
A Novel miRNA Screen Identifies miRNA-4454 as a Candidate Biomarker for Ventricular Fibrosis in Patients with Hypertrophic Cardiomyopathy

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. miRNA Isolation
2.3. miRNA Screening Using the nCounter Sprint Digital Analyzer
2.4. miRNA Validation by RT-qPCR
2.5. Data Analysis
3. Results
3.1. Baseline Characteristics of the Study Population
3.2. Differentially Expressed of Circulating miRNAs in Patients with HCM
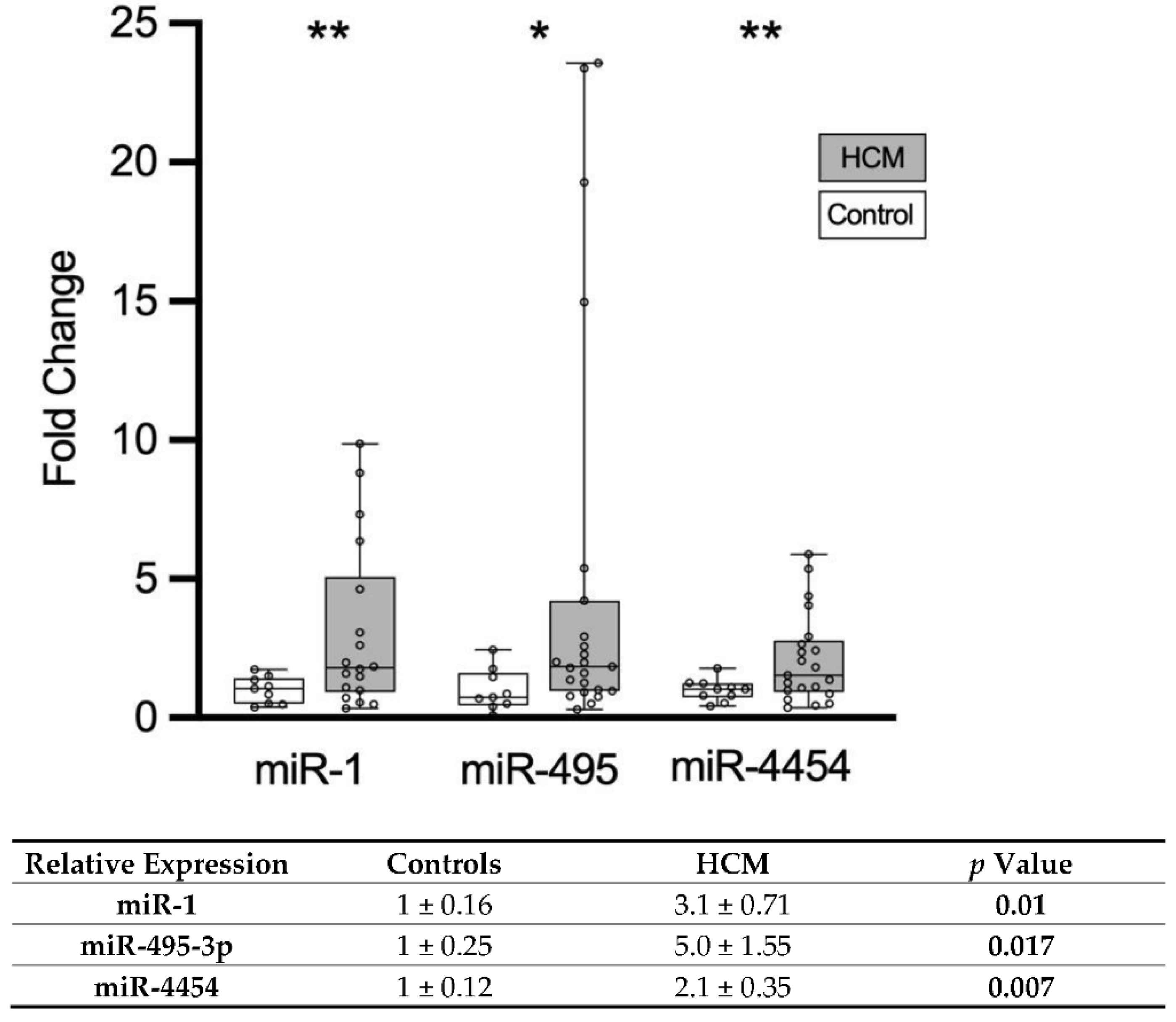
3.3. Validation of Circulating miRNAs in Patients with HCM
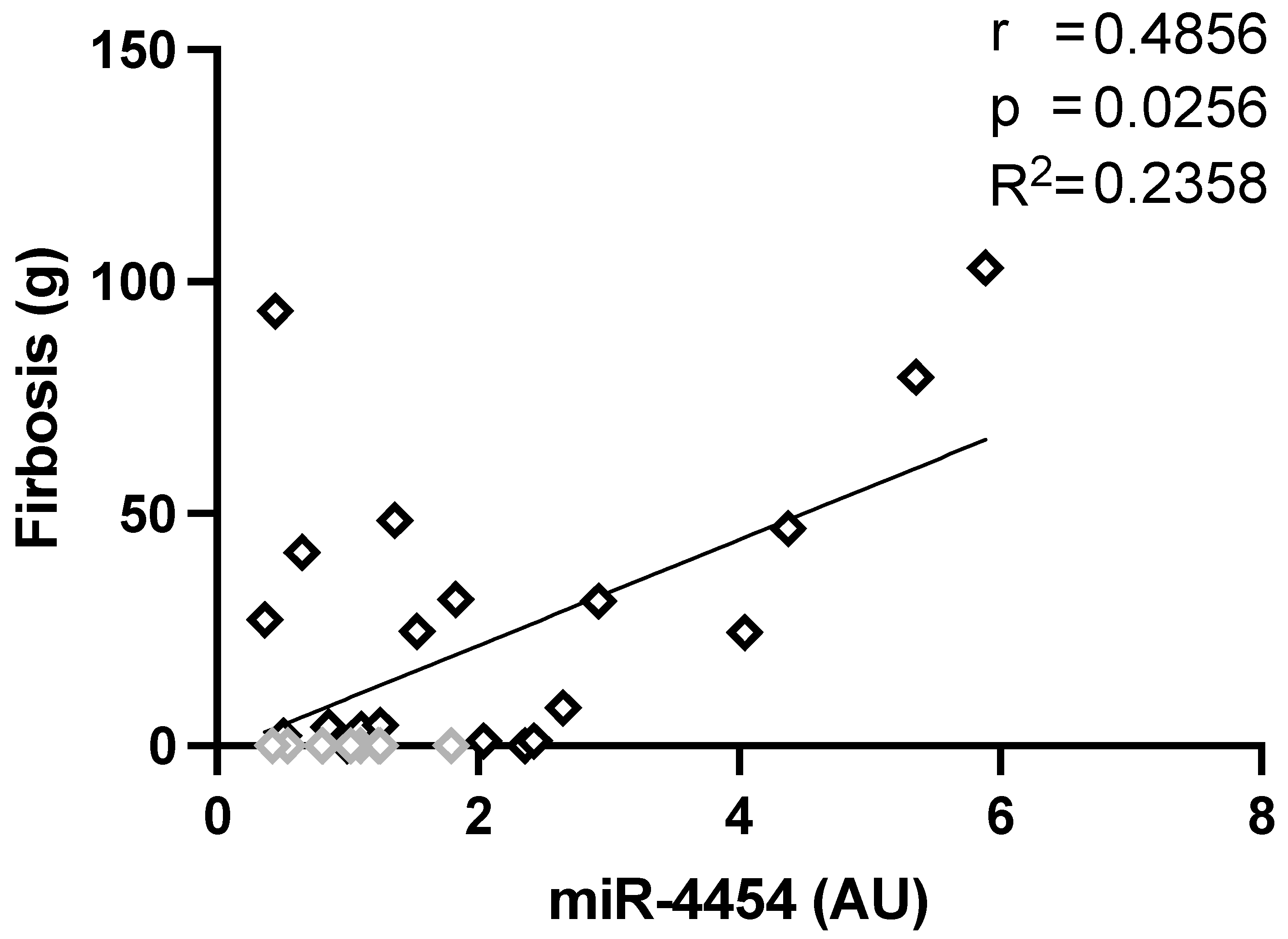
3.4. Correlation of miRNA Level and Clinical Parameters
4. Discussion
4.1. Main Findings of the Study
4.2. The nCounter Digital Analyzer
4.3. Expression Pattern of Circulating miRNA in Inherited HCM
4.4. MiR-4454 as a Putative Marker for Cardiac Fibrosis
4.5. Limitations of the Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maron, B.; Gardin, J.M.; Flack, J.M.; Gidding, S.S.; Kurosaki, T.T.; Bild, D.E. Prevalence of Hypertrophic Cardiomyopathy in a General Population of Young Adults. Circulation 1995, 92, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Semsarian, C.; Ingles, J.; Maron, M.S.; Maron, B.J. New Perspectives on the Prevalence of Hypertrophic Cardiomyopathy. J. Am. Coll. Cardiol. 2015, 65, 1249–1254. [Google Scholar] [CrossRef] [Green Version]
- Marian, A.J.; Braunwald, E. Hypertrophic cardiomyopathy: Genetics, pathogenesis, clinical manifestations, diagnosis, and therapy. Circ. Res. 2017, 121, 749–770. [Google Scholar] [CrossRef]
- Maron, B.J.; Maron, M.S. Hypertrophic cardiomyopathy. Lancet 2013, 381, 242–255. [Google Scholar] [CrossRef]
- Chim, S.S.C.; Shing, T.K.F.; Hung, E.C.W.; Leung, T.Y.; Lau, T.K.; Chiu, R.W.K.; Lo, Y.M.D. Detection and characterization of placental microRNAs in maternal plasma. Clin. Chem. 2008, 54, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilad, S.; Meiri, E.; Yogev, Y.; Benjamin, S.; Lebanony, D.; Yerushalmi, N.; Benjamin, H.; Kushnir, M.; Cholakh, H.; Melamed, N.; et al. Serum MicroRNAs Are Promising Novel Biomarkers. PLoS ONE 2008, 3, e3148. [Google Scholar] [CrossRef] [Green Version]
