
Accumulation of CD28null Senescent T-Cells Is Associated with Poorer Outcomes in COVID19 Patients


Abstract
:1. Introduction
2. Negative Consequences of CD28null Cells in Aging and Underlying Conditions
2.1. Aging
2.2. Diabetes
2.3. COPD
2.4. Hypertension
2.5. CVD
2.6. Cancer
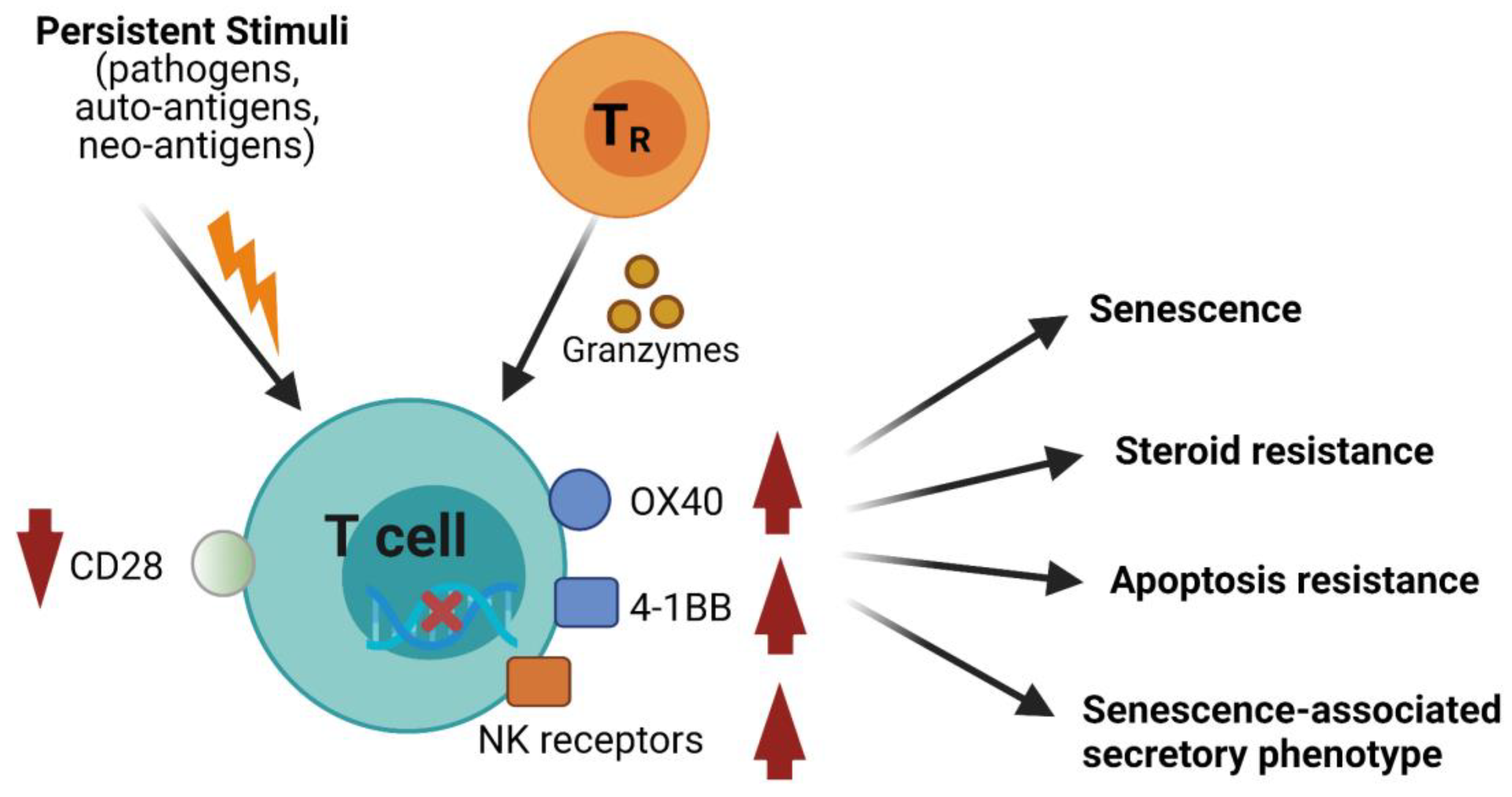
3. Mechanisms Underlying CD28null Cells-Associated Adverse Consequences
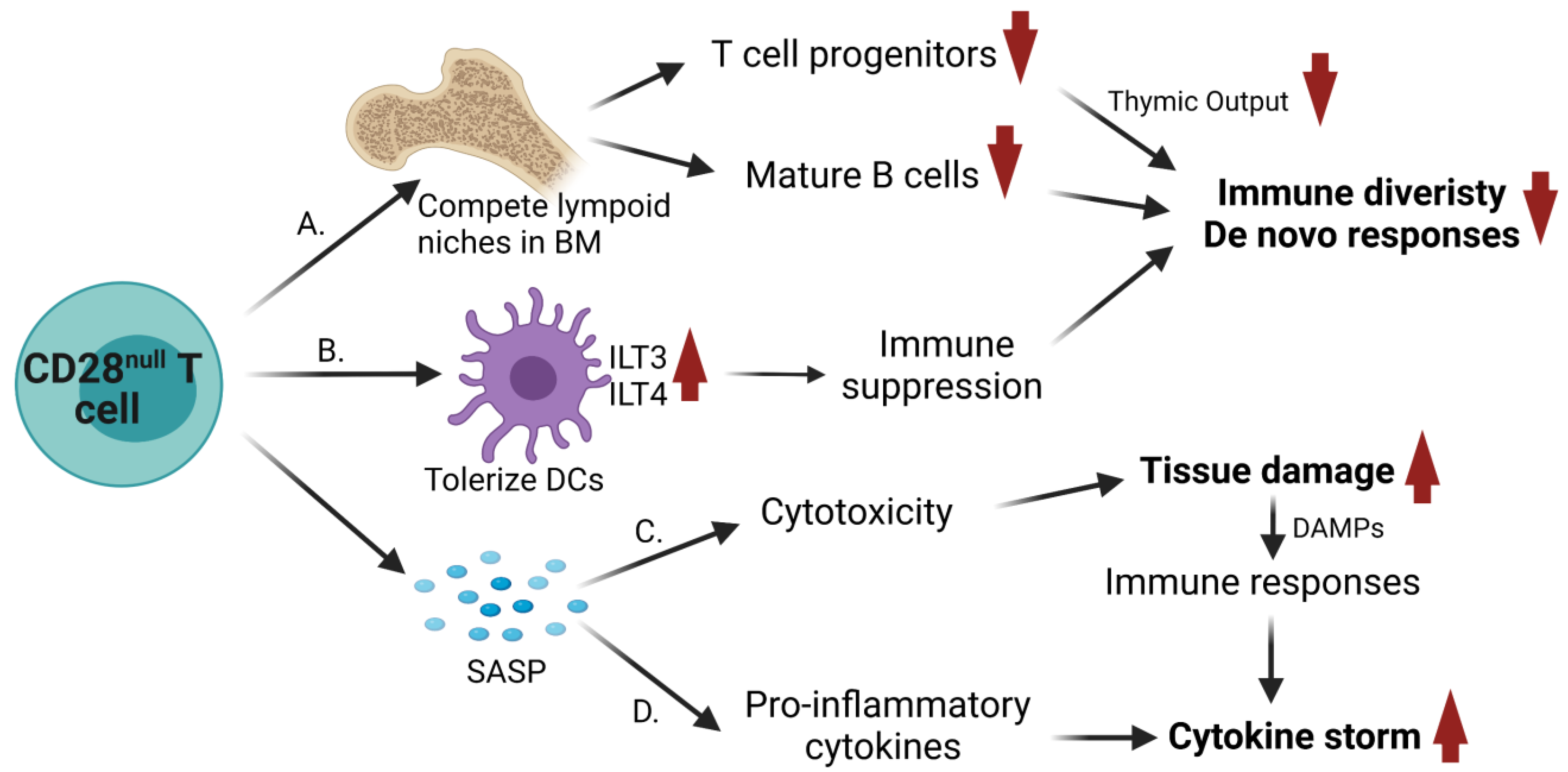
3.1. Decline of Lymphocytic Diversity and Naïve/Effector Pools
3.2. Immune Suppression
3.3. Direct Cytotoxicity
3.4. Contribution to Cytokine Release Syndrome
4. Potential Treatments
4.1. Re-sensitization to Apoptosis
4.2. Targeting the Costimulatory Pathways
4.3. Targeting the Maintenance of Senescent Cells
4.4. Preventing Senescence
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Schulte-Schrepping, J.; Reusch, N.; Paclik, D.; Baßler, K.; Schlickeiser, S.; Zhang, B.; Krämer, B.; Krammer, T.; Brumhard, S.; Bonaguro, L.; et al. Severe COVID-19 Is Marked by a Dysregulated Myeloid Cell Compartment. Cell 2020, 182, 1419–1440.e23. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Hou, H.; Yao, Y.; Wu, S.; Huang, M.; Ran, X.; Zhou, H.; Liu, Z.; Sun, Z. Systemically comparing host immunity between survived and deceased COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Liu, Y.; Yuan, J.; Wen, Y.; Xu, G.; Zhao, J.; Cheng, L.; Li, J.; Wang, X.; Wang, F.; et al. Single-cell landscape of bronchoalveolar immune cells in patients with COVID-19. Nat. Med. 2020, 26, 842–844. [Google Scholar] [CrossRef] [PubMed]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Péré, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.A.P.M.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tak-Yin Tsang, O.; et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef]
- Wu, D.; Yang, X.O. Dysregulation of Pulmonary Responses in Severe COVID-19. Viruses 2021, 13, 957. [Google Scholar] [CrossRef]
- Acuto, O.; Michel, F. CD28-mediated co-stimulation: A quantitative support for TCR signalling. Nat. Rev. Immunol. 2003, 3, 939–951. [Google Scholar] [CrossRef]
- Esensten, J.H.; Helou, Y.A.; Chopra, G.; Weiss, A.; Bluestone, J.A. CD28 Costimulation: From Mechanism to Therapy. Immunity 2016, 44, 973–988. [Google Scholar] [CrossRef] [Green Version]
- Weng, N.; Akbar, A.N.; Goronzy, J. CD28− T cells: Their role in the age-associated decline of immune function. Trends Immunol. 2009, 30, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Tarazona, R.; DelaRosa, O.; Alonso, C.; Ostos, B.; Espejo, J.; Peña, J.; Solana, R. Increased expression of NK cell markers on T lymphocytes in aging and chronic activation of the immune system reflects the accumulation of effector/senescent T cells. Mech. Ageing Dev. 2001, 121, 77–88. [Google Scholar] [CrossRef]
- Huff, W.X.; Kwon, J.H.; Henriquez, M.; Fetcko, K.; Dey, M. The Evolving Role of CD8+CD28− Immunosenescent T Cells in Cancer Immunology. Int. J. Mol. Sci. 2019, 20, 2810. [Google Scholar] [CrossRef] [Green Version]
- Dumitriu, I.E. The life (and death) of CD4 + CD28 null T cells in inflammatory diseases. Immunology 2015, 146, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Strioga, M.; Pasukoniene, V.; Characiejus, D. CD8+ CD28− and CD8+ CD57+ T cells and their role in health and disease. Immunology 2011, 134, 17–32. [Google Scholar] [CrossRef]
- Pangrazzi, L.; Weinberger, B. T cells, aging and senescence. Exp. Gerontol. 2020, 134, 110887. [Google Scholar] [CrossRef]
- Vallejo, A.N.; Schirmer, M.; Weyand, C.M.; Goronzy, J.J. Clonality and Longevity of CD4 + CD28 null T Cells Are Associated with Defects in Apoptotic Pathways. J. Immunol. 2000, 165, 6301–6307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaulding, C.; Guo, W.; Effros, R.B. Resistance to apoptosis in human CD8+ T cells that reach replicative senescence after multiple rounds of antigen-specific proliferation☆. Exp. Gerontol. 1999, 34, 633–644. [Google Scholar] [CrossRef]
- Schirmer, M.; Vallejo, A.N.; Weyand, C.M.; Goronzy, J.J. Resistance to apoptosis and elevated expression of Bcl-2 in clonally expanded CD4+CD28- T cells from rheumatoid arthritis patients. J. Immunol. 1998, 161, 1018–1025. [Google Scholar] [PubMed]
- Mikolajczyk, T.P.; Guzik, T.J. Adaptive Immunity in Hypertension. Curr. Hypertens. Rep. 2019, 21, 68. [Google Scholar] [CrossRef] [Green Version]
- Kyaw, T.; Tipping, P.; Toh, B.-H.; Bobik, A. Killer cells in atherosclerosis. Eur. J. Pharmacol. 2017, 816, 67–75. [Google Scholar] [CrossRef]
- Giubilato, S.; Liuzzo, G.; Brugaletta, S.; Pitocco, D.; Graziani, F.; Smaldone, C.; Montone, R.A.; Pazzano, V.; Pedicino, D.; Biasucci, L.M.; et al. Expansion of CD4+CD28null T-lymphocytes in diabetic patients: Exploring new pathogenetic mechanisms of increased cardiovascular risk in diabetes mellitus. Eur. Heart J. 2011, 32, 1214–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodge, G.; Hodge, S. Steroid Resistant CD8+CD28null NKT-Like Pro-inflammatory Cytotoxic Cells in Chronic Obstructive Pulmonary Disease. Front. Immunol. 2016, 7, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Hou, H.; Luo, Y.; Tang, G.; Wu, S.; Huang, M.; Liu, W.; Zhu, Y.; Lin, Q.; Mao, L.; et al. The laboratory tests and host immunity of COVID-19 patients with different severity of illness. JCI Insight 2020, 5, e137799. [Google Scholar] [CrossRef] [PubMed]
