Biochemical Pathways Leading to the Formation of Wyosine Derivatives in tRNA of Archaea


Abstract
:1. Introduction
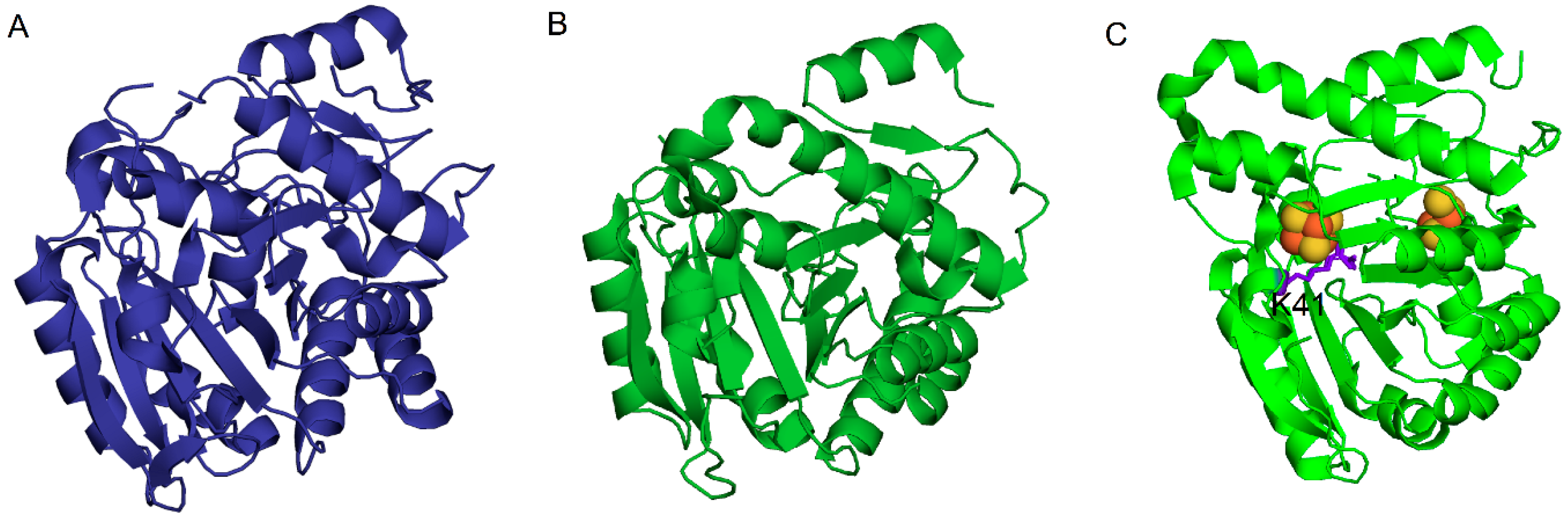
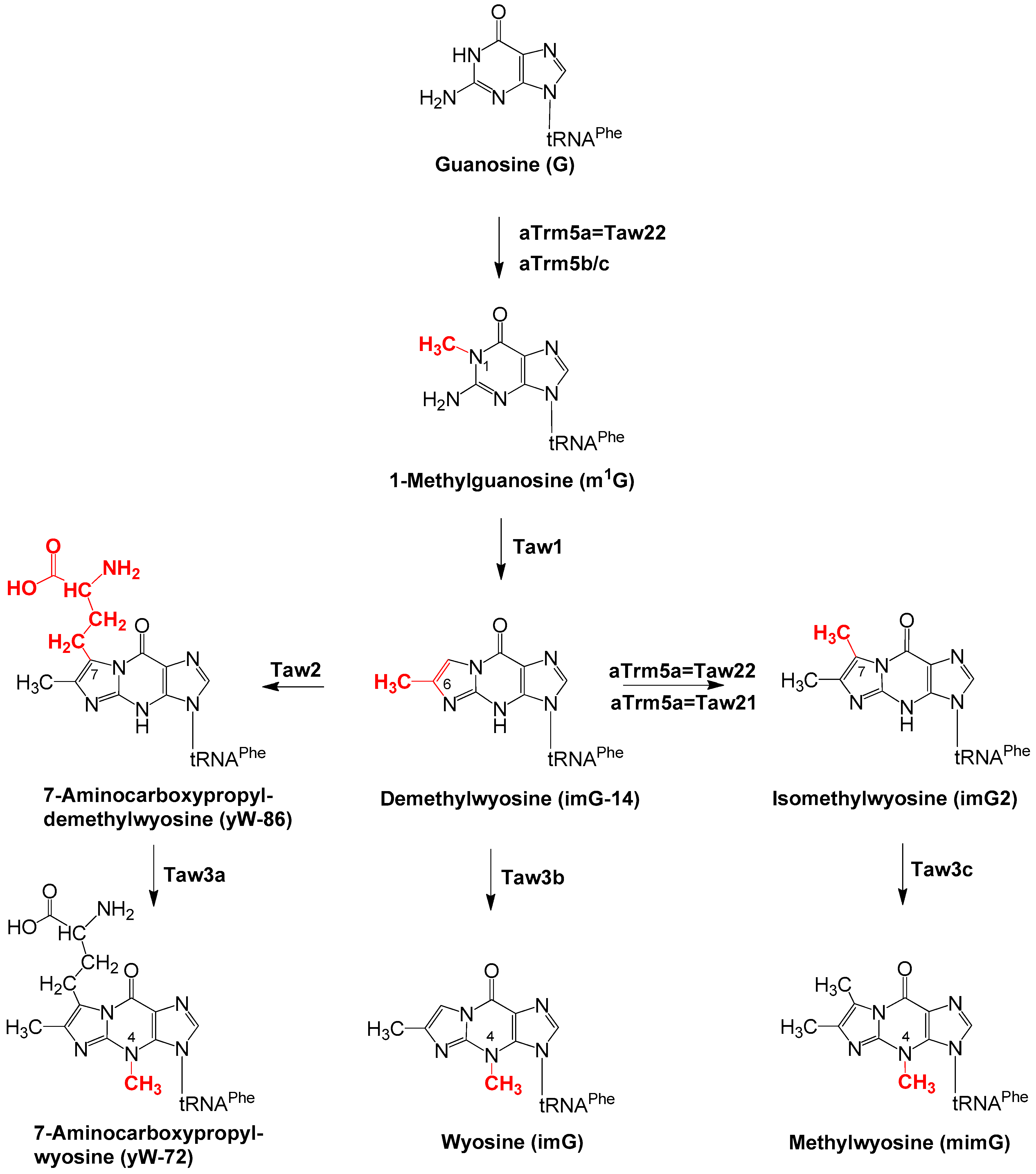
2. Enzymatic Formation of 1-Methylguanosine, Intermediate in the Pathway Leading to the Formation of Wyosine Derivatives
3. Enzymatic Biosynthesis of Wyosine Derivatives
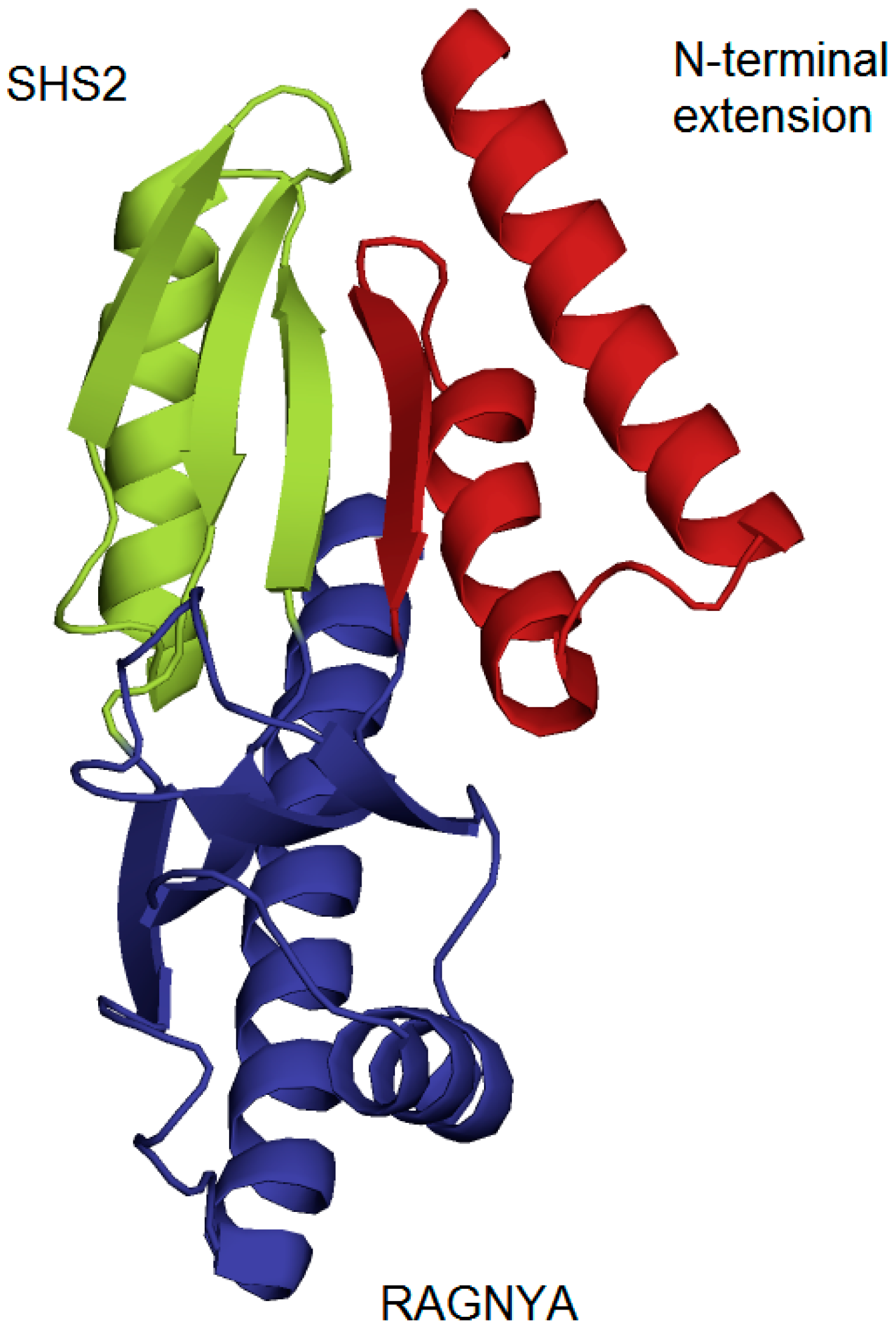
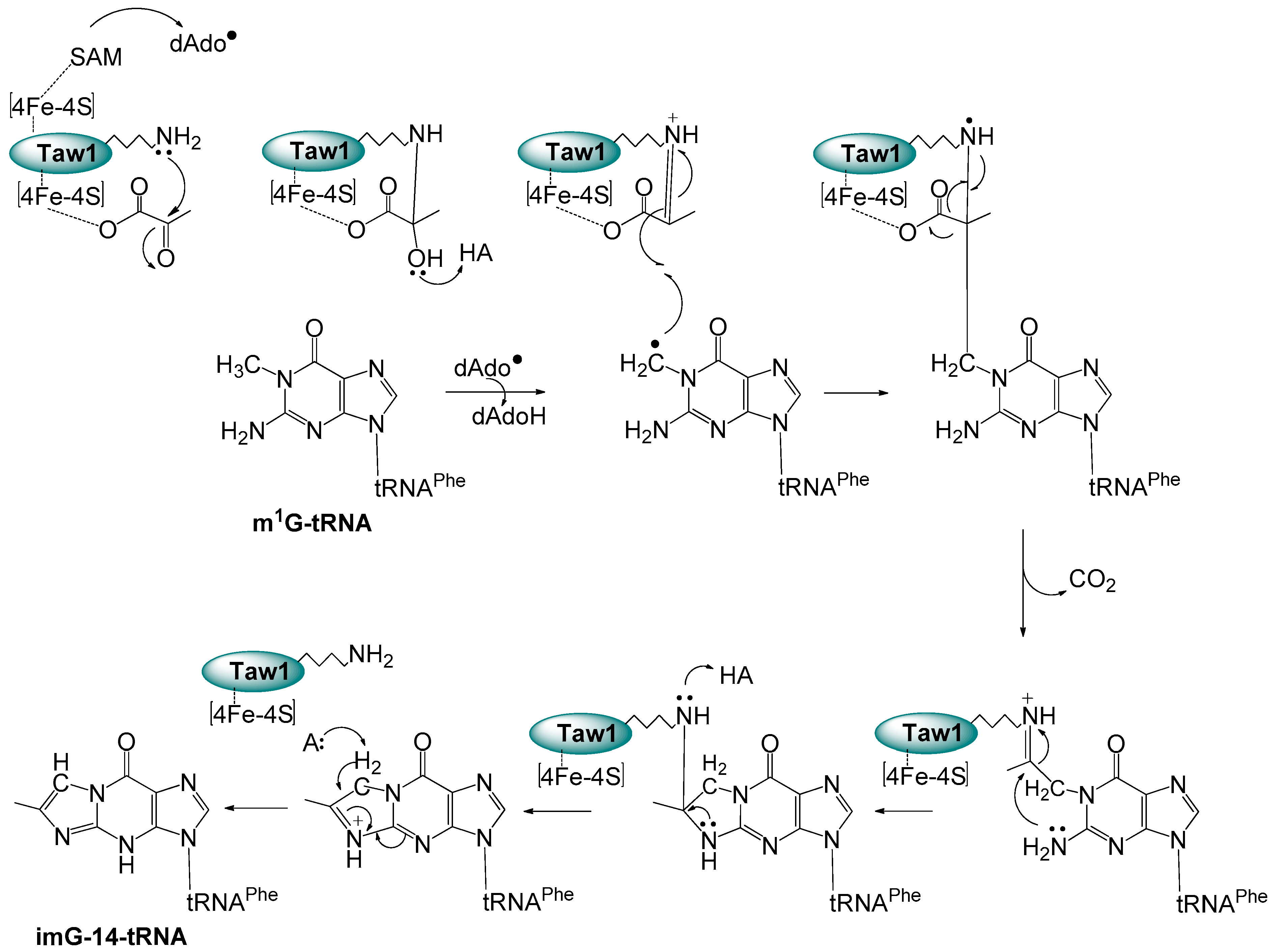
3.1. Formation of imG-14 by Taw1 Enzyme
3.2. Formation of yW-86 by the Taw2 Enzyme
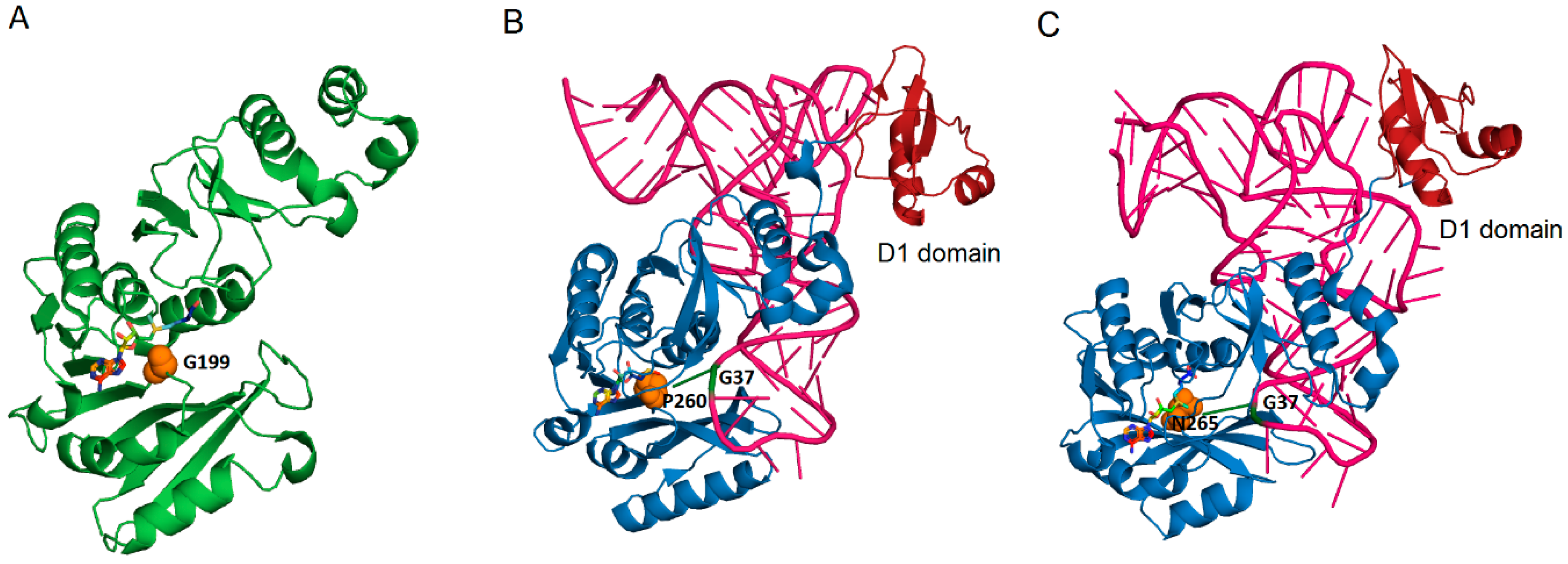
3.3. Formation of imG2 by Taw21/Taw22 (aTrm5) Enzymes
3.4. The Enzymatic Activity of Taw3 Proteins
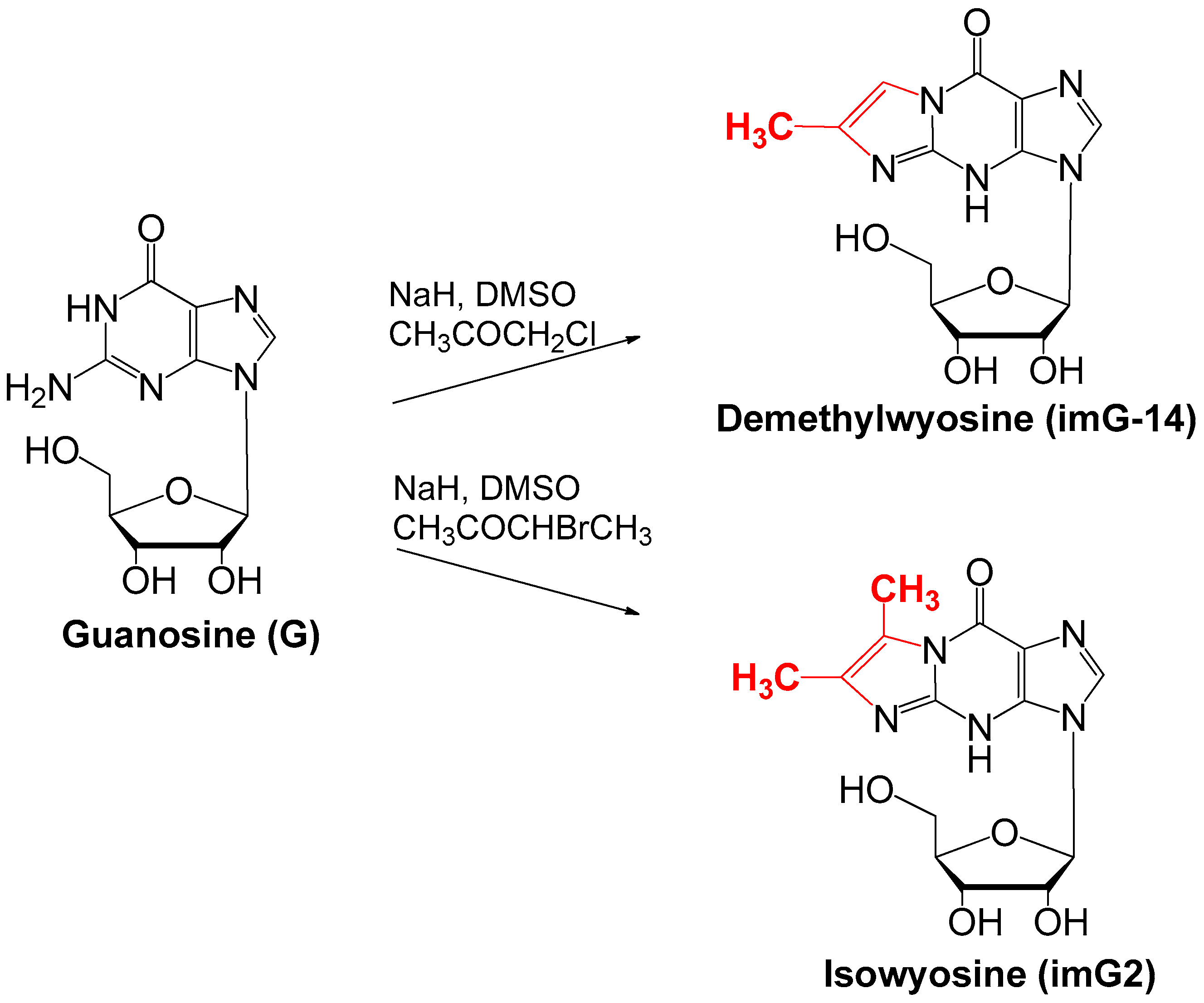
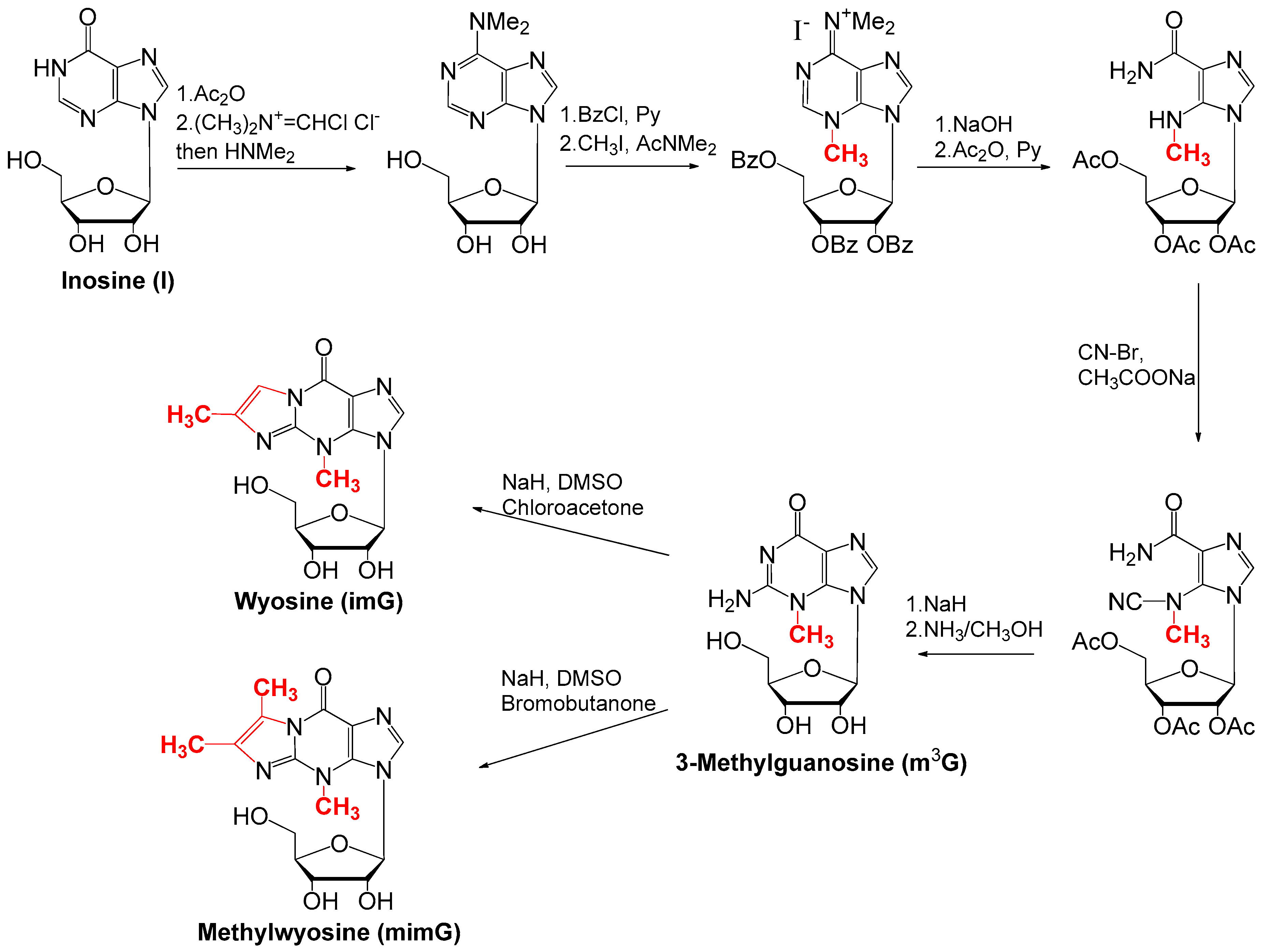
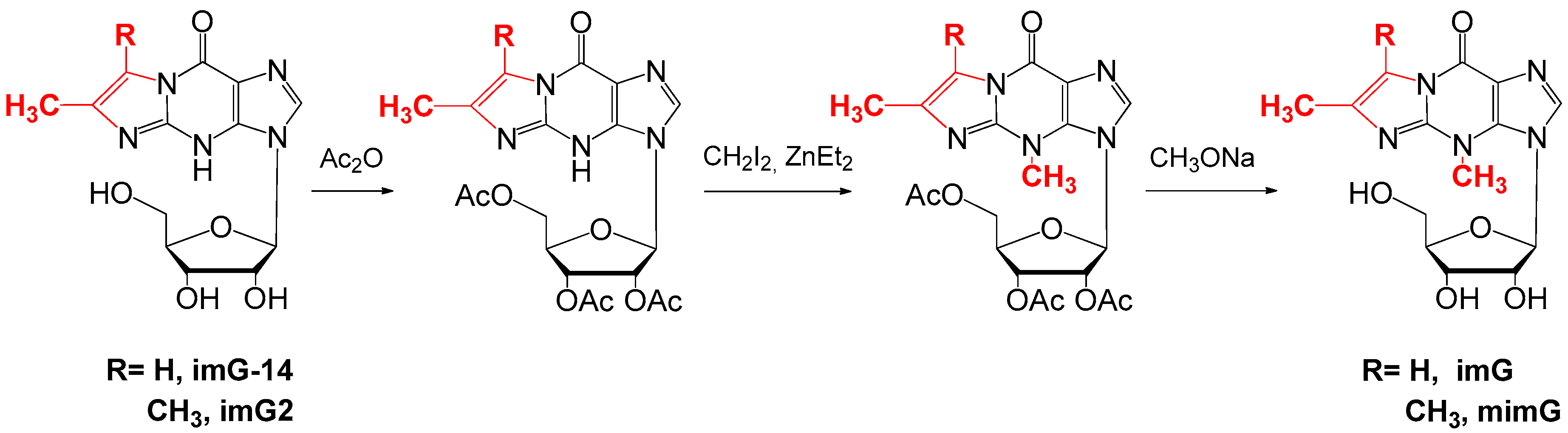
4. Chemical Synthesis of Wyosine Nucleosides
4.1. Synthesis of imG-14 and imG2
4.2. Synthesis of imG and mimG
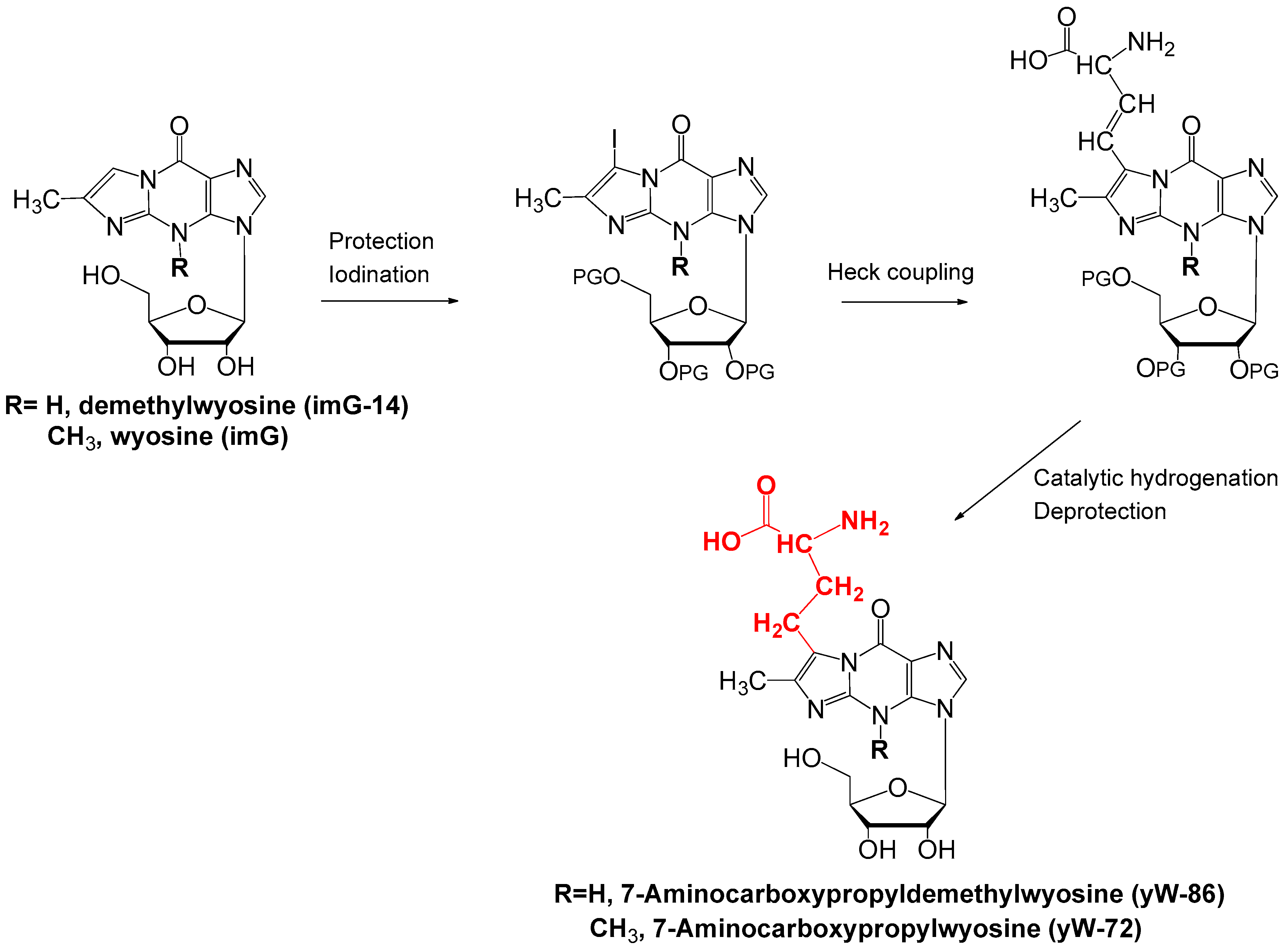
4.3. Synthesis of yW-86 and yW-72
5. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piatkowski, P.; Baginski, B.; Wirecki, T.K.; de Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A.; et al. MODOMICS: A database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef]
- Urbonavičius, J.; Meškys, R.; Grosjean, H. Biosynthesis of wyosine derivatives in tRNAPhe of Archaea: Role of a remarkable bifunctional tRNAPhe:m1G/imG2 methyltransferase. RNA 2014, 20, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Rajbhandary, U.L.; Chang, S.H. Studies on polynucleotides. J. Biol. Chem. 1968, 243, 598–608. [Google Scholar]
- Konevega, A.L.; Soboleva, N.G.; Makhno, V.I.; Semenkov, Y.P.; Wintermeyer, W.; Rodnina, M.V.; Katunin, V.I. Purine bases at position 37 of tRNA stabilize codon-anticodon interaction in the ribosomal A site by stacking and Mg2+-dependent interactions. RNA 2004, 10, 90–101. [Google Scholar] [CrossRef] [Green Version]
- Waas, W.F.; Druzina, Z.; Hanan, M.; Schimmel, P. Role of a tRNA base modification and its precursors in frameshifting in eukaryotes. J. Biol. Chem. 2007, 282, 26026–26034. [Google Scholar] [CrossRef] [Green Version]