- Hunter, M.P.; Ismail, N.; Zhang, X.; Aguda, B.D.; Lee, E.J.; Yu, L.; Xiao, T.; Schafer, J.; Lee, M.-L.T.; Schmittgen, T.D.; et al. Detection of microRNA Expression in Human Peripheral Blood Microvesicles. PLoS ONE 2008, 3, e3694. [Google Scholar] [CrossRef] [Green Version]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Peterson, A.; Noteboom, J.; O’Briant, K.C.; Allen, A.; et al. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Natl. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Sayed, D.; Hong, C.; Chen, I.Y.; Lypowy, J.; Abdellatif, M. MicroRNAs play an essential role in the development of cardiac hypertrophy. Circ. Res. 2007, 100, 416–424. [Google Scholar] [CrossRef] [Green Version]
- Shen, E.; Diao, X.; Wei, C.; Wu, Z.; Zhang, L.; Hu, B. MicroRNAs target gene and signaling pathway by bioinformatics analysis in the cardiac hypertrophy. Biochem. Biophys. Res. Commun. 2010, 397, 380–385. [Google Scholar] [CrossRef]
- Roma-Rodrigues, C.; Raposo, L.R.; Fernandes, A.R. MicroRNAs Based Therapy of Hypertrophic Cardiomyopathy: The Road Traveled So Far. BioMed Res. Int. 2015. [Google Scholar] [CrossRef]
- Schulte, C.; Barwari, T.; Joshi, A.; Zeller, T.; Mayr, M. Noncoding RNAs versus Protein Biomarkers in Cardiovascular Disease. Trends Mol. Med. 2020, 26, 583–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scolari, F.L.; Faganello, L.S.; Garbin, H.I.; Piva e Mattos, B.; Biolo, A. A systematic review of microRNAs in patients with hypertrophic cardiomyopathy. Int. J. Cardiol. 2021, 327, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Zamorano, J.L.; Anastasakis, A.; Borger, M.A.; Borggrefe, M.; Cecchi, F.; Charron, P.; Hagege, A.A.; Lafont, A.; Limongelli, G.; Mahrholdt, H.; et al. 2014 ESC Guidelines on diagnosis and management of hypertrophic cardiomyopathy. Eur. Heart J. 2014, 35, 2733–2779. [Google Scholar] [CrossRef]