- Anft, M.; Paniskaki, K.; Blazquez-Navarro, A.; Doevelaar, A.; Seibert, F.S.; Hölzer, B.; Skrzypczyk, S.; Kohut, E.; Kurek, J.; Zapka, J.; et al. COVID-19-Induced ARDS Is Associated with Decreased Frequency of Activated Memory/Effector T Cells Expressing CD11a++. Mol. Ther. 2020, 28, 2691–2702. [Google Scholar] [CrossRef]
- Qin, C.; Zhou, L.; Hu, Z.; Zhang, S.; Yang, S.; Tao, Y.; Xie, C.; Ma, K.; Shang, K.; Wang, W.; et al. Dysregulation of Immune Response in Patients With Coronavirus 2019 (COVID-19) in Wuhan, China. Clin. Infect. Dis. 2020, 71, 762–768. [Google Scholar] [CrossRef]
- De Biasi, S.; Meschiari, M.; Gibellini, L.; Bellinazzi, C.; Borella, R.; Fidanza, L.; Gozzi, L.; Iannone, A.; Lo Tartaro, D.; Mattioli, M.; et al. Marked T cell activation, senescence, exhaustion and skewing towards TH17 in patients with COVID-19 pneumonia. Nat. Commun. 2020, 11, 3434. [Google Scholar] [CrossRef]
- Naismith, E.; Pangrazzi, L.; Grasse, M.; Keller, M.; Miggitsch, C.; Weinberger, B.; Trieb, K.; Grubeck-Loebenstein, B. Peripheral antibody concentrations are associated with highly differentiated T cells and inflammatory processes in the human bone marrow. Immun. Ageing 2019, 16, 21. [Google Scholar] [CrossRef] [Green Version]
- Panossian, L. Telomere shortening in T cells correlates with Alzheimer’s disease status. Neurobiol. Aging 2003, 24, 77–84. [Google Scholar] [CrossRef]
- Pangrazzi, L.; Meryk, A.; Naismith, E.; Koziel, R.; Lair, J.; Krismer, M.; Trieb, K.; Grubeck-Loebenstein, B. “Inflamm-aging” influences immune cell survival factors in human bone marrow. Eur. J. Immunol. 2017, 47, 481–492. [Google Scholar] [CrossRef]
- Arnold, C.R.; Pritz, T.; Brunner, S.; Knabb, C.; Salvenmoser, W.; Holzwarth, B.; Thedieck, K.; Grubeck-Loebenstein, B. T cell receptor-mediated activation is a potent inducer of macroautophagy in human CD8+CD28+ T cells but not in CD8+CD28− T cells. Exp. Gerontol. 2014, 54, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Warrington, K.J.; Takemura, S.; Goronzy, J.J.; Weyand, C.M. CD4+,CD28- T cells in rheumatoid arthritis patients combine features of the innate and adaptive immune systems. Arthritis Rheum. 2001, 44, 13–20. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Y.; Richardson, B. Decreased DNA methyltransferase levels contribute to abnormal gene expression in “senescent” CD4+CD28− T cells. Clin. Immunol. 2009, 132, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Álvarez, B.; Rodríguez, R.M.; Schlangen, K.; Raneros, A.B.; Márquez-Kisinousky, L.; Fernández, A.F.; Díaz-Corte, C.; Aransay, A.M.; López-Larrea, C. Phenotypic characteristics of aged CD4 + CD28 null T lymphocytes are determined by changes in the whole-genome DNA methylation pattern. Aging Cell 2017, 16, 293–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarde, D.N.; Lorenzo-Arteaga, K.; Corley, K.P.; Cabrera, M.; Sarvetnick, N.E. CD28− CD8+ T cells are significantly reduced and correlate with disease duration in juveniles with type 1 diabetes. Hum. Immunol. 2014, 75, 1069–1074. [Google Scholar] [CrossRef] [Green Version]
- Mikulkova, Z.; Praksova, P.; Stourac, P.; Bednarik, J.; Strajtova, L.; Pacasova, R.; Belobradkova, J.; Dite, P.; Michalek, J. Numerical defects in CD8+CD28− T-suppressor lymphocyte population in patients with type 1 diabetes mellitus and multiple sclerosis. Cell. Immunol. 2010, 262, 75–79. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, S.R.; Han, D.H.; Yu, H.T.; Han, Y.D.; Kim, J.H.; Kim, S.H.; Lee, C.J.; Min, B.-H.; Kim, D.-H.; et al. Senescent T Cells Predict the Development of Hyperglycemia in Humans. Diabetes 2019, 68, 156–162. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.-S.; Kim, S.Y.; Kim, J.T.; Lee, Y.-S.; Moon, J.S.; Kim, M.; Kang, Y.E.; Joung, K.H.; Lee, J.H.; Kim, H.J.; et al. T-cell senescence contributes to abnormal glucose homeostasis in humans and mice. Cell Death Dis. 2019, 10, 249. [Google Scholar] [CrossRef] [Green Version]
- Phoksawat, W.; Jumnainsong, A.; Leelayuwat, N.; Leelayuwat, C. Aberrant NKG2D expression with IL-17 production of CD4+ T subsets in patients with type 2 diabetes. Immunobiology 2017, 222, 944–951. [Google Scholar] [CrossRef]
- El-Samahy, M.H.; Tantawy, A.A.G.; Adly, A.A.M.; Habeeb, N.M.; Ismail, E.A.R.; Hamed, G.M.; Eladawy, R. Expression of CD4 + CD28 null T lymphocytes in children and adolescents with type 1 diabetes mellitus: Relation to microvascular complications, aortic elastic properties, and carotid intima media thickness. Pediatr. Diabetes 2017, 18, 785–793. [Google Scholar] [CrossRef]
- Hodge, G.; Mukaro, V.; Reynolds, P.N.; Hodge, S. Role of increased CD8/CD28null T cells and alternative co-stimulatory molecules in chronic obstructive pulmonary disease. Clin. Exp. Immunol. 2011, 166, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Hodge, G.; Roscioli, E.; Jersmann, H.; Tran, H.B.; Holmes, M.; Reynolds, P.N.; Hodge, S. Steroid resistance in COPD is associated with impaired molecular chaperone Hsp90 expression by pro-inflammatory lymphocytes. Respir. Res. 2016, 17, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodge, G.; Tran, H.B.; Reynolds, P.N.; Jersmann, H.; Hodge, S. Lymphocyte senescence in COPD is associated with decreased sirtuin 1 expression in steroid resistant pro-inflammatory lymphocytes. Ther. Adv. Respir. Dis. 2020, 14, 175346662090528. [Google Scholar] [CrossRef] [PubMed]
