Heavily Armed Ancestors: CRISPR Immunity and Applications in Archaea with a Comparative Analysis of CRISPR Types in Sulfolobales


Abstract
:1. Introduction—Where There Is a Cell, There Is a Virus
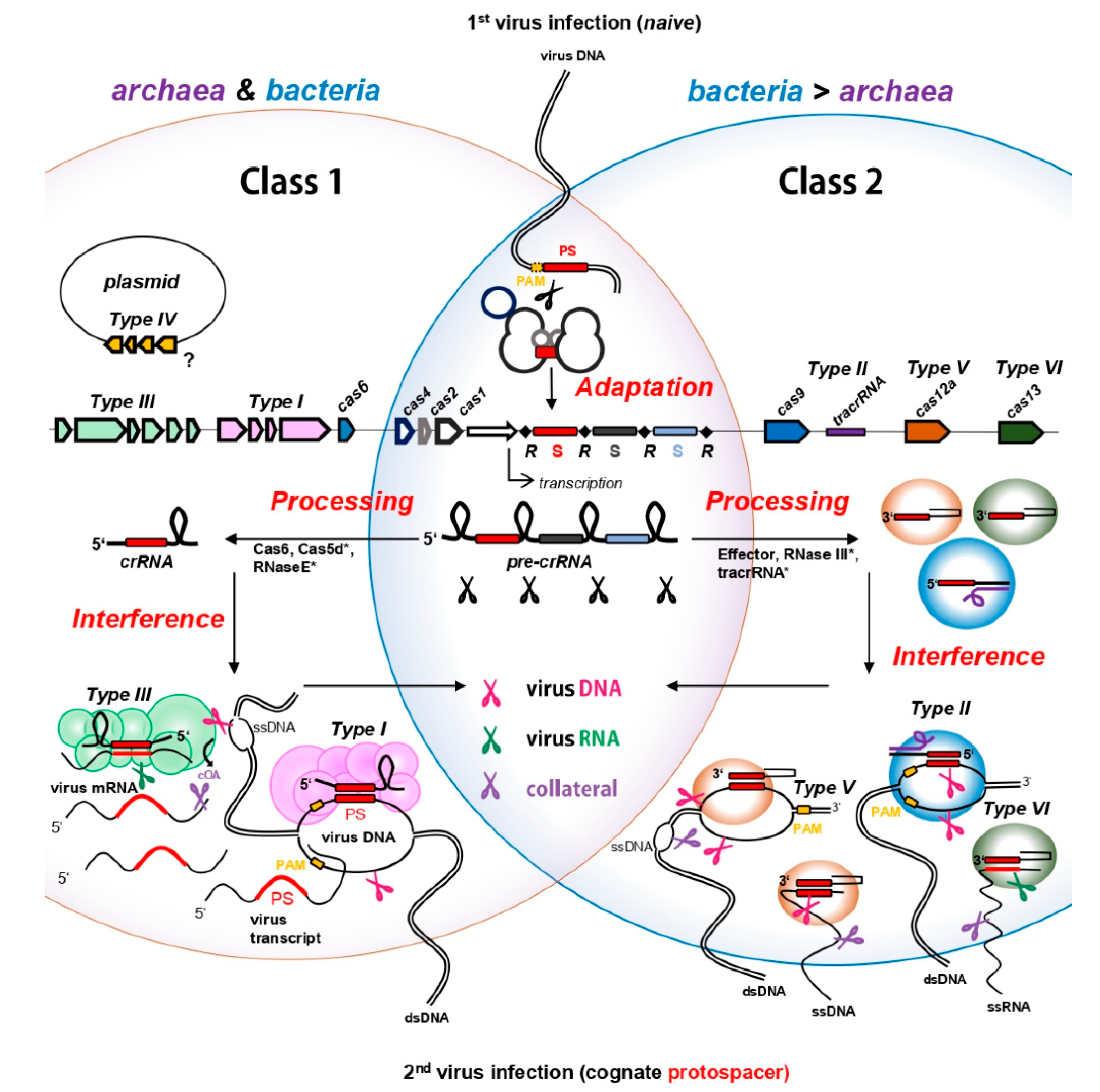
2. CRISPR Immunity in Prokaryotes—Archaea and Bacteria in the Ring with Viruses
2.1. CRISPR Adaptation—Know Your Enemy
2.2. Processing of crRNAs—Forging Weapons
2.3. CRISPR Interference-Counterstrike
2.3.1. CRISPR DNA Interference
2.3.2. “RNAttack”