- Schulz-Menger, J.; Bluemke, D.A.; Bremerich, J.; Flamm, S.D.; Fogel, M.A.; Friedrich, M.G.; Kim, R.J.; von Knobelsdorff-Brenkenhoff, F.; Kramer, C.M.; Pennell, D.J.; et al. Standardized image interpretation and post-processing in cardiovascular magnetic resonance—2020 update. J. Cardiovasc. Magn. Reson. 2020, 22, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Geiss, G.K.; Bumgarner, R.E.; Birditt, B.; Dahl, T.; Dowidar, N.; Dunaway, D.L.; Fell, H.P.; Ferree, S.; George, R.D.; Grogan, T.; et al. Direct multiplexed measurement of gene expression with color-coded probe pairs. Nat. Biotechnol. 2008, 26, 317–325. [Google Scholar] [CrossRef]
- Bauersachs, J. Regulation of myocardial fibrosis by micrornas. J. Cardiovasc. Pharmacol. 2010, 56, 454–459. [Google Scholar] [CrossRef]
- Derda, A.A.; Thum, S.; Lorenzen, J.M.; Bavendiek, U.; Heineke, J.; Keyser, B.; Stuhrmann, M.; Givens, R.C.; Kennel, P.J.; Christian Schulze, P.; et al. Blood-based microRNA signatures differentiate various forms of cardiac hypertrophy. Int. J. Cardiol. 2015, 196, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Ellims, A.H.; Moore, X.; White, D.A.; Taylor, A.J.; Chin-Dusting, J.; Dart, A.M. Circulating microRNAs as biomarkers for diffuse myocardial fibrosis in patients with hypertrophic cardiomyopathy. J. Transl. Med. 2015, 13, 314. [Google Scholar] [CrossRef]
- Roncarati, R.; Viviani Anselmi, C.; Losi, M.A.; Papa, L.; Cavarretta, E.; Da Costa Martins, P.; Contaldi, C.; Saccani Jotti, G.; Franzone, A.; Galastri, L.; et al. Circulating miR-29a, Among Other Up-Regulated MicroRNAs, Is the Only Biomarker for Both Hypertrophy and Fibrosis in Patients With Hypertrophic Cardiomyopathy. J. Am. Coll. Cardiol. 2014, 63, 920–927. [Google Scholar] [CrossRef] [Green Version]
- Ai, J.; Zhang, R.; Li, Y.; Pu, J.; Lu, Y.; Jiao, J.; Li, K.; Yu, B.; Li, Z.; Wang, R.; et al. Circulating microRNA-1 as a potential novel biomarker for acute myocardial infarction. Biochem. Biophys. Res. Commun. 2010, 391, 73–77. [Google Scholar] [CrossRef]
- D’Alessandra, Y.; Devanna, P.; Limana, F.; Straino, S.; Di Carlo, A.; Brambilla, P.G.; Rubino, M.; Carena, M.C.; Spazzafumo, L.; De Simone, M.; et al. Circulating microRNAs are new and sensitive biomarkers of myocardial infarction. Eur. Heart J. 2010, 31, 2765–2773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, S.; Chen, Y.; He, R.; Shi, Y.; Su, L. Rescuing infusion of miRNA-1 prevents cardiac remodeling in a heart-selective miRNA deficient mouse. Biochem. Biophys. Res. Commun. 