 8.2 | >100 | >100 | 73.7 ± 14.6 1 | 92.4 ± 3.3 | 13.6 ± 3.9 | 9.2 ± 7.0 2 | 3.5 ± 1.9 |
| 14 | 45.6 ± 12.2 | 100 ± 45 | 8.5 ± 1.0 | 65.3 ± 3.6 | >100 | >100 | >100 | >100 1 | 98.3 ± 4.1 | 34.4 ± 9.2 | 16.6 ± 12.1 2 | 1.8 ± 0.2 | |
| 21 | 76.0.0 ± 8.4 | 79.0 ± 7.3 | 8.1 ± 1.1 | 67.5 ± 15 | >100 | >100 | >100 | >100 1 | 88.4 ± 10.4 | 19.4 ± 11.9 | 16.4 ± 11.6 2 | 2.9 ± 2.6 | |
| 28 | 60.9 ± 19.1 | 80.8 ± 15.1 | 12.9 ± 5.4 | 58.2 ± 12 | >100 | >100 | >100 | >100 1 | >100 | >100 | 11.2 ± 9.0 2 | 3.6 ± 3.2 | |
| Green, NLFA | 7 | 84.1 ± 57.9 | >100 | 20.3 ± 4.5 | 86.3 ± 19.1 | >100 | >100 | >100 | 73.7 ± 14.6 | >100 | 11.6 ± 4.3 | 4.7 ± 15.6 2 | nd |
| 14 | >100 | >100 | 56.8 ± 21 | >100 | >100 | >100 | >100 | >100 1 | >100 | 16.3 ± 10.8 | 76.6 ± 19.7 2 | nd | |
| 21 | >100 | >100 | 35.4 ± 14.1 | >100 | >100 | >100 | >100 | >100 1 | >100 | 16.1 ± 4.2 | 5.5 ± 15.1 2 | nd | |
| 28 | >100 | >100 | 47.4 ± 17.9 | >100 | >100 | >100 | >100 | >100 1 | >100 | 20.9 ± 1.6 | 70.1 ± 15.3 2 | nd | |
| Diatom, PLFA | 7 | >100 | 35.3 ± 6.9 | 13.1 ± 6.2 | >100 | 52.5 ± 9.4 | 64.2 ± 24.4 | >100 | 19.6 ± 7.8 1 | nd | nd | 73.9 ± 16.7 | 26.2 ± 4.0 |
| 14 | >100 | 40.5 ± 4.0 | 17.9 ± 4.0 | >100 | 56.2 ± 8.2 | 93.9 ± 31.6 | >100 | 45.2 ± 11.6 1 | nd | nd | >100 | 22.4 ± 1.5 | |
| 21 | 68.0 ± 0.4 | 27.7 ± 2.4 | 9.7 ± 1.3 | >100 | 70.6 ± 8.3 | >100 | >100 | 86.1 ± 10.6 1 | nd | nd | >100 | 46.9 ± 1.7 | |
| 28 | >100 | 58.0 ± 7.8 | 31.5 ± 9.1 | >100 | >100 | >100 | >100 | 92.2 ± 42.5 1 | nd | nd | >100 | 54.3 ± 5.5 | |
| Diatom, NLFA | 7 | >100 | 63.2 ± 10.7 | 8.3 ± 4.1 | >100 | 67.2 ± 8.7 | 90.1 ± 33.6 | 90.5 ± 11.9 | nd | nd | nd | >100 | nd |
| 14 | >100 | 63.2 ± 20.6 | 17.7 ± 7.3 | >100 | 53.3 ± 8.4 | >100 | >100 | >100 1 | nd | nd | >100 | nd | |
| 21 | >100 | 24.9 ± 2.2 | 3,2 ± 0.5 | >100 | 62.4 ± 7.4 | 49.9 ± 23.5 | >100 | 85.5 ± 7.7 1 | nd | nd | >100 | nd | |
| 28 | >100 | 69.6 ± 28 | 13.7 ± 6.3 | >100 | >100 | >100 | >100 | nd | nd | nd | >100 | nd | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taipale, S.J.; Kers, E.; Peltomaa, E.; Loehr, J.; Kainz, M.J. Selective Fatty Acid Retention and Turnover in the Freshwater Amphipod Pallaseopsis quadrispinosa. Biomolecules 2021, 11, 478. https://doi.org/10.3390/biom11030478
Taipale SJ, Kers E, Peltomaa E, Loehr J, Kainz MJ. Selective Fatty Acid Retention and Turnover in the Freshwater Amphipod Pallaseopsis quadrispinosa. Biomolecules. 2021; 11(3):478. https://doi.org/10.3390/biom11030478
Chicago/Turabian StyleTaipale, Sami J., Erwin Kers, Elina Peltomaa, John Loehr, and Martin J. Kainz. 2021. "Selective Fatty Acid Retention and Turnover in the Freshwater Amphipod Pallaseopsis quadrispinosa" Biomolecules 11, no. 3: 478. https://doi.org/10.3390/biom11030478