Study of Synthesis Pathways of the Essential Polyunsaturated Fatty Acid 20:5n-3 in the Diatom Chaetoceros Muelleri Using 13C-Isotope Labeling

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
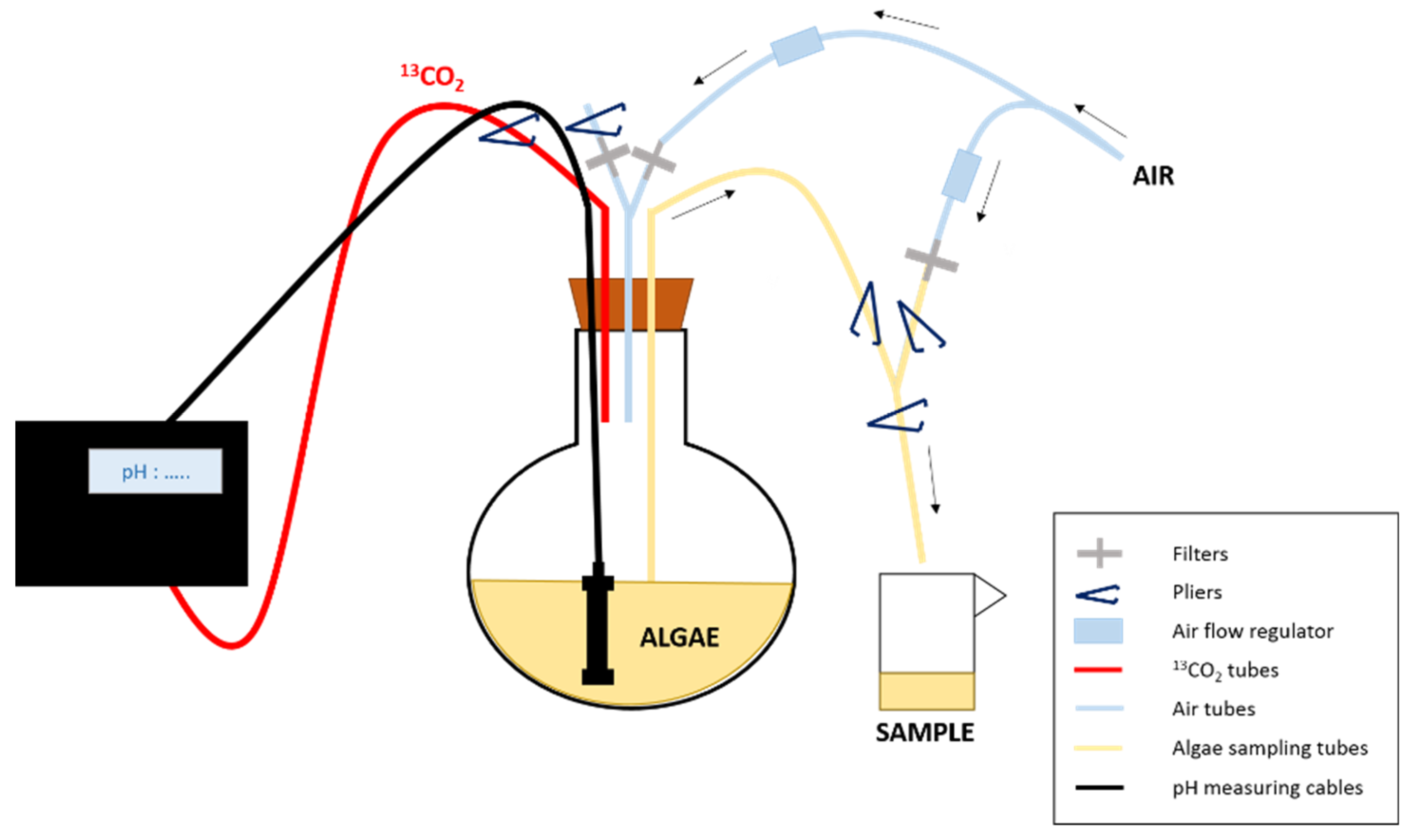
2.1. Algal Culture and Isotopic Labeling
2.2. Samples Collection
2.3. Flow Cytometry Analysis
2.4. POC Concentration and Bulk Carbon Isotopic Composition
2.5. DIC Concentrations and Bulk Carbon Isotopic Composition
2.6. Isotopic Data Processing
2.7. Fatty Acids Analysis
2.7.1. Lipid Extraction
2.7.2. Separation of Neutral and Polar Lipids
2.7.3. Transesterification of FAME
2.7.4. Fatty Acid Analysis by Gas Chromatography Flame Ionisation Detector (GC-FID)
2.7.5. Fatty Acids Compound-Specific Isotope Analysis
2.7.6. Compound-Specific Isotope Data Processing
2.8. Statistical Analysis
3. Results
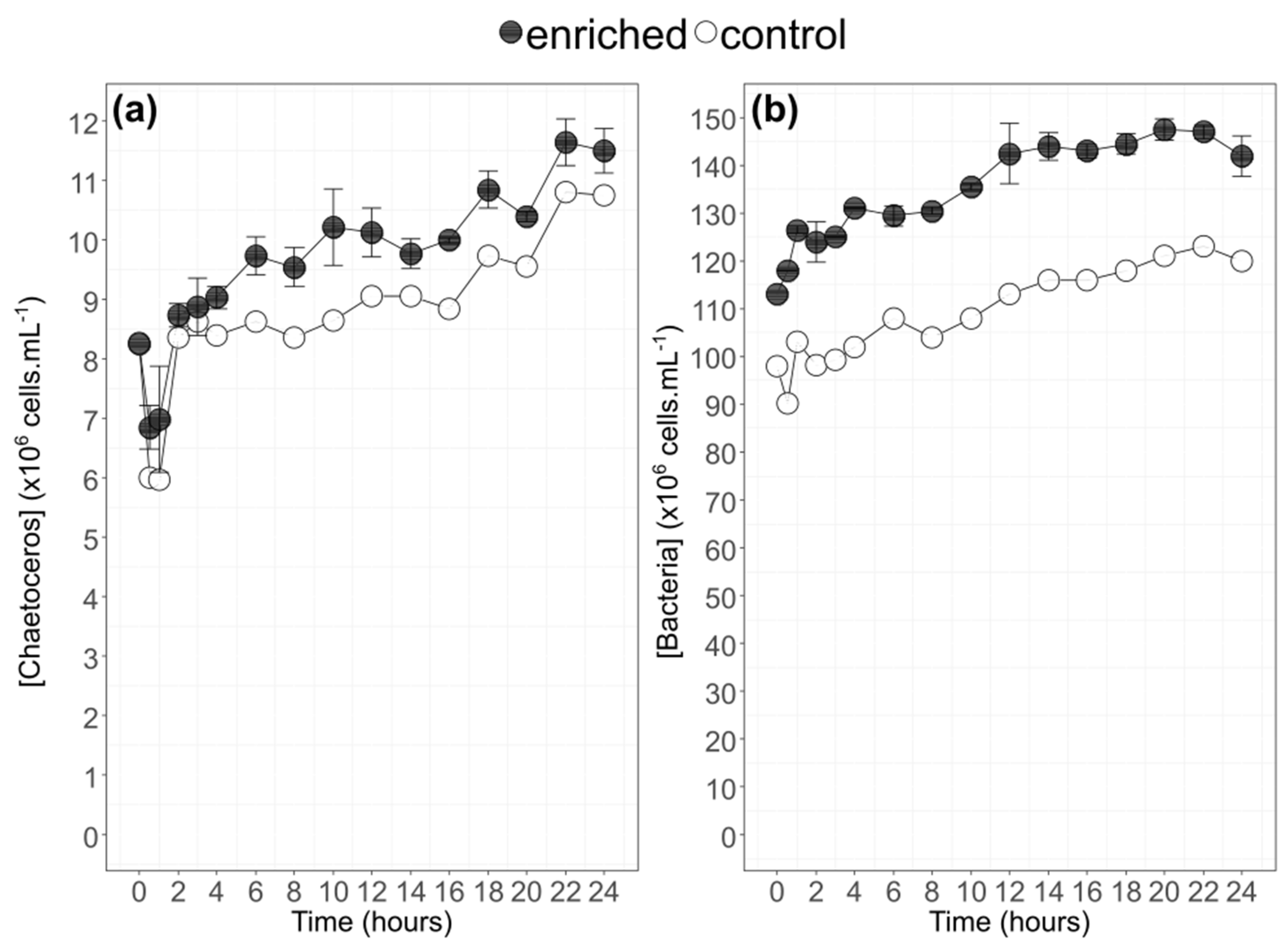
3.1. Algae Physiology During Growth
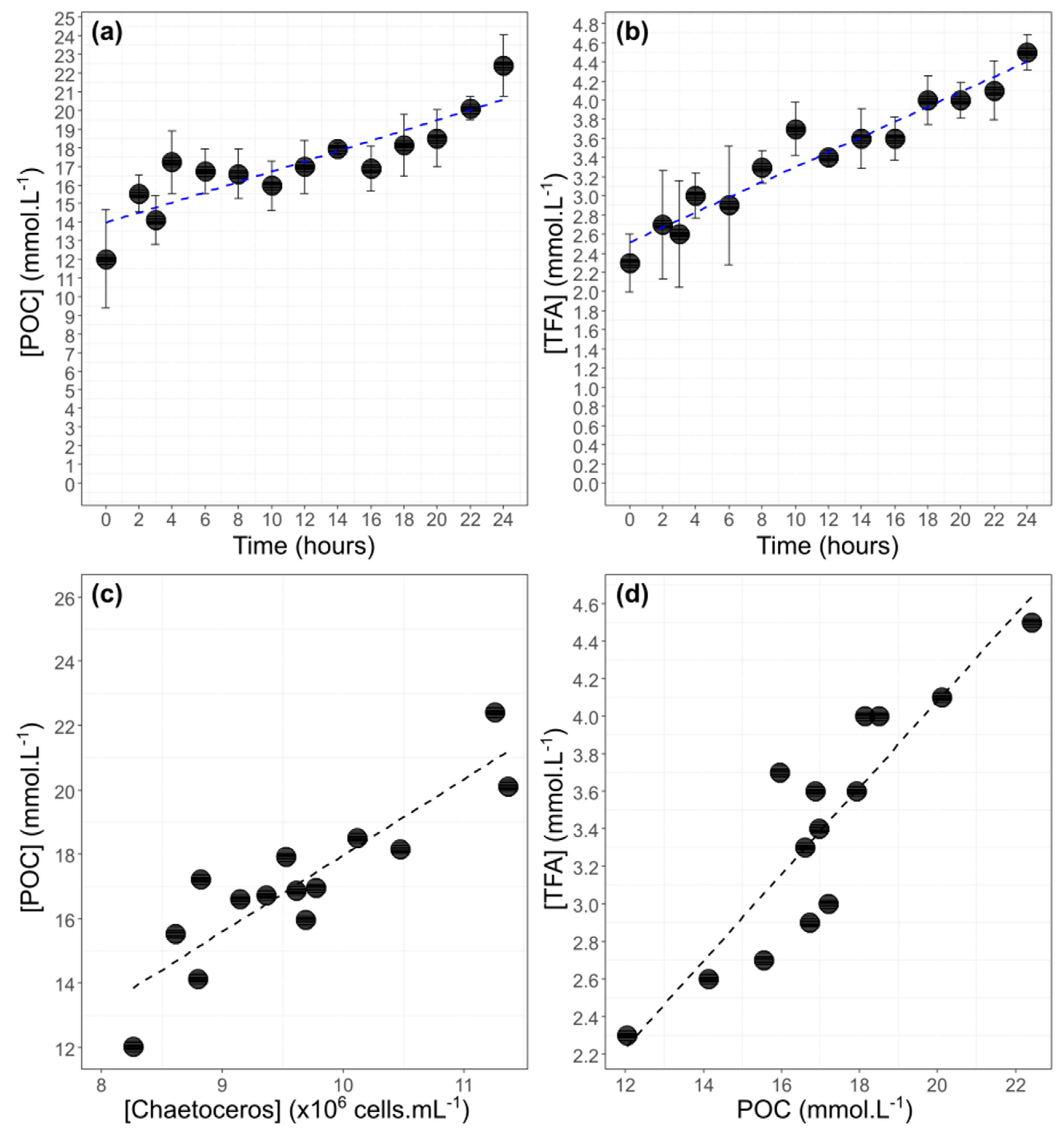
3.2. Correlation between POC Concentration, TFA Concentration, and Cell Abundance
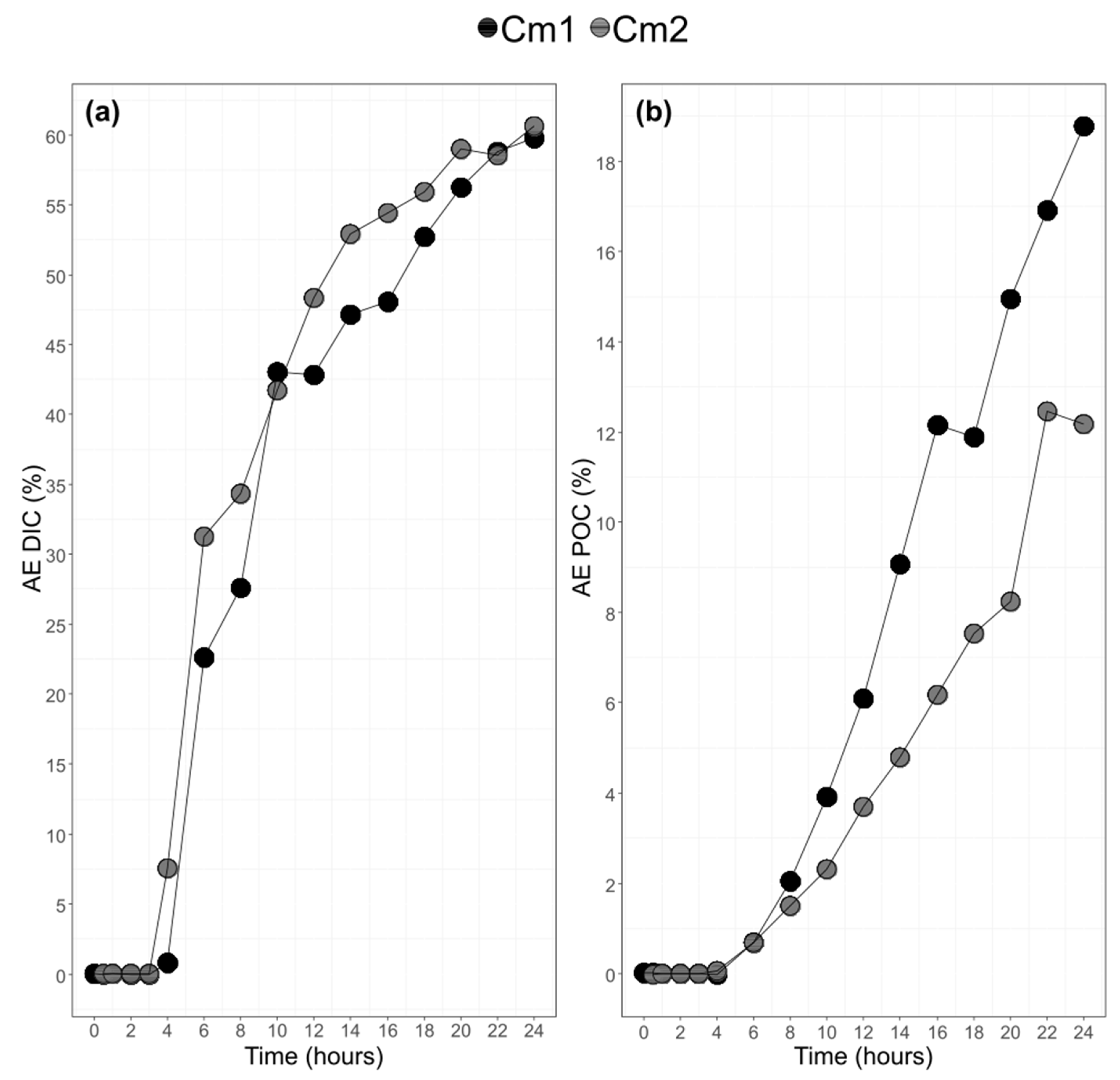
3.3. Bulk POC and DIC and their 13C-Labeling
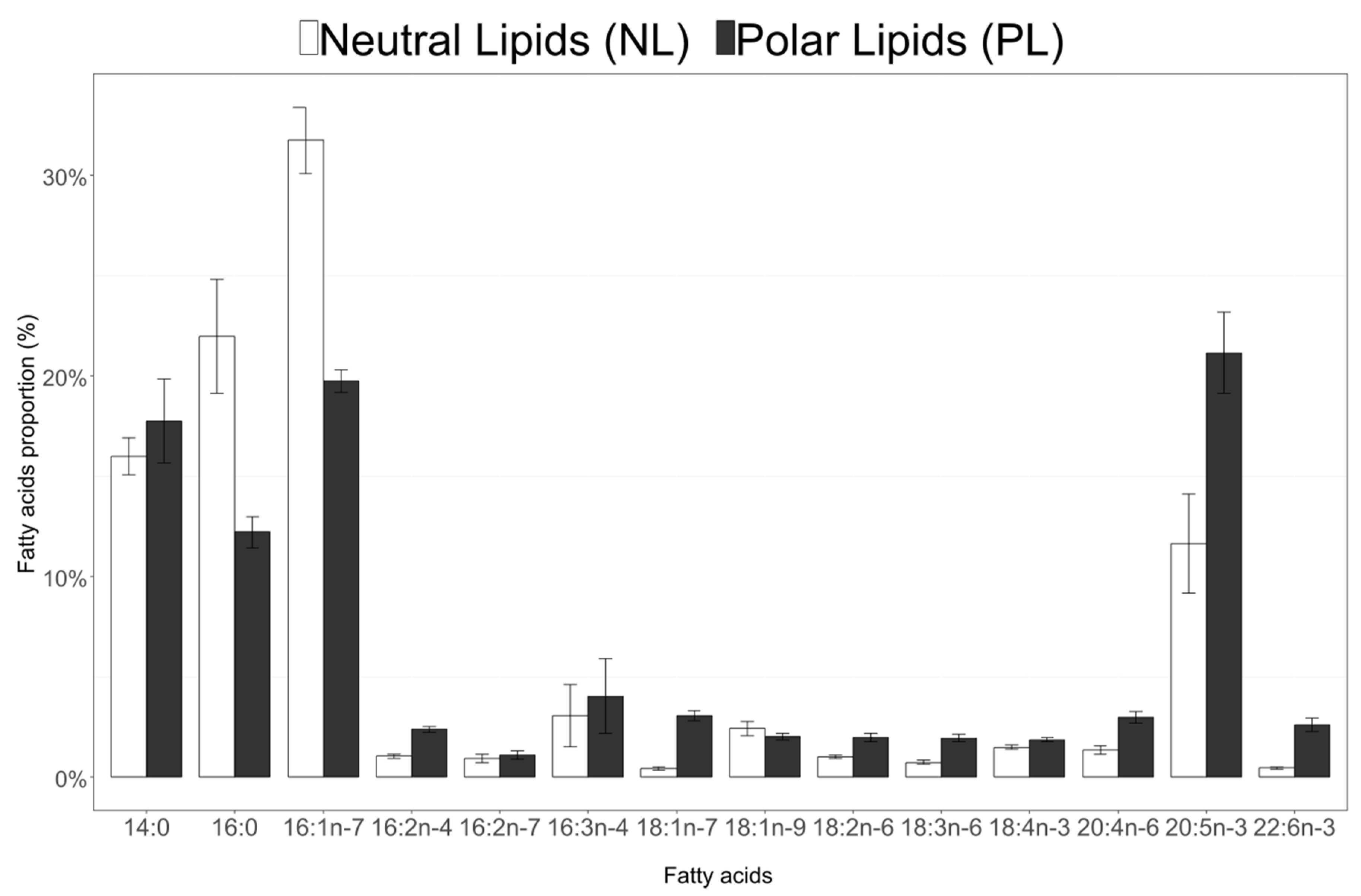
3.4. Fatty Acid Composition of Neutral and Polar Lipids in C. Muelleri
3.5. Variability of Fatty Acid Proportions during the Experiment
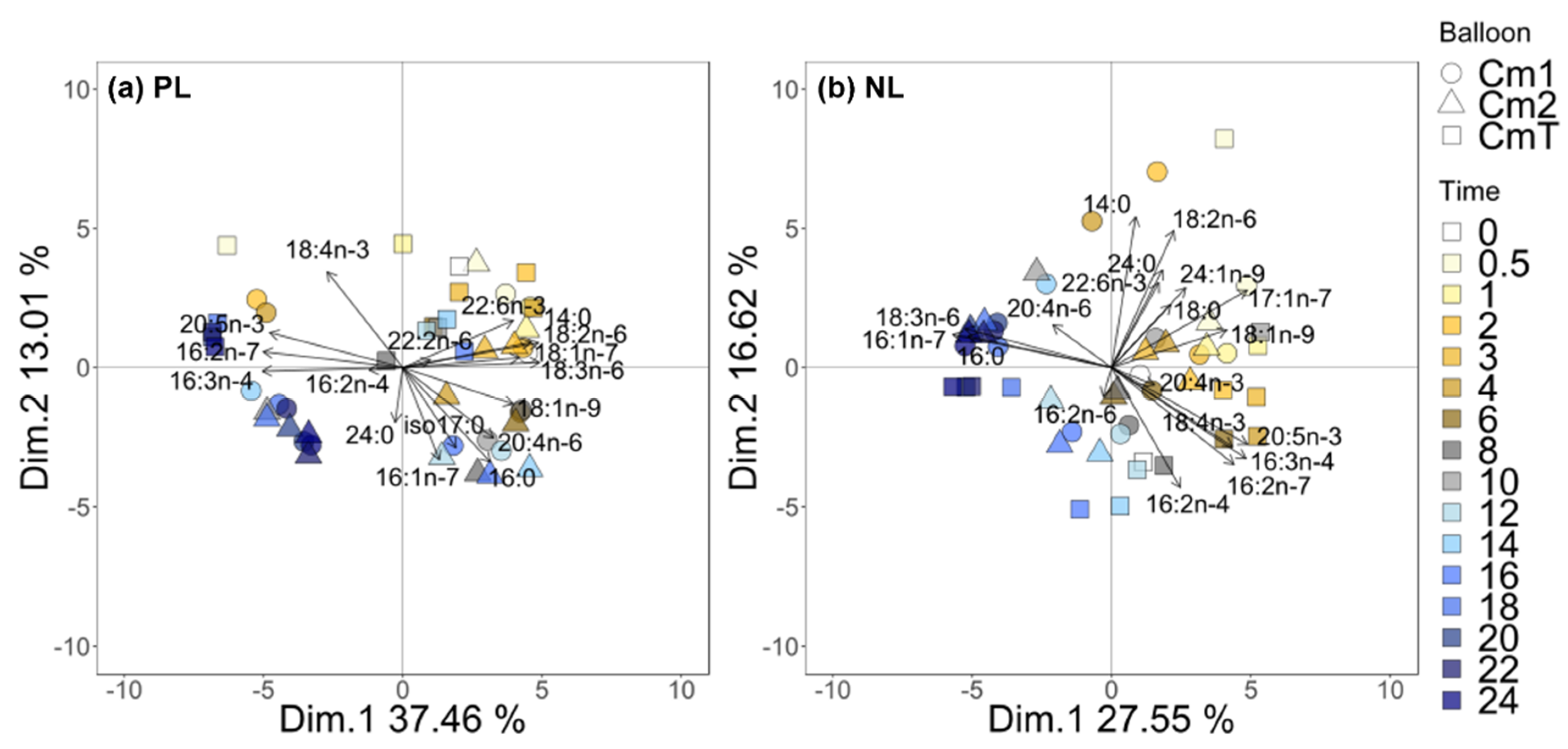
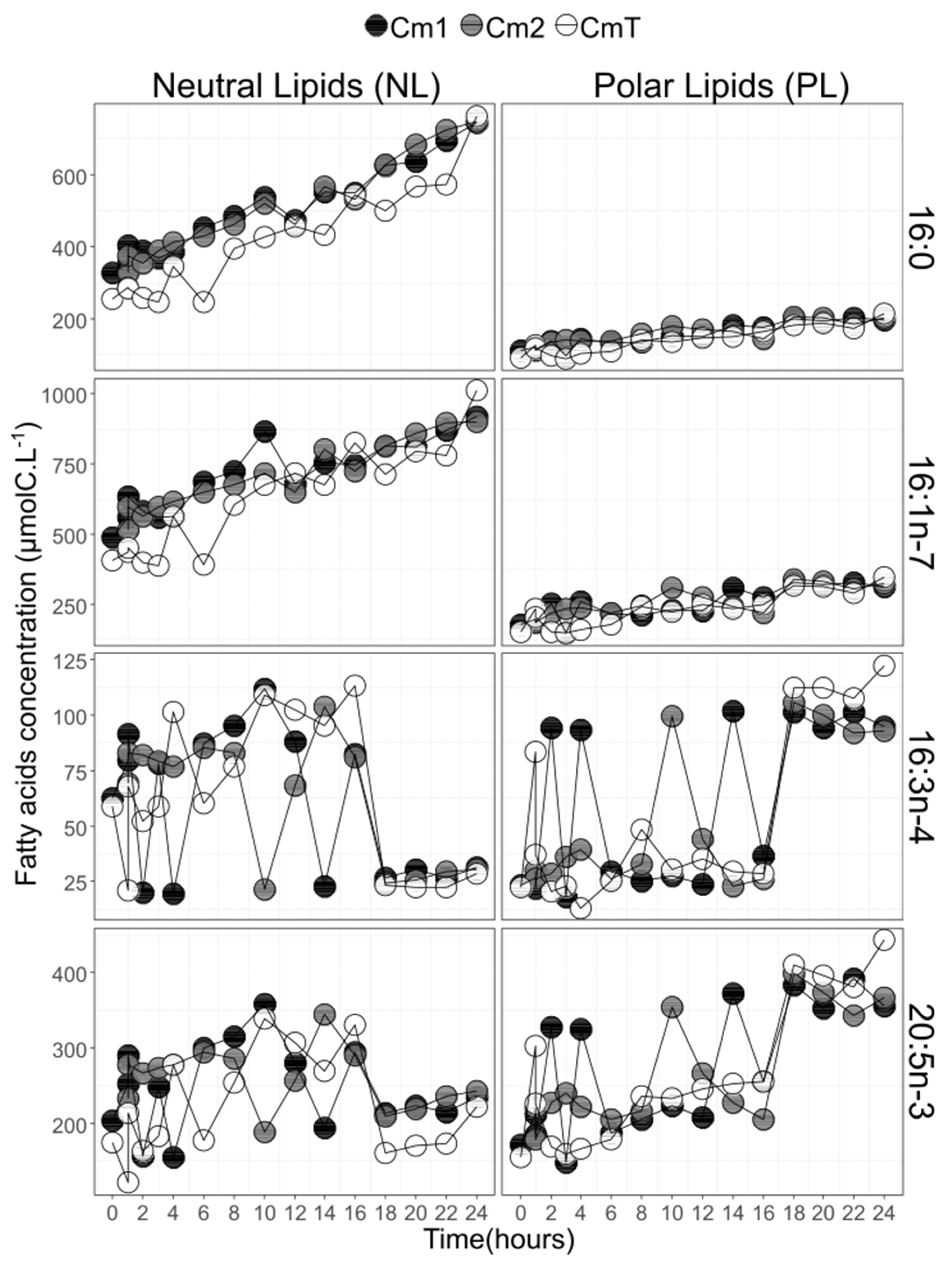
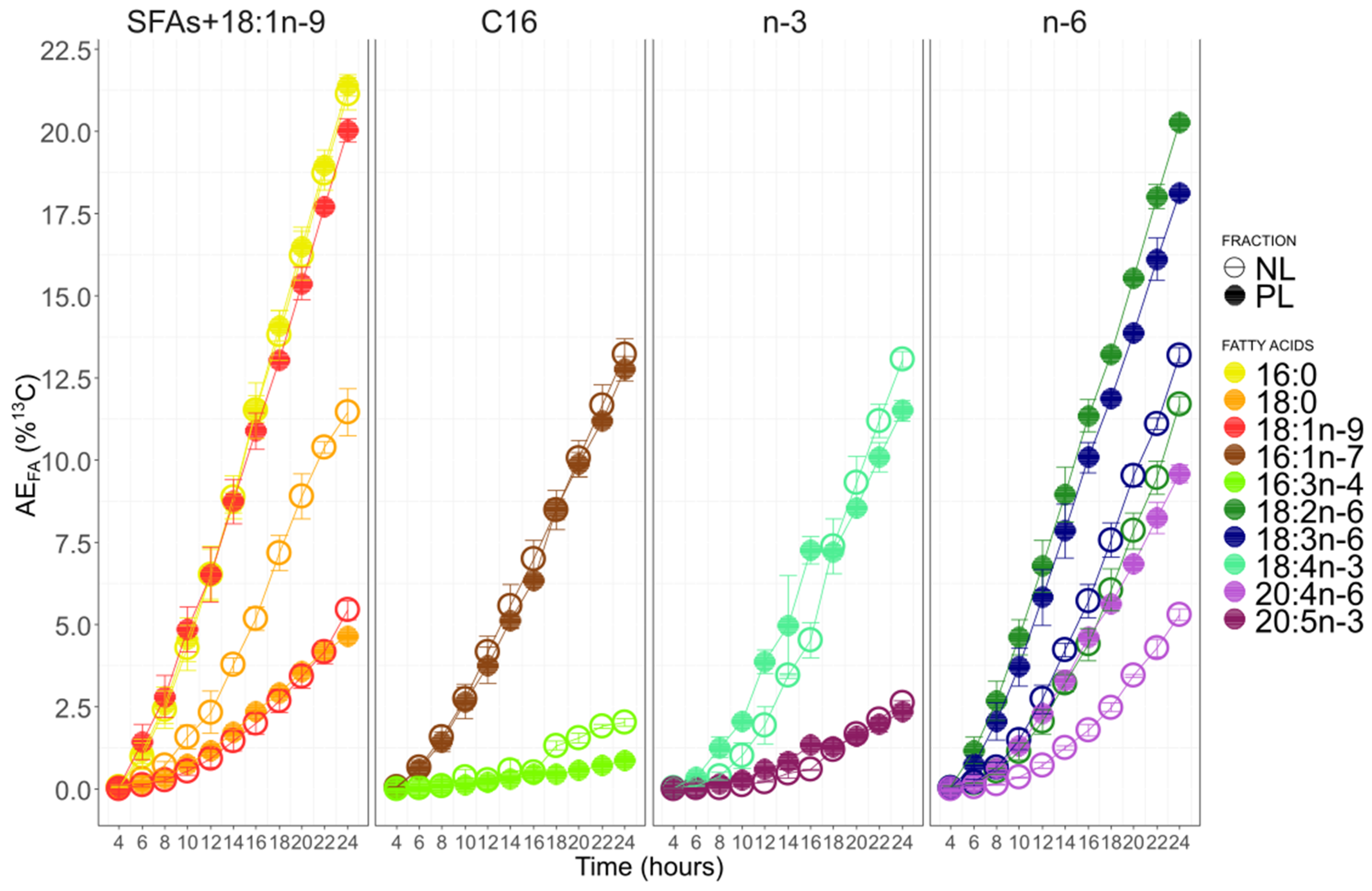
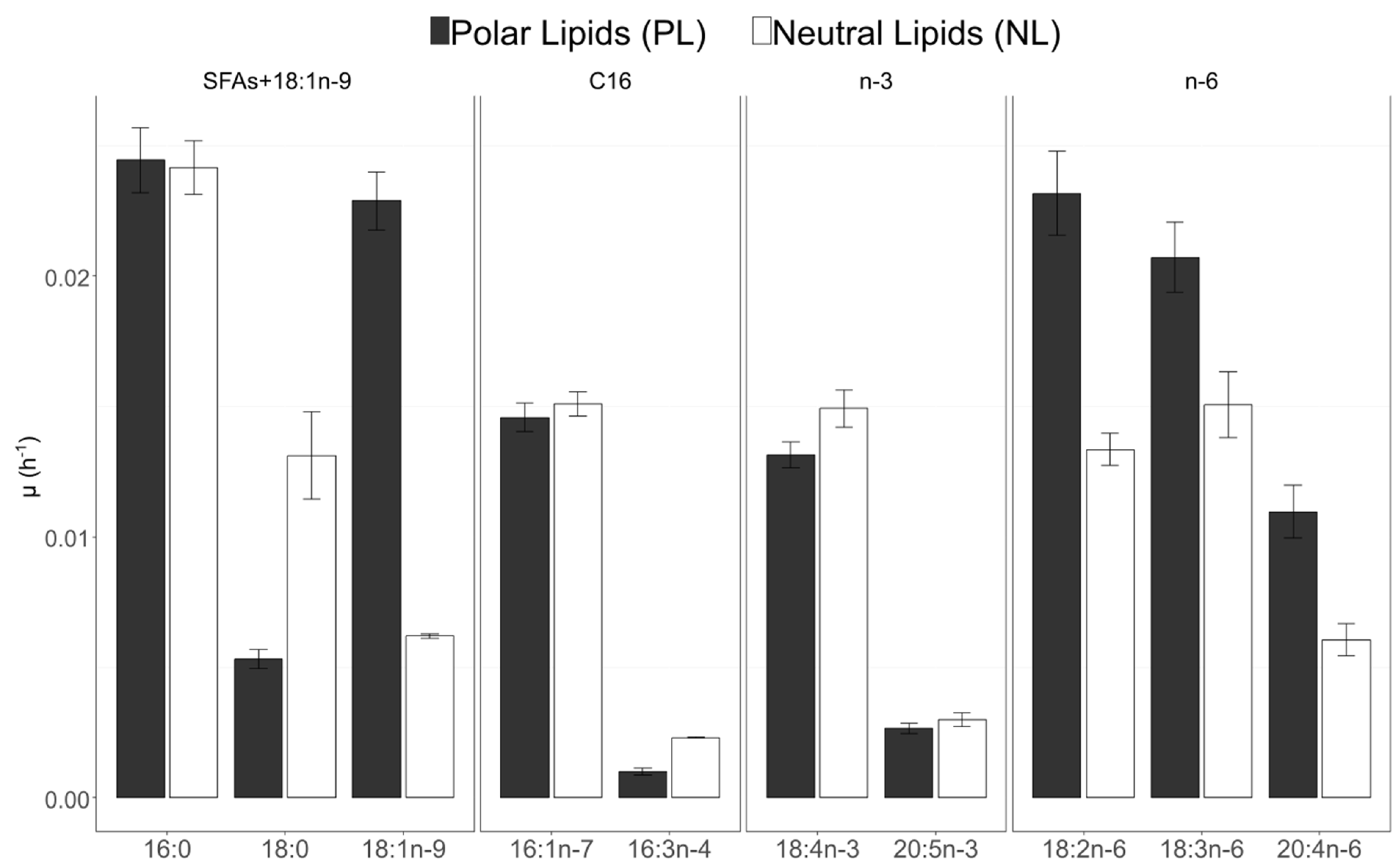
3.6. Fatty Acid 13C-Enrichment and Synthesis
4. Discussion
4.1. Growth and 13C Incorporation
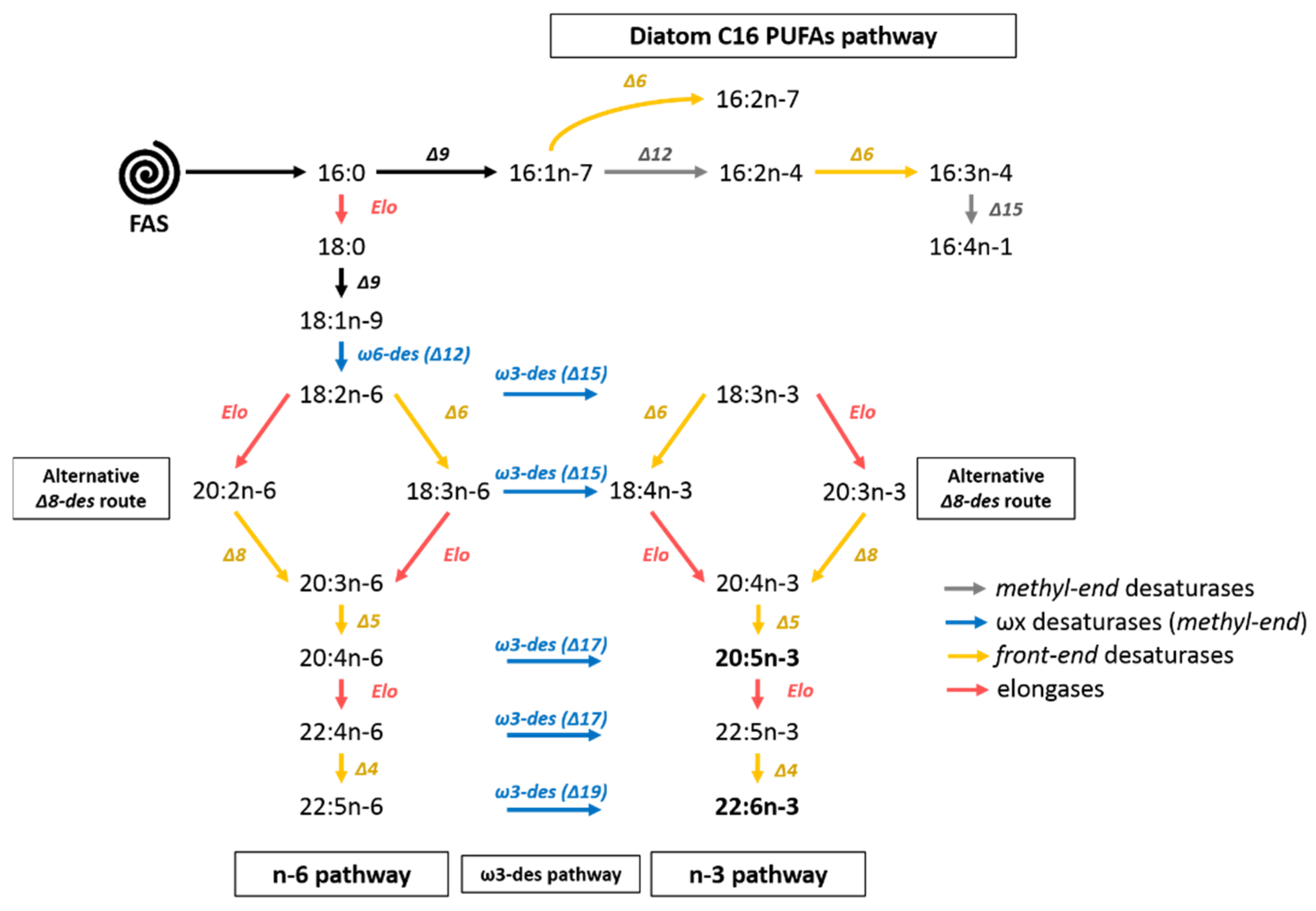
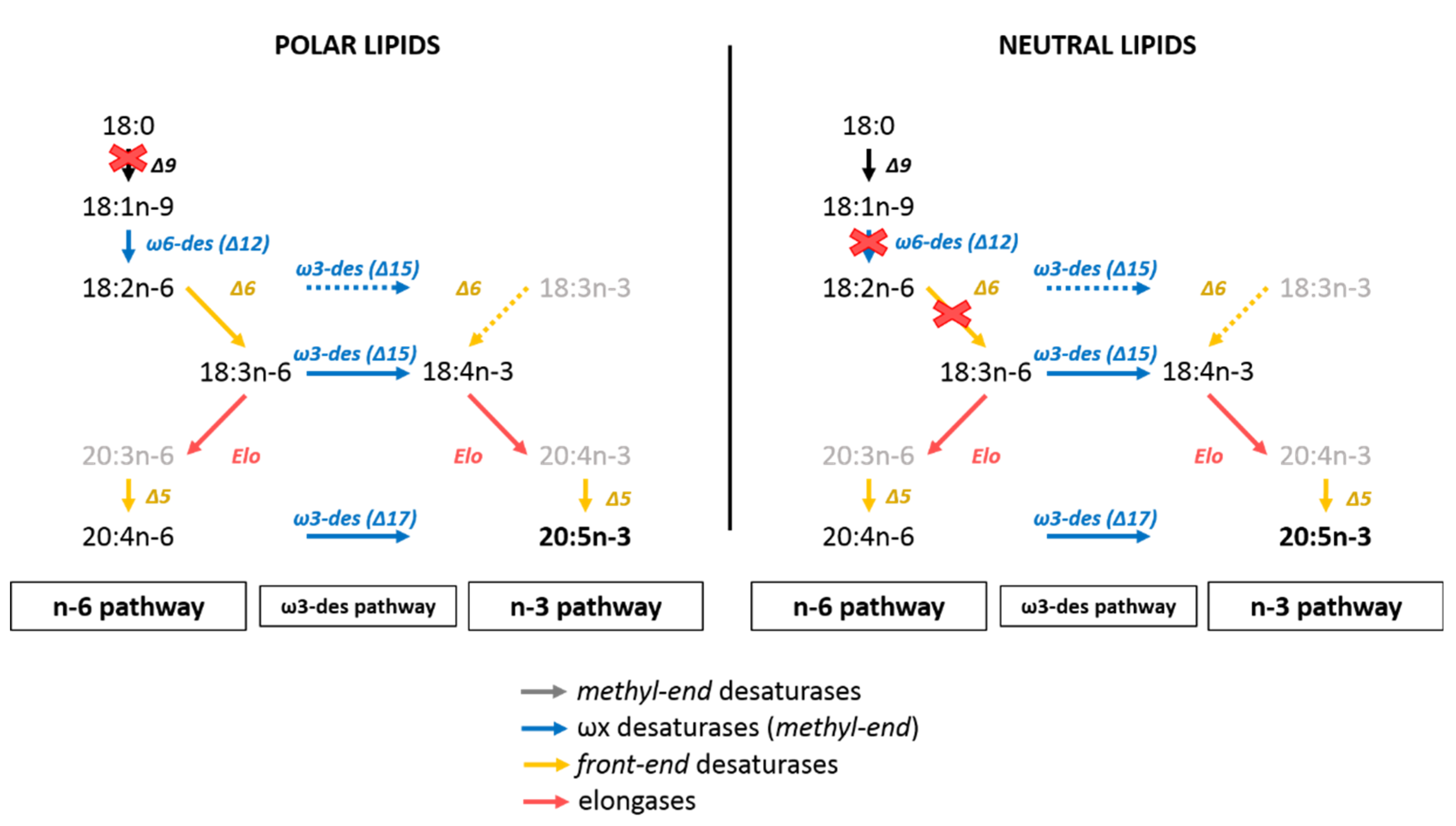
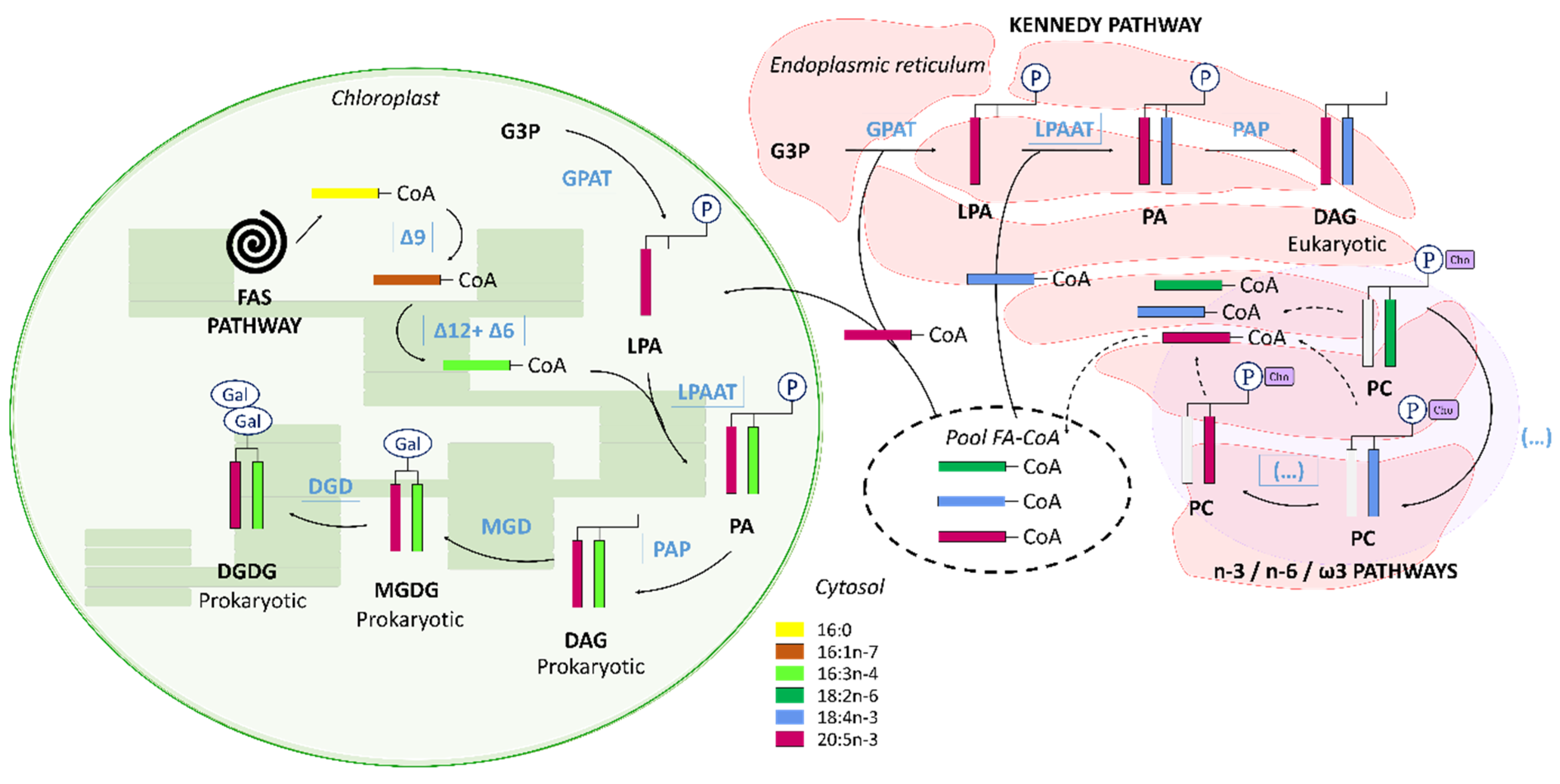
4.2. The C16 PUFAs Pathway in C. muelleri
4.3. EPA Synthesis Resulting from a Combination of n-3 and n-6 Pathways?
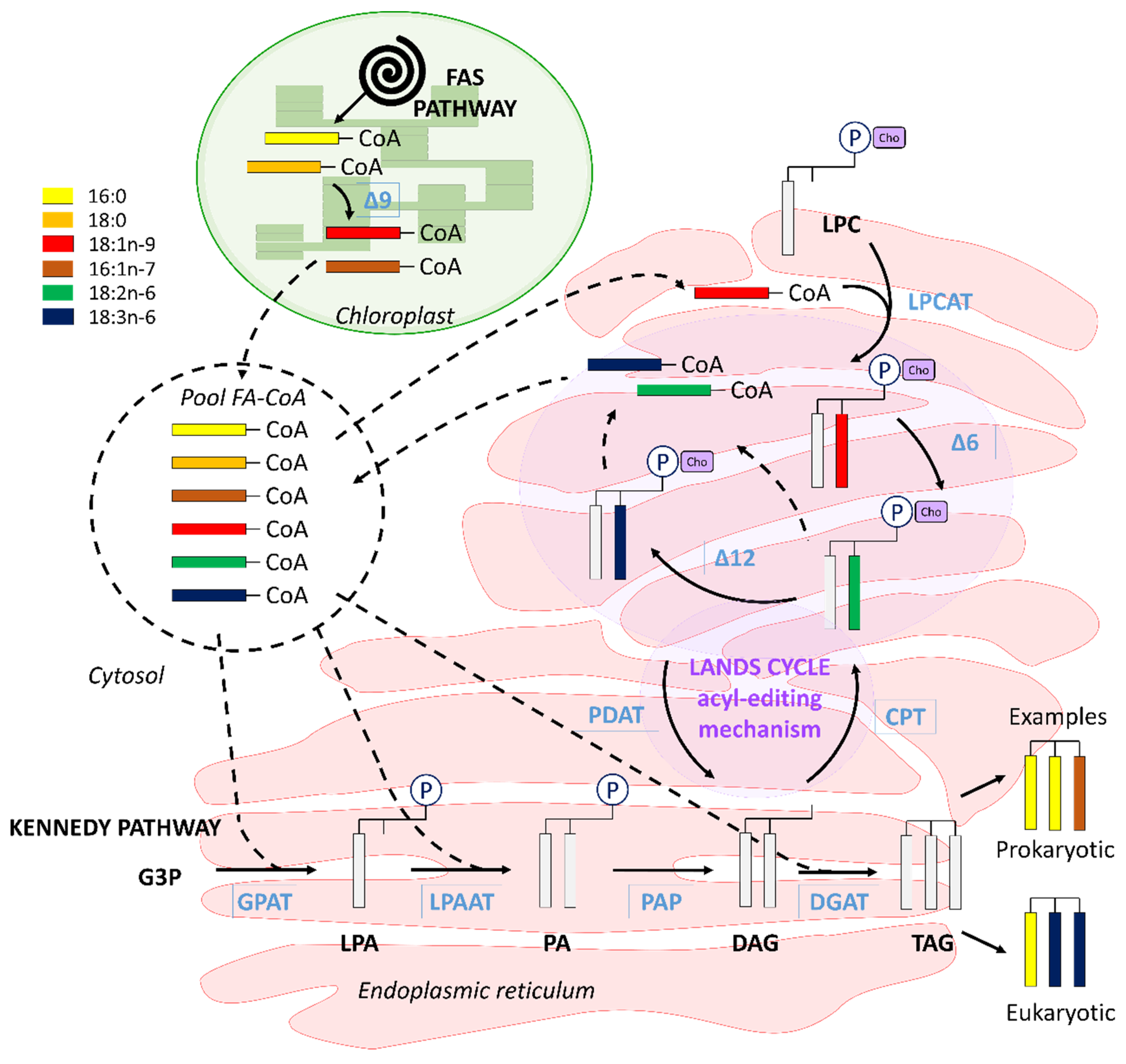
4.4. Interconnection between Polar and Neutral Pools in C. muelleri
4.5. C16 PUFAs and 20:5n-3: Linked on Glycerol Backbone in C. muelleri
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arts, M.T.; Ackman, R.G.; Holub, B.J. Essential fatty acids in aquatic ecosystems: A crucial link between diet and human health and evolution. Can. J. Fish. Aquat. Sci. 2001, 58, 122–137. [Google Scholar] [CrossRef]
- Mozzaffarian, D.; Wu, J.H.Y. (n-3) Fatty acids and cardiovascular health: Are effects of EPA and DHA shared or complementary? J. Nutr. 2012, 142, 614S–625S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, I.; Özogul, F.; Özogul, Y.; Regenstein, J.M. Marine bioactive compounds and their health benefits: A review: Importance of functional seafood. Compr. Rev. Food Sci. Food Saf. 2015, 14, 446–465. [Google Scholar] [CrossRef]
- Tocher, D.; Betancor, M.B.; Sprague, M.; Olsen, R.E.; Napier, J.A. Omega-3 long-chain polyunsaturated fatty acids, EPA and DHA: Bridging the gap between supply and demand. Nutrients 2019, 11, 89. [Google Scholar] [CrossRef] [Green Version]
- Dalsgaard, J.; John, M.S.; Kattner, G.; Müller-Navarra, D.; Hagen, W. Fatty acid trophic markers in the pelagic marine environment. Adv. Mar. Biol. 2003, 46, 225–340. [Google Scholar] [CrossRef]
- Dijkman, N.A.; Kromkamp, J.C. Phospholipid-derived fatty acids as chemotaxonomic markers for phytoplankton: Application for inferring phytoplankton composition. Mar. Ecol. Prog. Ser. 2006, 324, 113–125. [Google Scholar] [CrossRef] [Green Version]