2018, 495, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, X.; Chen, L.; Chen, K.; Zhou, J.; Song, J. MiR-1-3p that correlates with left ventricular function of HCM can serve as a potential target and differentiate HCM from DCM. J. Transl. Med. 2018, 16, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palacín, M.; Reguero, J.R.; Martín, M.; Molina, B.D.; Morís, C.; Alvarez, V.; Coto, E.; Díaz Molina, B.; Morís, C.; Alvarez, V.; et al. Profile of MicroRNAs Differentially Produced in Hearts from Patients with Hypertrophic Cardiomyopathy and Sarcomeric Mutations. Clin. Chem. 2011, 57, 1614–1616. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Han, Y.; Liu, J.; Jiang, F.; Hu, H.; Wang, Y.; Liu, Q.; Gong, Y.; Li, X. MIR-135b-5p and MIR-499a-3p Promote Cell Proliferation and Migration in Atherosclerosis by Directly Targeting MEF2C. Sci. Rep. 2015, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, C.; Naya, F. The Function of the MEF2 Family of Transcription Factors in Cardiac Development, Cardiogenomics, and Direct Reprogramming. J. Cardiovasc. Dev. Dis. 2016, 3, 26. [Google Scholar] [CrossRef] [Green Version]
- Czubryt, M.P.; Olson, E.N. Balancing Contractility and Energy Production: The Role of Myocyte Enhancer Factor 2 (MEF2) in Cardiac Hypertrophy. Recent Prog. Horm. Res. 2004, 59, 105–124. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhou, Y.; Wang, C. xia LncRNA-MIAT regulates fibrosis in hypertrophic cardiomyopathy (HCM) by mediating the expression of miR-29a-3p. J. Cell. Biochem. 2019, 120, 7265–7275. [Google Scholar] [CrossRef]
- Clark, A.L.; Maruyama, S.; Sano, S.; Accorsi, A.; Girgenrath, M.; Walsh, K.; Naya, F.J. miR-410 and miR-495 Are Dynamically Regulated in Diverse Cardiomyopathies and Their Inhibition Attenuates Pathological Hypertrophy. PLoS ONE 2016, 11, e0151515. [Google Scholar] [CrossRef] [Green Version]