- Carlson, B.A.; Mushinski, J.F.; Henderson, D.W.; Kwon, S.Y.; Crain, P.F.; Lee, B.J.; Hatfield, D.L. 1-Methylguanosine in place of Y base at position 37 in phenylalanine tRNA is responsible for its shiftiness in retroviral ribosomal frameshifting. Virology 2001, 279, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Noma, A.; Kirino, Y.; Ikeuchi, Y.; Suzuki, T. Biosynthesis of wybutosine, a hyper-modified nucleoside in eukaryotic phenylalanine tRNA. EMBO J. 2006, 25, 2142–2154. [Google Scholar] [CrossRef]
- Ohira, T.; Suzuki, T. Retrograde nuclear import of tRNA precursors is required for modified base biogenesis in yeast. Proc. Natl. Acad. Sci. USA 2011, 108, 10502–10507. [Google Scholar] [CrossRef] [Green Version]
- Itaya, T.; Kanai, T.; Sawada, T. Structure of wyosine, the condensed tricyclic nucleoside of Torula yeast phenylalanine transfer ribonucleic acid. Chem. Pharm. Bull. 2002, 50, 547–548. [Google Scholar] [CrossRef] [Green Version]
- Sample, P.J.; Kořený, L.; Paris, Z.; Gaston, K.W.; Rubio, M.A.T.; Fleming, I.M.; Hinger, S.; Horáková, E.; Limbach, P.A.; Lukeš, J.; et al. A common tRNA modification at an unusual location: The discovery of wyosine biosynthesis in mitochondria. Nucleic Acids Res. 2015, 43, 4262–4273. [Google Scholar] [CrossRef] [Green Version]
- White, B.N.; Tener, G.M. Properties of tRNAPhe from Drosophila. BBA Sect. Nucleic Acids Protein Synth. 1973, 312, 267–275. [Google Scholar] [CrossRef]
- Mazabraud, A. Deficiency of the peroxy-Y base in oocyte phenylalanine tRNA. FEBS Lett. 1979, 100, 235–240. [Google Scholar] [CrossRef] [Green Version]
- Keith, G.; Dirheimer, G. Primary structure of Bombyx mori posterior silkgland tRNAPhe. Biochem. Biophys. Res. Commun. 1980, 92, 109–115. [Google Scholar] [CrossRef]
- Mushinski, J.F.; Marini, M. Tumor-associated phenylalanyl transfer RNA found in a wide spectrum of rat and mouse tumors but absent in normal adult, fetal, and regenerating tissues. Cancer Res. 1979, 39. [Google Scholar]
- Rodriguez, V.; Chen, Y.; Elkahloun, A.; Dutra, A.; Pak, E.; Chandrasekharappa, S. Chromosome 8 BAC array comparative genomic hybridization and expression analysis identify amplification and overexpression of TRMT12 in breast cancer. Genes Chromosom. Cancer 2007, 46, 694–707. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, V.; Vasudevan, S.; Noma, A.; Carlson, B.A.; Green, J.E.; Suzuki, T.; Chandrasekharappa, S.C. Structure-function analysis of human TYW2 enzyme required for the biosynthesis of a highly modified wybutosine (yW) base in phenylalanine-tRNA. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Rosselló-Tortella, M.; Llinàs-Arias, P.; Sakaguchi, Y.; Miyauchi, K.; Davalos, V.; Setien, F.; Calleja-Cervantes, M.E.; Piñeyro, D.; Martínez-Gómez, J.; Guil, S.; et al. Epigenetic loss of the transfer RNA-modifying enzyme TYW2 induces ribosome frameshifts in colon cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 20785–20793. [Google Scholar] [CrossRef]
- De Crécy-Lagard, V.; Brochier-Armanet, C.; Urbonavičius, J.; Fernandez, B.; Phillips, G.; Lyons, B.; Noma, A.; Alvarez, S.; Droogmans, L.; Armengaud, J.; et al. Biosynthesis of wyosine derivatives in tRNA: An ancient and highly diverse pathway in Archaea. Mol. Biol. Evol. 2010, 27, 2062–2077. [Google Scholar] [CrossRef] [Green Version]
- Gupta, R. Halobacterium volcanii tRNAs: Identification of 41 tRNAs covering all amino acids, and the sequences of 33 class I tRNAs. J. Biol. Chem. 1984, 259, 9461–9471. [Google Scholar]
- McCloskey, J.A.; Graham, D.E.; Zhou, S.; Crain, P.F.; Ibba, M.; Konisky, J.; Söll, D.; Olsen, G.J. Post-transcriptional modification in archaeal tRNAs: Identities and phylogenetic relations of nucleotides from mesophilic and hyperthermophilic Methanococcales. Nucleic Acids Res. 2001, 29, 4699–4706. [Google Scholar] [CrossRef]
- Zhou, S.; Sitaramaiah, D.; Noon, K.R.; Guymon, R.; Hashizume, T.; McCloskey, J.A. Structures of two new “minimalist” modified nucleosides from archaeal tRNA. Bioorg. Chem. 2004, 32, 82–91. [Google Scholar] [CrossRef]
- Yu, N.; Jora, M.; Solivio, B.; Thakur, P.; Acevedo-Rocha, C.G.; Randau, L.; de Crécy-Lagard, V.; Addepalli, B.; Limbach, P.A. tRNA modification profiles and codon-decoding strategies in Methanocaldococcus jannaschii. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Hirata, A.; Suzuki, T.; Nagano, T.; Fujii, D.; Okamoto, M.; Sora, M.; Lowe, T.M.; Kanai, T.; Atomi, H.; Suzuki, T.; et al. Distinct modified nucleosides in tRNATrp from the hyperthermophilic Archaeon Thermococcus kodakarensis and requirement of tRNA m2G10/m22G10 methyltransferase (archaeal Trm11) for survival at high temperatures. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef]