- Reuss, N.; Poulsen, L. Evaluation of fatty acids as biomarkers for a natural plankton community. A field study of a spring bloom and a post-bloom period off West Greenland. Mar. Biol. 2002, 141, 423–434. [Google Scholar] [CrossRef]
- Henjes, J.; Assmy, P.; Klaas, C.; Verity, P.; Smetacek, V. Response of microzooplankton (protist and small copepods) to an iron-induced phytoplankton bloom in the Southern Ocean (EisenEx). Deep Sea Res. Part I 2007, 54, 363–384. [Google Scholar] [CrossRef]
- Hernando, M.; Schloss, I.R.; Almandoz, G.O.; Malanga, G.; Varela, D.E.; De Troch, M. Combined effects of temperature and salinity on fatty acid content and lipid damage in Antarctic phytoplankton. J. Exp. Mar. Biol. Ecol. 2018, 503, 120–128. [Google Scholar] [CrossRef]
- Nichols, P.D.; Skerratt, J.H.; Davidson, A.; Burton, H.; McMeekin, T.A. Lipid of culture Phaecystis pouchetii: Signature for food-web, biogeochemical and environmental studies in Antartica and the Southern Ocean. Phytochemistry 1991, 30, 3209–3214. [Google Scholar] [CrossRef]
- Becker, S.; Graeve, M.; Bischof, K. Photosynthesis and lipid composition of the Antarctic endemic rhodophyte Palmaria decipiens: Effects of changing light and temperature levels. Polar Biol. 2010, 33, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Christaki, U.; Obernosterer, I.; Van Wambeke, F.; Veldhuis, M.J.W.; Garcia, N.; Catala, P. Microbial food web structure in a naturally iron-fertilized area in the Southern Ocean (Kerguelen Plateau). Deep Sea Res. Part II Top. Stud. Oceanogr. 2008, 55, 706–719. [Google Scholar] [CrossRef]
- Corsolini, S.; Borghesi, N. A comparative assessment of fatty acids in Antarctic organisms from the Ross Sea: Occurrence and distribution. Chemosphere 2017, 174, 747–753. [Google Scholar] [CrossRef]
- Arao, T.; Yamada, M. Biosynthesis of polyunsaturated fatty acids in the marine diatom, Phaeodactylum tricornutum. Phytochemistry 1994, 35, 1177–1181. [Google Scholar] [CrossRef]
- Bell, M.V.; Tocher, D. Biosynthesis of polyunsaturated fatty acids in aquatic ecosystems: General pathways and new directions. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 211–239. [Google Scholar]
- Gurr, M.I.; Harwood, J.L.; Frayn, K.N. Fatty acids structure and metabolism: Fatty acids biosynthesis. In Lipid biochemistry; Science, B., Ed.; Blackwell Science: Hoboken, NJ, USA, 2002; pp. 21–59. [Google Scholar]
- Khozin-Goldberg, I. The Physiology of Microalgae-Lipid metabolism in microalgae. In The Physiology of Microalgae; Borowitska, M.A., Beardall, J., Raven, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 413–484. [Google Scholar]
- Metz, J.G.; Roessler, P.; Facciotti, D.; Levering, C.; Dittrich, F.; Lassner, M.; Valentine, R.; Lardizabal, K.; Domergue, F.; Yamada, A.; et al. Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science 2001, 293, 290–293. [Google Scholar] [CrossRef]
- Domergue, F.; Lerchl, J.; Zähringer, U.; Heinz, E. Cloning and functional characterization of Phaeodactylum tricornutum front-end desaturases involved in eicosapentaenoic acid biosynthesis. Eur. J. Biochem. 2002, 269, 4105–4113. [Google Scholar] [CrossRef]