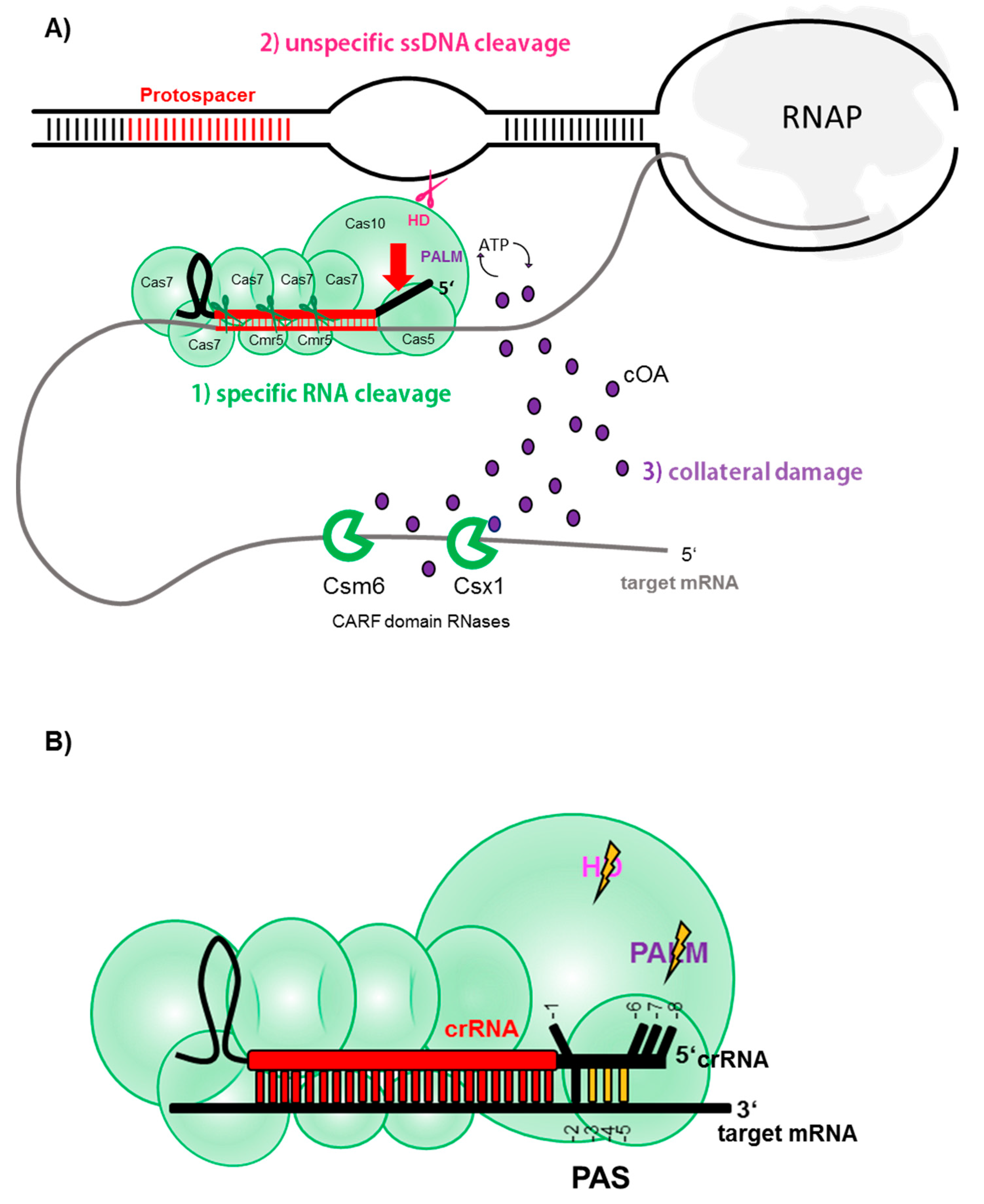
3. All Good Things Come in (Type) Threes—A Tripartite Immune Response of CRISPR Type III Systems
3.1. Specific RNA Cleavage
3.2. Unspecific ssDNA Cleavage
3.3. Collateral ssRNA Cleavage via cOA Signaling
4. CRISPR Research and Application in Archaeal Model Organisms
4.1. Experimental CRISPR Research in Archaeal Model Strains
4.1.1. Haloarchaea—Attack One’s Own Kind
4.1.2. Pyrococcus—Shaping CRISPR Crystals
4.1.3. Methanoarchaea—CRISPR Models on the Fast Lane?
4.1.4. Sulfolobales—The Virus Fighters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Archaeal Order | CRISPR Model Organism + | Physiology | CRISPR Types * | CRISPR Steps Studied $ | In Vivo Application |
|---|---|---|---|---|---|
| Thermococcales | Pyrococcus furiosus | hyperthermophilic, anaeorbic | COM: Type I-A, Type I-B, Type III-B | Adaptation [33] a, [180] b Processing [172] a, [58] b, RNA interference [175] a, [110] b DNA interference [179] a, [122] a,b cOA signaling & [147] a,b | |
| Pyrococcus horikoshii | hyperthermophilic, anaeorbic | OT3: Type I-A, Type I-B (x2), Type III-A | Processing [219] b | ||
| Thermococcus kodakarensis | hyperthermophilic, anaeorbic | KOD1: Type I-A, Type I-B | Processing [220] a DNA interference [220] a | CRISPR locus engineered to target invading plasmid [220] | |
| Thermococcus onnurineus | hyperthermophilic, anaeorbic | NA1: Type III-A, Type IV-C | DNA interference [126] b RNA interference [221] b cOA signaling [146] b | ||
| Methanosarcinales | Methanosarcina mazei | mesophilic, anaerobic | Go1: Type I-B, Type III-C | Processing [188] a,b cOA signaling [145] c,b | |
| Methanosarcina acetivorans | mesophilic, anaerobic | C2A: Type I-B, Type III-A | Cas9 genome editing * [222], dCas9 silencing * [223] | ||
| Methanococcales | Methanococcus maripaludis | mesophilic, anaerobic | C5: Type I-B | Processing [186] a,b DNA interference [190] c | |
| Methanocaldococcus jannaschii | hyperthermophilic, anaerobic | DSM 2661: Type I-A, partial Type III-A | DNA interference [182] b | ||
| Methanobacteriales | Methanothermobacter thermoautotrophicus | thermophilic, anaerobic | Type I-B, Type III-A, Type III-C | DNA interference [163] b | |
| Methanopyrales | Methanopyrus kandleri | hyperthermophilic, anaerobic | AV19: Type III-A, Type III-B | Processing [224] a | |
| Halobacteriales | Haloferax volcanii | mesophilic, halophilic, aerobic | DS2: Type I-B | Adaptation [169] a Processing [159] a DNA interference [161] a | CRISPRi: Type I-B gene silencing [225] |
| Haloferax mediterranei | mesophilic, halophilic, aerobic | ATCC 33500: Type I-B | Processing [160] a | ||
| Haloarcula hispanica | mesophilic, halophilic, aerobic | ATCC 33960: Type I-B | Adaptation [49] a DNA interference [164] a | Type I genome editing [226] | |
| Archaeoglobales | Archaeoglobus fulgidus | hyperthermophilic, anaerobic | DSM 4304: Type I-A (x2), Type III-B | Adaptation [227] b Processing [228] a RNA interference [107] b cOA signaling [145] b | |
| Sulfolobales # | Saccharolobus solfataricus | thermophilic, aerobic | P1: Type I-A (x3), Type III-B, Type III-D, partial Type III-B | Adaptation [229] a, [230] b Processing [231] a, [82] b DNA Interference [204] a, [232] b RNA Interference [119] a, [101] b cOA signaling [151] a, [138] b | Type III gene silencing [119] |
| Saccharolobus islandicus | thermophilic, aerobic | REY 15A: Type I-A, Type III-B (x2), | Adaptation [233] a, Processing [207] a, DNA Interference [204] a, [124] b RNA Interference [118] a, [124] b, cOAsignaling [141] b | Type III gene silencing [118], Type I genome editing [234], anti-CRISPR based virus editing [235] | |
| Sulfolobus acidocaldarius | thermophilic, aerobic | DSM 639: Type I-D, Type III-D | Processing [236] a | Type III gene silencing [237] | |
| Thermoproteales # | Thermoproteus tenax | hyperthermophilic, anaerobic | Kra 1: Type I-A, Type III-A, partial Type I-A | Processing [238] a DNA interference [239] b | |
| Pyrobaculum calidifontis | hyperthermophilic, anaerobic | JCM 11548: Type I-A, Type III-B (x2), | Processing [240] a |
4.2. CRISPR Application in Archaeal Models
5. A Hot Fuzz: CRISPR Immunity in Sulfolobales
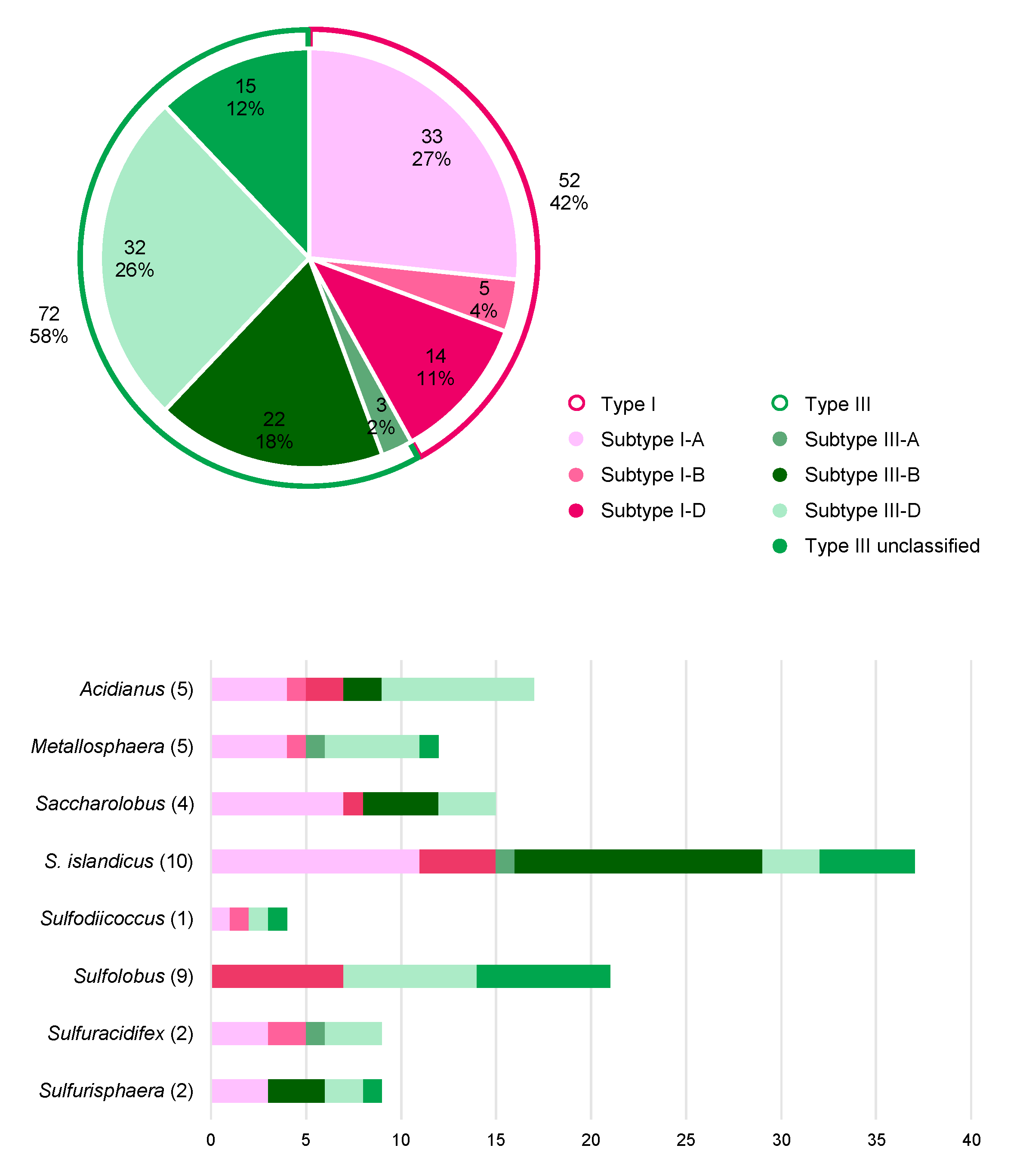
5.1. Distribution of CRISPR Types in Sulfolobales
5.2. CRISPR Arrays and Virus Matches in Sulfolobales

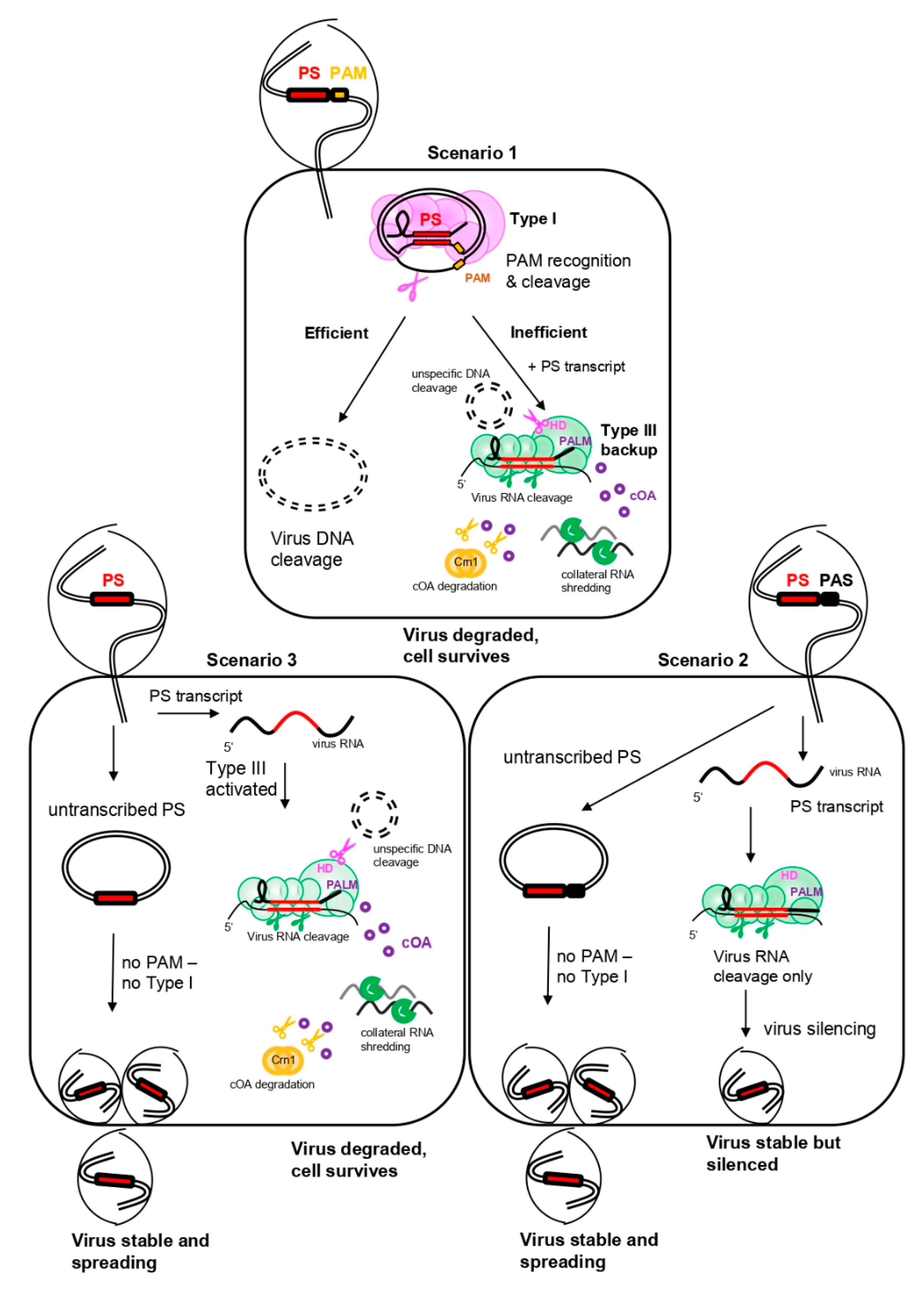
5.3. A General Scenario for CRISPR Interference in a Sulfolobales Cell

Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Parikka, K.J.; Le Romancer, M.; Wauters, N.; Jacquet, S. Deciphering the virus-to-prokaryote ratio (VPR): Insights into virus–host relationships in a variety of ecosystems. Biol. Rev. 2017, 92, 1081–1100. [Google Scholar] [CrossRef] [PubMed]
- Pietilä, M.K.; Demina, T.A.; Atanasova, N.S.; Oksanen, H.M.; Bamford, D.H. Archaeal viruses and bacteriophages: Comparisons and contrasts. Trends Microbiol. 2014, 22, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Forterre, P.; Prangishvili, D. The major role of viruses in cellular evolution: Facts and hypotheses. Curr. Opin. Virol. 2013, 3, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Lindell, D.; Sullivan, M.B.; Johnson, Z.I.; Tolonen, A.C.; Rohwer, F.; Chisholm, S.W. Transfer of photosynthesis genes to and from Prochlorococcus viruses. Proc. Natl. Acad. Sci. USA 2004, 101, 11013–11018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Wolf, Y.I. Evolution of microbes and viruses: A paradigm shift in evolutionary biology? Front. Cell. Infect. Microbiol. 2012, 2, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thingstad, T.F. Elements of a theory for the mechanisms controlling abundance, diversity, and biogeochemical role of lytic bacterial viruses in aquatic systems. Limnol. Oceanogr. 2000, 45, 1320–1328. [Google Scholar] [CrossRef]
- Brüssow, H.; Canchaya, C.; Hardt, W.; Bru, H. Phages and the Evolution of Bacterial Pathogens: From Genomic Rearrangements to Lysogenic Conversion Phages and the Evolution of Bacterial Pathogens: From Genomic Rearrangements to Lysogenic Conversion. Microbiol. Mol. Biol. Rev. 2004, 68, 560–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busby, B.; Kristensen, D.M.; Koonin, E.V. Contribution of phage-derived genomic islands to the virulence of facultative bacterial pathogens. Environ. Microbiol. 2013, 15, 307–312. [Google Scholar] [CrossRef]
