An Extract of Transgenic Senna obtusifolia L. hairy roots with Overexpression of PgSS1 Gene in Combination with Chemotherapeutic Agent Induces Apoptosis in the Leukemia Cell Line

 , ,
, ,  , and
, and
Abstract
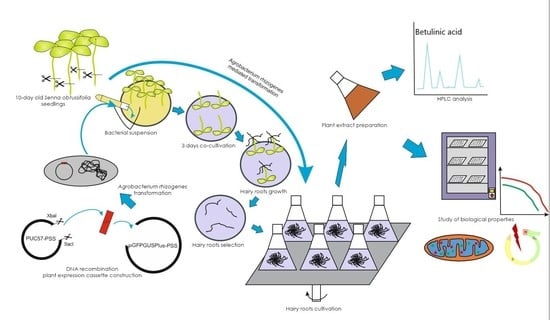
:
1. Introduction
2. Materials and Methods
2.1. Plant Expression Vector Construction
2.2. Agrobacterium Rhizogenes Competent Cells Preparation
2.3. Agrobacterium Rhizogenes Competent Cell Transformation
2.4. Plant Materials
2.5. Establishment of Plant In Vitro Cultures
2.6. Plant Material Transformation
2.7. PgSS1, GFP and A4 Hairy Roots Selection
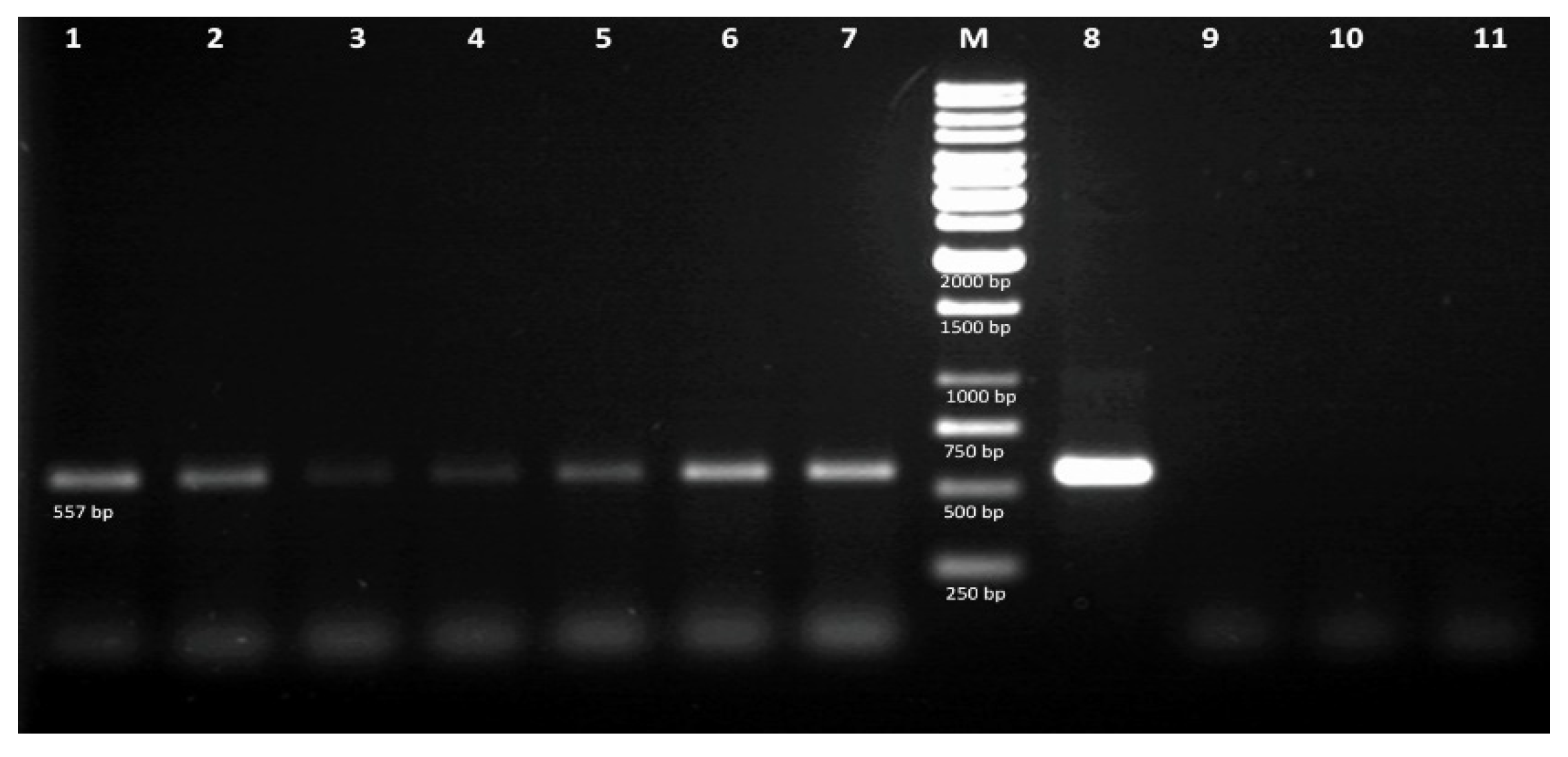
2.8. Molecular Analysis of Hairy Roots by PCR
2.9. β-Glucuronidase (GUSPlus) Histochemical Assay
2.10. GFP Expression Detection
2.11. Plant Extract Preparation
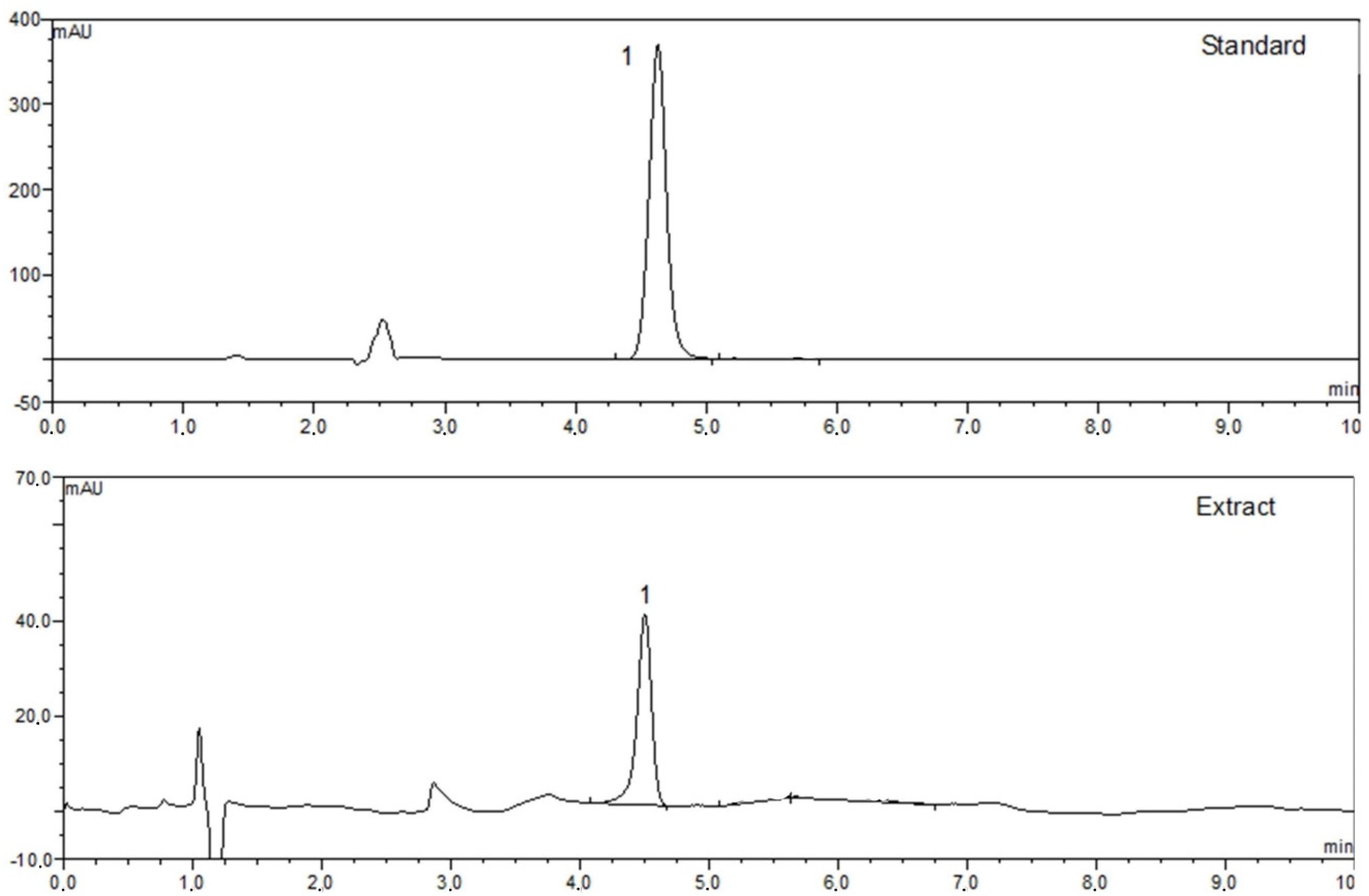
2.12. HPLC Analysis
2.13. Cell Culture
2.14. Cytotoxic Activity
2.15. Gene Expression
2.16. Cell Cycle and Apoptosis
2.17. Mitochondrial Membrane Potential (ΔΨm)
2.18. Statistical Analysis
3. Results
3.1. Generation of Transgenic Hairy Root Lines from S. obtusifolia
3.2. PCR analysis of Transgenic Hairy Roots
3.3. GUSPlus and GFP Detection
3.4. HPLC Analysis
3.5. Cytotoxic Activity of SOPSS1 and SOPSS2 S. obtusifolia Hairy Root Extracts in NALM6 Leukemia Cell Line
3.6. Gene Expression Analysis
3.7. Effects of SOPSS Transgenic Hairy Root Extracts (Alone or Combination with Doxorubicin) on Apoptosis and Cell Cycle in NALM6 Cells
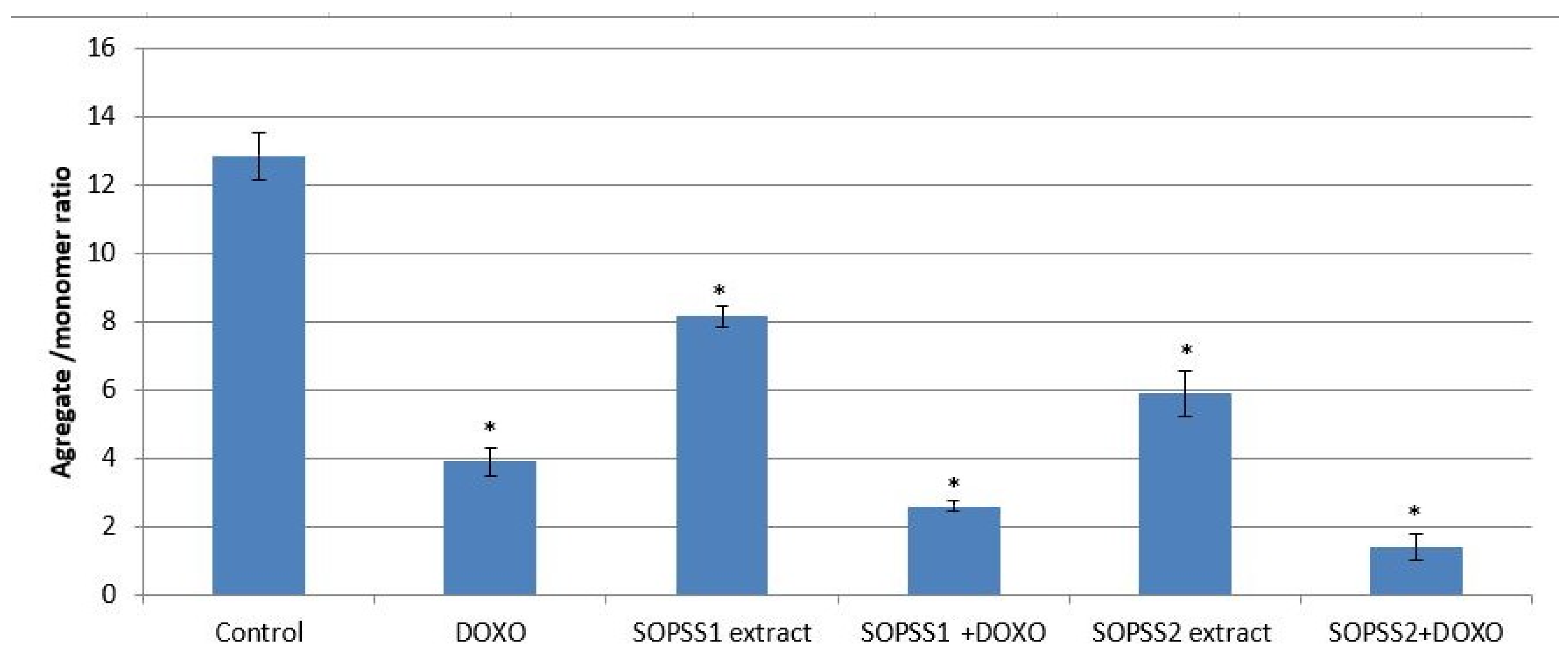
3.8. Effect of Transgenic SOPSS Root Extracts (Alone or Combination with Doxorubicin) on Mitochondrial Membrane Potential of NALM6 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019, 69, 363–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majolo, F.; de Oliveira Becker Delwing, L.K.; Marmitt, D.J.; Bustamante-Filho, I.C.; Goettert, M.I. Medicinal plants and bioactive natural compounds for cancer treatment: Important advances for drug discovery. Phytochem. Lett. 2019, 31, 196–207. [Google Scholar] [CrossRef]
- Shukla, S.; Mehta, A. Anticancer potential of medicinal plants and their phytochemicals: A review. Rev. Bras. Bot. 2015, 38, 199–210. [Google Scholar] [CrossRef]