- Qi, B.; Beaudoin, F.; Fraser, T.; Stobart, A.K.; Napier, J.A.; Lazarus, C.M. Identification of a cDNA encoding a novel C18-Δ9 polyunsaturated fatty acid-specific elongating activity from the docosahexaenoic acid (DHA)-producing microalga, Isochrysis galbana. Febs Lett. 2002, 510, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Robert, S.S.; Petrie, J.R.; Frampton, D.M.F.; Mansour, M.P.; Blackburn, S.I.; Nichols, P.D.; Green, A.G.; Singh, S.P. Isolation and characterization of genes from the marine microalga Pavlova salina encoding three front-end desaturases involved in docosahexaenoic acid biosynthesis. Phytochemistry 2007, 68, 785–796. [Google Scholar] [CrossRef]
- Sayanova, O.; Haslam, R.P.; Caleron, M.V.; Lopez, N.R.; Worthy, C.; Rooks, P.; Allen, M.J.; Napier, J.A. Identification and functional characterisation of genes encoding the omega-3 polyunsaturated fatty acid biosynthetic pathway from the coccolithophore Emiliania huxleyi. Phytochemistry 2011, 72, 594–600. [Google Scholar] [CrossRef]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Armenta, R.E.; Valentine, M.C. Single-cell oils as a source of omega-3 fatty acids: An overview of recent advances. J. Am. Oil Chem. Soc. 2013, 90, 167–182. [Google Scholar] [CrossRef]
- Ye, C.; Qiao, W.; Yu, X.; Ji, X.-J.; Huang, H.; Collier, J.L.; Liu, L. Reconstruction and analysis of the genome-scale metabolic model of Schizochytrium limacinum SR21 for docosahexaenoic acid production. BMC Genom. 2015, 16, 799–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauvermale, A.; Kuner, J.; Rosenzweig, B.; Guerra, D.; Diltz, S.; Metz, J.G. Fatty acid production in Schizochytrium sp.: Involvement of a polyunsaturated fatty acid synthase and a type I fatty acid synthase. Lipids 2006, 41, 739–747. [Google Scholar] [CrossRef] [PubMed]
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Resek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef] [Green Version]
- Caramujo, M.J.; Boschker, H.T.S.; Admiraal, W. Fatty acid profiles of algae mark the development and composition of harpacticoid copepods. Freshw. Biol. 2007, 53, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Hixson, S.M.; Arts, M.T. Climate warming is predicted to reduce omega-3, long-chain, polyunsaturated fatty acid production in phytoplankton. Glob. Chang. Biol. 2016, 22, 2744–2755. [Google Scholar] [CrossRef]
- de Carvalho, C.; Caramujo, M.J. Fatty acids as a tool to understand microbial diversity and their role in food webs of mediterranean temporary ponds. Molecules 2014, 19, 5570–5598. [Google Scholar] [CrossRef] [Green Version]
- Sayanova, O.; Mimouni, V.; Ulmann, L.; Morant, A.; Pasquet, V.; Schoefs, B.; Napier, J.A. Modulation of lipid biosynthesis by stress in diatoms. Philos. Trans. R. Soc. B 2017, 372, 14. [Google Scholar] [CrossRef] [Green Version]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Prog. Lipid Res. 2018, 70, 1–16. [Google Scholar] [CrossRef]
- Bates, P.D.; Browse, J. The significance of different diacylgycerol synthesis pathways on plant oil composition and bioengineering. Front. Plant. Sci. 2012, 3, 147. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, E.; Lupette, J. Relationship between acyl-lipid and sterol metabolisms in diatoms. Biochimie 2019, 169, 3–11. [Google Scholar] [CrossRef]
- Mülhroth, A.; Li, K.; Røkke, G.; Winge, P.; Olsen, Y.; Hohmann-Mariott, M.; Vadstein, O.; Bones, A. Pathways of lipid metabolism in marine algae, co-expression network, bottlenecks and candidate genes for enhanced production of EPA and DHA in species of Chromista. Mar. Drugs 2013, 11, 4662–4697. [Google Scholar] [CrossRef]