- Tock, M.R.; Dryden, D.T.F.; Tock, M.R.; Dryden, D.T.F. The biology of restriction and anti-restriction. Curr. Opin. Microbiol. 2005, 8, 466–472. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Micro. 2010, 8, 317–327. [Google Scholar] [CrossRef]
- Lopatina, A.; Tal, N.; Sorek, R. Abortive Infection: Bacterial Suicide as an Antiviral Immune Strategy. Annu. Rev. Virol. 2020, 7, 371–384. [Google Scholar] [CrossRef]
- Swarts, D.C.; Jore, M.M.; Westra, E.R.; Zhu, Y.; Janssen, J.H.; Snijders, A.P.; Wang, Y.; Patel, D.J.; Berenguer, J.; Brouns, S.J.J.; et al. DNA-guided DNA interference by a prokaryotic Argonaute. Nature 2014, 507, 258–261. [Google Scholar] [CrossRef] [Green Version]
- Goldfarb, T.; Sberro, H.; Weinstock, E.; Cohen, O.; Doron, S.; Charpak-Amikam, Y.; Afik, S.; Ofir, G.; Sorek, R. BREX is a novel phage resistance system widespread in microbial genomes. EMBO J. 2015, 34, 169–183. [Google Scholar] [CrossRef]
- Ofir, G.; Melamed, S.; Sberro, H.; Mukamel, Z.; Silverman, S.; Yaakov, G.; Doron, S.; Sorek, R. DISARM is a widespread bacterial defence system with broad anti-phage activities. Nat. Microbiol. 2018, 3, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Doron, S.; Melamed, S.; Ofir, G.; Leavitt, A.; Lopatina, A.; Keren, M.; Amitai, G.; Sorek, R. Systematic discovery of antiphage defense systems in the microbial pangenome. Science 2018, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordeeva, J.; Morozova, N.; Sierro, N.; Isaev, A.; Sinkunas, T.; Tsvetkova, K.; Matlashov, M.; Truncaite, L.; Morgan, R.D.; Ivanov, N.V.; et al. BREX system of Escherichia coli distinguishes self from non-self by methylation of a specific DNA site. Nucleic Acids Res. 2019, 47, 253–265. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Wolf, Y.I. Is evolution Darwinian or/and Lamarckian? Biol. Direct 2009, 4, 42. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, C.; Touchon, M.; Villeriot, N.; Vernadet, J.P.; Couvin, D.; Toffano-Nioche, C.; Vergnaud, G. CRISPRCasdb a successor of CRISPRdb containing CRISPR arrays and cas genes from complete genome sequences, and tools to download and query lists of repeats and spacers. Nucleic Acids Res. 2020, 48, D535–D544. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR–Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Jansen, R.; Embden, J.D.; Gaastra, W.; Schouls, L.M. Identification of genes that are associated with DNA repeats in prokaryotes. Mol. Microbiol. 2002, 43, 1565–1575. [Google Scholar] [CrossRef] [PubMed]
- Lillestøl, R.K.; Shah, S.A.; Brügger, K.; Redder, P.; Phan, H.; Christiansen, J.; Garrett, R. CRISPR families of the crenarchaeal genus Sulfolobus: Bidirectional transcription and dynamic properties. Mol. Microbiol. 2009, 72, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Pul, U.; Wurm, R.; Arslan, Z.; Geissen, R.; Hofmann, N.; Wagner, R. Identification and characterization of E. coli CRISPR-cas promoters and their silencing by H-NS. Mol. Microbiol. 2010, 75, 1495–1512. [Google Scholar] [CrossRef] [PubMed]
- Shmakov, S.A.; Wolf, Y.I.; Savitskaya, E.; Severinov, K.V.; Koonin, E.V. Mapping CRISPR spaceromes reveals vast host-specific viromes of prokaryotes. Commun. Biol. 2020, 3, 1–9. [Google Scholar] [CrossRef]
- Shmakov, S.A.; Sitnik, V.; Makarova, K.S.; Wolf, Y.I.; Severinov, K.V.; Koonin, E.V. The CRISPR spacer space is dominated by sequences from species-specific mobilomes. Mbio 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, K.H.; Haitjema, C.; Liu, X.; Ding, F.; Wang, H.; DeLisa, M.P.; Ke, A. Cas5d protein processes pre-crRNA and assembles into a cascade-like interference complex in subtype I-C/Dvulg CRISPR-Cas system. Structure 2012, 20, 1574–1584. [Google Scholar] [CrossRef] [Green Version]
- Behler, J.; Sharma, K.; Reimann, V.; Wilde, A.; Urlaub, H.; Hess, W.R. The host-encoded RNase E endonuclease as the crRNA maturation enzyme in a CRISPR–Cas subtype III-Bv system. Nat. Microbiol. 2018, 3, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Fuglsang, A.; Kjeldsen, A.L.; Sun, K.; Bhoobalan-Chitty, Y.; Peng, X. DNA targeting by subtype I-D CRISPR-Cas shows type I and type III features. Nucleic Acids Res. 2020, 48, 10470–10478. [Google Scholar] [CrossRef]
- Silas, S.; Mohr, G.; Sidote, D.J.; Markham, L.M.; Sanchez-Amat, A.; Bhaya, D.; Lambowitz, A.M.; Fire, A.Z. Direct CRISPR spacer acquisition from RNA by a natural reverse transcriptase-Cas1 fusion protein. Science 2016, 351, aad4234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.X.; Hunnewell, P.; Alfonse, L.E.; Carte, J.M.; Keston-Smith, E.; Sothiselvam, S.; Garrity, A.J.; Chong, S.; Makarova, K.S.; Koonin, E.V.; et al. Functionally diverse type V CRISPR-Cas systems. Science 2019, 363, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef] [Green Version]
- Modell, J.W.; Jiang, W.; Marraffini, L.A. CRISPR–Cas systems exploit viral DNA injection to establish and maintain adaptive immunity. Nature 2017, 544, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Levy, A.; Goren, M.G.; Yosef, I.; Auster, O.; Manor, M.; Amitai, G.; Edgar, R.; Qimron, U.; Sorek, R. CRISPR adaptation biases explain preference for acquisition of foreign DNA. Nature 2015, 520, 505–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiimori, M.; Garrett, S.C.; Chambers, D.P.; Glover, C.V.C.; Graveley, B.R.; Terns, M.P. Role of free DNA ends and protospacer adjacent motifs for CRISPR DNA uptake in pyrococcus furiosus. Nucleic Acids Res. 2017, 45, 11281–11294. [Google Scholar] [CrossRef] [Green Version]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Almendros, C. Short motif sequences determine the targets of the prokaryotic CRISPR defence system. Microbiology 2009, 155, 733–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarts, D.C.; Mosterd, C.; van Passel, M.W.J.; Brouns, S.J.J. CRISPR interference directs strand specific spacer acquisition. PLoS ONE 2012, 7, e35888. [Google Scholar] [CrossRef]
- McGinn, J.; Marraffini, L.A. Molecular mechanisms of CRISPR–Cas spacer acquisition. Nat. Rev. Microbiol. 2019, 17, 7–12. [Google Scholar] [CrossRef]
- Wang, J.; Li, J.; Zhao, H.; Sheng, G.; Wang, M.; Yin, M.; Wang, Y. Structural and Mechanistic Basis of PAM-Dependent Spacer Acquisition in CRISPR-Cas Systems. Cell 2015, 163, 840–853. [Google Scholar] [CrossRef] [Green Version]
- Nuñez, J.K.; Kranzusch, P.J.; Noeske, J.; Wright, A.V.; Davies, C.W.; Doudna, J.A. Cas1–Cas2 complex formation mediates spacer acquisition during CRISPR–Cas adaptive immunity. Nat. Struct. Mol. Biol. 2014, 21, 528–534. [Google Scholar] [CrossRef]