- Siena, L.; Gjomarkaj, M.; Elliot, J.; Pace, E.; Bruno, A.; Baraldo, S.; Saetta, M.; Bonsignore, M.R.; James, A. Reduced apoptosis of CD8+ T-Lymphocytes in the airways of smokers with mild/moderate COPD. Respir. Med. 2011, 105, 1491–1500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoetzenecker, K.; Mitterbauer, A.; Guenova, E.; Schweiger, T.; Altmann, P.; Zimmermann, M.; Hofbauer, H.; Beer, L.; Klepetko, W.; Ankersmit, H.J. High levels of lung resident CD4+CD28null cells in COPD: Implications of autoimmunity. Wien. Klin. Wochenschr. 2013, 125, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Lambers, C.; Hacker, S.; Posch, M.; Hoetzenecker, K.; Pollreisz, A.; Lichtenauer, M.; Klepetko, W.; Jan Ankersmit, H. T cell senescence and contraction of T cell repertoire diversity in patients with chronic obstructive pulmonary disease. Clin. Exp. Immunol. 2009, 155, 466–475. [Google Scholar] [CrossRef]
- Gackowska, L.; Michałkiewicz, J.; Niemirska, A.; Helmin-Basa, A.; Kłosowski, M.; Kubiszewska, I.; Obrycki, Ł.; Szalecki, M.; Wierzbicka, A.; Kułaga, Z.; et al. Loss of CD31 receptor in CD4+ and CD8+ T-cell subsets in children with primary hypertension is associated with hypertension severity and hypertensive target organ damage. J. Hypertens. 2018, 36, 2148–2156. [Google Scholar] [CrossRef] [PubMed]
- Youn, J.-C.; Yu, H.T.; Lim, B.J.; Koh, M.J.; Lee, J.; Chang, D.-Y.; Choi, Y.S.; Lee, S.-H.; Kang, S.-M.; Jang, Y.; et al. Immunosenescent CD8 + T Cells and C-X-C Chemokine Receptor Type 3 Chemokines Are Increased in Human Hypertension. Hypertension 2013, 62, 126–133. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Liu, Y.; Qiu, Y.; Zhang, J.; Sun, J.; Zhou, Z.; Wang, Z.; Zeng, P.; Tao, J.; He, J. Circulating senescent angiogenic T cells are linked with endothelial dysfunction and systemic inflammation in hypertension. J. Hypertens. 2021, 39, 970–978. [Google Scholar] [CrossRef]
- Yu, H.T.; Youn, J.; Kim, J.H.; Seong, Y.; Park, S.; Kim, H.C.; Lee, W.; Park, S.; Shin, E. Arterial Stiffness Is Associated With Cytomegalovirus-Specific Senescent CD8 + T Cells. J. Am. Heart Assoc. 2017, 6, e006535. [Google Scholar] [CrossRef]
- Bergström, I.; Backteman, K.; Lundberg, A.; Ernerudh, J.; Jonasson, L. Persistent accumulation of interferon-γ-producing CD8+CD56+ T cells in blood from patients with coronary artery disease. Atherosclerosis 2012, 224, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Téo, F.H.; de Oliveira, R.T.D.; Mamoni, R.L.; Ferreira, M.C.S.; Nadruz, W.; Coelho, O.R.; de Lara Fernandes, J.; Blotta, M.H.S.L. Characterization of CD4+CD28null T cells in patients with coronary artery disease and individuals with risk factors for atherosclerosis. Cell. Immunol. 2013, 281, 11–19. [Google Scholar] [CrossRef]
- Tomas, L.; Bengtsson, E.; Andersson, L.; Badn, W.; Tengryd, C.; Persson, A.; Edsfeldt, A.; Nilsson, P.M.; Schiopu, A.; Nilsson, J.; et al. Low Levels of CD4 + CD28 null T Cells at Baseline Are Associated With First-Time Coronary Events in a Prospective Population-Based Case-Control Cohort. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 426–436. [Google Scholar] [CrossRef] [Green Version]
- Sun, Z.; Ye, H.; Tang, B.; Shen, X.; Wu, X.; Zhong, H.; Song, W. Prevalence of circulating CD4+CD28null T cells is associated with early atherosclerotic damage in patients with end-stage renal disease undergoing hemodialysis. Hum. Immunol. 2013, 74, 6–13. [Google Scholar] [CrossRef]
- El-Menoufy, M.A.; El-Kak, A.E.-A.A.; Ahmed, M.A. Unusual CD4+CD28− T lymphocyte subset is implicated in the pathogenesis of early atherosclerosis in patients with rheumatoid arthritis. Egypt. Rheumatol. 2019, 41, 105–109. [Google Scholar] [CrossRef]
- Betjes, M.G.H.; Weimar, W.; Litjens, N.H.R. Circulating CD4 + CD28null T Cells May Increase the Risk of an Atherosclerotic Vascular Event Shortly after Kidney Transplantation. J. Transplant. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lopez, P.; Rodriguez-Carrio, J.; Martinez-Zapico, A.; Caminal-Montero, L.; Suarez, A. Senescent profile of angiogenic T cells from systemic lupus erythematosus patients. J. Leukoc. Biol. 2016, 99, 405–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sayed, N.M.; Abdel-Rahman, S.M.; Esmat, I.; Nammas, W. CD4+CD28null T cells in acute coronary syndrome: Lower with ST-elevation myocardial infarction. Scand. Cardiovasc. J. 2015, 49, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Koller, L.; Richter, B.; Goliasch, G.; Blum, S.; Korpak, M.; Zorn, G.; Brekalo, M.; Maurer, G.; Wojta, J.; Pacher, R.; et al. CD4+CD28null cells are an independent predictor of mortality in patients with heart failure. Atherosclerosis 2013, 230, 414–416. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, D.; Zhang, Y.; Chen, Z.; Zhu, W.; Zhang, B.; Wang, Z.; Le, H. Changes of CD4+CD25+FOXP3+ and CD8+CD28− regulatory T cells in non-small cell lung cancer patients undergoing surgery. Int. Immunopharmacol. 2014, 18, 255–261. [Google Scholar] [CrossRef]
- Meloni, F.; Morosini, M.; Solari, N.; Passadore, I.; Nascimbene, C.; Novo, M.; Ferrari, M.; Cosentino, M.; Marino, F.; Pozzi, E.; et al. Foxp3 Expressing CD4+ CD25+ and CD8+CD28− T Regulatory Cells in the Peripheral Blood of Patients with Lung Cancer and Pleural Mesothelioma. Hum. Immunol. 2006, 67, 1–12. [Google Scholar] [CrossRef]
- Song, G.; Wang, X.; Jia, J.; Yuan, Y.; Wan, F.; Zhou, X.; Yang, H.; Ren, J.; Gu, J.; Lyerly, H.K. Elevated level of peripheral CD8+CD28− T lymphocytes are an independent predictor of progression-free survival in patients with metastatic breast cancer during the course of chemotherapy. Cancer Immunol. Immunother. 2013, 62, 1123–1130. [Google Scholar] [CrossRef]
