Enhanced Biomass Yield of and Saccharification in Transgenic Tobacco Over-Expressing β-Glucosidase

Abstract
:1. Introduction
2. Materials and Methods
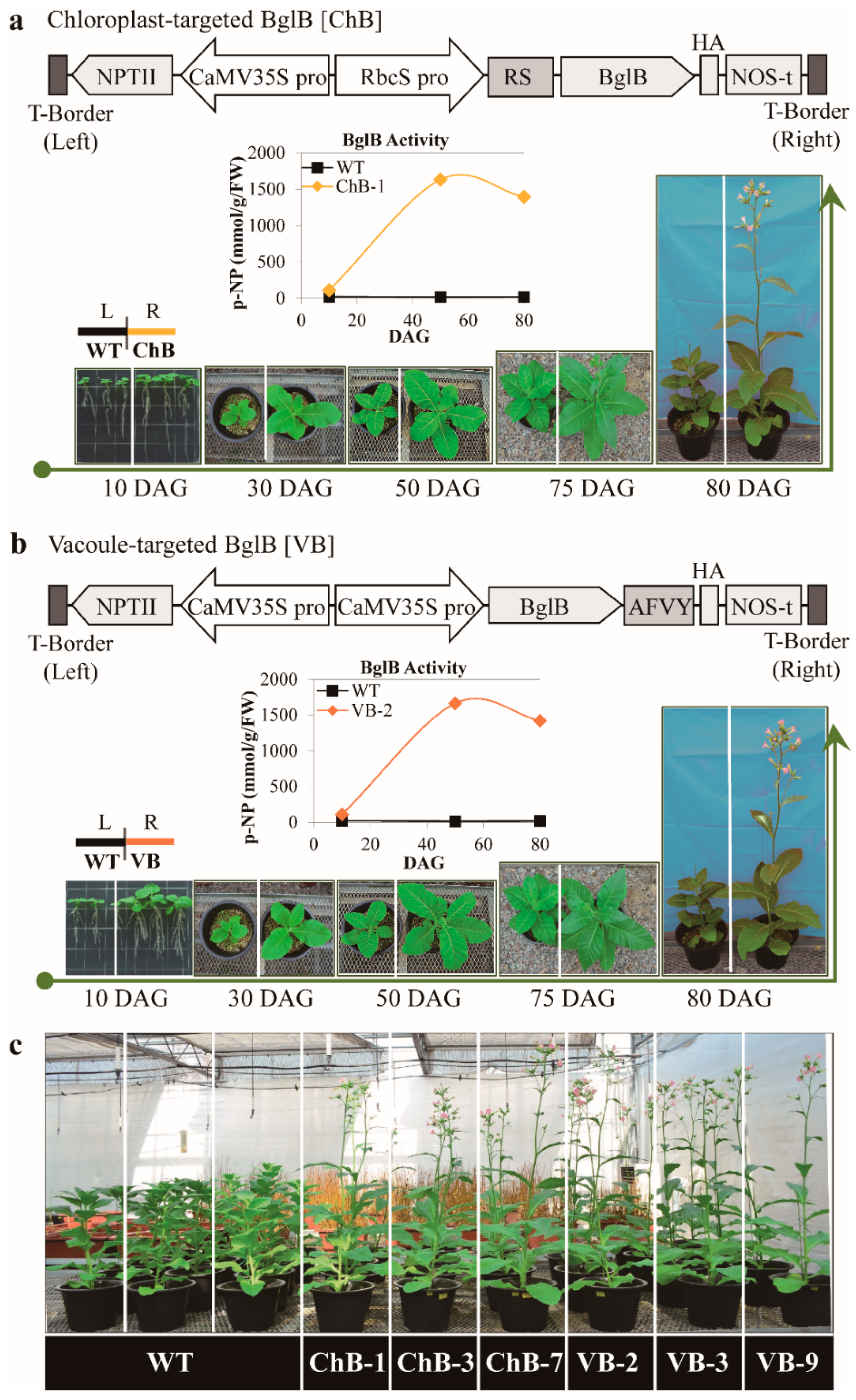
2.1. Collection of Transgenic Tobacco Plants Overexpressing Thermostable β-Glucosidase T. maritima BglB Targeting Chloroplasts and Vacuoles, Planting Conditions, and Measurement of Phenotypic Characteristics
2.2. Measurement of β-Glucosidase Enzymatic Activities
2.3. mRNA Microarray and Transcriptional Expression Levels
2.4. Carbohydrate Contents and Composition Analysis
2.5. Pretreatment by Mild Acid and Saccharification
2.6. Techno-Economic Assessment—Costs for Planting and Biomass Yields
3. Results and Discussion
3.1. Vector Construction for the Overexpression of Thermostable β-Glucosidase T. maritima BglB Targeted to the Chloroplasts and Vacuoles in Tobacco Plants and β-Glucosidase Enzymatic Activity
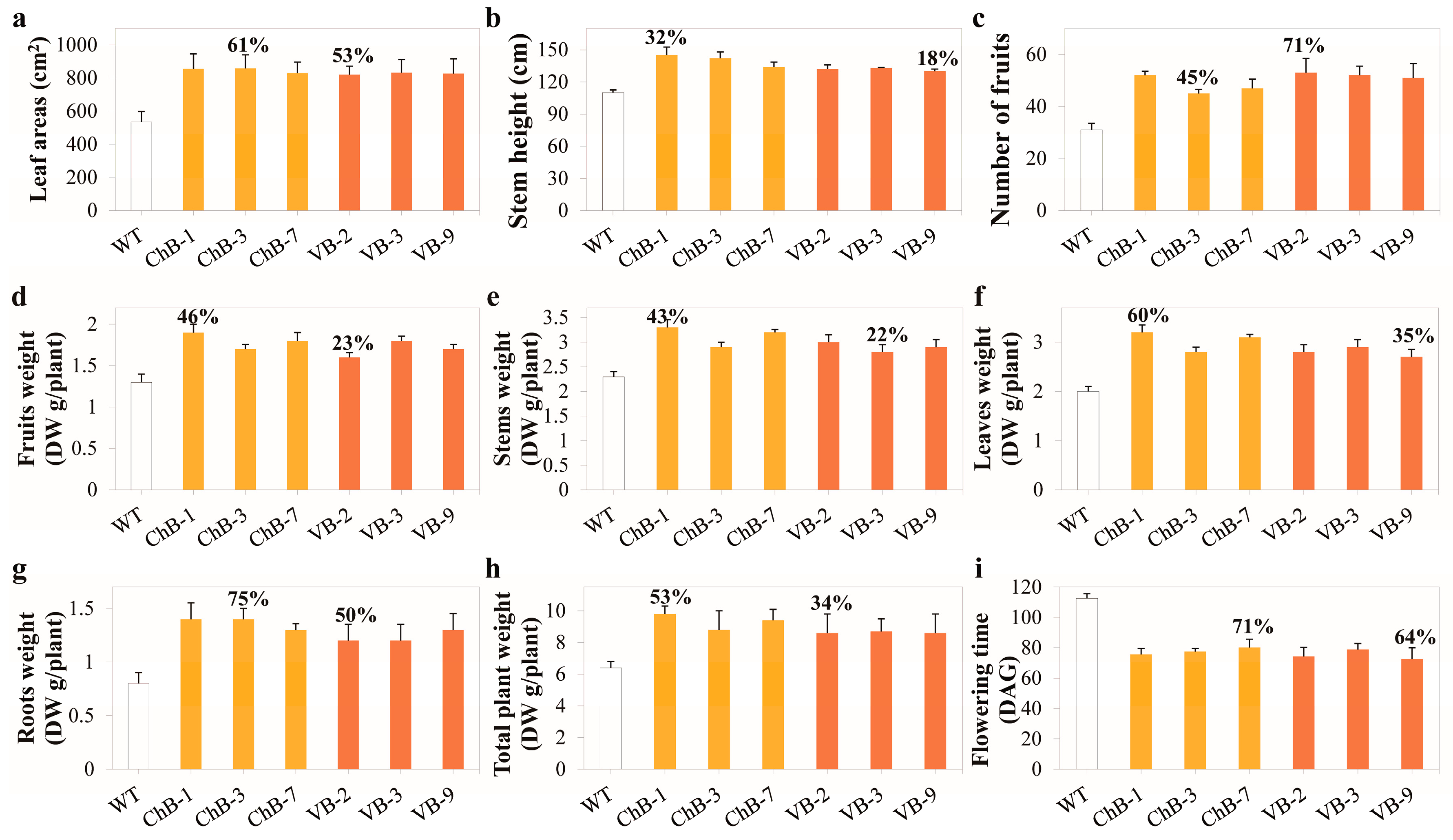
3.2. Phenotypes of Transgenic Plants Overexpressing T. maritima BglB Targeted to the Chloroplasts and Vacuoles Enhanced Growth and Biomass Yield
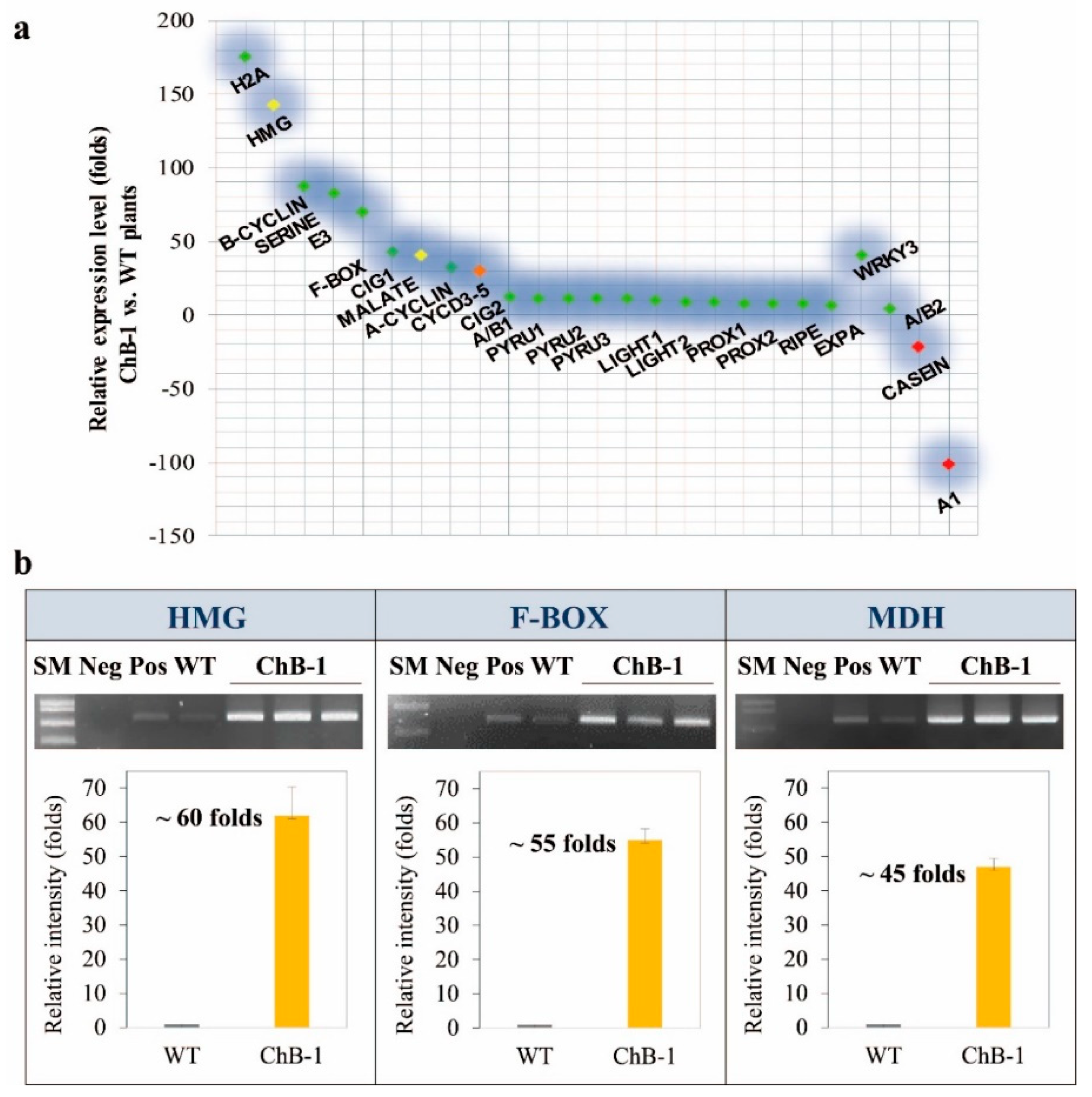
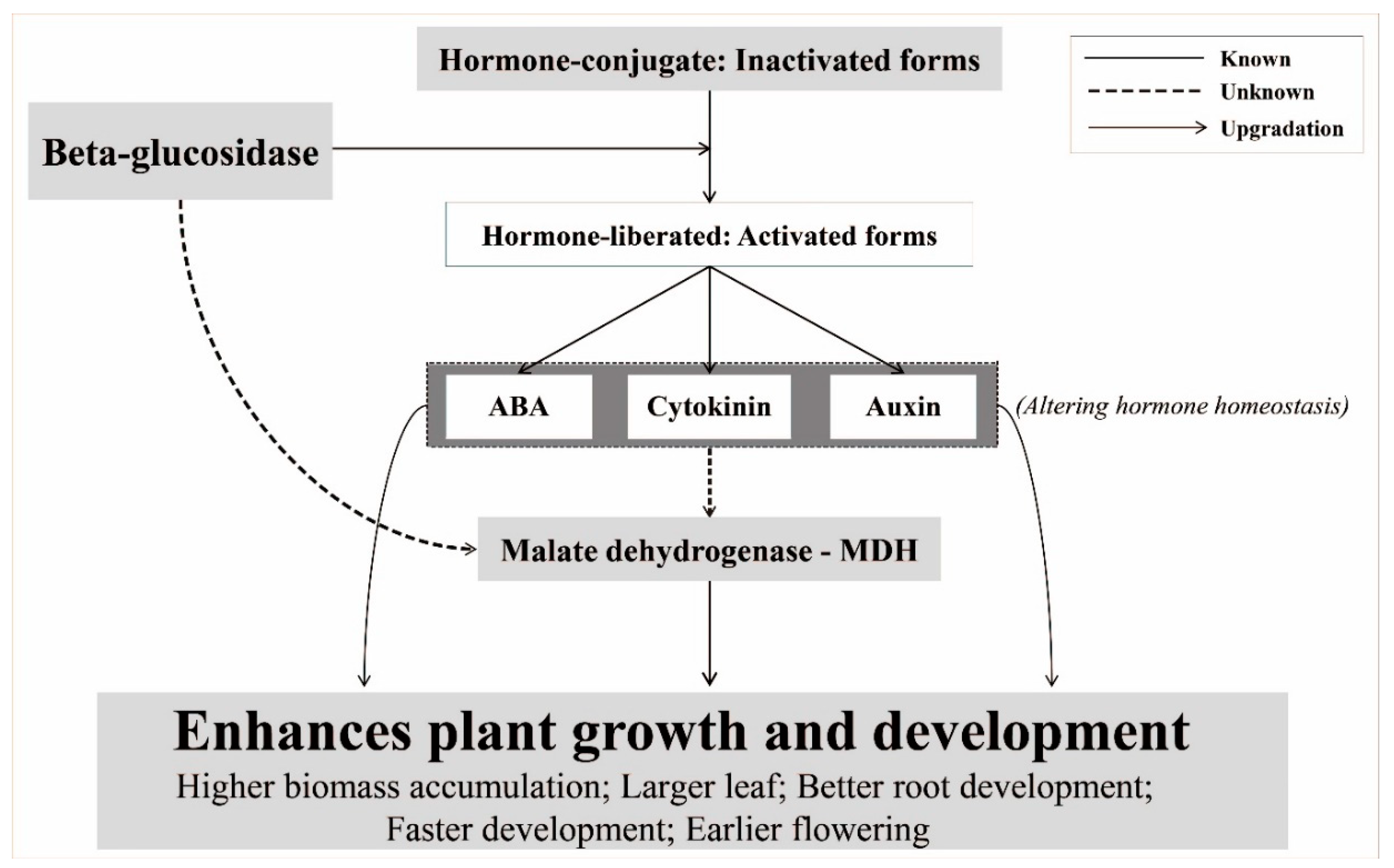
3.3. Involvement of HMG, F-Box, and MDH in the Transgenic Plants
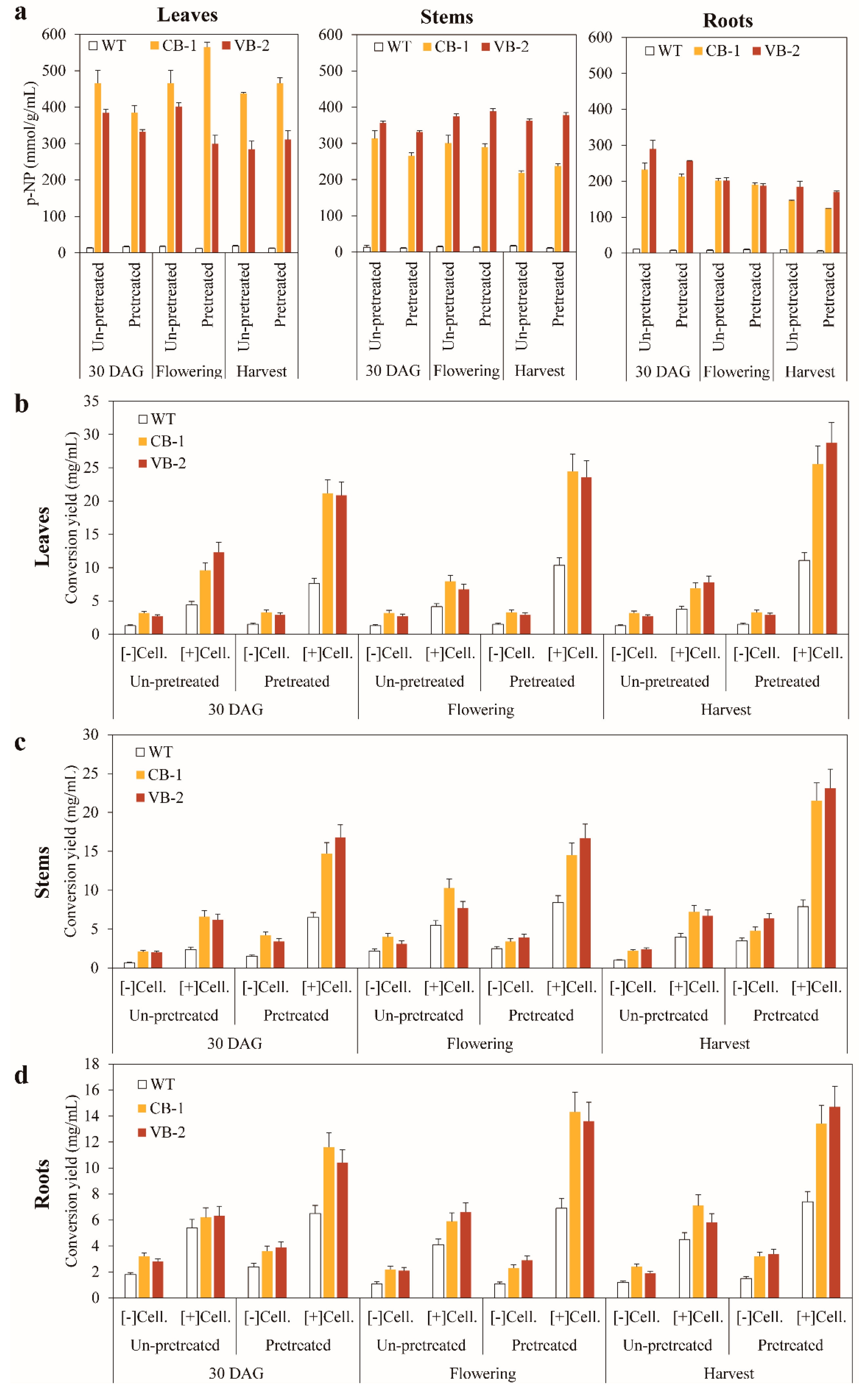
3.4. Enhanced Saccharification in the Enzymatic Conversion of Biomass from Transgenic Plants Expressing β-Glucosidase T. maritima BglB
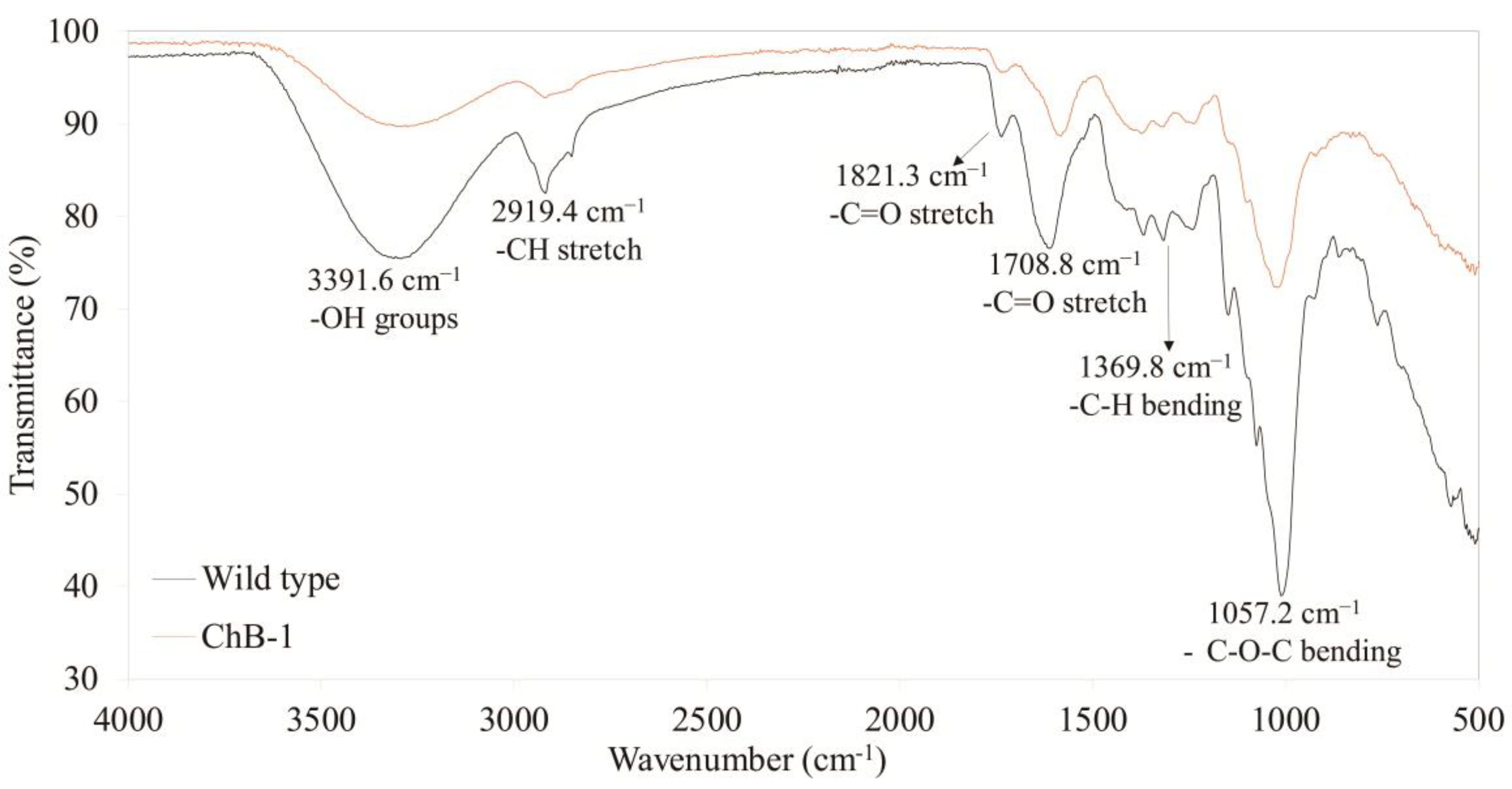
3.5. Difference in the Fourier Transformed Infrared (FT-IR) Spectra between the Transgenic and Wild-Type Tobacco Plants