- Grissa, I.; Vergnaud, G.; Pourcel, C. The CRISPRdb database and tools to display CRISPRs and to generate dictionaries of spacers and repeats. Bioinformatics 2007, 8. [Google Scholar] [CrossRef] [Green Version]
- Nuñez, J.K.; Bai, L.; Harrington, L.B.; Hinder, T.L.; Doudna, J.A. CRISPR Immunological Memory Requires a Host Factor for Specificity. Mol. Cell 2016, 62, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Yoganand, K.N.R.; Sivathanu, R.; Nimkar, S.; Anand, B. Asymmetric positioning of Cas1-2 complex and Integration Host Factor induced DNA bending guide the unidirectional homing of protospacer in CRISPR-Cas type I-E system. Nucleic Acids Res. 2017, 45, 367–381. [Google Scholar] [CrossRef]
- Rollie, C.; Graham, S.; Rouillon, C.; White, M.F. Prespacer processing and specific integration in a type I-A CRISPR system. Nucleic Acids Res. 2018, 46, 1007–1020. [Google Scholar] [CrossRef] [Green Version]
- Heler, R.; Samai, P.; Modell, J.W.; Weiner, C.; Goldberg, G.W.; Bikard, D.; Marraffini, L.A. Cas9 specifies functional viral targets during CRISPR–Cas adaptation. Nature 2015, 519, 199–202. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Zhou, Y.; Taylor, D.W.; Sashital, D.G. Cas4-Dependent Prespacer Processing Ensures High-Fidelity Programming of CRISPR Arrays. Mol. Cell 2018, 70, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Shiimori, M.; Garrett, S.C.; Graveley, B.R.; Terns, M.P. Cas4 Nucleases Define the PAM, Length, and Orientation of DNA Fragments Integrated at CRISPR Loci. Mol. Cell 2018, 70, 814–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kieper, S.N.; Almendros, C.; Behler, J.; McKenzie, R.E.; Nobrega, F.L.; Haagsma, A.C.; Vink, J.N.A.; Hess, W.R.; Brouns, S.J.J. Cas4 Facilitates PAM-Compatible Spacer Selection during CRISPR Adaptation. Cell Rep. 2018, 22, 3377–3384. [Google Scholar] [CrossRef]
- Zhang, Z.; Pan, S.; Liu, T.; Li, Y.; Peng, N. Cas4 nucleases can effect specific integration of CRISPR spacers. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Liu, Z.; Ye, Q.; Pan, S.; Wang, X.; Li, Y.; Peng, W.; Liang, Y.; She, Q.; Peng, N. Coupling transcriptional activation of CRISPR-Cas system and DNA repair genes by Csa3a in Sulfolobus islandicus. Nucleic Acids Res. 2017, 45, 8978–8992. [Google Scholar] [CrossRef]
- Li, M.; Wang, R.; Zhao, D.; Xiang, H. Adaptation of the Haloarcula hispanica CRISPR-Cas system to a purified virus strictly requires a priming process. Nucleic Acids Res. 2014, 42, 2483–2492. [Google Scholar] [CrossRef]
- Counts, J.A.; Willard, D.J.; Kelly, R.M. Life in hot acid: A genome-based reassessment of the archaeal order Sulfolobales. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Dillard, K.E.; Brown, M.W.; Johnson, N.V.; Xiao, Y.; Dolan, A.; Hernandez, E.; Dahlhauser, S.D.; Kim, Y.; Myler, L.R.; Anslyn, E.V.; et al. Assembly and Translocation of a CRISPR-Cas Primed Acquisition Complex. Cell 2018, 175, 934–946. [Google Scholar] [CrossRef] [Green Version]
- Staals, R.H.J.; Jackson, S.A.; Biswas, A.; Brouns, S.J.J.; Brown, C.M.; Fineran, P.C. Interference-driven spacer acquisition is dominant over naive and primed adaptation in a native CRISPR-Cas system. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mohr, G.; Silas, S.; Stamos, J.L.; Makarova, K.S.; Markham, L.M.; Yao, J.; Lucas-Elío, P.; Sanchez-Amat, A.; Fire, A.Z.; Koonin, E.V.; et al. A Reverse Transcriptase-Cas1 Fusion Protein Contains a Cas6 Domain Required for Both CRISPR RNA Biogenesis and RNA Spacer Acquisition. Mol. Cell 2018, 72, 700–714. [Google Scholar] [CrossRef] [Green Version]
- González-Delgado, A.; Mestre, M.R.; Martínez-Abarca, F.; Toro, N. Spacer acquisition from RNA mediated by a natural reverse transcriptase-Cas1 fusion protein associated with a type III-D CRISPR–Cas system in Vibrio vulnificus. Nucleic Acids Res. 2019, 47, 10202–10211. [Google Scholar] [CrossRef]
- Toro, N.; Mestre, M.R.; Martínez-Abarca, F.; González-Delgado, A. Recruitment of Reverse Transcriptase-Cas1 Fusion Proteins by Type VI-A CRISPR-Cas Systems. Front. Microbiol. 2019, 10, 2160. [Google Scholar] [CrossRef]
- Özcan, A.; Pausch, P.; Linden, A.; Wulf, A.; Schühle, K.; Heider, J.; Urlaub, H.; Heimerl, T.; Bange, G.; Randau, L. Type IV CRISPR RNA processing and effector complex formation in Aromatoleum aromaticum. Nat. Microbiol. 2019, 4, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Taylor, H.N.; Warner, E.E.; Armbrust, M.J.; Crowley, V.M.; Olsen, K.J.; Jackson, R.N. Structural basis of Type IV CRISPR RNA biogenesis by a Cas6 endoribonuclease. RNA Biol. 2019, 16, 1438–1447. [Google Scholar] [CrossRef]
- Carte, J.; Wang, R.; Li, H.; Terns, R.M.; Terns, M.P. Cas6 is an endoribonuclease that generates guide RNAs for invader defense in prokaryotes. Genes Dev. 2008, 3489–3496. [Google Scholar] [CrossRef] [Green Version]
- Haurwitz, R.E.; Jinek, M.; Wiedenheft, B.; Zhou, K.; Doudna, J.A. Sequence- and structure-specific RNA processing by a CRISPR endonuclease. Science 2010, 329, 1355–1358. [Google Scholar] [CrossRef] [Green Version]
- Przybilski, R.; Richter, C.; Gristwood, T.; Clulow, J.S.; Vercoe, R.B.; Fineran, P.C. Csy4 is responsible for CRISPR RNA processing in Pectobacterium atrosepticum. RNA Biol. 2011, 8, 517–528. [Google Scholar] [CrossRef] [Green Version]
- Sokolowski, R.D.; Graham, S.; White, M.F. Cas6 specificity and CRISPR RNA loading in a complex CRISPR-Cas system. Nucleic Acids Res. 2014, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Garside, E.L.; Schellenberg, M.J.; Gesner, E.M.; Bonanno, J.B.; Sauder, J.M.; Burley, S.K.; Almo, S.C.; Mehta, G.; MacMillan, A.M. Cas5d processes pre-crRNA and is a member of a larger family of CRISPR RNA endonucleases. RNA 2012, 18, 2020–2028. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Li, H. Recognition and cleavage of a nonstructured CRISPR RNA by its processing endoribonuclease Cas6. Structure 2013, 21, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Behler, J.; Hess, W.R. Approaches to study CRISPR RNA biogenesis and the key players involved. Methods 2020, 172, 12–26. [Google Scholar] [CrossRef]
- Fonfara, I.; Richter, H.; Bratovič, M.; Le Rhun, A.; Charpentier, E. The CRISPR-associated DNA-cleaving enzyme Cpf1 also processes precursor CRISPR RNA. Nature 2016, 532, 517–521. [Google Scholar] [CrossRef]