- Shin, V.Y.; Ng, E.K.O.; Chan, V.W.; Kwong, A.; Chu, K.-M. A three-miRNA signature as promising non-invasive diagnostic marker for gastric cancer. Mol. Cancer 2015, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Zhang, Q.; Yang, X.; Qian, S.Y.; Guo, B. Vitamin D enhances the efficacy of irinotecan through miR-627-mediated inhibition of intratumoral drug metabolism. Mol. Cancer Ther. 2016, 15, 2086–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Kong, X.; Jiang, S.; Liao, W.; Zhang, Z.; Song, J.; Liang, Y.; Zhang, W. miR-627/HMGB1/NF-κB regulatory loop modulates TGF-β1-induced pulmonary fibrosis. J. Cell. Biochem. 2019, 120, 2983–2993. [Google Scholar] [CrossRef]
- Ruan, L.; Yang, Y.; Huang, Y.; Ding, L.; Zhang, C.; Wu, X. Functional prediction of miR-3144-5p in human cardiac myocytes based on transcriptome sequencing and bioinformatics. Medicine 2017, 96, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Cai, Q.; Zhang, X.; Zhang, H.; Zhong, Y.; Xu, C.; Li, Y. Two less common human microRNAs miR-875 and miR-3144 target a conserved site of E6 oncogene in most high-risk human papillomavirus subtypes. Protein Cell 2015, 6, 575–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amarilyo, G.; Pillar, N.; Ben-Zvi, I.; Weissglas-Volkov, D.; Zalcman, J.; Harel, L.; Livneh, A.; Shomron, N. Analysis of microRNAs in familial Mediterranean fever. PLoS ONE 2018, 13, e0197829. [Google Scholar] [CrossRef]
- Nakamura, A.; Rampersaud, Y.R.; Sharma, A.; Lewis, S.J.; Wu, B.; Datta, P.; Sundararajan, K.; Endisha, H.; Rossomacha, E.; Rockel, J.S.; et al. Identification of microRNA-181a-5p and microRNA-4454 as mediators of facet cartilage degeneration. JCI Insight 2016, 1, e86820. [Google Scholar] [CrossRef]
- Liu, H.; Qin, H.; Chen, G.X.; Liang, M.Y.; Rong, J.; Yao, J.P.; Wu, Z.K. Comparative expression profiles of microRNA in left and right atrial appendages from patients with rheumatic mitral valve disease exhibiting sinus rhythm or atrial fibrillation. J. Transl. Med. 2014, 12, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Z.; Wu, H.; Zhong, W.; Zhang, Q.; Yu, Z. Expression profiling and bioinformatics analysis of circulating microRNAs in patients with acute myocardial infarction. J. Clin. Lab. Anal. 2020, 34, e23099. [Google Scholar] [CrossRef]