- Björk, G.R.; Wikström, P.M.; Byström, A.S. Prevention of translational frameshifting by the modified nucleoside 1-methylguanosine. Science 1989, 244, 986–989. [Google Scholar] [CrossRef]
- Urbonavičius, J.; Qian, Q.; Durand, J.M.B.; Hagervall, T.G.; Björk, G.R. Improvement of reading frame maintenance is a common function for several tRNA modifications. EMBO J. 2001, 20, 4863–4873. [Google Scholar] [CrossRef] [Green Version]
- Goto-Ito, S.; Ito, T.; Yokoyama, S. Trm5 and TrmD: Two enzymes from distinct origins catalyze the identical tRNA modification, m1G37. Biomolecules 2017, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Urbonavičius, J.; Rutkiene, R.; Lopato, A.; Tauraite, D.; Stankevičiute, J.; Aučynaite, A.; Kaliniene, L.; Van Tilbeurgh, H.; Meškys, R. Evolution of tRNAPhe:imG2 methyltransferases involved in the biosynthesis of wyosine derivatives in archaea. RNA 2016, 22, 1871–1883. [Google Scholar] [CrossRef] [Green Version]
- Bujnicki, J.M. Phylogenomic analysis of 16S rRNA:(guanine-N2) methyltransferases suggests new family members and reveals highly conserved motifs and a domain structure similar to other nucleic acid amino-methyltransferases. FASEB J. 2000, 14, 2365–2368. [Google Scholar] [CrossRef]
- Goto-Ito, S.; Ito, T.; Ishii, R.; Muto, Y.; Bessho, Y.; Yokoyama, S. Crystal structure of archaeal tRNA(m1 G37)methyltransferase aTrm5. Proteins Struct. Funct. Genet. 2008, 72, 1274–1289. [Google Scholar] [CrossRef]
- Goto-Ito, S.; Ito, T.; Kuratani, M.; Bessho, Y.; Yokoyama, S. Tertiary structure checkpoint at anticodon loop modification in tRNA functional maturation. Nat. Struct. Mol. Biol. 2009, 16, 1109–1115. [Google Scholar] [CrossRef]
- Wu, J.; Jia, Q.; Wu, S.; Zeng, H.; Sun, Y.; Wang, C.; Ge, R.; Xie, W. The crystal structure of the Pyrococcus abyssi mono-functional methyltransferase PaTrm5b. Biochem. Biophys. Res. Commun. 2017, 493, 240–245. [Google Scholar] [CrossRef]
- Sofia, H.J.; Chen, G.; Hetzler, B.G.; Reyes-Spindola, J.F.; Miller, N.E. Radical SAM, a novel protein superfamily linking unresolved steps in familiar biosynthetic pathways with radical mechanisms: Functional characterization using new analysis and information visualization methods. Nucleic Acids Res. 2001, 29, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Young, A.P.; Bandarian, V. Pyruvate is the source of the two carbons that are required for formation of the imidazoline ring of 4-demethylwyosine. Biochemistry 2011, 50, 10573–10575. [Google Scholar] [CrossRef] [Green Version]