- East-Seletsky, A.; O’Connell, M.R.; Knight, S.C.; Burstein, D.; Cate, J.H.D.; Tjian, R.; Doudna, J.A. Two distinct RNase activities of CRISPR-C2c2 enable guide-RNA processing and RNA detection. Nature 2016, 538, 270–273. [Google Scholar] [CrossRef]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR–Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [Green Version]
- McBride, T.; Schwartz, E.; Kumar, A.; Taylor, D.; Fineran, P.; Fagerlund, R. Diverse CRISPR-Cas complexes require independent translation of small and large subunits from a single gene. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gleditzsch, D.; Pausch, P.; Müller-Esparza, H.; Özcan, A.; Guo, X.; Bange, G.; Randau, L. PAM identification by CRISPR-Cas effector complexes: Diversified mechanisms and structures. RNA Biol. 2019, 16, 504–517. [Google Scholar] [CrossRef]
- Leenay, R.T.; Beisel, C.L. Deciphering, Communicating, and Engineering the CRISPR PAM. J. Mol. Biol. 2017, 429, 177–191. [Google Scholar] [CrossRef] [Green Version]
- Westra, E.R.; Semenova, E.; Datsenko, K.A.; Jackson, R.N.; Wiedenheft, B.; Severinov, K.; Brouns, S.J.J. Type I-E CRISPR-Cas Systems Discriminate Target from Non-Target DNA through Base Pairing-Independent PAM Recognition. PLoS Genet. 2013, 9, e1003742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westra, E.R.; van Erp, P.B.G.; Künne, T.; Wong, S.P.; Staals, R.H.J.; Seegers, C.L.C.; Bollen, S.; Jore, M.M.; Semenova, E.; Severinov, K.; et al. CRISPR Immunity Relies on the Consecutive Binding and Degradation of Negatively Supercoiled Invader DNA by Cascade and Cas3. Mol. Cell 2012, 46, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Redding, S.; Sternberg, S.H.; Marshall, M.; Gibb, B.; Bhat, P.; Guegler, C.K.; Wiedenheft, B.; Doudna, J.A.; Greene, E.C. Surveillance and Processing of Foreign DNA by the Escherichia coli CRISPR-Cas System. Cell 2015, 163, 854–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jore, M.M.; Lundgren, M.; van Duijn, E.; Bultema, J.B.; Westra, E.R.; Waghmare, S.P.; Wiedenheft, B.; Pul, U.; Wurm, R.; Wagner, R.; et al. Structural basis for CRISPR RNA-guided DNA recognition by Cascade. Nat. Struct. Mol. Biol. 2011, 18, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Sinkunas, T.; Gasiunas, G.; Fremaux, C.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas3 is a single-stranded DNA nuclease and ATP-dependent helicase in the CRISPR/Cas immune system. EMBO J. 2011, 30, 1335–1342. [Google Scholar] [CrossRef] [Green Version]
- Mulepati, S.; Bailey, S. Structural and biochemical analysis of nuclease domain of clustered regularly interspaced short palindromic repeat (CRISPR)-associated protein 3 (Cas3). J. Biol. Chem. 2011, 286, 31896–31903. [Google Scholar] [CrossRef] [Green Version]
- Brouns, S.J.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.H.; Snijders, A.P.L.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; Van Der Oost, J. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Luo, M.; Dolan, A.E.; Liao, M.; Ke, A. Structure basis for RNA-guided DNA degradation by Cascade and Cas3. Science 2018, 361, eaat0839. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, S.; Terns, M.P. CRISPR RNA-guided DNA cleavage by reconstituted Type I-A immune effector complexes. Extremophiles 2019, 23, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Majumdar, S.; Ligon, M.; Skinner, W.C.; Terns, R.M.; Terns, M.P. Target DNA recognition and cleavage by a reconstituted Type I-G CRISPR-Cas immune effector complex. Extremophiles 2017, 21, 95–107. [Google Scholar] [CrossRef] [Green Version]
- Lintner, N.G.; Kerou, M.; Brumfield, S.K.; Graham, S.; Liu, H.; Naismith, J.H.; Sdano, M.; Peng, N.; She, Q.; Copié, V.; et al. Structural and functional characterization of an archaeal clustered regularly interspaced short palindromic repeat (CRISPR)-associated complex for antiviral defense (CASCADE). J. Biol. Chem. 2011, 286, 21643–21656. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Zetsche, B.; Gootenberg, J.S.; Abudayyeh, O.O.; Slaymaker, I.M.; Makarova, K.S.; Essletzbichler, P.; Volz, S.E.; Joung, J.; Van Der Oost, J.; Regev, A.; et al. Cpf1 Is a Single RNA-Guided Endonuclease of a Class 2 CRISPR-Cas System. Cell 2015, 163, 759–771. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Ma, E.; Harrington, L.B.; Da Costa, M.; Tian, X.; Palefsky, J.M.; Doudna, J.A. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar] [CrossRef] [Green Version]
- Koonin, E.V.; Zhang, F. Coupling immunity and programmed cell suicide in prokaryotes: Life-or-death choices. BioEssays 2017, 39, e201600186. [Google Scholar] [CrossRef] [Green Version]
- Crowley, V.M.; Catching, A.; Taylor, H.N.; Borges, A.L.; Metcalf, J.; Bondy-Denomy, J.; Jackson, R.N. A Type IV-A CRISPR-Cas System in Pseudomonas aeruginosa Mediates RNA-Guided Plasmid Interference In Vivo. Cris. J. 2019, 2, 434–440. [Google Scholar] [CrossRef]
- Pinilla-Redondo, R.; Mayo-Muñoz, D.; Russel, J.; Garrett, R.A.; Randau, L.; Sørensen, S.J.; Shah, S.A. Type IV CRISPR-Cas systems are highly diverse and involved in competition between plasmids. Nucleic Acids Res. 2020, 48, 2000–2012. [Google Scholar] [CrossRef] [Green Version]
- Harrington, L.B.; Burstein, D.; Chen, J.S.; Paez-Espino, D.; Ma, E.; Witte, I.P.; Cofsky, J.C.; Kyrpides, N.C.; Banfield, J.F.; Doudna, J.A. Programmed DNA destruction by miniature CRISPR-Cas14 enzymes. Science 2018, 362, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Burstein, D.; Harrington, L.B.; Strutt, S.C.; Probst, A.J.; Anantharaman, K.; Thomas, B.C.; Doudna, J.A.; Banfield, J.F. New CRISPR-Cas systems from uncultivated microbes. Nature 2017, 542, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Smargon, A.A.; Cox, D.B.T.; Pyzocha, N.K.; Zheng, K.; Slaymaker, I.M.; Gootenberg, J.S.; Abudayyeh, O.A.; Essletzbichler, P.; Shmakov, S.; Makarova, K.S.; et al. Cas13b Is a Type VI-B CRISPR-Associated RNA-Guided RNase Differentially Regulated by Accessory Proteins Csx27 and Csx28. Mol. Cell 2017, 65, 618–630. [Google Scholar] [CrossRef] [Green Version]
- Yan, W.X.; Chong, S.; Zhang, H.; Makarova, K.S.; Koonin, E.V.; Cheng, D.R.; Scott, D.A. Cas13d Is a Compact RNA-Targeting Type VI CRISPR Effector Positively Modulated by a WYL-Domain-Containing Accessory Protein. Mol. Cell 2018, 70, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Ye, Y.; Ye, W.; Perčulija, V.; Jiang, H.; Chen, Y.; Li, Y.; Chen, J.; Lin, J.; Wang, S.; et al. Two HEPN domains dictate CRISPR RNA maturation and target cleavage in Cas13d. Nat. Commun. 2019, 10, 2544. [Google Scholar] [CrossRef] [Green Version]
