
β-Dystroglycan Restoration and Pathology Progression in the Dystrophic mdx Mouse: Outcome and Implication of a Clinically Oriented Study with a Novel Oral Dasatinib Formulation

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vivo Procedures
2.1.1. Animal Groups, Drug Formulation, and Treatment Schedule
2.1.2. Forelimb Grip Strength Test and Treadmill Exhaustion Test
2.1.3. Isometric Plantar Flexor Torque
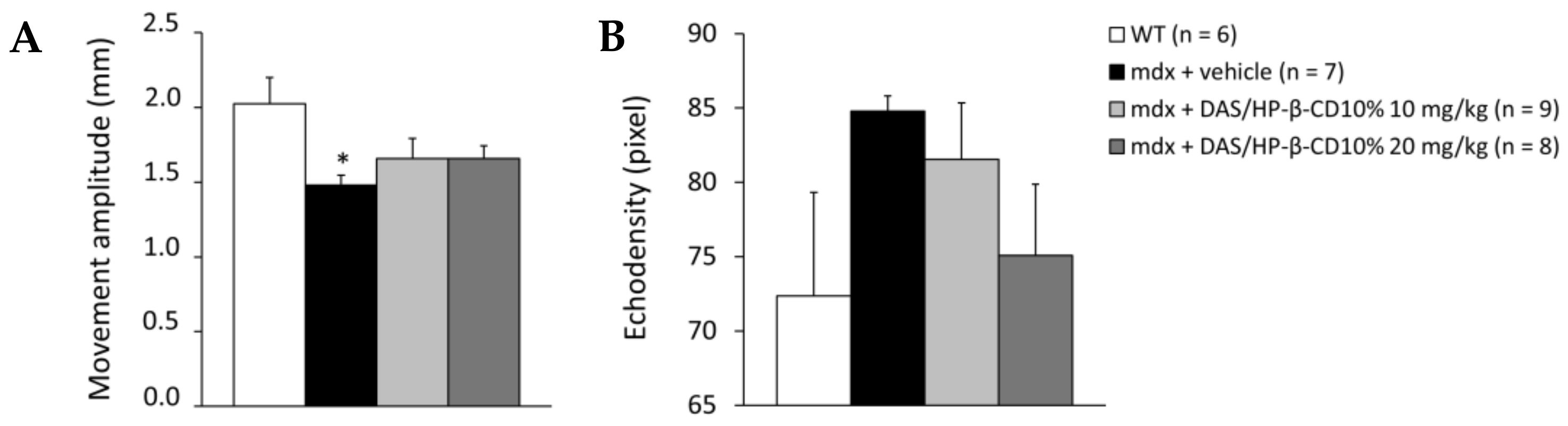
2.1.4. Diaphragm Ultrasonography
2.2. Ex Vivo Procedures
2.2.1. Sample Collection, Processing, and Storage
2.2.2. Pharmacokinetic Analysis
2.2.3. Protein Expression Analysis by Immunoprecipitation and Western Blot
2.2.4. Isometric and Eccentric Contraction Recordings of Isolated Muscles
2.2.5. Muscle Histology
2.2.6. Spectrophotometric Determination of CK and LDH Plasma Levels
2.3. Statistical Analysis
3. Results
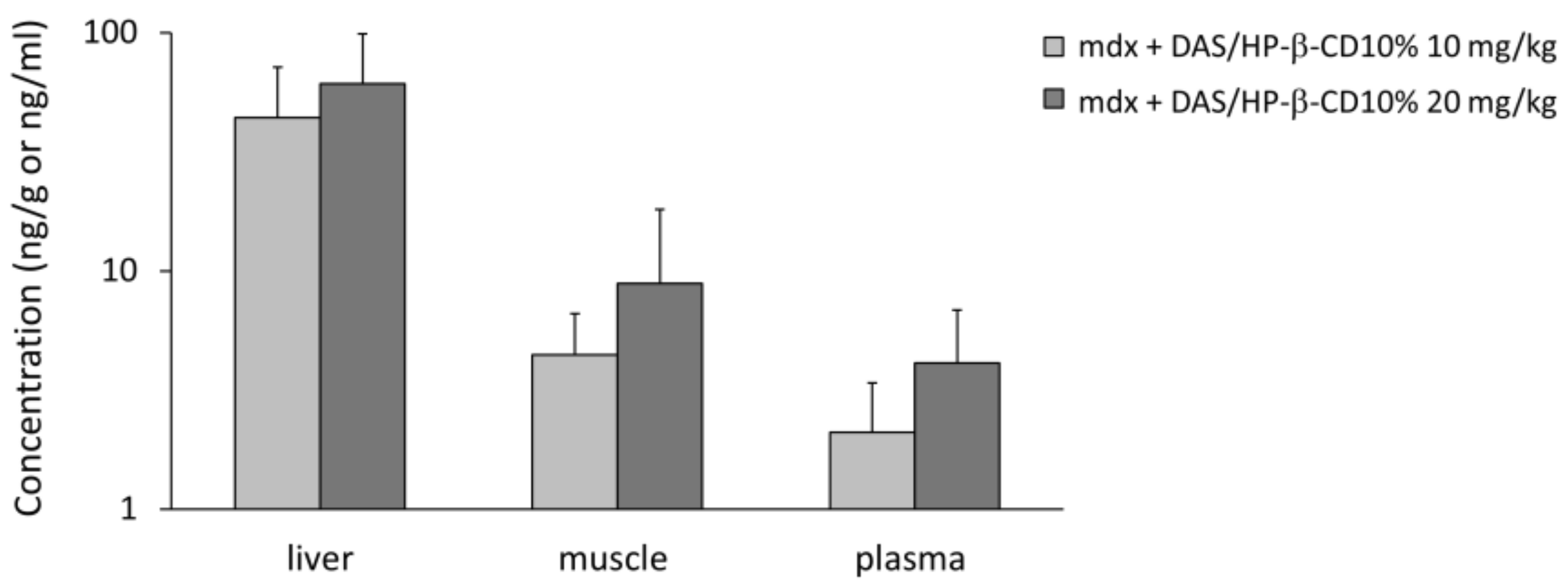
3.1. DAS/HP-β-CD 10% Oral Complex Distribution in Plasma and Tissues of mdx Mice
3.2. Effect of DAS/HP-β-CD 10% on β-DG Protein Levels in mdx DIA and GC Muscles
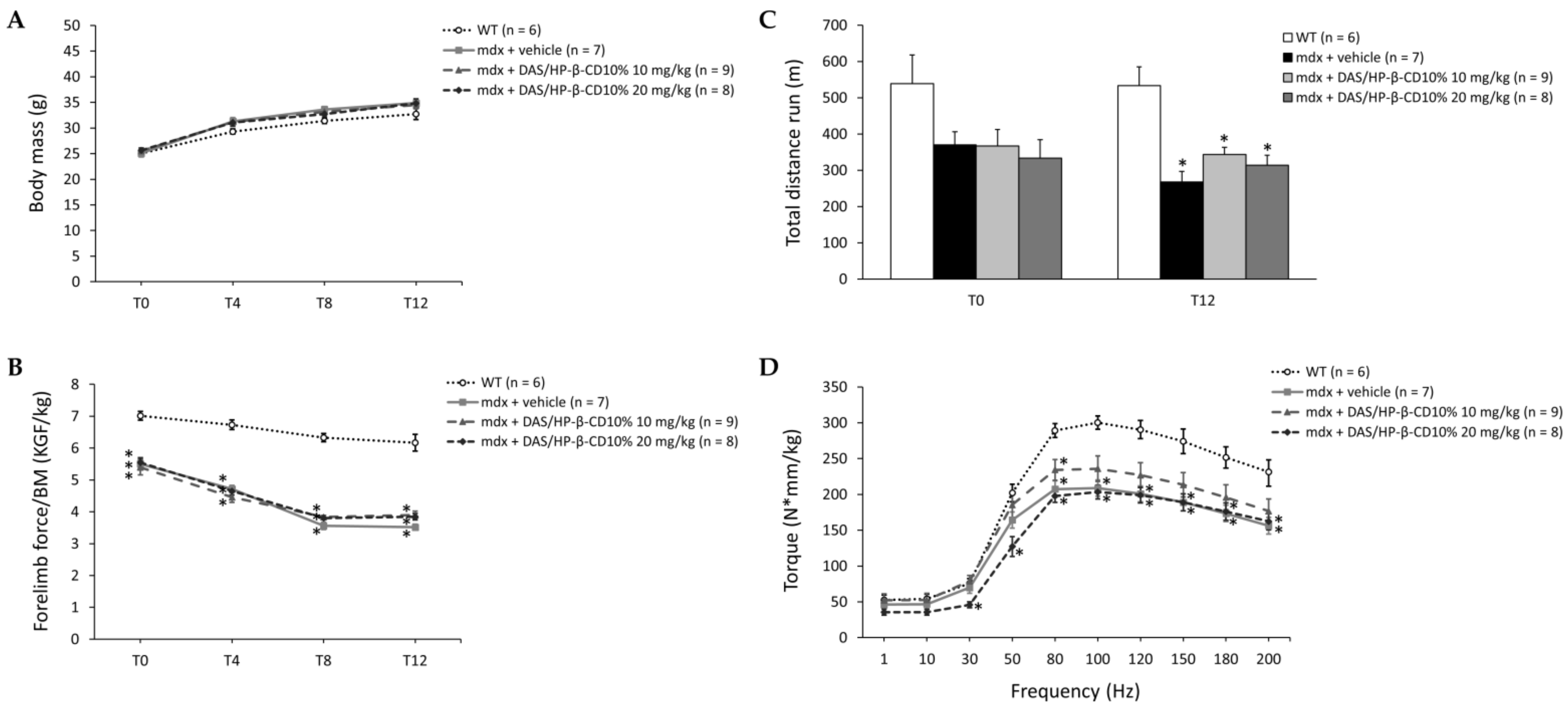
3.3. Effect of DAS/HP-β-CD 10% on In Vivo Readouts in Dystrophic Mice
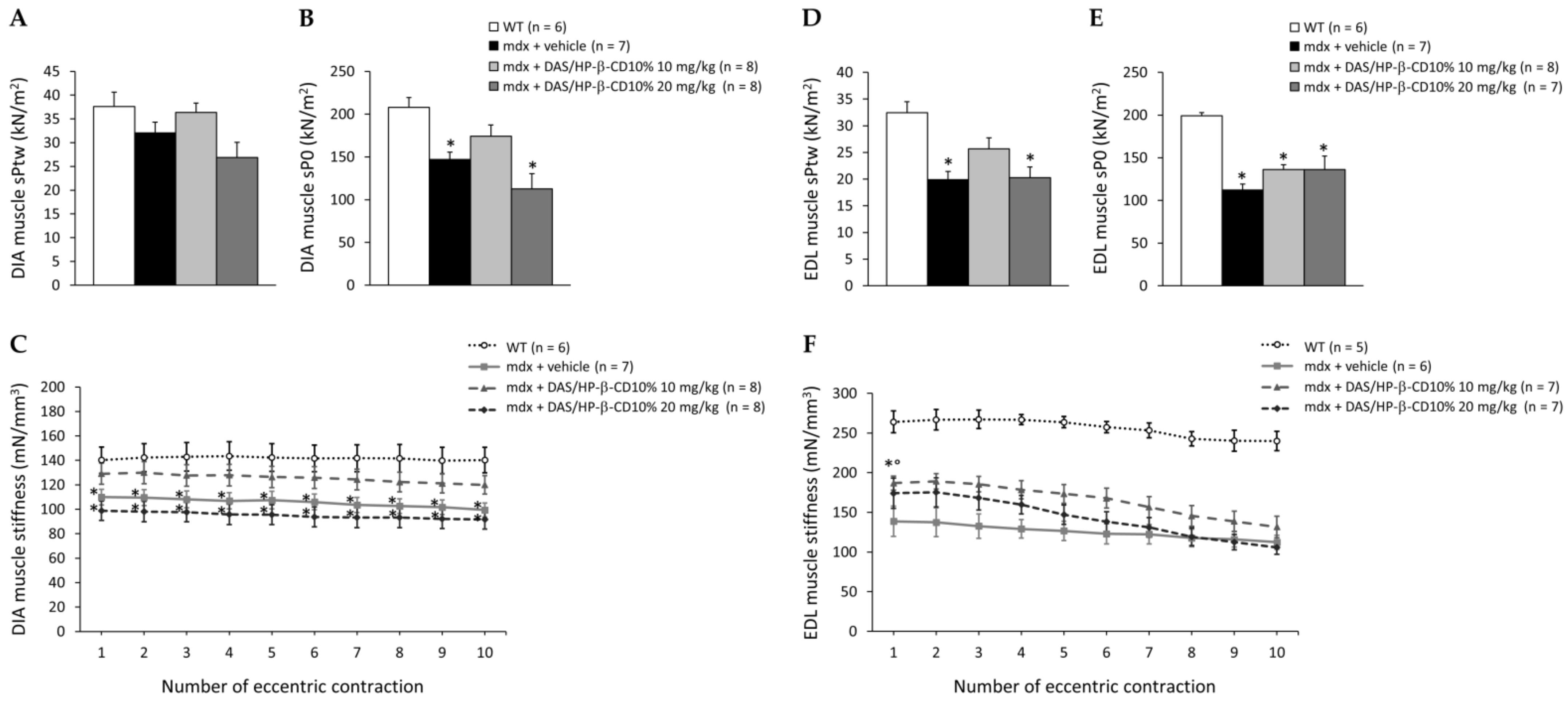
3.4. Effects of DAS/HP-β-CD 10% on Ex Vivo Contraction of mdx DIA and EDL Muscles
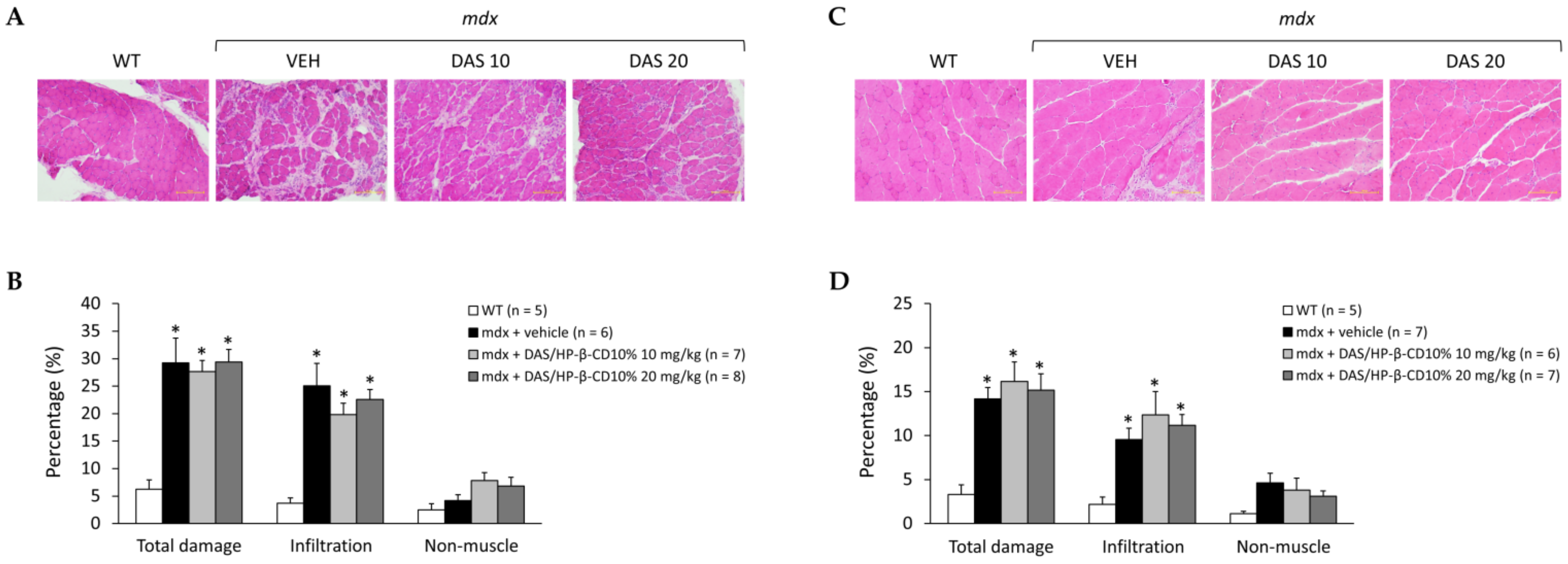
3.5. Effects of DAS/HP-β-CD 10% on Histopathology of mdx DIA and GC Muscles
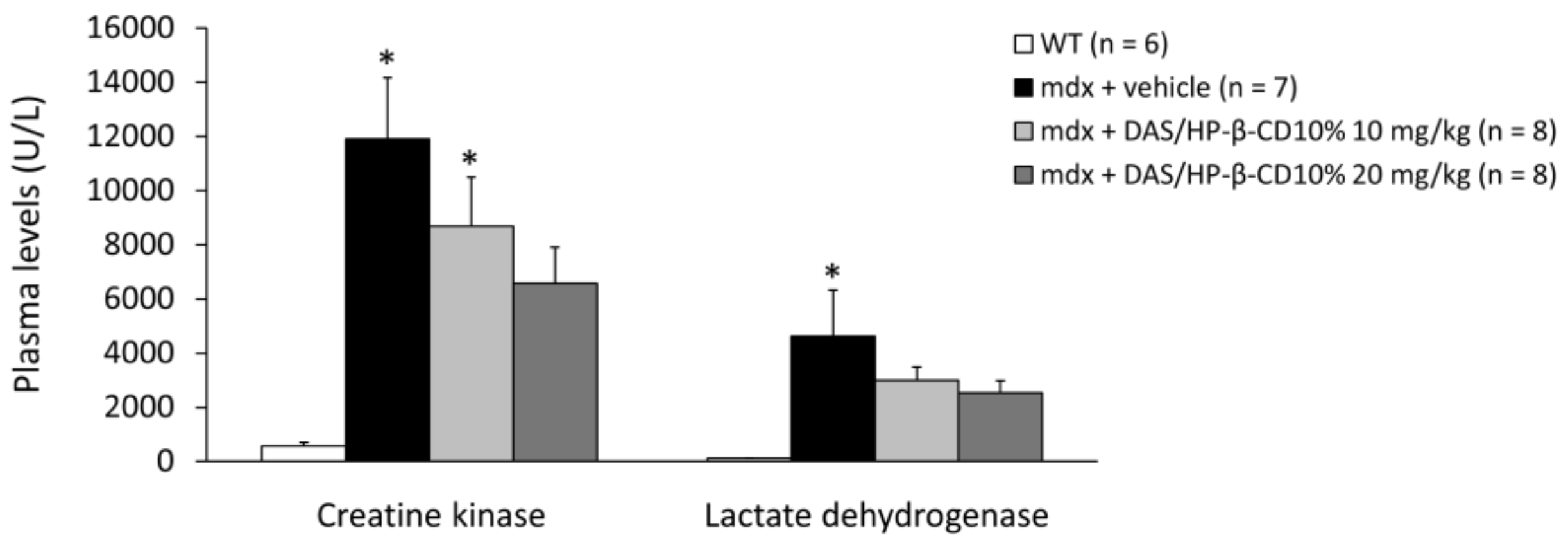
3.6. Effects of DAS/HP-β-CD 10% on Hind Limb Muscles and Vital Organs Mass, and Plasma Biomarkers of Muscle Damage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crisafulli, S.; Sultana, J.; Fontana, A.; Salvo, F.; Messina, S.; Trifirò, G. Global epidemiology of Duchenne muscular dystrophy: An updated systematic review and meta-analysis. Orphanet J. Rare Dis. 2020, 15, 141. [Google Scholar] [CrossRef]