- Mus, F.; Toussaint, J.-P.; Cooksey, K.E.; Fields, M.W.; Gerlach, R.; Peyton, B.M.; Carlson, R.P. Physiological and molecular analysis of carbon source supplementation and pH stress-induced lipid accumulation in the marine diatom Phaeodactylum tricornutum. Appl. Microbiol. Biotechnol. 2013, 97, 3625–3642. [Google Scholar] [CrossRef] [Green Version]
- Lands, W.E.M. Metabolism of glycerolipides: A comparison of lecithin and triglycerides synthesis. J. Biol. Chem. 1958, 231, 883–888. [Google Scholar]
- Wang, L.; Shen, W.; Kazachkov, M.; Chen, G.; Chen, Q.; Carlsson, A.S.; Stymne, S.; Weselake, R.J.; Zou, J. Metabolic interactions between the Lands cycle and the Kennedy pathway of glycerolipid synthesis in Arabidopsis developing seeds. Plant. Cell 2012, 24, 4652–4669. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, N.M.D.; Parisien, A.; Wang, B.; Lan, C.Q. Enhancement of lipid production using biochemical, genetic and transcription factor engineering approaches. J. Biotechnol. 2009, 141, 31–41. [Google Scholar] [CrossRef]
- Gong, Y.; Wan, X.; Jiang, M.; Hu, C.; Hu, H.; Huang, F. Metabolic engineering of microorganisms to produce omega-3 very long chain polyunsaturated fatty acids. Prog. Lipid Res. 2014, 56, 19–35. [Google Scholar] [CrossRef]
- Xiong, W.; Liu, L.; Wu, C.; Yang, C.; Wu, Q. 13C-tracer and Gas Chromatography-Mass Spectrometry analyses reveal metabolic flux distribution in the oleaginous microalga Chlorella protothecoides. Plant. Physiol. 2010, 154, 1001–1011. [Google Scholar] [CrossRef] [Green Version]
- Cui, J.; Diao, J.; Sun, T.; Shi, M.; Liu, L.; Wang, F.; Chen, L.; Zhang, W. 13C Metabolic Flux Analysis of enhanced lipid accumulation modulated by ethanolamine in Crypthecodinium cohnii. Front. Microbiol. 2018, 9, 956. [Google Scholar] [CrossRef]
- Martzolff, A.; Cahoreau, E.; Cogne, G.; Peyriga, L.; Portais, J.-C.; Dechandol, E.; Le Grand, F.; Massou, S.; Gonçalves, O.; Pruvost, J.; et al. Photobioreactor design for isotopic non-stationnary 13C-metabolic flux analysis (INST 13C-MFA) under photoautotrophic conditions. Biotechnol. Bioeng. 2012, 109, 3030–3040. [Google Scholar] [CrossRef] [Green Version]
- Grosse, J.; Van Breugel, P.; Boschker, H.T.S. Tracing carbon fixation in phytoplankton-compound specific and total 13C incorporation rates: 13C uptake into macromolecules. Limnol. Oceanogr. Methods 2015, 13, 288–302. [Google Scholar] [CrossRef] [Green Version]
- Menzel, R.; Ngosong, C.; Ruess, L. Isotopologue profiling enables insights into dietary routing and metabolism of trophic biomarker fatty acids. Chemoecology 2017, 27, 101–114. [Google Scholar] [CrossRef]
- Wei, X.; Shi, B.; Koo, I.; Yin, X.; Lorkiewicz, P.; Suhail, H.; Rattan, R.; Giri, S.; McClain, C.J.; Zhang, X. Analysis of stable isotope assisted metabolomics data acquired by GC-MS. Anal. Chim. Acta 2017, 980, 25–32. [Google Scholar] [CrossRef]
- Lima, V.F.; Perez de Souza, L.; Williams, T.C.R.; Fernie, A.R.; Daloso, D.M. Gas chromatography-mass spectrometry based 13C labeling studies in plant metabolomics. In Plant Metabolomics; Antonio, C., Ed.; Springer: New York, NY, USA, 2018; Volume 1778, pp. 47–58. [Google Scholar]