- Young, A.P.; Bandarian, V. Mechanistic studies of the radical S-adenosyl-L-methionine enzyme 4-demethylwyosine synthase reveal the site of hydrogen atom abstraction. Biochemistry 2015, 54, 3569–3572. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, Y.; Noma, A.; Suzuki, T.; Senda, M.; Senda, T.; Ishitani, R.; Nureki, O. Crystal structure of the radical SAM enzyme catalyzing tricyclic modified base formation in tRNA. J. Mol. Biol. 2007, 372, 1204–1214. [Google Scholar] [CrossRef]
- Goto-Ito, S.; Ishii, R.; Ito, T.; Shibata, R.; Fusatomi, E.; Sekine, S.I.; Bessho, Y.; Yokoyama, S. Structure of an archaeal TYW1, the enzyme catalyzing the second step of wye-base biosynthesis. Acta Crystallogr. Sect. D Biol. Crystallogr. 2007, 63, 1059–1068. [Google Scholar] [CrossRef]
- Kathirvelu, V.; Perche-Letuvée, P.; Latour, J.-M.; Atta, M.; Forouhar, F.; Gambarelli, S.; Garcia-Serres, R. Spectroscopic evidence for cofactor–substrate interaction in the radical-SAM enzyme TYW1. Dalt. Trans. 2017, 46, 13211. [Google Scholar] [CrossRef]
- Perche-Letuvée, P.; Molle, T.; Forouhar, F.; Mulliez, E.; Atta, M. Wybutosine biosynthesis: Structural and mechanistic overview. RNA Biol. 2014, 11, 1508–1518. [Google Scholar] [CrossRef] [Green Version]
- Grell, T.A.J.; Young, A.P.; Drennan, C.L.; Bandarian, V. Biochemical and structural characterization of a Schiff base in the radical-mediated biosynthesis of 4-demethylwyosine by TYW1. J. Am. Chem. Soc. 2018, 140, 6842–6852. [Google Scholar] [CrossRef]
- Young, A.P.; Bandarian, V. TYW1: A radical SAM enzyme involved in the biosynthesis of wybutosine bases. In Methods in Enzymology; Academic Press Inc.: Cambridge, MA, USA, 2018; Volume 606, pp. 119–153. ISBN 9780128127940. [Google Scholar]
- Bandarian, V. Radical SAM Enzymes; Bandarian, V., Ed.; Academic Press Inc.: Cambridge, MA, USA, 2018; Volume 606, ISBN 9780128127940. [Google Scholar]
- Matte-Tailliez, O.; Zivanovic, Y.; Forterre, P. Mining archaeal proteomes for eukaryotic proteins with novel functions: The PACE case. Trends Genet. 2000, 16, 533–536. [Google Scholar] [CrossRef]