- Duan, D.; Goemans, N.; Takeda, S.; Mercuri, E.; Aartsma-Rus, A. Duchenne muscular dystrophy. Nat. Rev. Dis. Primers 2021, 7, 13. [Google Scholar] [CrossRef]
- Birnkrant, D.J.; Bushby, K.; Bann, C.M.; Apkon, S.D.; Blackwell, A.; Brumbaugh, D.; Case, L.E.; Clemens, P.R.; Hadjiyannakis, S.; Pandya, S.; et al. DMD Care Considerations Working Group. Diagnosis and management of Duchenne muscular dystrophy, part 1: Diagnosis, and neuromuscular, rehabilitation, endocrine, and gastrointestinal and nutritional management. Lancet Neurol. 2018, 17, 251–267. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.J.; Winder, S.J. The inside and out of dystroglycan post-translational modification. Neuromuscul. Disord. 2012, 22, 959–965. [Google Scholar] [CrossRef]
- Mantuano, P.; Sanarica, F.; Conte, E.; Morgese, M.G.; Capogrosso, R.F.; Cozzoli, A.; Fonzino, A.; Quaranta, A.; Rolland, J.F.; De Bellis, M.; et al. Effect of a long-term treatment with metformin in dystrophic mdx mice: A reconsideration of its potential clinical interest in Duchenne muscular dystrophy. Biochem. Pharmacol. 2018, 154, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Kourakis, S.; Timpani, C.A.; Campelj, D.G.; Hafner, P.; Gueven, N.; Fischer, D.; Rybalka, E. Standard of care versus new-wave corticosteroids in the treatment of Duchenne muscular dystrophy: Can we do better? Orphanet J. Rare Dis. 2021, 16, 117. [Google Scholar] [CrossRef] [PubMed]
- Bouviere, J.; Fortunato, R.S.; Dupuy, C.; Werneck-de-Castro, J.P.; Carvalho, D.P.; Louzada, R.A. Exercise-Stimulated ROS Sensitive Signaling Pathways in Skeletal Muscle. Antioxidants 2021, 10, 537. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, S.; Pelosi, L.; Piemonte, F.; Travaglini, L.; Forcina, L.; Catteruccia, M.; Petrini, S.; Verardo, M.; D’Amico, A.; Musarò, A.; et al. Oxidative stress in Duchenne muscular dystrophy: Focus on the NRF2 redox pathway. Hum. Mol. Genet. 2017, 26, 2781–2790. [Google Scholar] [CrossRef]