- Rousseaux, C.; Gregg, W. Interannual variation in phytoplankton primary production at a global scale. Remote Sens. 2014, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- González-Fernández, C.; Toullec, J.; Lambert, C.; Le Goïc, N.; Seoane, M.; Moriceau, B.; Huvet, A.; Berchel, M.; Vincent, D.; Courcot, L.; et al. Do transparent exopolymeric particles (TEP) affect the toxicity of nanoplastics on Chaetoceros neogracile? Environ. Pollut. 2019, 250, 873–882. [Google Scholar] [CrossRef]
- Tracy, B.P.; Gaida, S.M.; Papoutsakis, E.T. Flow cytometry for bacteria: Enabling metabolic engineering, synthetic biology and the elucidation of complex phenotypes. Curr. Opin. Biotechnol. 2010, 21, 85–99. [Google Scholar] [CrossRef]
- Le Goïc, N.; Hegaret, H.; Boulais, M.; Béguel, J.-P.; Lambert, C.; Fabioux, C.; Soudant, P. Flow cytometric assessment of morphology, viability, and production of Reactive Oxygen Species of Crassostrea gigas oocytes. Application to toxic Dinoflagellate (Alexandrium minutum) exposure. Cytom. Part. A 2014, 85A, 1049–1056. [Google Scholar] [CrossRef] [Green Version]
- Seoane, M.; González-Fernández, C.; Soudant, P.; Huvet, A.; Esperanza, M.; Cid, A.; Paul-Pont, I. Polystyrene microbeads modulate the energy metabolism of the marine diatom Chaetoceros neogracile. Environ. Pollut. 2019, 251, 362–371. [Google Scholar] [CrossRef]
- Lelong, A.; Haberkorn, H.; Le Goïc, N.; Hégaret, H.; Soudant, P. A new insight into allelopathic effects of Alexandrium minutum on photosynthesis and respiration of the Diatom Chaetoceros neogracile revealed by photosynthetic-performance analysis and flow cytometry. Microb. Ecol. 2011, 62, 919–930. [Google Scholar] [CrossRef]
- Trull, T.W.; Davies, D.M.; Dehairs, F.; Cavagna, A.J.; Lasbleiz, M.; Laurenceau-Cornec, E.C.; Ovidio, F.; Planchon, F.; Leblanc, K.; Quéguiner, B.; et al. Chemometric perspectives on plankton community responses to natural iron fertilisation over and downstream of the Kerguelen Plateau in the Southern Ocean. Biogeosciences 2015, 12, 1029–1056. [Google Scholar] [CrossRef] [Green Version]
- Brooks, P.D.; Geilmann, H.; Werner, R.A.; Brand, W. Improved precision of coupled 13C and 15N measurements from single samples using an elemental analyser. Rapid Commun. Mass Spectrom. 2003, 12, 1924–1926. [Google Scholar] [CrossRef]
- United States Environmental Protection Agency, U.S.E. Methods for the Determination of Chemical Substances in Marine and Estuarine Environmental Matrices; US Environmental Protection Agency: Washington, DC, USA, 1997; p. 199. [Google Scholar]
- Assayag, N.; Rivé, K.; Ader, M.; Jézéquel, D.; Agrinier, P. Improved method for isotopic and quantitative analysis of dissolved inorganic carbon in natural water samples. Rapid Commun. Mass Spectrom. 2006, 20, 2243–2251. [Google Scholar] [CrossRef]
- Larsson, C.; Ahlstrøm, Ø.; Junghans, P.; Jensen, R.B.; Blache, D.; Tauson, A.-H. The oral [13C]bicarbonate technique for measurement of short-term energy expenditure of sled dogs and their physiological response to diets with different fat:carbohydrate ratios. J. Nutr. Sci. 2015, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Grand, F.; Soudant, P.; Siah, A.; Tremblay, P.; Marty, Y.; Kraffe, E. Disseminated Neoplasia in the soft-shell clam Mya arenaria: Membrane lipid composition and functional parameters of circulating cells. Lipids 2014, 49, 807–818. [Google Scholar] [CrossRef] [PubMed]