- Abudayyeh, O.O.; Gootenberg, J.S.; Essletzbichler, P.; Han, S.; Joung, J.; Belanto, J.J.; Verdine, V.; Cox, D.B.T.; Kellner, M.J.; Regev, A.; et al. RNA targeting with CRISPR–Cas13. Nature 2017, 550, 280–284. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Li, X.; Ma, J.; Li, Z.; You, L.; Wang, J.; Wang, M.; Zhang, X.; Wang, Y. The Molecular Architecture for RNA-Guided RNA Cleavage by Cas13a. Cell 2017, 170, 714–726. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patchsung, M.; Jantarug, K.; Pattama, A.; Aphicho, K.; Suraritdechachai, S.; Meesawat, P.; Sappakhaw, K.; Leelahakorn, N.; Ruenkam, T.; Wongsatit, T.; et al. Clinical validation of a Cas13-based assay for the detection of SARS-CoV-2 RNA. Nat. Biomed. Eng. 2020. [Google Scholar] [CrossRef]
- Liu, L.; Li, X.; Wang, J.; Wang, M.; Chen, P.; Yin, M.; Li, J.; Sheng, G.; Wang, Y. Two Distant Catalytic Sites Are Responsible for C2c2 RNase Activities. Cell 2017, 168, 121–134. [Google Scholar] [CrossRef] [Green Version]
- Meeske, A.J.; Nakandakari-Higa, S.; Marraffini, L.A. Cas13-induced cellular dormancy prevents the rise of CRISPR-resistant bacteriophage. Nature 2019, 570, 241–245. [Google Scholar] [CrossRef]
- Rouillon, C.; Zhou, M.; Zhang, J.; Politis, A.; Beilsten-Edmands, V.; Cannone, G.; Graham, S.; Robinson, C.V.; Spagnolo, L.; White, M.F. Structure of the CRISPR interference complex CSM reveals key similarities with cascade. Mol. Cell 2013, 52, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Rouillon, C.; Kerou, M.; Reeks, J.; Brugger, K.; Reimann, J.; Cannone, G.; Liu, H.; Albers, S.; Naismith, H.; et al. Structure and mechanism of the CMR complex for CRISPR- mediated antiviral immunity. Mol. Cell 2012, 45, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Vestergaard, G.; Garrett, R.A.; Shah, S. CRISPR adaptive immune systems of Archaea. RNA Biol. 2014, 11, 156–167. [Google Scholar] [CrossRef] [Green Version]
- Pan, S.; Li, Q.; Deng, L.; Jiang, S.; Jin, X.; Peng, N.; Liang, Y.; She, Q.; Li, Y. A seed motif for target RNA capture enables efficient immune defence by a type III-B CRISPR-Cas system. RNA Biol. 2019, 16, 1166–1178. [Google Scholar] [CrossRef] [PubMed]
- Sofos, N.; Feng, M.; Stella, S.; Pape, T.; Fuglsang, A.; Lin, J.; Huang, Q.; Li, Y.; She, Q.; Montoya, G. Structures of the Cmr-β Complex Reveal the Regulation of the Immunity Mechanism of Type III-B CRISPR-Cas. Mol. Cell 2020. [Google Scholar] [CrossRef] [PubMed]
- Spilman, M.; Cocozaki, A.; Hale, C.; Shao, Y.; Ramia, N.; Terns, R.; Terns, M.; Li, H.; Stagg, S. Structure of an RNA silencing complex of the CRISPR-Cas immune system. Mol. Cell 2013, 52, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Staals, R.H.J.; Agari, Y.; Maki-Yonekura, S.; Zhu, Y.; Taylor, D.W.; van Duijn, E.; Barendregt, A.; Vlot, M.; Koehorst, J.J.; Sakamoto, K.; et al. Structure and Activity of the RNA-Targeting Type III-B CRISPR-Cas Complex of Thermus thermophilus. Mol. Cell 2013, 52, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Osawa, T.; Inanaga, H.; Sato, C.; Numata, T. Crystal Structure of the CRISPR-Cas RNA Silencing Cmr Complex Bound to a Target Analog. Mol. Cell 2015, 58, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Marraffini, L.A.; Sontheimer, E.J. Self versus non-self discrimination during CRISPR RNA-directed immunity. Nature 2010, 463, 568–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR Interference Limits Horizontal Targeting DNA. Science 2008, 322, 1843–1845. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Zhao, P.; Olson, S.; Duff, M.O.; Graveley, B.R.; Wells, L.; Terns, R.M.; Terns, M.P. RNA-guided RNA cleavage by a CRISPR RNA-Cas protein complex. Cell 2009, 139, 945–956. [Google Scholar] [CrossRef] [Green Version]
- Staals, R.H.J.; Zhu, Y.; Taylor, D.W.; Kornfeld, J.E.; Sharma, K.; Barendregt, A.; Koehorst, J.J.; Vlot, M.; Neupane, N.; Varossieau, K.; et al. RNA Targeting by the Type III-A CRISPR-Cas Csm Complex of Thermus thermophilus. Mol. Cell 2014, 56, 518–530. [Google Scholar] [CrossRef] [Green Version]
- Tamulaitis, G.; Kazlauskiene, M.; Manakova, E.; Venclovas, Č.; Nwokeoji, A.O.; Dickman, M.J.; Horvath, P.; Siksnys, V. Programmable RNA Shredding by the Type III-A CRISPR-Cas System of Streptococcus thermophilus. Mol. Cell 2014, 56, 506–517. [Google Scholar] [CrossRef] [Green Version]
- Estrella, M.A.; Kuo, F.-T.; Bailey, S. RNA-activated DNA cleavage by the Type III-B CRISPR–Cas effector complex. Genes Dev. 2016, 30, 460–470. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Graham, S.; Tello, A.; Liu, H.; White, M.F. Multiple nucleic acid cleavage modes in divergent type III CRISPR systems. Nucleic Acids Res. 2016. [Google Scholar] [CrossRef] [Green Version]
- Benda, C.; Ebert, J.; Scheltema, R.A.; Schiller, H.B.; Baumgärtner, M.; Bonneau, F.; Mann, M.; Conti, E. Structural Model of a CRISPR RNA-Silencing Complex Reveals the RNA-Target Cleavage Activity in Cmr4. Mol. Cell 2014, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Cocozaki, A.; Li, H.; Terns, R.M.; Terns, M.P. Target RNA capture and cleavage by the Cmr type III-B CRISPR–Cas effector complex. Genes Dev. 2014, 28, 2432–2443. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Ye, K. Cmr4 is the slicer in the RNA-targeting Cmr CRISPR complex. Nucleic Acids Res. 2014, 43, 1257–1267. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Feng, M.; Feng, X.; Liang, Y.X.; She, Q. An archaeal CRISPR type III-B system exhibiting distinctive RNA targeting features and mediating dual RNA and DNA interference. Nucleic Acids Res. 2015, 43, 406–417. [Google Scholar] [CrossRef]
- Zebec, Z.; Manica, A.; Zhang, J.; White, M.F.; Schleper, C. CRISPR-mediated targeted mRNA degradation in the archaeon Sulfolobus solfataricus. Nucleic Acids Res. 2014, 42, 5280–5288. [Google Scholar] [CrossRef] [Green Version]
- Deng, L.; Garrett, R.A.; Shah, S.A.; Peng, X.; She, Q. A novel interference mechanism by a type IIIB CRISPR-Cmr module in Sulfolobus. Mol. Microbiol. 2013, 87, 1088–1099. [Google Scholar] [CrossRef]
- Goldberg, G.W.; Jiang, W.; Bikard, D.; Marraffini, L. A Conditional tolerance of temperate phages via transcription-dependent CRISPR-Cas targeting. Nature 2014, 514, 633–637. [Google Scholar] [CrossRef] [Green Version]