- Makarova, K.S.; Sorokin, A.V.; Novichkov, P.S.; Wolf, Y.I.; Koonin, E.V. Clusters of orthologous genes for 41 archaeal genomes and implications for evolutionary genomics of archaea. Biol. Direct 2007, 2, 33. [Google Scholar] [CrossRef] [Green Version]
- Takakura, M.; Ishiguro, K.; Akichika, S.; Miyauchi, K.; Suzuki, T. Biogenesis and functions of aminocarboxypropyluridine in tRNA. Nat. Commun. 2019, 10, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Meyer, B.; Immer, C.; Kaiser, S.; Sharma, S.; Yang, J.; Watzinger, P.; Weiß, L.; Kotter, A.; Helm, M.; Seitz, H.-M.; et al. Identification of the 3-amino-3-carboxypropyl (acp) transferase enzyme responsible for acp3U formation at position 47 in Escherichia coli tRNAs. Nucleic Acids Res. 2020, 48, 1435–1450. [Google Scholar] [CrossRef]
- Umitsu, M.; Nishimasu, H.; Noma, A.; Suzuki, T.; Ishitani, R.; Nureki, O. Structural basis of AdoMet-dependent aminocarboxypropyl transfer reaction catalyzed by tRNA-wybutosine synthesizing enzyme, TYW2. Proc. Natl. Acad. Sci. USA 2009, 106, 15616–15621. [Google Scholar] [CrossRef] [Green Version]
- Phillips, G.; de Crécy-Lagard, V. Biosynthesis and function of tRNA modifications in Archaea. Curr. Opin. Microbiol. 2011, 14, 335–341. [Google Scholar] [CrossRef]
- Wang, C.; Jia, Q.; Chen, R.; Wei, Y.; Li, J.; Ma, J.; Xie, W. Crystal structures of the bifunctional tRNA methyltransferase Trm5a. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Jia, Q.; Zeng, J.; Chen, R.; Xie, W. Structural insight into the methyltransfer mechanism of the bifunctional Trm5. Sci. Adv. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Currie, M.A.; Brown, G.; Wong, A.; Ohira, T.; Sugiyama, K.; Suzuki, T.; Yakunin, A.F.; Jia, Z. Structural and functional characterization of the TYW3/Taw3 class of SAM-dependent methyltransferases. RNA 2017, 23, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, S.; Kurosawa, N. Development of the multiple gene knockout system with one-step PCR in thermoacidophilic crenarchaeon Sulfolobus acidocaldarius. Archaea 2017, 2017, 7459310. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Cooper, T.E.; Krause, D.J.; Whitaker, R.J. Augmenting the genetic toolbox for Sulfolobus islandicus with a stringent positive selectable marker for agmatine prototrophy. Appl. Environ. Microbiol. 2013, 79, 5539–5549. [Google Scholar] [CrossRef] [Green Version]