- Mathieu-Resuge, M.; Schaal, G.; Kraffe, E.; Corvaisier, R.; Lebeau, O.; Lluch-Costa, S.E.; Salgado García, R.L.; Kainz, M.J.; Le Grand, F. Different particle sources in a bivalve species of a coastal lagoon: Evidence from stable isotopes, fatty acids, and compound-specific stable isotopes. Mar. Biol. 2019, 166, 89–101. [Google Scholar] [CrossRef]
- Volkman, J.K.; Jeffrey, S.W.; Nichols, P.D.; Rogers, G.I.; Garland, C.D. Fatty acid and lipid composition of 10 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1989, 128, 219–240. [Google Scholar] [CrossRef]
- Hatate, H.; Ohgai, M.; Murase, N.; Miyake, N.; Suzuki, N. Accumulation of fatty acids in Chaetoceros gracilis (Bacillariophyceae) during stationary growth phase. Fish. Sci. 1998, 64, 578–581. [Google Scholar] [CrossRef] [Green Version]
- Vazhappilly, R.; Chen, F. Eicosapentaenoic acid and docosahexaenoic acid production potential of microalgae and their heterotrophic growth. J. Am. Oil Chem. Soc. 1998, 75, 393–397. [Google Scholar] [CrossRef]
- Pernet, F.; Tremblay, R.; Demers, E.; Roussy, M. Variation of lipid class and fatty acid composition of Chaetoceros muelleri and Isochrysis sp. grown in a semicontinuous system. Aquaculture 2003, 221, 393–406. [Google Scholar] [CrossRef]
- Vaezi, R.; Napier, J.A.; Sayanova, O. Identification and functional characterization of genes encoding omega-3 polyunsaturated fatty acid biosynthetic activities from unicellular microalgae. Mar. Drugs 2013, 11, 5116–5129. [Google Scholar] [CrossRef] [Green Version]
- Domergue, F.; Spiekermann, P.; Lerchl, J.; Beckmann, C.; Kilian, O.; Kroth, P.G.; Boland, W.; Zahringer, U.; Heinz, E. New insight into Phaeodactylum tricornutum fatty acid metabolism. Cloning and functional characterization of plastidial and microsomal Δ12-fatty acid desaturases. Plant. Physiol. 2003, 131, 1648–1660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desvillettes, C.; Bec, A. Formation and transfer of fatty acids in aquatic microbial food webs: Role of heterotrophic protists. In Lipids in Aquatic Ecosystems; Arts, M.T., Kainz, M., Eds.; Springer: New York, NY, USA, 2009; pp. 25–42. [Google Scholar]
- d’Ippolito, G.; Cutignano, A.; Briante, R.; Febbraio, F.; Cimino, G.; Fontana, A. New C16 fatty-acid-based oxylipin pathway in the marine diatom Thalassiosira rotula. Org. Biomol. Chem. 2005, 3, 4065. [Google Scholar] [CrossRef]
- d’Ippolito, G.; Tucci, S.; Cutignano, A.; Romano, G.; Cimino, G.; Miralto, A.; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom Skeletonema costatum. Biochim. Et Biophys. Acta (Bba) Mol. Cell Biol. Lipids 2004, 1686, 100–107. [Google Scholar] [CrossRef]
- Cutignano, A.; d’Ippolito, G.; Romano, G.; Lamari, N.; Cimino, G.; Febbraio, F.; Nucci, R.; Fontana, A. chloroplastic glycolipids fuel aldehyde biosynthesis in the marine diatom Thalassiosira rotula. ChemBioChem 2006, 7, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Lebl, T.; Yan, L.; Smith, V.J. Isolation and structural characterisation of two antibacterial free fatty acids from the marine diatom, Phaeodactylum tricornutum. Appl. Microbiol. Biotechnol. 2008, 81, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Khozin-Goldberg, I.; Iskandarov, U.; Cohen, Z. LC-PUFA from photosynthetic microalgae: Occurrence, biosynthesis, and prospects in biotechnology. Appl. Microbiol. Biotechnol. 2011, 91, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.-Y.; Chen, F. Heterotrophic production of eicosapentaenoic acid by microalgae. Biotechnol. Adv. 2003, 21, 273–294. [Google Scholar] [CrossRef]