- Vakrou, S.; Fukunaga, R.; Foster, D.B.; Sorensen, L.; Liu, Y.; Guan, Y.; Woldemichael, K.; Pineda-Reyes, R.; Liu, T.; Tardiff, J.C.; et al. Allele-specific differences in transcriptome, miRNome, and mitochondrial function in two hypertrophic cardiomyopathy mouse models. JCI Insight 2018, 3, e94493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasari, S.; Pandhiri, T.; Grassi, T.; Visscher, D.W.; Multinu, F.; Agarwal, K.; Mariani, A.; Shridhar, V.; Mitra, A.K. Signals from the metastatic niche regulate early and advanced ovarian cancer metastasis through miR-4454 downregulation. Mol. Cancer Res. 2020, 18, 1202–1217. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, L.; Marley, R.; Miellet, S.; Hartley, P.S. The impact of SPARC on age-related cardiac dysfunction and fibrosis in Drosophila. Exp. Gerontol. 2018, 109, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, A.D.; Baicu, C.F.; Rentz, T.J.; Van Laer, A.O.; Bonnema, D.D.; Zile, M.R. Age-dependent alterations in fibrillar collagen content and myocardial diastolic function: Role of SPARC in post-synthetic procollagen processing. Am. J. Physiol. -Hear. Circ. Physiol. 2010, 298, 614–622. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| HCM | Controls | p Value | |
|---|---|---|---|
| Total (n) | 24 | 11 | |
| Age (y) | 54 ± 14 | 49 ± 9 | 0.16 |
| Sex (female) | 12 (50%) | 6 (55%) | >0.99 |
| Height (cm) | 170 ± 10 | 173 ± 12 | 0.45 |
| Weight (kg) | 80 ± 14 | 75 ± 15 | 0.43 |
| Septum thickness (mm) | 23 ± 6 | 10 ± 1 | <0.001 |
| Inferolateral wall thickness (mm) | 16 ± 4 | 9 ± 1 | <0.001 |
| Max. LVOT gradient (mmHg) | 29 ± 29 (19; 29; 29%) | 5 ± 2 | <0.001 |
| Fibrosis (g) | 26 ± 30 | 0 | <0.001 |
| Hs-cTnT (pg/mL) | 32 ± 63 | 4 ± 3 | 0.04 |
| NT-proBNP (ng/mL) | 1646 ± 2150 | 49 ± 33 | 0.002 |
| AF (n) | 4 (17%) | 0 (0%) | 0.28 |
| VT/Syncope (n) | 14 (58%) | 0 (0%) | <0.001 |
| Blood pressure (mmHg) | 126/74 | 127/79 | 0.87/0.21 |
| Heart Rate (bpm) | 69 ± 13 | 68 ± 9 | 0.80 |
| miRNA | Controls | HCM | p Value |
|---|---|---|---|
| miR-1 | 31 ± 3 | 58 ± 10 | 0.02 |
| miR-3144 | 61 ± 5 | 143 ± 30 | 0.01 |
| miR-4454 | 62 ± 8 | 123 ± 25 | 0.03 |
| miR-495-3p | 74 ± 9 | 154 ± 30 | 0.02 |
| miR-499a-5p | 75 ± 9 | 133 ± 24 | 0.04 |
| miR-627-3p | 34 ± 3 | 70 ± 14 | 0.02 |
| miR-133a | 28 ± 5 | 51 ± 13 | 0.11 |
| miR-199a-5p | 21 ± 2 | 36 ± 7 | 0.06 |