- Di, J.; Liu, M.; Fan, Y.; Gao, P.; Wang, Z.; Jiang, B.; Su, X. Phenotype molding of T cells in colorectal cancer by single-cell analysis. Int. J. Cancer 2020, 146, 2281–2295. [Google Scholar] [CrossRef] [Green Version]
- Casado, J.G.; Soto, R.; DelaRosa, O.; Peralbo, E.; del Carmen Munoz-Villanueva, M.; Rioja, L.; Peña, J.; Solana, R.; Tarazona, R. CD8 T cells expressing NK associated receptors are increased in melanoma patients and display an effector phenotype. Cancer Immunol. Immunother. 2005, 54, 1162–1171. [Google Scholar] [CrossRef]
- Arasanz, H.; Zuazo, M.; Bocanegra, A.; Gato, M.; Martínez-Aguillo, M.; Morilla, I.; Fernández, G.; Hernández, B.; López, P.; Alberdi, N.; et al. Early Detection of Hyperprogressive Disease in Non-Small Cell Lung Cancer by Monitoring of Systemic T Cell Dynamics. Cancers 2020, 12, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escarra-Senmarti, M.; Bueno-Topete, M.R.; Jave-Suarez, L.F.; Gomez-Bañuelos, E.; Gutierrez-Franco, J.; Vega-Magaña, N.; Aguilar-Lemarroy, A.; Pereira-Suarez, A.L.; Haramati, J.; del Toro-Arreola, S. Loss of CD28 within CD4+ T cell subsets from cervical cancer patients is accompanied by the acquisition of intracellular perforin, and is further enhanced by NKG2D expression. Immunol. Lett. 2017, 182, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Fabbri, E. Inflammageing: Chronic inflammation in ageing, cardiovascular disease, and frailty. Nat. Rev. Cardiol. 2018, 15, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Vallejo, A.N.; Brandes, J.C.; Weyand, C.M.; Goronzy, J.J. Modulation of CD28 expression: Distinct regulatory pathways during activation and replicative senescence. J. Immunol. 1999, 162, 6572–6579. [Google Scholar]
- Focosi, D.; Bestagno, M.; Burrone, O.; Petrini, M. CD57 + T lymphocytes and functional immune deficiency. J. Leukoc. Biol. 2010, 87, 107–116. [Google Scholar] [CrossRef]
- Broux, B.; Mizee, M.R.; Vanheusden, M.; van der Pol, S.; van Horssen, J.; Van Wijmeersch, B.; Somers, V.; de Vries, H.E.; Stinissen, P.; Hellings, N. IL-15 Amplifies the Pathogenic Properties of CD4 + CD28 − T Cells in Multiple Sclerosis. J. Immunol. 2015, 194, 2099–2109. [Google Scholar] [CrossRef] [Green Version]
- Childs, B.G.; Gluscevic, M.; Baker, D.J.; Laberge, R.-M.; Marquess, D.; Dananberg, J.; van Deursen, J.M. Senescent cells: An emerging target for diseases of ageing. Nat. Rev. Drug Discov. 2017, 16, 718–735. [Google Scholar] [CrossRef] [Green Version]
- Pal, R.; Bhadada, S.K. COVID-19 and diabetes mellitus: An unholy interaction of two pandemics. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 513–517. [Google Scholar] [CrossRef]
- Affinati, A.H.; Wallia, A.; Gianchandani, R.Y. Severe hyperglycemia and insulin resistance in patients with SARS-CoV-2 infection: A report of two cases. Clin. Diabetes Endocrinol. 2021, 7, 8. [Google Scholar] [CrossRef]
- Hodge, G.; Jersmann, H.; Tran, H.B.; Holmes, M.; Reynolds, P.N.; Hodge, S. Lymphocyte senescence in COPD is associated with loss of glucocorticoid receptor expression by pro-inflammatory/cytotoxic lymphocytes. Respir. Res. 2015, 16, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Jin, Y.; Ji, W.; Yang, H.; Chen, S.; Zhang, W.; Duan, G. Endothelial activation and dysfunction in COVID-19: From basic mechanisms to potential therapeutic approaches. Signal Transduct. Target. Ther. 2020, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- López, P.; Rodríguez-Carrio, J.; Martínez-Zapico, A.; Pérez-Álvarez, Á.I.; Suárez-Díaz, S.; Mozo, L.; Benavente, L.; Caminal-Montero, L.; Suárez, A. Low-density granulocytes and monocytes as biomarkers of cardiovascular risk in systemic lupus erythematosus. Rheumatology 2020, 59, 1752–1764. [Google Scholar] [CrossRef]
- Katsoularis, I.; Fonseca-Rodríguez, O.; Farrington, P.; Lindmark, K.; Fors Connolly, A.-M. Risk of acute myocardial infarction and ischaemic stroke following COVID-19 in Sweden: A self-controlled case series and matched cohort study. Lancet 2021, 398, 599–607. [Google Scholar] [CrossRef]
- Modin, D.; Claggett, B.; Sindet-Pedersen, C.; Lassen, M.C.H.; Skaarup, K.G.; Jensen, J.U.S.; Fralick, M.; Schou, M.; Lamberts, M.; Gerds, T.; et al. Acute COVID-19 and the Incidence of Ischemic Stroke and Acute Myocardial Infarction. Circulation 2020, 142, 2080–2082. [Google Scholar] [CrossRef]
- Reading, J.L.; Gálvez-Cancino, F.; Swanton, C.; Lladser, A.; Peggs, K.S.; Quezada, S.A. The function and dysfunction of memory CD8 + T cells in tumor immunity. Immunol. Rev. 2018, 283, 194–212. [Google Scholar] [CrossRef]
- Woroniecka, K.I.; Rhodin, K.E.; Chongsathidkiet, P.; Keith, K.A.; Fecci, P.E. T-cell Dysfunction in Glioblastoma: Applying a New Framework. Clin. Cancer Res. 2018, 24, 3792–3802. [Google Scholar] [CrossRef] [Green Version]
- Tsukishiro, T.; Donnenberg, A.D.; Whiteside, T.L. Rapid turnover of the CD8 + CD28—T-cell subset of effector cells in the circulation of patients with head and neck cancer. Cancer Immunol. Immunother. 2003, 52, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Ma, C.; Hsueh, E.C.; Dou, J.; Mo, W.; Liu, S.; Han, B.; Huang, Y.; Zhang, Y.; Varvares, M.A.; et al. TLR8 signaling enhances tumor immunity by preventing tumor-induced T-cell senescence. EMBO Mol. Med. 2014, 6, 1294–1311. [Google Scholar] [CrossRef] [PubMed]