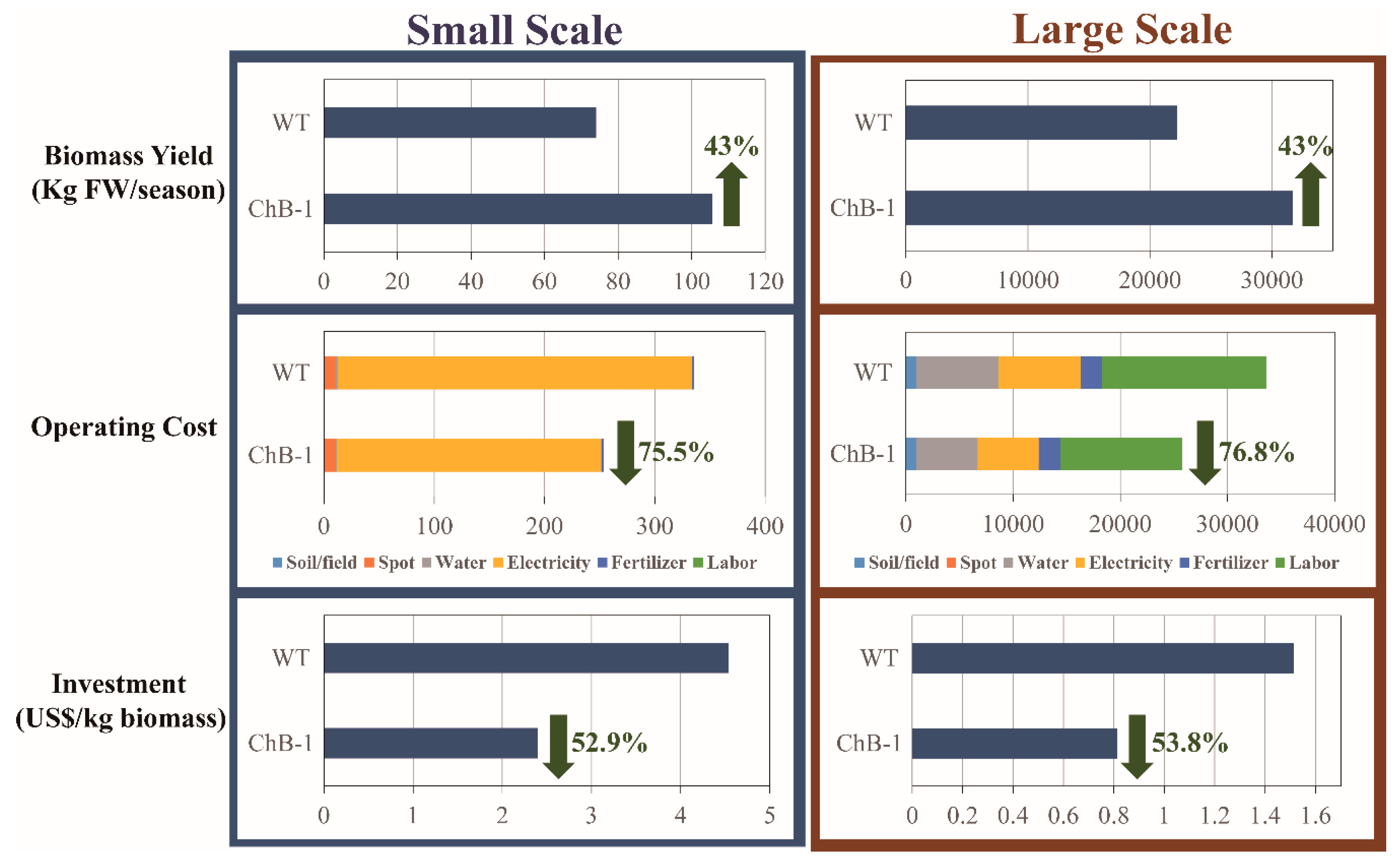
3.6. Techno–Economic Assessment of the Increase in Biomass Production in the Transgenic Tobacco Plants as Compared with the Wild-Type Tobacco Plants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nguyen, Q.A.; Cho, E.J.; Trinh, L.T.P.; Jeong, J.S.; Bae, H.J. Development of an integrated process to produce d-mannose and bioethanol from coffee residue waste. Bioresour. Technol. 2017, 244, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.A.; Cho, E.J.; Lee, D.S.; Bae, H.J. Development of an advanced integrative process to create valuable biosugars including manno-oligosaccharides and mannose from spent coffee grounds. Bioresour. Technol. 2019, 272, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; Scheller, H.V.; Pauly, M. Engineering of plant cell walls for enhanced biofuel production. Curr. Opin. Plant Biol. 2015, 25, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sticklen, M.B. Plant genetic engineering for biofuel production: Towards affordable cellulosic ethanol. Nat. Rev. Gen. 2008, 9, 433–443. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, C.; Hu, H.; Li, Y.; Sun, D.; Wang, Y.; Peng, L. Genetic modification of plant cell walls to enhance biomass yield and biofuel production in bioenergy crops. Biotechnol. Adv. 2016, 34, 997–1017. [Google Scholar] [CrossRef]
- Jin, S.; Kanagaraj, A.; Verma, D.; Lange, T.; Daniell, H. Release of hormones from conjugates: Chloroplast expression of beta-glucosidase results in elevated phytohormone levels associated with significant increase in biomass and protection from aphids or whiteflies conferred by sucrose esters. Plant Physiol. 2011, 155, 222–235. [Google Scholar] [CrossRef]