- Yang, L.; Yang, C.; Li, C.; Zhao, Q.; Liu, L.; Fang, X.; Chen, X.Y. Recent advances in biosynthesis of bioactive compounds in traditional Chinese medicinal plants. Sci. Bull. 2016, 61, 3–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, H. Brilliant future of phytomedicines in the light of latest technological developments. J. Phytopharm. 2015, 4, 58–60. [Google Scholar]
- Moraes, D.F.C.; de Mesquita, L.S.S.; do Amaral, F.M.M.; de Sousa Ribeiro, M.N.; Malik, S. Anticancer drugs from plants. In Biotechnology and Production of Anti-Cancer Compounds; Springer International Publishing: New York, NY, USA, 2017; pp. 121–142. ISBN 9783319538808. [Google Scholar]
- de Azevedo, F.P.; de Souza Conceição, A. (Leguminosae: Caesalpinioideae) na serra geral de licínio de almeida, Bahia, Brasil. Acta Sci. Biol. Sci. 2017, 39, 95–112. [Google Scholar] [CrossRef] [Green Version]
- Rakib, M.; Ansari, A.A.; Arif, M.; Ahmad, A. A review of phytochemical and biological studies on cassia obtusifolia linn. In folklore medicine of eastern Uttar Pradesh. World J. Pharm. Res. 2018, 7, 191–201. [Google Scholar]
- Yagi, S.M.; el Tigani, S.; Adam, S.E.I. Toxicity of Senna obtusifolia fresh and fermented leaves (kawal), Senna alata leaves and some products from Senna alata on rats. Phytother. Res. 1998, 12, 324–330. [Google Scholar] [CrossRef]
- Sudi, I.Y.; Ksgbiya, D.M.; Muluh, E.K.; Clement, A. Nutritional and phytochemical screening of Senna obtusifolia indigenous to Mubi, Nigeria. Adv. Appl. Sci. Res. 2011, 2, 432–437. [Google Scholar]
- Hordyjewska, A.; Ostapiuk, A.; Horecka, A.; Kurzepa, J. Betulin and betulinic acid: Triterpenoids derivatives with a powerful biological potential. Phytochem. Rev. 2019, 18, 929–951. [Google Scholar] [CrossRef] [Green Version]
- Isah, T.; Umar, S.; Mujib, A.; Sharma, M.P.; Rajasekharan, P.E.; Zafar, N.; Frukh, A. Secondary metabolism of pharmaceuticals in the plant in vitro cultures: Strategies, approaches, and limitations to achieving higher yield. Plant Cell Tissue Organ Cult. 2018, 132, 239–265. [Google Scholar] [CrossRef]
- Kowalczyk, T.; Wieczfinska, J.; Skała, E.; Śliwiński, T.; Sitarek, P. Transgenesis as a tool for the efficient production of selected secondary metabolites from in vitro plant cultures. Plants 2020, 9, 132. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Ko, K. Production of recombinant anti-cancer vaccines in plants. Biomol. Ther. 2017, 25, 345–353. [Google Scholar] [CrossRef] [Green Version]