- Fornara, O.; Odeberg, J.; Wolmer Solberg, N.; Tammik, C.; Skarman, P.; Peredo, I.; Stragliotto, G.; Rahbar, A.; Söderberg-Nauclér, C. Poor survival in glioblastoma patients is associated with early signs of immunosenescence in the CD4 T-cell compartment after surgery. Oncoimmunology 2015, 4, e1036211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert, C. A decade of immune-checkpoint inhibitors in cancer therapy. Nat. Commun. 2020, 11, 3801. [Google Scholar] [CrossRef] [PubMed]
- Darvin, P.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Immune checkpoint inhibitors: Recent progress and potential biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.; Gross, S.; Kirchberger, M.C.; Erdmann, M.; Schuler, G.; Heinzerling, L. Senescence markers: Predictive for response to checkpoint inhibitors. Int. J. Cancer 2019, 144, 1147–1150. [Google Scholar] [CrossRef]
- Falanga, A.; Marchetti, M.; Vignoli, A. Coagulation and cancer: Biological and clinical aspects. J. Thromb. Haemost. 2013, 11, 223–233. [Google Scholar] [CrossRef]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Warnock, M.; Harbaugh, A.; Yalavarthi, S.; Gockman, K.; Zuo, M.; Madison, J.A.; Knight, J.S.; Kanthi, Y.; Lawrence, D.A. Plasma tissue plasminogen activator and plasminogen activator inhibitor-1 in hospitalized COVID-19 patients. Sci. Rep. 2021, 11, 1580. [Google Scholar] [CrossRef]
- Marelli-Berg, F.M.; Fu, H.; Vianello, F.; Tokoyoda, K.; Hamann, A. Memory T-cell trafficking: New directions for busy commuters. Immunology 2010, 130, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, F.; Gebhardt, T. Bone Marrow T Cells and the Integrated Functions of Recirculating and Tissue-Resident Memory T Cells. Front. Immunol. 2016, 7, 51. [Google Scholar] [CrossRef] [Green Version]
- Plaumann, J.; Engelhardt, M.; Awwad, M.H.S.; Echchannaoui, H.; Amman, E.; Raab, M.S.; Hillengass, J.; Halama, N.; Neuber, B.; Müller-Tidow, C.; et al. IL-10 inducible CD8+ regulatory T-cells are enriched in patients with multiple myeloma and impact the generation of antigen-specific T-cells. Cancer Immunol. Immunother. 2018, 67, 1695–1707. [Google Scholar] [CrossRef] [PubMed]
- Di Rosa, F.; Santoni, A. Memory T-cell competition for bone marrow seeding. Immunology 2003, 108, 296–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goronzy, J.J.; Fulbright, J.W.; Crowson, C.S.; Poland, G.A.; O’Fallon, W.M.; Weyand, C.M. Value of Immunological Markers in Predicting Responsiveness to Influenza Vaccination in Elderly Individuals. J. Virol. 2001, 75, 12182–12187. [Google Scholar] [CrossRef] [Green Version]
- Manavalan, J.S.; Rossi, P.C.; Vlad, G.; Piazza, F.; Yarilina, A.; Cortesini, R.; Mancini, D.; Suciu-Foca, N. High expression of ILT3 and ILT4 is a general feature of tolerogenic dendritic cells. Transpl. Immunol. 2003, 11, 245–258. [Google Scholar] [CrossRef]
- Chang, C.C.; Ciubotariu, R.; Manavalan, J.S.; Yuan, J.; Colovai, A.I.; Piazza, F.; Lederman, S.; Colonna, M.; Cortesini, R.; Dalla-Favera, R.; et al. Tolerization of dendritic cells by TS cells: The crucial role of inhibitory receptors ILT3 and ILT4. Nat. Immunol. 2002, 3, 237–243. [Google Scholar] [CrossRef]
- Köstlin, N.; Ostermeir, A.-L.; Spring, B.; Schwarz, J.; Marmé, A.; Walter, C.B.; Poets, C.F.; Gille, C. HLA-G promotes myeloid-derived suppressor cell accumulation and suppressive activity during human pregnancy through engagement of the receptor ILT4. Eur. J. Immunol. 2017, 47, 374–384. [Google Scholar] [CrossRef]
- de Goeje, P.L.; Bezemer, K.; Heuvers, M.E.; Dingemans, A.-M.C.; Groen, H.J.; Smit, E.F.; Hoogsteden, H.C.; Hendriks, R.W.; Aerts, J.G.; Hegmans, J.P. Immunoglobulin-like transcript 3 is expressed by myeloid-derived suppressor cells and correlates with survival in patients with non-small cell lung cancer. Oncoimmunology 2015, 4, e1014242. [Google Scholar] [CrossRef] [Green Version]
- Gregori, S.; Tomasoni, D.; Pacciani, V.; Scirpoli, M.; Battaglia, M.; Magnani, C.F.; Hauben, E.; Roncarolo, M.-G. Differentiation of type 1 T regulatory cells (Tr1) by tolerogenic DC-10 requires the IL-10–dependent ILT4/HLA-G pathway. Blood 2010, 116, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Immunosenescence: The potential role of myeloid-derived suppressor cells (MDSC) in age-related immune deficiency. Cell. Mol. Life Sci. 2019, 76, 1901–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Huang, X.; Hsueh, E.C.; Zhang, Q.; Ma, C.; Zhang, Y.; Varvares, M.A.; Hoft, D.F.; Peng, G. Human regulatory T cells induce T-lymphocyte senescence. Blood 2012, 120, 2021–2031. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Ma, C.; Hsueh, E.C.; Eickhoff, C.S.; Zhang, Y.; Varvares, M.A.; Hoft, D.F.; Peng, G. Tumor-Derived γδ Regulatory T Cells Suppress Innate and Adaptive Immunity through the Induction of Immunosenescence. J. Immunol. 2013, 190, 2403–2414. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.; Zhang, D.; Chowdhury, D.; Martinvalet, D.; Keefe, D.; Shi, L.; Lieberman, J. Granzyme A, which causes single-stranded DNA damage, targets the double-strand break repair protein Ku70. EMBO Rep. 2006, 7, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Grossman, W.J.; Verbsky, J.W.; Barchet, W.; Colonna, M.; Atkinson, J.P.; Ley, T.J. Human T Regulatory Cells Can Use the Perforin Pathway to Cause Autologous Target Cell Death. Immunity 2004, 21, 589–601. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Mo, W.; Ye, J.; Li, L.; Zhang, Y.; Hsueh, E.C.; Hoft, D.F.; Peng, G. Regulatory T cells trigger effector T cell DNA damage and senescence caused by metabolic competition. Nat. Commun. 2018, 9, 249. [Google Scholar] [CrossRef]