- Nguyen, Q.A.; Lee, D.S.; Jung, J.; Bae, H.J. Phenotypic changes in transgenic tobacco plants overexpressing vacuole-targeted Thermotoga maritima BglB related to elevated levels of liberated hormones. Front. Bioeng. Biotechnol. 2015, 3, 181–192. [Google Scholar] [CrossRef] [Green Version]
- South, P.F.; Cavanagh, A.P.; Liu, H.W.; Ort, D.R. Synthetic glycolate metabolism pathways stimulate crop growth and productivity in the field. Science 2019, 363, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Brzobohatý, B.; Moore, I.; Kristoffersen, P.; Bako, L.; Campos, N.; Schell, J.; Palme, K. Release of Active Cytokinin by a beta-Glucosidase Localized to the Maize Root Meristem. Science 1993, 262, 1051–1054. [Google Scholar] [CrossRef]
- Ketudat Cairns, J.R.; Esen, A. β-Glucosidases. Cell. Mol. Life Sci. 2010, 67, 3389–3405. [Google Scholar] [CrossRef]
- Ketudat Cairns, J.R.; Mahong, B.; Baiya, S.; Jeon, J.S. β-Glucosidases: Multitasking, moonlighting or simply misunderstood? Plant Sci. 2015, 241, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, Z.; Huo, Y.-Y.; Bao, L.; Gao, B.; Xiao, P.; Hu, X.; Xu, X.-W.; Li, J. Structural and functional insights into CmGH1, a novel GH39 family β-glucosidase from deep-sea bacterium. Front. Microbiol. 2019, 10, 2922–2933. [Google Scholar] [CrossRef] [PubMed]