- Chahel, A.A.; Zeng, S.; Yousaf, Z.; Liao, Y.; Yang, Z.; Wei, X.; Ying, W. Plant-specific transcription factor LrTCP4 enhances secondary metabolite biosynthesis in Lycium ruthenicum hairy roots. Plant Cell Tissue Organ Cult. 2019, 136, 323–337. [Google Scholar] [CrossRef]
- Mendoza-Poudereux, I.; Muñoz-Bertomeu, J.; Navarro, A.; Arrillaga, I.; Segura, J. Enhanced levels of S-linalool by metabolic engineering of the terpenoid pathway in spike lavender leaves. Plant Cell Tissue Organ Cult. 2014, 23, 136–144. [Google Scholar] [CrossRef]
- Häkkinen, S.T.; Moyano, E.; Cusidó, R.M.; Oksman-Caldentey, K.M. Exploring the metabolic stability of engineered hairy roots after 16 years maintenance. Front. Plant Sci. 2016, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sitarek, P.; Kowalczyk, T.; Rijo, P.; Białas, A.J.; Wielanek, M.; Wysokińska, H.; Garcia, C.; Toma, M.; Śliwiński, T.; Skała, E. Over-Expression of AtPAP1 transcriptional factor enhances phenolic acid production in transgenic roots of Leonurus sibiricus L. and their biological activities. Mol. Biotechnol. 2018, 60, 74–82. [Google Scholar] [CrossRef]
- Ni, X.; Wen, S.; Wang, X.; Wang, W.; Xu, H.; Kai, G. Enhancement of camptothecin production in Camptotheca acuminata hairy roots by overexpressing ORCA3 gene. J. App. Pharm. Sci. 2011, 1, 85–88. [Google Scholar]
- Murashige, T.; Skoog, F. Murashige1962 Revised.Pdf. Physiol. Plant. 1962, 15, 474–497. [Google Scholar]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: B-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. Embo J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef] [PubMed]
- Pai, S.R.; Nimbalkar, M.S.; Pawar, N.V.; Dixit, G.B. Optimization of extraction techniques and quantification of betulinic Acid (BA) by RP-HPLC method from Ancistrocladus heyneanus Wall. Ex Grah. Ind. Crop. Prod. 2011, 34, 1458–1464. [Google Scholar] [CrossRef]