- Wright, C.R.; Allsopp, G.L.; Addinsall, A.B.; McRae, N.L.; Andrikopoulos, S.; Stupka, N. A Reduction in Selenoprotein S Amplifies the Inflammatory Profile of Fast-Twitch Skeletal Muscle in the mdx Dystrophic Mouse. Mediat. Inflamm. 2017, 2017, 7043429. [Google Scholar] [CrossRef]
- Sadasivam, G.; Willmann, R.; Lin, S.; Erb-Vögtli, S.; Kong, X.C.; Rüegg, M.A.; Fuhrer, C. Src-family kinases stabilize the neuromuscular synapse in vivo via protein interactions, phosphorylation, and cytoskeletal linkage of acetylcholine receptors. J. Neurosci. 2005, 25, 10479–10493. [Google Scholar] [CrossRef] [Green Version]
- Giannoni, E.; Taddei, M.L.; Chiarugi, P. Src redox regulation: Again in the front line. Free Radic Biol. Med. 2010, 49, 516–527. [Google Scholar] [CrossRef]
- Camerino, G.M.; Cannone, M.; Giustino, A.; Massari, A.M.; Capogrosso, R.F.; Cozzoli, A.; De Luca, A. Gene expression in mdx mouse muscle in relation to age and exercise: Aberrant mechanical-metabolic coupling and implications for pre-clinical studies in Duchenne muscular dystrophy. Hum. Mol. Genet. 2014, 23, 5720–5732. [Google Scholar] [CrossRef] [Green Version]
- Ilsley, J.L.; Sudol, M.; Winder, S.J. The interaction of dystrophin with beta-dystroglycan is regulated by tyrosine phosphorylation. Cell Signal. 2001, 13, 625–632. [Google Scholar] [CrossRef]
- Pal, R.; Palmieri, M.; Loehr, J.A.; Li, S.; Abo-Zahrah, R.; Monroe, T.O.; Thakur, P.B.; Sardiello, M.; Rodney, G.G. Src-dependent impairment of autophagy by oxidative stress in a mouse model of Duchenne muscular dystrophy. Nat. Commun. 2014, 5, 4425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipscomb, L.; Piggott, R.W.; Emmerson, T.; Winder, S.J. Dasatinib as a treatment for Duchenne muscular dystrophy. Hum. Mol. Genet. 