- Morant, A.V.; Jørgensen, K.; Jørgensen, C.; Paquette, S.M.; Sánchez-Pérez, R.; Møller, B.L.; Bak, S. β-Glucosidases as detonators of plant chemical defense. Phytochemistry 2008, 69, 1795–1813. [Google Scholar] [CrossRef] [PubMed]
- Lindén, P.; Keech, O.; Stenlund, H.; Gardeström, P.; Moritz, T. Reduced mitochondrial malate dehydrogenase activity has a strong effect on photorespiratory metabolism as revealed by 13C labelling. J. Exp. Bot. 2016, 67, 3123–3135. [Google Scholar] [CrossRef] [Green Version]
- Van der Merwe, M.J.; Osorio, S.; Moritz, T.; Nunes-Nesi, A.; Fernie, A.R. Decreased mitochondrial activities of malate dehydrogenase and fumarase in tomato lead to altered root growth and architecture via diverse mechanisms. Plant Physiol. 2009, 149, 653–669. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.F.; Zhao, Y.; Yi, Q.; Li, K.Z.; Yu, Y.X.; Chen, L.M. Overexpression of malate dehydrogenase in transgenic tobacco leaves: Enhanced malate synthesis and augmented Al-resistance. Act. Phys. Plan. 2010, 32, 1209–1220. [Google Scholar] [CrossRef]
- Kiran, N.S.; Benková, E.; Reková, A.; Dubová, J.; Malbeck, J.; Palme, K.; Brzobohatý, B. Retargeting a maize β-glucosidase to the vacuole – Evidence from intact plants that zeatin-O-glucoside is stored in the vacuole. Phytochemistry 2012, 79, 67–77. [Google Scholar] [CrossRef]
- Li, Y.; Liu, P.; Huang, J.; Zhang, R.; Hu, A.; Feng, S.; Wang, Y.; Wang, L.; Xia, T.; Peng, L. Mild chemical pretreatments are sufficient for bioethanol production in transgenic rice straws overproducing glucosidase. Green Chem. 2018, 20, 2047–2056. [Google Scholar] [CrossRef]