- Elmore, J.R.; Sheppard, N.F.; Ramia, N.; Deighan, T.; Li, H.; Terns, R.M.; Terns, M.P. Bipartite recognition of target RNAs activates DNA cleavage by the Type III-B CRISPR–Cas system. Genes Dev. 2016, 30, 447–459. [Google Scholar] [CrossRef] [Green Version]
- Samai, P.; Pyenson, N.; Jiang, W.; Goldberg, G.W.; Hatoum-Aslan, A.; Marraffini, L.A. Co-transcriptional DNA and RNA Cleavage during Type III CRISPR-Cas Immunity. Cell 2015, 161, 1164–1174. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Li, Y.; Deng, L.; Feng, M.; Peng, W.; Hallstrøm, S.; Zhang, J.; Peng, N.; Liang, Y.X.; White, M.F.; et al. A type III-B CRISPR-Cas effector complex mediating massive target DNA destruction. Nucleic Acids Res. 2016, gkw1274. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Ma, J.; Wang, J.; Artamonova, D.; Wang, M.; Liu, L.; Xiang, H.; Severinov, K.; Zhang, X.; Wang, Y. Structure Studies of the CRISPR-Csm Complex Reveal Mechanism of Co-transcriptional Interference. Cell 2019, 176, 239–253. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.-Y.; An, Y.; Park, K.-H.; Lee, M.-H.; Oh, B.-H.; Woo, E. Crystal structure of the Csm1 subunit of the Csm complex and its single-stranded DNA-specific nuclease activity. Structure 2015, 23, 782–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazlauskiene, M.; Tamulaitis, G.; Kostiuk, G.; Venclovas, Č.; Siksnys, V. Spatiotemporal Control of Type III-A CRISPR-Cas Immunity: Coupling DNA Degradation with the Target RNA Recognition. Mol. Cell 2016, 62, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.Y.; Liu, J.-J.; Aditham, A.J.; Nogales, E.; Doudna, J.A. Target preference of Type III-A CRISPR-Cas complexes at the transcription bubble. Nat. Commun. 2019, 10, 3001. [Google Scholar] [CrossRef]
- Manica, A.; Zebec, Z.; Steinkellner, J.; Schleper, C. Unexpectedly broad target recognition of the CRISPR-mediated virus defence system in the archaeon Sulfolobus solfataricus. Nucleic Acids Res. 2013, 1–9. [Google Scholar] [CrossRef]
- Makarova, K.S.; Anantharaman, V.; Grishin, N.V.; Koonin, E.V.; Aravind, L. CARF and WYL domains: Ligand-binding regulators of prokaryotic defense systems. Front. Genet. 2014, 5, 102. [Google Scholar] [CrossRef]
- Niewoehner, O.; Jinek, M. Structural basis for the endoribonuclease activity of the type III-A CRISPR-associated protein Csm6. RNA 2016, 22, 318–329. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, N.F.; Glover, C.V.C.; Terns, R.M.; Terns, M.P. The CRISPR-associated Csx1 protein of Pyrococcus furiosus is an adenosine-specific endoribonuclease. RNA 2016, 22, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Pan, S.; López-Méndez, B.; Montoya, G.; She, Q. Allosteric regulation of Csx1, a type IIIB-associated CARF domain ribonuclease by RNAs carrying a tetraadenylate tail. Nucleic Acids Res. 2017, 45, 10740–10750. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Samai, P.; Marraffini, L.A. Degradation of Phage Transcripts by CRISPR-Associated RNases Enables Type III CRISPR-Cas Immunity. Cell 2016, 164, 710–721. [Google Scholar] [CrossRef] [Green Version]
- Niewoehner, O.; Garcia-Doval, C.; Rostøl, J.T.; Berk, C.; Schwede, F.; Bigler, L.; Hall, J.; Marraffini, L.A.; Jinek, M. Type III CRISPR–Cas systems produce cyclic oligoadenylate second messengers. Nature 2017, 548, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskiene, M.; Kostiuk, G.; Venclovas, Č.; Tamulaitis, G.; Siksnys, V. A cyclic oligonucleotide signaling pathway in type III CRISPR-Cas systems. Science 2017, 357, 605–609. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Horvath, P.; Moineau, S.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; et al. Evolution and classification of the CRISPR-Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef] [Green Version]
- Rouillon, C.; Athukoralage, J.S.; Graham, S.; Grüschow, S.; White, M.F. Control of cyclic oligoadenylate synthesis in a type III CRISPR system. Elife 2018, 7. [Google Scholar] [CrossRef]
- Molina, R.; Stella, S.; Feng, M.; Sofos, N.; Jauniskis, V.; Pozdnyakova, I.; López-Méndez, B.; She, Q.; Montoya, G. Structure of Csx1-cOA4 complex reveals the basis of RNA decay in Type III-B CRISPR-Cas. Nat. Commun. 2019, 10, 4302. [Google Scholar] [CrossRef]
- Grüschow, S.; Athukoralage, J.S.; Graham, S.; Hoogeboom, T.; White, M.F. Cyclic oligoadenylate signalling mediates Mycobacterium tuberculosis CRISPR defence. Nucleic Acids Res. 2019, 47, 9259–9270. [Google Scholar] [CrossRef]
- Han, W.; Stella, S.; Zhang, Y.; Guo, T.; Sulek, K.; Peng-Lundgren, L.; Montoya, G.; She, Q. A Type III-B Cmr effector complex catalyzes the synthesis of cyclic oligoadenylate second messengers by cooperative substrate binding. Nucleic Acids Res. 2018, 46, 10319–10330. [Google Scholar] [CrossRef] [Green Version]
- McMahon, S.A.; Zhu, W.; Graham, S.; Rambo, R.; White, M.F.; Gloster, T.M. Structure and mechanism of a Type III CRISPR defence DNA nuclease activated by cyclic oligoadenylate. Nat. Commun. 2020, 11, 14–17. [Google Scholar] [CrossRef] [Green Version]
- Rostøl, J.T.; Marraffini, L.A. Non-specific degradation of transcripts promotes plasmid clearance during type III-A CRISPR–Cas immunity. Nat. Microbiol. 2019, 4, 656–662. [Google Scholar] [CrossRef]
- Athukoralage, J.S.; Rouillon, C.; Graham, S.; Grüschow, S.; White, M.F. Ring nucleases deactivate type III CRISPR ribonucleases by degrading cyclic oligoadenylate. Nature 2018, 562, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Athukoralage, J.S.; McQuarrie, S.; Grüschow, S.; Graham, S.; Gloster, T.M.; White, M.F. Tetramerisation of the crispr ring nuclease crn3/csx3 facilitates cyclic oligoadenylate cleavage. Elife 2020, 9, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Jones, R.; Yang, G.; Ouerfelli, O.; Patel, D.J. CRISPR-Cas III-A Csm6 CARF Domain Is a Ring Nuclease Triggering Stepwise cA4 Cleavage with ApA > p Formation Terminating RNase Activity. Mol. Cell 2019, 75, 944–956. [Google Scholar] [CrossRef]