- Sitarek, P.; Synowiec, E.; Kowalczyk, T.; Śliwiński, T.; Skała, E. An in vitro estimation of the cytotoxicity and genotoxicity of root extract from Leonurus sibiricus L. overexpressing AtPAP1 against different cancer cell lines. Molecules 2018, 23, 2049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dehghan, E.; Reed, D.W.; Covello, P.S.; Hasanpour, Z.; Palazon, J.; Oksman-Caldentey, K.M.; Ahmadi, F.S. Genetically engineered hairy root cultures of Hyoscyamus senecionis and H. muticus: Ploidy as a promising parameter in the metabolic engineering of tropane alkaloids. Plant Cell Rep. 2017, 36, 1615–1626. [Google Scholar] [CrossRef]
- Wei, T.; Gao, Y.; Deng, K.; Zhang, L.; Yang, M.; Liu, X.; Qi, C.; Wang, C.; Song, W.; Zhang, Y.; et al. Enhancement of tanshinone production in Salvia miltiorrhiza hairy root cultures by metabolic engineering. Plant Methods 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Miklaszewska, M.; Banaś, A.; Królicka, A. Metabolic engineering of fatty alcohol production in transgenic hairy roots of Crambe abyssinica. Biotechnol. Bioeng. 2017, 114, 1275–1282. [Google Scholar] [CrossRef]
- Shi, M.; Luo, X.; Ju, G.; Li, L.; Huang, S.; Zhang, T.; Wang, H.; Kai, G. Enhanced diterpene tanshinone accumulation and bioactivity of transgenic Salvia miltiorrhiza hairy roots by pathway engineering. J. Agric. Food Chem. 2016, 64, 2523–2530. [Google Scholar] [CrossRef]
- Sharma, M.; Koul, A.; Sharma, D.; Kaul, S.; Swamy, M.K.; Dhar, M.K. Metabolic Engineering Strategies for Enhancing the Production of Bio-active Compounds from Medicinal Plants. In Natural Bio-active Compounds: Volume 3: Biotechnology, Bioengineering, and Molecular Approaches; Akhtar, M.S., Swamy, M.K., Eds.; Springer Singapore: Singapore, 2019; pp. 287–316. ISBN 978-981-13-7438-8. [Google Scholar]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In vitro plant tissue culture: Means for production of biological active compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef]
- Gwak, Y.S.; Han, J.Y.; Adhikari, P.B.; Ahn, C.H.; Choi, Y.E. Heterologous production of a ginsenoside saponin (compound K) and its precursors in transgenic tobacco impairs the vegetative and reproductive growth. Planta 2017, 245, 1105–1119. [Google Scholar] [CrossRef]
- Yao, J.; Weng, Y.; Dickey, A.; Wang, K.Y. Plants as factories for human pharmaceuticals: Applications and challenges. Int. J. Mol. Sci. 2015, 16, 28549–28565. [Google Scholar] [CrossRef]