- Park, S.-H.; Ong, R.G.; Sticklen, M. Strategies for the production of cell wall-deconstructing enzymes in lignocellulosic biomass and their utilization for biofuel production. Plant Biotechnol. J. 2016, 14, 1329–1344. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Pandey, A.; Ganansounou, E. Genetic modification: A tool for enhancing beta-glucosidase production for biofuel application. Bioresour. Technol. 2017, 245, 1352–1361. [Google Scholar] [CrossRef]
- Jung, S.; Lee, D.-S.; Kim, Y.-O.; Joshi, C.; Bae, H.-J. Improved recombinant cellulase expression in chloroplast of tobacco through promoter engineering and 5′ amplification promoting sequence. Plant Mol. Biol. 2013, 83, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.A.; Yang, J.; Bae, H.-J. Bioethanol production from individual and mixed agricultural biomass residues. Ind. Crop. Prod. 2017, 95, 718–725. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass; National Renewable Energy Laboratory: Golden, CO, USA, 2008.
- Jung, S.; Kim, S.; Bae, H.; Lim, H.-S.; Bae, H.-J. Expression of thermostable bacterial β-glucosidase (BglB) in transgenic tobacco plants. Bioresour. Technol. 2010, 101, 7144–7150. [Google Scholar] [CrossRef] [PubMed]
- Biswal, A.K.; Atmodjo, M.A.; Li, M.; Baxter, H.L.; Yoo, C.G.; Pu, Y.; Lee, Y.-C.; Mazarei, M.; Black, I.M.; Hang, J.-Y.; et al. Sugar release and growth of biofuel crops are improved by downregulation of pectin biosynthesis. Nat. Biotechnol. 2018, 36, 249–257. [Google Scholar] [CrossRef]