- Kmiecik, J.; Poli, A.; Brons, N.H.C.; Waha, A.; Eide, G.E.; Enger, P.Ø.; Zimmer, J.; Chekenya, M. Elevated CD3+ and CD8+ tumor-infiltrating immune cells correlate with prolonged survival in glioblastoma patients despite integrated immunosuppressive mechanisms in the tumor microenvironment and at the systemic level. J. Neuroimmunol. 2013, 264, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.P.K.; Dey, M.; Neculai, D.; Cao, C.; Dever, T.E.; Sicheri, F. Structure of the Dual Enzyme Ire1 Reveals the Basis for Catalysis and Regulation in Nonconventional RNA Splicing. Cell 2008, 132, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Dumitriu, I.E.; Baruah, P.; Finlayson, C.J.; Loftus, I.M.; Antunes, R.F.; Lim, P.; Bunce, N.; Carlos Kaski, J. High Levels of Costimulatory Receptors OX40 and 4-1BB Characterize CD4 + CD28 null T Cells in Patients With Acute Coronary Syndrome. Circ. Res. 2012, 110, 857–869. [Google Scholar] [CrossRef] [Green Version]
- Verneris, M.R.; Karami, M.; Baker, J.; Jayaswal, A.; Negrin, R.S. Role of NKG2D signaling in the cytotoxicity of activated and expanded CD8+ T cells. Blood 2004, 103, 3065–3072. [Google Scholar] [CrossRef] [Green Version]
- Prajapati, K.; Perez, C.; Rojas, L.B.P.; Burke, B.; Guevara-Patino, J.A. Functions of NKG2D in CD8+ T cells: An opportunity for immunotherapy. Cell. Mol. Immunol. 2018, 15, 470–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.; Song, J.-W.; Wang, S.-Y.; Cao, W.-J.; Wang, X.-W.; Zhou, M.-J.; Yang, T.; Zhou, C.-B.; Hou, J.; Zhang, J.-Y.; et al. Changes of Damage Associated Molecular Patterns in COVID-19 Patients. Infect. Dis. Immun. 2021, 1, 20–27. [Google Scholar] [CrossRef]
- Cicco, S.; Cicco, G.; Racanelli, V.; Vacca, A. Neutrophil Extracellular Traps (NETs) and Damage-Associated Molecular Patterns (DAMPs): Two Potential Targets for COVID-19 Treatment. Mediators Inflamm. 2020, 2020, 7527953. [Google Scholar] [CrossRef]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.N.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef] [Green Version]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef]
- Wu, D.; Yang, X.O. TH17 responses in cytokine storm of COVID-19: An emerging target of JAK2 inhibitor Fedratinib. J. Microbiol. Immunol. Infect. 2020, 53, 368–370. [Google Scholar] [CrossRef]
- Hojyo, S.; Uchida, M.; Tanaka, K.; Hasebe, R.; Tanaka, Y.; Murakami, M.; Hirano, T. How COVID-19 induces cytokine storm with high mortality. Inflamm. Regen. 2020, 40, 37. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine Storm in COVID-19: The Current Evidence and Treatment Strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hao, Y.; Ou, W.; Ming, F.; Liang, G.; Qian, Y.; Cai, Q.; Dong, S.; Hu, S.; Wang, W.; et al. Serum interleukin-6 is an indicator for severity in 901 patients with SARS-CoV-2 infection: A cohort study. J. Transl. Med. 2020, 18, 406. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, P.; Zhang, L.; Wang, F. Serum Interleukin-6 Concentrations and the Severity of COVID-19 Pneumonia: A Retrospective Study at a Single Center in Bengbu City, Anhui Province, China, in January and February 2020. Med. Sci. Monit. 2020, 26, e926941. [Google Scholar] [CrossRef] [PubMed]
- Kovalcsik, E.; Antunes, R.F.; Baruah, P.; Kaski, J.C.; Dumitriu, I.E. Proteasome-Mediated Reduction in Proapoptotic Molecule Bim Renders CD4 + CD28 null T Cells Resistant to Apoptosis in Acute Coronary Syndrome. Circulation 2015, 131, 709–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugaletta, S. Novel anti-inflammatory effect of statins: Reduction of CD4+CD28null T lymphocyte frequency in patients with unstable angina. Heart 2006, 92, 249–250. [Google Scholar] [CrossRef] [Green Version]
- Link, A.; Selejan, S.; Hewera, L.; Walter, F.; Nickenig, G.; Böhm, M. Rosuvastatin induces apoptosis in CD4+CD28null T cells in patients with acute coronary syndromes. Clin. Res. Cardiol. 2011, 100, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.J.; Guan, J.; Trtchounian, A.; Hunninghake, G.M.; Kaimal, R.; Desai, M.; Kozikowski, L.-A.; DeSouza, L.; Mogan, S.; Liu, K.D.; et al. Association of Elevated Plasma Interleukin-18 Level With Increased Mortality in a Clinical Trial of Statin Treatment for Acute Respiratory Distress Syndrome. Crit. Care Med. 2019, 47, 1089–1096. [Google Scholar] [CrossRef]
- Goldstein, M.R.; Poland, G.A.; Graeber, C.W. Are certain drugs associated with enhanced mortality in COVID-19? QJM Int. J. Med. 2020, 113, 509–510. [Google Scholar] [CrossRef] [Green Version]
- Dashti-Khavidaki, S.; Khalili, H. Considerations for Statin Therapy in Patients with COVID-19. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2020, 40, 484–486. [Google Scholar] [CrossRef] [Green Version]
- Daniels, L.B.; Sitapati, A.M.; Zhang, J.; Zou, J.; Bui, Q.M.; Ren, J.; Longhurst, C.A.; Criqui, M.H.; Messer, K. Relation of Statin Use Prior to Admission to Severity and Recovery Among COVID-19 Inpatients. Am. J. Cardiol. 2020, 136, 149–155. [Google Scholar] [CrossRef]
- Chow, R.; Im, J.; Chiu, N.; Chiu, L.; Aggarwal, R.; Lee, J.; Choi, Y.-G.; Prsic, E.H.; Shin, H.J. The protective association between statins use and adverse outcomes among COVID-19 patients: A systematic review and meta-analysis. PLoS ONE 2021, 16, e0253576. [Google Scholar] [CrossRef]