2016, 25, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Sanarica, F.; Mantuano, P.; Conte, E.; Cozzoli, A.; Capogrosso, R.F.; Giustino, A.; Cutrignelli, A.; Cappellari, O.; Rolland, J.F.; De Bellis, M.; et al. Proof-of-concept validation of the mechanism of action of Src tyrosine kinase inhibitors in dystrophic mdx mouse muscle: In vivo and in vitro studies. Pharmacol. Res. 2019, 145, 104260. [Google Scholar] [CrossRef]
- Cutrignelli, A.; Sanarica, F.; Lopalco, A.; Lopedota, A.; Laquintana, V.; Franco, M.; Boccanegra, B.; Mantuano, P.; De Luca, A.; Denora, N. Dasatinib/HP-β-CD Inclusion Complex Based Aqueous Formulation as a Promising Tool for the Treatment of Paediatric Neuromuscular Disorders. Int. J. Mol. Sci. 2019, 20, 591. [Google Scholar] [CrossRef] [Green Version]
- Kimmelman, J.; Mogil, J.S.; Dirnagl, U. Distinguishing between exploratory and confirmatory preclinical research will improve translation. PLoS Biol. 2014, 12, e1001863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willmann, R.; Luca, A.; Nagaraju, K.; Rüegg, M.A. Best Practices and Standard Protocols as a Tool to Enhance Translation for Neuromuscular Disorders. J. Neuromuscul. Dis. 2015, 2, 113–117. [Google Scholar] [CrossRef] [Green Version]
- Willmann, R.; Lee, J.; Turner, C.; Nagaraju, K.; Aartsma-Rus, A.; Wells, D.J.; Wagner, K.R.; Csimma, C.; Straub, V.; Grounds, M.D.; et al. Improving translatability of preclinical studies for neuromuscular disorders: Lessons from the TREAT-NMD Advisory Committee for Therapeutics (TACT). Dis. Model Mech. 2020, 13, dmm042903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Sert, N.P.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. Br. J. Pharmacol. 2020, 177, 3617–3624. [Google Scholar] [CrossRef] [PubMed]