- Kang, R.; Chen, R.; Zhang, Q.; Hou, W.; Wu, S.; Cao, L.; Huang, J.; Yu, T.; Fan, X.; Yan, Z.; et al. HMGB1 in health and disease. Mol. Asp. Med. 2014, 40, 1–116. [Google Scholar] [CrossRef] [Green Version]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Env. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- O’Hara, L.E.; Paul, M.J.; Wingler, A. How Do Sugars Regulate Plant Growth and Development? New Insight into the Role of Trehalose-6-Phosphate. Mol. Plant 2013, 6, 261–274. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-F.; Zhao, J.; Zhang, J.; Dai, Z.-H.; Zhang, L.-G. Ectopic Expression of the Chinese Cabbage Malate Dehydrogenase Gene Promotes Growth and Aluminum Resistance in Arabidopsis. Front. Plant Sci. 2016, 7, 1180–1191. [Google Scholar] [CrossRef] [Green Version]
- Tomaz, T.; Bagard, M.; Pracharoenwattana, I.; Lindén, P.; Lee, C.P.; Carroll, A.J.; Ströher, E.; Smith, S.M.; Gardeström, P.; Millar, A.H. Mitochondrial Malate Dehydrogenase Lowers Leaf Respiration and Alters Photorespiration and Plant Growth in Arabidopsis. Plant Physiol. 2010, 154, 1143–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesfaye, M.; Temple, S.J.; Allan, D.L.; Vance, C.P.; Samac, D.A. Overexpression of Malate Dehydrogenase in Transgenic Alfalfa Enhances Organic Acid Synthesis and Confers Tolerance to Aluminum. Plant Physiol. 2001, 127, 1836–1844. [Google Scholar] [CrossRef]
- Xia, W.; Xu, A.; Qian, L.; Shi, P.; Bai, Y.; Luo, H.; Ma, R.; Yao, B. Engineering a highly active thermophilic β-glucosidase to enhance its pH stability and saccharification performance. Biotechnol. Biofuels 2016, 9, 147–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Heading | % | WT | ChB-1 | ChB-3 | ChB-7 | VB-2 | VB-3 | VB-9 |