- Daniels, L.B.; Ren, J.; Kumar, K.; Bui, Q.M.; Zhang, J.; Zhang, X.; Sawan, M.A.; Eisen, H.; Longhurst, C.A.; Messer, K. Relation of prior statin and anti-hypertensive use to severity of disease among patients hospitalized with COVID-19: Findings from the American Heart Association’s COVID-19 Cardiovascular Disease Registry. PLoS ONE 2021, 16, e0254635. [Google Scholar] [CrossRef]
- Herold, M.J.; McPherson, K.G.; Reichardt, H.M. Glucocorticoids in T cell apoptosis and function. Cell. Mol. Life Sci. 2006, 63, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Hodge, G.; Holmes, M.; Jersmann, H.; Reynolds, P.N.; Hodge, S. The drug efflux pump Pgp1 in pro-inflammatory lymphocytes is a target for novel treatment strategies in COPD. Respir. Res. 2013, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Kirkland, J.L.; Tchkonia, T. Senolytic drugs: From discovery to translation. J. Intern. Med. 2020, 288, 518–536. [Google Scholar] [CrossRef]
- Kirkland, J.L.; Tchkonia, T. Cellular Senescence: A Translational Perspective. EBioMedicine 2017, 21, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Justice, J.N.; Nambiar, A.M.; Tchkonia, T.; LeBrasseur, N.K.; Pascual, R.; Hashmi, S.K.; Prata, L.; Masternak, M.M.; Kritchevsky, S.B.; Musi, N.; et al. Senolytics in idiopathic pulmonary fibrosis: Results from a first-in-human, open-label, pilot study. EBioMedicine 2019, 40, 554–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickson, L.J.; Langhi Prata, L.G.P.; Bobart, S.A.; Evans, T.K.; Giorgadze, N.; Hashmi, S.K.; Herrmann, S.M.; Jensen, M.D.; Jia, Q.; Jordan, K.L.; et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine 2019, 47, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Tchkonia, T.; Pirtskhalava, T.; Gower, A.C.; Ding, H.; Giorgadze, N.; Palmer, A.K.; Ikeno, Y.; Hubbard, G.B.; Lenburg, M.; et al. The Achilles’ heel of senescent cells: From transcriptome to senolytic drugs. Aging Cell 2015, 14, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Pirtskhalava, T.; Farr, J.N.; Weigand, B.M.; Palmer, A.K.; Weivoda, M.M.; Inman, C.L.; Ogrodnik, M.B.; Hachfeld, C.M.; Fraser, D.G.; et al. Senolytics improve physical function and increase lifespan in old age. Nat. Med. 2018, 24, 1246–1256. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Ribeiro, M.L.; Gambero, A. The Impact of Polyphenols-Based Diet on the Inflammatory Profile in COVID-19 Elderly and Obese Patients. Front. Physiol. 2021, 11, 612268. [Google Scholar] [CrossRef] [PubMed]
- Topp, M.S.; Riddell, S.R.; Akatsuka, Y.; Jensen, M.C.; Blattman, J.N.; Greenberg, P.D. Restoration of CD28 Expression in CD28− CD8+ Memory Effector T Cells Reconstitutes Antigen-induced IL-2 Production. J. Exp. Med. 2003, 198, 947–955. [Google Scholar] [CrossRef] [Green Version]
- Warrington, K.J.; Vallejo, A.N.; Weyand, C.M.; Goronzy, J.J. CD28 loss in senescent CD4+ T cells: Reversal by interleukin-12 stimulation. Blood 2003, 101, 3543–3549. [Google Scholar] [CrossRef] [Green Version]
- Bryl, E.; Vallejo, A.N.; Weyand, C.M.; Goronzy, J.J. Down-Regulation of CD28 Expression by TNF-α. J. Immunol. 2001, 167, 3231–3238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryl, E.; Vallejo, A.N.; Matteson, E.L.; Witkowski, J.M.; Weyand, C.M.; Goronzy, J.J. Modulation of CD28 expression with anti-tumor necrosis factor α therapy in rheumatoid arthritis. Arthritis Rheum. 2005, 52, 2996–3003. [Google Scholar] [CrossRef] [PubMed]
- Rizzello, V.; Liuzzo, G.; Brugaletta, S.; Rebuzzi, A.; Biasucci, L.M.; Crea, F. Modulation of CD4 + CD28 null T Lymphocytes by Tumor Necrosis Factor-α Blockade in Patients With Unstable Angina. Circulation 2006, 113, 2272–2277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierer, M.; Rossol, M.; Kaltenhäuser, S.; Arnold, S.; Häntzschel, H.; Baerwald, C.; Wagner, U. Clonal expansions in selected TCR BV families of rheumatoid arthritis patients are reduced by treatment with the TNFα inhibitors etanercept and infliximab. Rheumatol. Int. 2011, 31, 1023–1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarsi, M.; Ziglioli, T.; Airo, P. Decreased Circulating CD28-negative T Cells in Patients with Rheumatoid Arthritis Treated with Abatacept Are Correlated with Clinical Response. J. Rheumatol. 2010, 37, 911–916. [Google Scholar] [CrossRef]
- Gómez-García, L.; Ramírez-Assad, C.; Vargas, A.; Massó, F.; Sánchez-Muñoz, F.; Márquez-Velasco, R.; Amezcua-Guerra, L.M.; Bojalil, R. Reduced numbers of circulating CD28-negative CD4+ cells in patients with rheumatoid arthritis chronically treated with abatacept. Int. J. Rheum. Dis. 2013, 16, 469–471. [Google Scholar] [CrossRef]
- Herndler-Brandstetter, D.; Landgraf, K.; Tzankov, A.; Jenewein, B.; Brunauer, R.; Laschober, G.T.; Parson, W.; Kloss, F.; Gassner, R.; Lepperdinger, G.; et al. The impact of aging on memory T cell phenotype and function in the human bone marrow. J. Leukoc. Biol. 2012, 91, 197–205. [Google Scholar] [CrossRef]