| miR-29a | 21 ± 3 | 32 ± 6 | 0.09 |
| miR-1 | miR-495-3p | miR-4454 | |
|---|---|---|---|
| Fibrosis (LGE-CMR) g | r = −0.21 p = 0.29 | r = −0.12 p = 0.51 | r = 0.56 p = 0.001 |
| LV-septal wall thickness mm | r = 0.2 p = 0.32 | r = −0.07 p = 0.71 | r = 0.38 p = 0.03 |
| LV mass g | r = 0.21 p = 0.33 | r = 0.08 p = 0.7 | r = 0.34 p = 0.09 |
| LVOT Gradient mmHg | r = −0.1 p = 0.61 | r = −0.03 p = 0.88 | r = 0.32 p = 0.08 |
| hsTNT pg/mL | r = −0.11 p = 0.61 | r = −0.07 p = 0.71 | r = 0.01 p = 0.97 |
| NT-proBNP ng/L | r = −0.13 p = 0.51 | r = −0.14 p = 0.44 | r = 0.07 p = 0.69 |
| Age years | r = 0.03 p = 0.87 | r = 0.03 p = 0.88 | r = 0.06 p = 0.77 |
| Sex m/f | r = −0.28 p = 0.16 | r = −0.07 p = 0.7 | r = 0.19 p = 0.29 |
| Height m | r = 0.28 p = 0.16 | r = −0.01 p = 0.95 | r = −0.15 p = 0.43 |
| Weight kg | r = 0.26 p = 0.19 | r = −0.13 p = 0.49 | r = 0.32 p = 0.09 |
| BSA (m/100)2 | r = 0.28 p = 0.16 | r = −0.01 p = 0.96 | r = −0.16 p = 0.4 |
| AF yes/no | r = 0.03 p = 0.88 | r = 0.07 p = 0.7 | r = −0.25 p = 0.18 |
| VT/Synkope yes/no | r = −0.02 p = 0.94 | r = −0.1 p = 0.58 | r = 0.28 p =0.12 |
| miR-495-3p | r = 0.59 p = 0.001 | – | r = 0.11 p = 0.58 |
| miR-1 | – | r = 0.59 p = 0.001 | r = 0.06 p = 0.79 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thottakara, T.; Lund, N.; Krämer, E.; Kirchhof, P.; Carrier, L.; Patten, M. A Novel miRNA Screen Identifies miRNA-4454 as a Candidate Biomarker for Ventricular Fibrosis in Patients with Hypertrophic Cardiomyopathy. Biomolecules 2021, 11, 1718. https://doi.org/10.3390/biom11111718
Thottakara T, Lund N, Krämer E, Kirchhof P, Carrier L, Patten M. A Novel miRNA Screen Identifies miRNA-4454 as a Candidate Biomarker for Ventricular Fibrosis in Patients with Hypertrophic Cardiomyopathy. Biomolecules. 2021; 11(11):1718. https://doi.org/10.3390/biom11111718
Chicago/Turabian StyleThottakara, Tilo, Natalie Lund, Elisabeth Krämer, Paulus Kirchhof, Lucie Carrier, and Monica Patten. 2021. "A Novel miRNA Screen Identifies miRNA-4454 as a Candidate Biomarker for Ventricular Fibrosis in Patients with Hypertrophic Cardiomyopathy" Biomolecules 11, no. 11: 1718. https://doi.org/10.3390/biom11111718