|---|---|---|---|---|---|---|---|---|
| Leaves | Rhamnose | 2.2 ± 0.2 | 2.3 ± 0.3 | 2.2 ± 0.2 | 2.7 ± 0.3 | 2.5 ± 0.3 | 2.5 ± 0.4 | 2.0 ± 0.0 |
| Arabinose | 1.3 ± 0.0 | 1.5 ± 0.1 | 1.3 ± 0.1 | 1.6 ± 0.2 | 1.0 ± 0.0 | 1.1 ± 0.2 | 1.3 ± 0.1 | |
| Xylose | 1.5 ± 0.2 | 1.6 ± 0.1 | 1.5 ± 0.1 | 1.5 ± 0.2 | 1.4 ± 0.1 | 1.5 ± 0.1 | 1.2 ± 0.1 | |
| Mannose | 0.6 ± 0.0 | 1.1 ± 0.2 | 1.1 ± 0.1 | 0.6 ± 0.0 | 0.6 ± 0.0 | 0.4 ± 0.0 | 1.0 ± 0.0 | |
| Galactose | 2.2 ± 0.0 | 2.0 ± 0.0 | 2.0 ± 0.0 | 2.1 ± 0.1 | 2.1 ± 0.0 | 2.0 ± 0.0 | 2.0 ± 0.1 | |
| Glucose | 46.3 ± 1.1 | 46.0 ± 2.1 | 45.1 ± 2.0 | 45.1 ± 1.7 | 45.8 ± 5.1 | 45.1 ± 2.5 | 45.3 ± 1.1 | |
| Total | 54.1 ± 1.3 | 54.5 ± 3.1 | 53.2 ± 3.0 | 53.6 ± 2.1 | 53.4 ± 4.8 | 52.6 ± 3.3 | 52.8 ± 1.5 | |
| Stems | Rhamnose | 1.9 ± 0.1 | 1.3 ± 0.2 | 1.0 ± 0.1 | 1.3 ± 0.1 | 1.2 ± 0.0 | 1.1 ± 0.1 | 1.3 ± 0.2 |
| Arabinose | 0.8 ± 0.0 | 0.9 ± 0.0 | 1.1 ± 0.0 | 1.1 ± 0.0 | 1.3 ± 0.1 | 0.7 ± 0.1 | 1.2 ± 0.0 | |
| Xylose | 5.4 ± 0.5 | 5.5 ± 0.4 | 6.3 ± 0.2 | 7.2 ± 1.4 | 6.4 ± 0.7 | 5.4 ± 0.2 | 6.0 ± 0.2 | |
| Mannose | 1.1 ± 0.2 | 1.4 ± 0.1 | 1.3 ± 0.0 | 1.3 ± 0.2 | 1.0 ± 0.1 | 1.3 ± 0.0 | 1.3 ± 0.1 | |
| Galactose | 1.3 ± 0.4 | 1.5 ± 0.2 | 1.2 ± 0.1 | 1.3 ± 0.0 | 1.2 ± 0.1 | 1.2 ± 0.0 | 1.2 ± 0.1 | |
| Glucose | 45.1 ± 2.7 | 45.3 ± 1.3 | 45.8 ± 2.1 | 46.2 ± 1.4 | 45.8 ± 1.7 | 46.1 ± 2.9 | 44.1 ± 1.8 | |