- Jamwal, K.; Bhattacharya, S.; Puri, S. Plant growth regulator mediated consequences of secondary metabolites in medicinal plants. J. Appl. Res. Med. Aromat. Plants 2018, 9, 26–38. [Google Scholar] [CrossRef]
- Halder, M.; Sarkar, S.; Jha, S. Elicitation: A biotechnological tool for enhanced production of secondary metabolites in hairy root cultures. Eng. Life Sci. 2019, 19, 880–895. [Google Scholar] [CrossRef] [Green Version]
- Gharari, Z.; Bagheri, K.; Danafar, H.; Sharafi, A. Enhanced flavonoid production in hairy root cultures of Scutellaria bornmuelleri by elicitor induced over-expression of MYB7 and FNSП2 genes. Plant Physiol. Biochem. 2020, 148, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Salmanzadeh, M.; Sabet, M.S.; Moieni, A.; Homaee, M. Heterologous expression of an acid phosphatase gene and phosphate limitation leads to substantial production of chicoric acid in Echinacea purpurea transgenic hairy roots. Planta 2020, 251. [Google Scholar] [CrossRef]
- Ko, K.S.; Ebizuka, Y.; Noguchi, H.; Sankawa, U. Production of polyketide pigments in hairy root cultures of Cassia plants. Chem. Pharm. Bull. 1995, 43, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Aminfar, Z.; Rabiei, B.; Tohidfar, M.; Mirjalili, M.H. Identification of key genes involved in the biosynthesis of triterpenic acids in the mint family. Sci. Rep. 2019. [Google Scholar] [CrossRef]
- Ali-Seyed, M.; Jantan, I.; Vijayaraghavan, K.; Bukhari, S.N.A. Betulinic Acid: Recent Advances in Chemical Modifications, Effective Delivery, and Molecular Mechanisms of a Promising Anticancer Therapy. Chem. Biol. Drug Des. 2016, 87, 517–536. [Google Scholar] [CrossRef]
- Lu, T.; Wei, D.; Yu, K.; Ma, D.; Xiong, J.; Fang, Q.; Wang, J. Betulinic acid restores imatinib sensitivity in BCR-ABL1 kinase−independent, imatinib-resistant chronic myeloid leukemia by increasing HDAC3 ubiquitination and degradation. Ann. N. Y. Acad. Sci. 2020. [Google Scholar] [CrossRef]
- Alakurtti, S.; Mäkelä, T.; Koskimies, S.; Yli-Kauhaluoma, J. Pharmacological properties of the ubiquitous natural product betulin. Eur. J. Pharm. Sci. 2006, 29, 1–13. [Google Scholar] [CrossRef]
- Guo, H.; Chang, Z.; Yang, R.; Guo, D.; Zheng, J. Anthraquinones from hairy root cultures of Cassia obtusifolia. Phytochemistry 1998, 49, 1623–1625. [Google Scholar] [CrossRef]