- Brunner, S.; Herndler-Brandstetter, D.; Arnold, C.R.; Wiegers, G.J.; Villunger, A.; Hackl, M.; Grillari, J.; Moreno-Villanueva, M.; Bürkle, A.; Grubeck-Loebenstein, B. Upregulation of miR-24 is associated with a decreased DNA damage response upon etoposide treatment in highly differentiated CD8 + T cells sensitizing them to apoptotic cell death. Aging Cell 2012, 11, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Meryk, A.; Grasse, M.; Balasco, L.; Kapferer, W.; Grubeck-Loebenstein, B.; Pangrazzi, L. Antioxidants N-Acetylcysteine and Vitamin C Improve T Cell Commitment to Memory and Long-Term Maintenance of Immunological Memory in Old Mice. Antioxidants 2020, 9, 1152. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Moghaddam, A.B.; Amini, S.; Keramati, M.R.; Zarmehri, A.M.; Alamdari, A.H.; Damsaz, M.; Banpour, H.; Yarahmadi, A.; Koliakos, G. Application of methylene blue -vitamin C –N-acetyl cysteine for treatment of critically ill COVID-19 patients, report of a phase-I clinical trial. Eur. J. Pharmacol. 2020, 885, 173494. [Google Scholar] [CrossRef]
- Lanna, A.; Henson, S.M.; Escors, D.; Akbar, A.N. The kinase p38 activated by the metabolic regulator AMPK and scaffold TAB1 drives the senescence of human T cells. Nat. Immunol. 2014, 15, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Passos, J.F.; Von Zglinicki, T. Oxygen free radicals in cell senescence: Are they signal transducers? Free Radic. Res. 2006, 40, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Blättler, S.M.; Rencurel, F.; Kaufmann, M.R.; Meyer, U.A. In the regulation of cytochrome P450 genes, phenobarbital targets LKB1 for necessary activation of AMP-activated protein kinase. Proc. Natl. Acad. Sci. USA 2007, 104, 1045–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slobodnyuk, K.; Radic, N.; Ivanova, S.; Llado, A.; Trempolec, N.; Zorzano, A.; Nebreda, A.R. Autophagy-induced senescence is regulated by p38α signaling. Cell Death Dis. 2019, 10, 376. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Factors | CD28null Subset | Adverse Effects |
|---|---|---|
| Aging | CD8+ | ↓Naïve T-cell pool → ↓Antigenic diversity → ↓Immune response [10,14] ↓B cell Population [27] ↑T-cell senescence, inferred [12] ↑Progression of Alzheimer’s disease [28] ↑IL-6; ↑IFN-γ → ↑IL-15 [29] ↑CD94/NKG2 → ↑Cytotoxicity [10,14] ↓Autophagy [30] |
| CD4+ | ↑NK receptors → ↑Inflammation and cytotoxicity [10,13,31] ↓Dnmt1and Dnmt3a → ↑KIR, perforin, and CD70 [32] ↑CX3CR1 [33] | |
| Diabetes | CD8+ | ↓Immune response in children [34] ↓Immune response in adults [35] ↑Risk of developing hyperglycemia [36] ↑ROS; ↑Glycolysis → ↑T2D development [37] |
| CD4+ | ↑Risk of acute coronary syndrome [13] ↑IL-17 in CD4+ CD28null NKG2D+ T cells → ↑Systemic inflammation; ↑HbA1c [38] ↑HbA1c and urinary albumin creatinine ratio [39] ↑Risk of cardiovascular events [21] | |
| COPD | CD8+ | ↑Pro-inflammatory cytokines, granzyme and perforin [40] ↓Glucocorticoid receptor → ↑Steroid resistance [22] ↓ Glucocorticoid receptor and Hsp90 → ↑IFNγ [41] ↓SIRT1 → ↑IFNγ, TNFα, steroid resistance, and disease severity [42] ↓Apoptosis; ↑Potential of tissue injury [43] |
| CD4+ | ↑IFNγ → matrix metalloproteinases and tissue destruction; ↑Apoptosis of lung epithelium [44] ↑Natural killer-like T-cell receptors CD94 and CD158; ↑Intracellular perforin and granzyme B; ↑TNF increase and IFNγ only increase at early disease [45] | |
| Hypertension | CD8+ | ↑Development of left ventricular hypertrophy (LVH) [46] ↑Granzyme B and Perforin; ↑CXCR3 chemokines, MIG, IP-10 and I-TAC; ↑IFNγ and TNFα [47] |
| CD4+ | ↑IFNγ, IL-6, IL-17, and TNFα; ↑Granzyme, and perforin; ↓Flow-mediated dilation [48] | |
| CVD | CD8+ | ↑IFNγ and TNFα [49] ↑IFNγ [50] |
| CD4+ | ↑Risk of plaque instability, acute coronary syndromes, and stroke [10] ↑IFNγ and TNFα; ↑Perforin, granzyme A, and granzyme B [51] ↑Cytotoxins and cytokines [20] ↑Risk of complications in follow-up surgeries; ↓Risk of first-time coronary event [52] End-stage renal disease: ↑IFNγ; ↓IL-4; ↑Granzyme B and perforin; ↓Flow-mediated vasodilation; ↑Carotid-intima media thickness (cIMT) [53] Pediatric T1D: ↑Aortic stiffness, and cIMT [39] Rheumatoid arthritis: ↑cIMT; ↓FMEDD [54] Kidney transplantation: ↑Risk of an atherosclerotic vascular event [55] Systemic lupus erythematosus: ↑Anti-dsDNA and anti-SSA/Ro; ↑ TNFα, IL-8, IFNα, and B lymphocyte stimulator [56] Acute coronary syndrome: ↑Non-ST-segment elevation ACS (NSTEACS) versus ST-segment elevation myocardial infarction (STEMI) [57] Chronic heart failure: ↑Mortality rate [58] | |
| Cancer | CD8+ | Non-small lung cancer: ↑Poor prognosis [59] Lung cancer: ↑Foxp3 mRNA in CD8+ CD28null T-cells; ↓Immune response, inferred [60] Metastatic breast cancer: ↑IL-6 and IL-10; ↓Progression-free survival [61] Colorectal cancer: ↑Suppression of cytotoxic function of T-cells; ↑Suppression of T-cell proliferation [62] Melanoma: ↑NK receptors, CD94, NKG2A, CD56, CD57, CD16, and CD244; ↑Perforin [63] |
| CD4+ | Non-small lung cancer (with immunotherapy): ↑Risk of hyperprogressive disease after anti-PD-1/PD-L1 immunotherapy [64] Cervical cancer: ↑NKG2D; ↑Perforin [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coleman, M.J.; Zimmerly, K.M.; Yang, X.O. Accumulation of CD28null Senescent T-Cells Is Associated with Poorer Outcomes in COVID19 Patients. Biomolecules 2021, 11, 1425. https://doi.org/10.3390/biom11101425
Coleman MJ, Zimmerly KM, Yang XO. Accumulation of CD28null Senescent T-Cells Is Associated with Poorer Outcomes in COVID19 Patients. Biomolecules. 2021; 11(10):1425. https://doi.org/10.3390/biom11101425
Chicago/Turabian StyleColeman, Mia J., Kourtney M. Zimmerly, and Xuexian O. Yang. 2021. "Accumulation of CD28null Senescent T-Cells Is Associated with Poorer Outcomes in COVID19 Patients" Biomolecules 11, no. 10: 1425. https://doi.org/10.3390/biom11101425