- Fan, J.; Yan, C.; Xu, C. Phospholipid:diacylglycerol acyltransferase-mediated triacylglycerol biosynthesis is crucial for protection against fatty acid-induced cell death in growing tissues of Arabidopsis. Plant. J. 2013, 76, 930–942. [Google Scholar] [CrossRef]
- Fan, J.; Yan, C.; Zhang, X.; Xu, C. Dual role for phospholipid:diacylglycerol acyltransferase: Enhancing fatty acid synthesis and diverting fatty acids from membrane lipids to triacylglycerol in Arabidopsis leaves. Plant. Cell 2013, 25, 3506–3518. [Google Scholar] [CrossRef] [Green Version]
- Dolch, L.-J.; Maréchal, E. Inventory of fatty acid desaturases in the pennate diatom Phaeodactylum tricornutum. Mar. Drugs 2015, 13, 1317–1339. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-Y.; Lu, Y.; Zheng, J.-W.; Liu, W.-D. Biochemical and genetic engineering of diatoms for polyunsaturated fatty acid biosynthesis. Mar. Drugs 2014, 12, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.T.; Zendejas, F.J.; Lane, P.D.; Gaucher, S.; Simmons, B.A.; Lane, T.W. Triacylglycerol accumulation and profiling in the model diatoms Thalassiosira pseudonana and Phaeodactylum tricornutum (Baccilariophyceae) during starvation. J. Appl. Phycol. 2009, 21, 669–681. [Google Scholar] [CrossRef]
- Danielewicz, M.A.; Anderson, L.A.; Franz, A.K. Triacylglycerol profiling of marine microalgae by mass spectrometry. J. Lipid Res. 2011, 52, 2101–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roughan, P.G.; Holland, R.; Slack, C.R. The role of chloroplasts and microsomal fractions in polar-lipid synthesis from [1-14C]acetate by cell-free preparations from spinach (Spinacia oleracea) leaves. Biochem. J. 1980, 188, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinz, E.; Roughan, P.G. Similarities and differences in lipid metabolism of chloroplast isolated from 18:3 and 16:3 plants. Plant. Physiol. 1983, 72, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Benning, C. Chloroplast lipid synthesis and lipid trafficking through ER–plastid membrane contact sites. Biochem. Soc. Trans. 2012, 40, 457–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Description | Nature | Analysis | δ13C (‰) | SD |
|---|---|---|---|---|
| IAEA-CH6 | Sucrose (C12H22O11) | 13C-POC | −10.45 | 0.03 |
| IAEA-600 | Caffeine (C8H10N4O2) | 13C-POC | −27.77 | 0.04 |
| Acetanilide | Acetanilide (C8H9NO) | 13C-POC | 29.53 | 0.01 |
| CA21 (in-house std) | Calcium carbonate (CaCO3) | 13C-DIC | +1.47 | |
| Na2CO3 (in-house std) | Sodium carbonate | 13C-DIC | −6.88 | |
| NaHCO3 (in-house std) | Sodium bicarbonate | 13C-DIC | −5.93 |
| Group | Fatty Acids Concerned |
|---|---|
| C16 PUFAs pathway | 16:2n-7/16:2n-4/16:3n-4/16:4n-1 |
| n-3 pathway | 18:3n-3/18:4n-3/20:4n-3/20:5n-3/22:5n-3/22:6n-3 |
| n-6 pathway | 18:2n-6/18:3n-6/20:2n-6/20:3n-6/20:4n-6/22:2n-6/22:5n-6 |
| FAS and post FAS | 16:0/18:0/16:1n-7/18:1n-9 |
| Bacterial FA | 15:0/17:0/iso15:0/iso17:0/17:1n-7 |
| FL1-FDA ACTIVE (%) | FL1-SYTOX ALIVE (%) | FL1-BODIPY (a.u) | FL3 (a.u) | SSC (a.u) | FSC (a.u) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 93 | ± | 4 | 99 | ± | 1 | 160 | ± | 20 | 263 | ± | 2 | 48 | ± | 1 | 149 | ± | 0 |