| Total | 55.6 ± 3.1 | 55.9 ± 2.1 | 56.7 ± 3.5 | 58.4 ± 2.4 | 56.9 ± 3.7 | 55.8 ± 3.4 | 55.1 ± 2.6 | |
| Roots | Rhamnose | 1.2 ± 0.0 | 1.2 ± 0.1 | 1.5 ± 0.0 | 1.2 ± 0.0 | 1.5 ± 0.2 | 1.5 ± 0.3 | 1.1 ± 0.1 |
| Arabinose | 0.9 ± 0.0 | 1.1 ± 0.1 | 1.0 ± 0.0 | 1.1 ± 0.0 | 1.2 ± 0.2 | 0.9 ± 0.1 | 0.7 ± 0.2 | |
| Xylose | 6.5 ± 1.3 | 7.0 ± 0.2 | 6.8 ± 0.5 | 6.5 ± 0.8 | 6.4 ± 0.8 | 7.1 ± 0.2 | 7.1 ± 0.3 | |
| Mannose | 1.0 ± 0.1 | 1.2 ± 0.1 | 1.3 ± 0.0 | 1.4 ± 0.1 | 0.8 ± 0.0 | 1.3 ± 0.1 | 1.2 ± 0.1 | |
| Galactose | 1.3 ± 0.1 | 1.0 ± 0.0 | 1.2 ± 0.1 | 1.2 ± 0.1 | 0.7 ± 0.0 | 0.8 ± 0.0 | 0.9 ± 0.0 | |
| Glucose | 45.7 ± 2.8 | 46.0 ± 3.9 | 45.3 ± 2.5 | 45.1 ± 0.9 | 44.9 ± 0.7 | 45.8 ± 2.4 | 45.9 ± 4.1 | |
| Total | 56.6 ± 3.8 | 57.5±4.1 | 57.1±3.3 | 56.5±1.6 | 55.5±3.8 | 57.4±4.1 | 57.2±4.8 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, E.J.; Nguyen, Q.A.; Lee, Y.G.; Song, Y.; Park, B.J.; Bae, H.-J. Enhanced Biomass Yield of and Saccharification in Transgenic Tobacco Over-Expressing β-Glucosidase. Biomolecules 2020, 10, 806. https://doi.org/10.3390/biom10050806
Cho EJ, Nguyen QA, Lee YG, Song Y, Park BJ, Bae H-J. Enhanced Biomass Yield of and Saccharification in Transgenic Tobacco Over-Expressing β-Glucosidase. Biomolecules. 2020; 10(5):806. https://doi.org/10.3390/biom10050806
Chicago/Turabian StyleCho, Eun Jin, Quynh Anh Nguyen, Yoon Gyo Lee, Younho Song, Bok Jae Park, and Hyeun-Jong Bae. 2020. "Enhanced Biomass Yield of and Saccharification in Transgenic Tobacco Over-Expressing β-Glucosidase" Biomolecules 10, no. 5: 806. https://doi.org/10.3390/biom10050806