- Kowalczyk, T.; Sitarek, P.; Skała, E.; Toma, M.; Wielanek, M.; Pytel, D.; Wieczfińska, J.; Szemraj, J.; Śliwiński, T. Induction of apoptosis by in vitro and in vivo plant extracts derived from Menyanthes trifoliata L. in human cancer cells. Cytotechnology 2019, 71, 165–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, J.W.; Jeong, J.H.; Shin, C.G.; Lo, S.C.; Han, S.S.; Yu, K.W.; Harada, E.; Han, J.Y.; Choi, Y.E. Overexpression of squalene synthase in Eleutherococcus senticosus increases phytosterol and triterpene accumulation. Phytochemistry 2005, 66, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Jeong, J.H.; Seo, J.W.; Shin, C.G.; Kim, Y.S.; In, J.G.; Yang, D.C.; Yi, J.S.; Choi, Y.E. Enhanced triterpene and phytosterol biosynthesis in Panax ginseng overexpressing squalene synthase gene. Plant Cell Physiol. 2004, 45, 976–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirjalili, M.H.; Moyano, E.; Bonfill, M.; Cusido, R.M.; Palazón, J. Overexpression of the Arabidopsis thaliana squalene synthase gene in Withania coagulans hairy root cultures. Biol. Plant. 2011, 55, 357–360. [Google Scholar] [CrossRef]
- Deshpande, S.R.; Shankar Naik, B. Cytotoxicity of stem extracts of selected Cassia species against HeLa and breast cancer cell lines in vitro. Asian J. Pharm. Clin. Res. 2017, 10, 80–82. [Google Scholar]
- Srihari, R.; Richard, A.; Nandashree, K.; Karigar, C.S.; Niranjali Devaraj, S.; Dhananjaya, B.L. Evaluating the antiproliferative potential of methonolic leaf extract of Cassia nigricans. Int. J. Pharm. Pharm. Sci. 2015, 7, 154–155. [Google Scholar]
- Levy, A.; Lewis, A. Cassia alata leaf extract induces cytotoxicity in A549 lung cancer cells via a mechanism that is caspase 8 dependent TT-El extracto de la hoja de cassia alata induce citotoxicidad en las células A549 del cáncer pulmonar humano a través de un mecanismo. West Indian Med. J. 2011, 60, 608–614. [Google Scholar]
- Schötterl, S.; Miemietz, J.T.; Ilina, E.I.; Wirsik, N.M.; Ehrlich, I.; Gall, A.; Huber, S.M.; Lentzen, H.; Mittelbronn, M.; Naumann, U. Mistletoe-based drugs work in synergy with radio-chemotherapy in the treatment of glioma in vitro and in vivo in glioblastoma bearing mice. Evid. Based Complement. Altern. Med. 2019, 2019, 1376140. [Google Scholar]