- Capogrosso, R.F.; Mantuano, P.; Uaesoontrachoon, K.; Cozzoli, A.; Giustino, A.; Dow, T.; Srinivassane, S.; Filipovic, M.; Bell, C.; Vandermeulen, J.; et al. Ryanodine channel complex stabilizer compound S48168/ARM210 as a disease modifier in dystrophin-deficient mdx mice: Proof-of-concept study and independent validation of efficacy. FASEB J. 2018, 32, 1025–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capogrosso, R.F.; Cozzoli, A.; Mantuano, P.; Camerino, G.M.; Massari, A.M.; Sblendorio, V.T.; De Bellis, M.; Tamma, R.; Giustino, A.; Nico, B.; et al. Assessment of resveratrol, apocynin and taurine on mechanical-metabolic uncoupling and oxidative stress in a mouse model of duchenne muscular dystrophy: A comparison with the gold standard, α-methyl prednisolone. Pharmacol. Res. 2016, 106, 101–113. [Google Scholar] [CrossRef]
- Capogrosso, R.F.; Mantuano, P.; Cozzoli, A.; Sanarica, F.; Massari, A.M.; Conte, E.; Fonzino, A.; Giustino, A.; Rolland, J.F.; Quaranta, A.; et al. Contractile efficiency of dystrophic mdx mouse muscle: In vivo and ex vivo assessment of adaptation to exercise of functional end points. J. Appl. Physiol. 2017, 122, 828–843. [Google Scholar] [CrossRef]
- Mele, A.; Mantuano, P.; De Bellis, M.; Rana, F.; Sanarica, F.; Conte, E.; Morgese, M.G.; Bove, M.; Rolland, J.F.; Capogrosso, R.F.; et al. A long-term treatment with taurine prevents cardiac dysfunction in mdx mice. Transl. Res. 2019, 204, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Mele, A.; Mantuano, P.; Fonzino, A.; Rana, F.; Capogrosso, R.F.; Sanarica, F.; Rolland, J.F.; Cappellari, O.; De Luca, A. Ultrasonography validation for early alteration of diaphragm echodensity and function in the mdx mouse model of Duchenne muscular dystrophy. PLoS ONE 2021, 16, e0245397. [Google Scholar] [CrossRef]