| 0.5 | 90 | ± | 4 | 99 | ± | 1 | 215 | ± | 23 | 274 | ± | 20 | 50 | ± | 2 | 152 | ± | 1 |
| 1 | 93 | ± | 3 | 99 | ± | 0 | 233 | ± | 31 | 276 | ± | 18 | 56 | ± | 10 | 151 | ± | 2 |
| 2 | 93 | ± | 2 | 100 | ± | 0 | 209 | ± | 11 | 275 | ± | 19 | 57 | ± | 10 | 153 | ± | 1 |
| 3 | 93 | ± | 2 | 100 | ± | 0 | 245 | ± | 25 | 263 | ± | 3 | 53 | ± | 2 | 152 | ± | 1 |
| 4 | 92 | ± | 1 | 98 | ± | 2 | 248 | ± | 26 | 260 | ± | 2 | 53 | ± | 1 | 152 | ± | 2 |
| 6 | 92 | ± | 2 | 98 | ± | 2 | 194 | ± | 17 | 260 | ± | 2 | 50 | ± | 2 | 152 | ± | 1 |
| 8 | 90 | ± | 1 | 98 | ± | 3 | 248 | ± | 17 | 260 | ± | 2 | 53 | ± | 2 | 153 | ± | 3 |
| 10 | 90 | ± | 1 | 98 | ± | 2 | 157 | ± | 2 | 262 | ± | 3 | 52 | ± | 3 | 153 | ± | 1 |
| 12 | 89 | ± | 4 | 99 | ± | 1 | 163 | ± | 26 | 262 | ± | 4 | 51 | ± | 2 | 154 | ± | 0 |
| 14 | 89 | ± | 2 | 97 | ± | 4 | 276 | ± | 24 | 261 | ± | 4 | 53 | ± | 0 | 156 | ± | 1 |
| 16 | 86 | ± | 3 | 97 | ± | 3 | 284 | ± | 9 | 261 | ± | 4 | 52 | ± | 2 | 155 | ± | 1 |
| 18 | 87 | ± | 2 | 98 | ± | 3 | 163 | ± | 11 | 259 | ± | 6 | 52 | ± | 3 | 156 | ± | 3 |
| 20 | 88 | ± | 4 | 99 | ± | 2 | 308 | ± | 9 | 258 | ± | 6 | 56 | ± | 1 | 155 | ± | 1 |
| 22 | 87 | ± | 4 | 96 | ± | 5 | 269 | ± | 16 | 258 | ± | 5 | 55 | ± | 2 | 154 | ± | 1 |
| 24 | 88 | ± | 3 | 99 | ± | 2 | 239 | ± | 5 | 257 | ± | 6 | 54 | ± | 1 | 154 | ± | 2 |
| Neutral Lipids | Polar Lipids | |||||||
|---|---|---|---|---|---|---|---|---|
| Cm1 | Cm2 | Cm1 | Cm2 | |||||
| Fatty acid B/Fatty acid A | Mean | SD | Mean | SD | Mean | SD | Mean | SD |
| 18:1n-9/18:0 | 0.37 | 0.03 | 0.41 | 0.05 | 5.72 | 2.16 | 5.89 | 2.14 |
| 18:2n-6/18:1n-9 | 2.25 | 0.15 | 2.03 | 0.27 | 1.00 | 0.06 | 0.98 | 0.07 |
| 18:3n-6/18:2n-6 | 1.23 | 0.14 | 1.26 | 0.11 | 0.83 | 0.10 | 0.85 | 0.08 |
| 20:4n-6/18:3n-6 | 0.33 | 0.05 | 0.29 | 0.09 | 0.43 | 0.09 | 0.41 | 0.10 |
| 20:5n-3/18:4n-3 | 0.18 | 0.05 | 0.13 | 0.05 | 0.18 | 0.02 | 0.15 | 0.04 |
| 18:4n-3/18:3n-6 | 0.77 | 0.20 | 0.82 | 0.20 | 0.62 | 0.06 | 0.61 | 0.08 |
| 20:5n-3/20:4n-6 | 0.40 | 0.09 | 0.37 | 0.12 | 0.26 | 0.03 | 0.23 | 0.04 |
| 16:1n-7/16:0 | 0.63 | 0.03 | 0.63 | 0.01 | 0.58 | 0.03 | 0.57 | 0.03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Remize, M.; Planchon, F.; Loh, A.N.; Le Grand, F.; Bideau, A.; Le Goic, N.; Fleury, E.; Miner, P.; Corvaisier, R.; Volety, A.; et al. Study of Synthesis Pathways of the Essential Polyunsaturated Fatty Acid 20:5n-3 in the Diatom Chaetoceros Muelleri Using 13C-Isotope Labeling. Biomolecules 2020, 10, 797. https://doi.org/10.3390/biom10050797
Remize M, Planchon F, Loh AN, Le Grand F, Bideau A, Le Goic N, Fleury E, Miner P, Corvaisier R, Volety A, et al. Study of Synthesis Pathways of the Essential Polyunsaturated Fatty Acid 20:5n-3 in the Diatom Chaetoceros Muelleri Using 13C-Isotope Labeling. Biomolecules. 2020; 10(5):797. https://doi.org/10.3390/biom10050797
Chicago/Turabian StyleRemize, Marine, Frédéric Planchon, Ai Ning Loh, Fabienne Le Grand, Antoine Bideau, Nelly Le Goic, Elodie Fleury, Philippe Miner, Rudolph Corvaisier, Aswani Volety, and et al. 2020. "Study of Synthesis Pathways of the Essential Polyunsaturated Fatty Acid 20:5n-3 in the Diatom Chaetoceros Muelleri Using 13C-Isotope Labeling" Biomolecules 10, no. 5: 797. https://doi.org/10.3390/biom10050797