- Tayeh, Z.; Ofir, R. Asteriscus graveolens extract in combination with cisplatin/etoposide/doxorubicin suppresses lymphoma cell growth through induction of caspase-3 dependent apoptosis. Int. J. Mol. Sci. 2018, 19, 2219. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.; Tyagi, A.K.; Singh, R.P.; Chan, D.C.F.; Agarwal, R. Synergistic anti-cancer effects of grape seed extract and conventional cytotoxic agent doxorubicin against human breast carcinoma cells. Breast Cancer Res. Treat. 2004, 85, 1–12. [Google Scholar] [CrossRef]
- Park, E.J.; Kwon, H.K.; Choi, Y.M.; Shin, H.J.; Choi, S. Doxorubicin induces cytotoxicity through upregulation of perk-dependent ATF3. PLoS ONE 2012, 7, e44990. [Google Scholar] [CrossRef] [Green Version]
- Hutagaol, S.H.; Satria, D. Combination effect of ethylacetate extract leaves of Moringa oleifera L. and doxorubicin against MCF-7 cell lines. J. Innov. Pharm. Biol. Sci. 2018, 5, 42–46. [Google Scholar]
- De Oliveira Júnior, R.G.; Christiane Adrielly, A.F.; da Silva Almeida, J.R.G.; Grougnet, R.; Thiéry, V.; Picot, L. Sensitization of tumor cells to chemotherapy by natural products: A systematic review of preclinical data and molecular mechanisms. Fitoterapia 2018, 129, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Watari, H.; Abualmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and molecular targeting therapy in cancer. BioMed Res. Int. 2014, 2014, 150845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speidel, D. Transcription-independent p53 apoptosis: An alternative route to death. Trends Cell Biol. 2010, 20, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Şoica, C.M.; Dehelean, C.A.; Peev, C.; Aluas, M.; Zupkó, I.; Kása, P.; Alexa, E. Physico-chemical comparison of betulinic acid, betulin and birch bark extract and in vitro investigation of their cytotoxic effects towards skin epidermoid carcinoma (A431), breast carcinoma (MCF7) and cervix adenocarcinoma (HeLa) cell lines. Nat. Prod. Res. 2012, 26, 968–974. [Google Scholar] [CrossRef]
- Drag, M.; Surowiak, P.; Malgorzata, D.Z.; Dietel, M.; Lage, H.; Oleksyszyn, J. Comparision of the cytotoxic effects of birch bark extract, betulin and betulinic acid towards human gastric carcinoma and pancreatic carcinoma drug-sensitive and drug-resistant cell lines. Molecules 2009, 14, 1639–1651. [Google Scholar] [CrossRef]