- Mantuano, P.; Bianchini, G.; Cappellari, O.; Boccanegra, B.; Conte, E.; Sanarica, F.; Mele, A.; Camerino, G.M.; Brandolini, L.; Allegretti, M.; et al. Ergogenic Effect of BCAAs and L-Alanine Supplementation: Proof-of-Concept Study in a Murine Model of Physiological Exercise. Nutrients 2020, 12, 2295. [Google Scholar] [CrossRef]
- Whitehead, N.P.; Bible, K.L.; Kim, M.J.; Odom, G.L.; Adams, M.E.; Froehner, S.C. Validation of ultrasonography for non-invasive assessment of diaphragm function in muscular dystrophy. J. Physiol. 2016, 594, 7215–7227. [Google Scholar] [CrossRef] [Green Version]
- Cappellari, O.; Mantuano, P.; De Luca, A. “The Social Network” and Muscular Dystrophies: The Lesson Learnt about the Niche Environment as a Target for Therapeutic Strategies. Cells 2020, 9, 1659. [Google Scholar] [CrossRef]
- Duckett, D.R.; Cameron, M.D. Metabolism considerations for kinase inhibitors in cancer treatment. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1175–1193. [Google Scholar] [CrossRef] [Green Version]
- Toussaint, M.; Davidson, Z.; Bouvoie, V.; Evenepoel, N.; Haan, J.; Soudon, P. Dysphagia in Duchenne muscular dystrophy: Practical recommendations to guide management. Disabil. Rehabil. 2016, 38, 2052–2062. [Google Scholar] [CrossRef] [Green Version]
- Rao, S.; Larroque-Lombard, A.L.; Peyrard, L.; Thauvin, C.; Rachid, Z.; Williams, C.; Jean-Claude, B.J. Target modulation by a kinase inhibitor engineered to induce a tandem blockade of the epidermal growth factor receptor (EGFR) and c-Src: The concept of type III combi-targeting. PLoS ONE 2015, 10, e0117215. [Google Scholar] [CrossRef] [Green Version]