- Maezato, Y.; Dana, K.; Blum, P. Engineering thermoacidophilic archaea using linear DNA recombination. Methods Mol. Biol. 2011, 765, 435–445. [Google Scholar] [CrossRef]
- Edmonds, C.G.; Crain, P.F.; Gupta, R.; Hashizume, T.; Hocart, C.H.; Kowalak, J.A.; Pomerantz, S.C.; Stetter, K.O.; McCloskey, J.A. Posttranscriptional modification of tRNA in thermophilic archaea (archaebacteria). J. Bacteriol. 1991, 173, 3138–3148. [Google Scholar] [CrossRef] [Green Version]
- Mccloskey, J.A.; Liu, X.-H.; Crain, P.F.; Bruenger, E.; Guymon, R.; Hashizume, T.; Stetter, K. 0 Posttranscriptional modification of transfer RNA in the submarine hyperthermophile Pyrolobus fumarii. Nucleic Acids Symp. Ser. 2000, 44, 267–268. [Google Scholar] [CrossRef] [Green Version]
- Kasai, H.; Goto, M.; Ikeda, K.; Zama, M.; Mizuno, Y.; Takemura, S.; Matsuura, S.; Sugimoto, T.; Goto, T. Structure of wye (Yt base) and wyosine (Yt) from Torulopsis utilis phenylalanine transfer ribonucleic acid. Biochemistry 1976, 15, 898–904. [Google Scholar] [CrossRef]
- Golankiewicz, B.; Folkman, W. Methylation of desmethyl analogue of Y nudeosides. Wyosine from guanosine. Nucleic Acids Res. 1983, 1, 5243. [Google Scholar] [CrossRef] [Green Version]
- Itaya, T.; Morisue, M.; Takeda, M.; Kumazawa, Y. Synthesis of 7-methyl-3-b-D-ribofuranosylwye, the putative structure for the hypermodified nucleoside isolated from archaebacterial transfer ribonucleic acids. Chem. Pharm. Bull. 1990, 38, 2656–2661. [Google Scholar] [CrossRef] [Green Version]
- Jahnz-Wechmann, Z.; Framski, G.R.; Januszczyk, P.A.; Boryski, J. Base-modified nucleosides: Etheno derivatives. Front. Chem. 2016, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Itaya, T.; Watanabe, T.; Matsumoto, H. A simple synthesis of 3-β-D-ribofuranosylwye and the stability of its glycosidic bond. J. Chem. Soc. Chem. Commun. 1980, 1158–1159. [Google Scholar] [CrossRef]