- Ehrhardt, H.; Fulda, S.; Führer, M.; Debatin, K.M.; Jeremias, I. Betulinic acid-induced apoptosis in leukemia cells. Leukemia 2004, 18, 1406–1412. [Google Scholar] [CrossRef]
- Ehrhardt, H.; Höfig, I.; Wachter, F.; Obexer, P.; Fulda, S.; Terziyska, N.; Jeremias, I. NOXA as critical mediator for drug combinations in polychemotherapy. Cell Death Dis. 2012, 3, e327. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Construct | Number of Seedlings Used for Transformation | Number of Seedlings with Hairy Root Induction | Hairy Root Induction Efficiency (%) |
|---|---|---|---|
| 35S::PgSS1 | 60 | 48 | 80 |
| 35S::GFP | 60 | 52 | 87 |
| A4 hairy roots | 60 | 55 | 92 |
| Roots Extracts from Various Clones of S. obtusifolia | Betulinic Acid mg/g DW |
|---|---|
| SOA41 | 2.7795 ± 0.0168 a |
| SOA42 | 1.9482 ± 0.0172 b |
| SOGFP1 | 4.0882 ± 0.2986 b |
| SOGFP2 | 4.7339 ± 0.0254 a |
| SOGFP3 | 3.9875 ± 0.0238 b |
| SOPSS1 | 15.7548 ± 0.4891 b |
| SOPSS2 | 22.7144 ± 0.0267 a |
| SOPSS3 | 8.6489 ± 0.3644 f |
| SOPSS4 | 5.9987 ± 0.0457 g |
| SOPSS5 | 14.4669 ± 0.0514 c |
| SOPSS6 | 10.9742 ± 0.092 d |
| SOPSS7 | 9.0251 ± 0.0494 e |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalczyk, T.; Sitarek, P.; Toma, M.; Picot, L.; Wielanek, M.; Skała, E.; Śliwiński, T. An Extract of Transgenic Senna obtusifolia L. hairy roots with Overexpression of PgSS1 Gene in Combination with Chemotherapeutic Agent Induces Apoptosis in the Leukemia Cell Line. Biomolecules 2020, 10, 510. https://doi.org/10.3390/biom10040510
Kowalczyk T, Sitarek P, Toma M, Picot L, Wielanek M, Skała E, Śliwiński T. An Extract of Transgenic Senna obtusifolia L. hairy roots with Overexpression of PgSS1 Gene in Combination with Chemotherapeutic Agent Induces Apoptosis in the Leukemia Cell Line. Biomolecules. 2020; 10(4):510. https://doi.org/10.3390/biom10040510
Chicago/Turabian StyleKowalczyk, Tomasz, Przemysław Sitarek, Monika Toma, Laurent Picot, Marzena Wielanek, Ewa Skała, and Tomasz Śliwiński. 2020. "An Extract of Transgenic Senna obtusifolia L. hairy roots with Overexpression of PgSS1 Gene in Combination with Chemotherapeutic Agent Induces Apoptosis in the Leukemia Cell Line" Biomolecules 10, no. 4: 510. https://doi.org/10.3390/biom10040510