- Cullen, M.J.; Walsh, J.; Stevenson, S.A.; Rothery, S.; Severs, N.J. Co-localization of dystrophin and beta-dystroglycan demonstrated in en face view by double immunogold labeling of freeze-fractured skeletal muscle. J. Histochem. Cytochem. 1998, 46, 945–954. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vieyra, I.A.; Vásquez-Limeta, A.; González-Ramírez, R.; Morales-Lázaro, S.L.; Mondragón, M.; Mondragón, R.; Ortega, A.; Winder, S.J.; Cisneros, B. A role for β-dystroglycan in the organization and structure of the nucleus in myoblasts. Biochim. Biophys Acta 2013, 1833, 698–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, D.; Yokota, K.; Gouda, M.; Narumi, Y.; Ohmoto, H.; Nishiwaki, E.; Akita, K.; Kirii, Y. Activity-based kinase profiling of approved tyrosine kinase inhibitors. Genes Cells 2013, 18, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R., Jr. Src protein-tyrosine kinase structure, mechanism, and small molecule inhibitors. Pharm. Res. 2015, 94, 9–25. [Google Scholar] [CrossRef]
- Bouitbir, J.; Panajatovic, M.V.; Frechard, T.; Roos, N.J.; Krähenbühl, S. Imatinib and Dasatinib Provoke Mitochondrial Dysfunction Leading to Oxidative Stress in C2C12 Myotubes and Human RD Cells. Front. Pharmacol. 2020, 11, 1106. [Google Scholar] [CrossRef]
- Politano, L. Read-through approach for stop mutations in Duchenne muscular dystrophy. Update Acta Myol. 2021, 40, 43–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Centronucleated Fibers (%) | |
|---|---|---|
| DIA | GC | |
| WT (n = 5) | 1.02 ± 0.32 | 1.95 ± 0.44 |
| mdx + vehicle (n = 5) | 26.1 ± 2.7 * | 68.2 ± 3.4 * |
| mdx + DAS/HP-β-CD10% 10 mg/kg (n = 7) | 23.6 ± 4.7 * | 76.1 ± 2.7 * |
| mdx + DAS/HP-β-CD10% 20 mg/kg (n = 8) | 21.1 ± 2.9 * | 74.3 ± 3.7 * |
| Group | Mass of Hind Limb Muscles/BM (mg/g) | Mass of Vital Organs/BM (mg/g) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| TA | EDL | QUAD | GC | SOL | Liver | Heart | Kidneys | Spleen | Brain | |
| WT (n = 6) | 1.57 ± 0.07 | 0.60 ± 0.22 | 6.72 ± 0.57 | 5.34 ± 0.15 | 0.27 ± 0.02 | 42.2 ± 1.5 | 4.78 ± 0.17 | 6.71 ± 0.15 | 2.81 ± 0.10 | 13.6 ± 0.23 |