- Itaya, T.; Matsumoto, H.; Watanabe, T. Synthesis of 5-(methylamino)-1-β-D-ribofuranosylimidazole-4-carboxamide, a synthetic intermediate for 3-methyl-9-β-D-ribofuranosylpurines. Chem. Pharm. Bull. 1982, 30, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Bazin, H.; Zhou, X.X.; Glemarec, C.; Chattopadhyaya, J. An efficient synthesis of y-nucleoside (wyosine) by regiospecific methylation of N4-desmethylwyosine using organozinc reagent. Tetrahedron Lett. 1987, 28, 3275–3278. [Google Scholar] [CrossRef]
- Hienzsch, A.; Deiml, C.; Reiter, V.; Carell, T. Total synthesis of the hypermodified RNA bases wybutosine and hydroxywybutosine and their quantification together with other modified RNA bases in plant materials. Chem.-A Eur. J. 2013, 19, 4244–4248. [Google Scholar] [CrossRef]
- Itaya, T.; Morisue, M.; Shimomichi, M.; Ozasa, M.; Shimizu, S.; Nakagawa, S. Synthesis of 3-β-D-ribofuranosylwybutine, the most probable structure for the hypermodified nucleoside isolated from yeast phenylalanine transfer ribonucleic acids. J. Chem. Soc. Perkin Trans. 1 1994, 2759–2765. [Google Scholar] [CrossRef]
- Itaya, T.; Kanai, T. Synthesis and structure of the hypermodified nucleoside of rat liver phenylalanine transfer ribonucleic acid. Chem. Pharm. Bull. 2002, 50, 1318–1326. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Archaea | Wyosine Derivatives * | taw3 Genes | Predicted Taw3 Enzymatic Activities |
|---|---|---|---|
| P. abyssi P. furiosus P. horikoshii | yW-72 imG mimG | 2 | a/b/c |
| M. jannaschii M. litotrophus | imG | 1 | b |
| S. solfataricus T. neutropylus P. calidifontis P. islandicum A. pernix | mimG | 1 | c |
| A. fulgidus | ? (no mimG) | 1 | a, b, or a/b |
| P. fumarii | imG mimG | 2 | b and c |
| S. hydrogenophyla S. marinus | imG mimG | 1 | b/c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbonavičius, J.; Tauraitė, D. Biochemical Pathways Leading to the Formation of Wyosine Derivatives in tRNA of Archaea. Biomolecules 2020, 10, 1627. https://doi.org/10.3390/biom10121627
Urbonavičius J, Tauraitė D. Biochemical Pathways Leading to the Formation of Wyosine Derivatives in tRNA of Archaea. Biomolecules. 2020; 10(12):1627. https://doi.org/10.3390/biom10121627
Chicago/Turabian StyleUrbonavičius, Jaunius, and Daiva Tauraitė. 2020. "Biochemical Pathways Leading to the Formation of Wyosine Derivatives in tRNA of Archaea" Biomolecules 10, no. 12: 1627. https://doi.org/10.3390/biom10121627