| mdx + vehicle (n = 7) | 2.30 ± 0.10 * | 0.44 ± 0.01 | 9.95 ± 0.37 * | 6.36 ± 0.11 * | 0.34 ± 0.03 | 49.8 ± 1.4 * | 4.79 ± 0.11 | 6.41 ± 0.08 | 3.03 ± 0.09 | 11.9 ± 0.67 |
| mdx + DAS/HP-β-CD10% 10 mg/kg (n = 8) | 2.27 ± 0.06 * | 0.37 ± 0.02 | 9.86 ± 0.20 * | 6.14 ± 0.11 * | 0.37 ± 0.03 | 47.7 ± 1.0 * | 4.78 ± 0.12 | 6.89 ± 0.53 | 3.13 ± 0.08 | 12.1 ± 0.32 |
| mdx + DAS/HP-β-CD10% 20 mg/kg (n = 8) | 2.23 ± 0.06 * | 0.37 ± 0.02 | 9.48 ± 0.32 * | 6.21 ± 0.16 * | 0.39 ± 0.03 | 47.8 ± 1.3 * | 5.01 ± 0.11 | 6.37 ± 0.11 | 2.97 ± 0.10 | 11.4 ± 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mantuano, P.; Boccanegra, B.; Conte, E.; De Bellis, M.; Cirmi, S.; Sanarica, F.; Cappellari, O.; Arduino, I.; Cutrignelli, A.; Lopedota, A.A.; et al. β-Dystroglycan Restoration and Pathology Progression in the Dystrophic mdx Mouse: Outcome and Implication of a Clinically Oriented Study with a Novel Oral Dasatinib Formulation. Biomolecules 2021, 11, 1742. https://doi.org/10.3390/biom11111742
Mantuano P, Boccanegra B, Conte E, De Bellis M, Cirmi S, Sanarica F, Cappellari O, Arduino I, Cutrignelli A, Lopedota AA, et al. β-Dystroglycan Restoration and Pathology Progression in the Dystrophic mdx Mouse: Outcome and Implication of a Clinically Oriented Study with a Novel Oral Dasatinib Formulation. Biomolecules. 2021; 11(11):1742. https://doi.org/10.3390/biom11111742
Chicago/Turabian StyleMantuano, Paola, Brigida Boccanegra, Elena Conte, Michela De Bellis, Santa Cirmi, Francesca Sanarica, Ornella Cappellari, Ilaria Arduino, Annalisa Cutrignelli, Angela Assunta Lopedota, and et al. 2021. "β-Dystroglycan Restoration and Pathology Progression in the Dystrophic mdx Mouse: Outcome and Implication of a Clinically Oriented Study with a Novel Oral Dasatinib Formulation" Biomolecules 11, no. 11: 1742. https://doi.org/10.3390/biom11111742