- Foster, K.; Grüschow, S.; Bailey, S.; White, M.F.; Terns, M.P. Regulation of the RNA and DNA nuclease activities required for Pyrococcus furiosus Type III-B CRISPR-Cas immunity. Nucleic Acids Res. 2020, 48, 4418–4434. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Doval, C.; Schwede, F.; Berk, C.; Rostøl, J.T.; Niewoehner, O.; Tejero, O.; Hall, J.; Marraffini, L.A.; Jinek, M. Activation and self-inactivation mechanisms of the cyclic oligoadenylate-dependent CRISPR ribonuclease Csm6. Nat. Commun. 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athukoralage, J.S.; Graham, S.; Grüschow, S.; Rouillon, C.; White, M.F. A Type III CRISPR Ancillary Ribonuclease Degrades Its Cyclic Oligoadenylate Activator. J. Mol. Biol. 2019, 431, 2894–2899. [Google Scholar] [CrossRef]
- Zhao, R.; Yang, Y.; Zheng, F.; Zeng, Z.; Feng, W.; Jin, X.; Wang, J.; Yang, K.; Liang, Y.X.; She, Q.; et al. A Membrane-Associated DHH-DHHA1 Nuclease Degrades Type III CRISPR Second Messenger. Cell Rep. 2020, 32. [Google Scholar] [CrossRef]
- Athukoralage, J.S.; McMahon, S.A.; Zhang, C.; Grüschow, S.; Graham, S.; Krupovic, M.; Whitaker, R.J.; Gloster, T.M.; White, M.F. An anti-CRISPR viral ring nuclease subverts type III CRISPR immunity. Nature 2020, 577, 572–575. [Google Scholar] [CrossRef]
- Bhoobalan-Chitty, Y.; Johansen, T.B.; Di Cianni, N.; Peng, X. Inhibition of Type III CRISPR-Cas Immunity by an Archaeal Virus-Encoded Anti-CRISPR Protein. Cell 2019, 179, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Jia, N.; Mo, C.Y.; Wang, C.; Eng, E.T.; Marraffini, L.A.; Patel, D.J. Type III-A CRISPR-Cas Csm Complexes: Assembly, Periodic RNA Cleavage, DNase Activity Regulation, and Autoimmunity. Mol. Cell 2019, 73, 264–277. [Google Scholar] [CrossRef] [Green Version]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mojica, F.J.M.; Juez, G.; Rodriguez-Valera, F. Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Mol. Microbiol. 1993, 9, 613–621. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Ferrer, C.; Juez, G.; Rodríguez-Valera, F. Long stretches of short tandem repeats are present in the largest replicons of the Archaea Haloferax mediterranei and Haloferax volcanii and could be involved in replicon partitioning. Mol. Microbiol. 1995, 17, 85–93. [Google Scholar] [CrossRef]
- Mojica, F.J.M.; Rodriguez-Valera, F. The discovery of CRISPR in archaea and bacteria. FEBS J. 2016, 283, 3162–3169. [Google Scholar] [CrossRef] [PubMed]
- Mojica, F.J.M.; Díez-Villaseñor, C.; García-Martínez, J.; Soria, E. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 2005, 60, 174–182. [Google Scholar] [CrossRef]
- Brendel, J.; Stoll, B.; Lange, S.J.; Sharma, K.; Lenz, C.; Stachler, A.E.; Maier, L.K.; Richter, H.; Nickel, L.; Schmitz, R.A.; et al. A complex of cas proteins 5, 6, and 7 is required for the biogenesis and stability of clustered regularly interspaced short palindromic repeats (CRISPR)-derived RNAs (crRNAs) in haloferax volcanii. J. Biol. Chem. 2014, 289, 7164–7177. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liu, H.; Han, J.; Liu, J.; Wang, R.; Zhao, D.; Zhou, J.; Xiang, H. Characterization of CRISPR RNA biogenesis and Cas6 cleavage-mediated inhibition of a provirus in the haloarchaeon Haloferax mediterranei. J. Bacteriol. 2013, 195, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Fischer, S.; Maier, L.K.; Stoll, B.; Brendel, J.; Fischer, E.; Pfeiffer, F.; Dyall-Smith, M.; Marchfelder, A. An archaeal immune system can detect multiple protospacer adjacent motifs (PAMs) to target invader DNA. J. Biol. Chem. 2012, 287, 33351–33365. [Google Scholar] [CrossRef] [Green Version]
- Maier, L.K.; Lange, S.J.; Stoll, B.; Haas, K.A.; Fischer, S.; Fischer, E.; Duchardt-Ferner, E.; Wöhnert, J.; Backofen, R.; Marchfelder, A. Essential requirements for the detection and degradation of invaders by the Haloferax volcanii CRISPR/Cas system I-B. RNA Biol. 2013, 10, 865–874. [Google Scholar] [CrossRef] [Green Version]
- Cass, S.D.B.; Haas, K.A.; Stoll, B.; Alkhnbashi, O.S.; Sharma, K.; Urlaub, H.; Backofen, R.; Marchfelder, A.; Bolt, E.L. The role of Cas8 in type I CRISPR interference. Biosci. Rep. 2015, 35. [Google Scholar] [CrossRef]
- Li, M.; Wang, R.; Xiang, H. Haloarcula hispanica CRISPR authenticates PAM of a target sequence to prime discriminative adaptation. Nucleic Acids Res. 2014, 42, 7226–7235. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, M.; Gong, L.; Hu, S.; Xiang, H. DNA motifs determining the accuracy of repeat duplication during CRISPR adaptation in Haloarcula hispanica. Nucleic Acids Res. 2016, 44, 4266–4277. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Gong, L.; Zhao, D.; Zhou, J.; Xiang, H. The spacer size of I-B CRISPR is modulated by the terminal sequence of the protospacer. Nucleic Acids Res. 2017, 45, 4642–4654. [Google Scholar] [CrossRef] [Green Version]
- Gong, L.; Li, M.; Cheng, F.; Zhao, D.; Chen, Y.; Xiang, H. Primed adaptation tolerates extensive structural and size variations of the CRISPR RNA guide in Haloarcula hispanica. Nucleic Acids Res. 2019, 47, 5880–5891. [Google Scholar] [CrossRef]
- Stachler, A.-E.; Turgeman-Grott, I.; Shtifman-Segal, E.; Allers, T.; Marchfelder, A.; Gophna, U. High tolerance to self-targeting of the genome by the endogenous CRISPR-Cas system in an archaeon. Nucleic Acids Res. 2017, 45, 5208–5216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachler, A.-E.; Wörtz, J.; Alkhnbashi, O.S.; Turgeman-Grott, I.; Smith, R.; Allers, T.; Backofen, R.; Gophna, U.; Marchfelder, A. Adaptation induced by self-targeting in a type I-B CRISPR-Cas system. J. Biol. Chem. 2020. [Google Scholar] [CrossRef]
- Turgeman-Grott, I.; Joseph, S.; Marton, S.; Eizenshtein, K.; Naor, A.; Soucy, S.M.; Stachler, A.E.; Shalev, Y.; Zarkor, M.; Reshef, L.; et al. Pervasive acquisition of CRISPR memory driven by inter-species mating of archaea can limit gene transfer and influence speciation. Nat. Microbiol. 2019, 4, 177–186. [Google Scholar] [CrossRef]
- Naor, A.; Lapierre, P.; Mevarech, M.; Papke, R.T.; Gophna, U. Low species barriers in halophilic archaea and the formation of recombinant hybrids. Curr. Biol. 2012, 22, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.; Kleppe, K.; Terns, R.M.; Terns, M.P. Prokaryotic silencing (psi)RNAs in Pyrococcus furiosus. RNA 2008, 14, 2572–2579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carte, J.; Pfister, N.T.; Compton, M.M.; Terns, R.M.; Terns, M.P. Binding and cleavage of CRISPR RNA by Cas6. RNA 2010, 16, 2181–2188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Preamplume, G.; Terns, M.P.; Terns, R.M.; Li, H. Interaction of the Cas6 riboendonuclease with CRISPR RNAs: Recognition and cleavage. Structure 2011, 19, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Hale, C.R.; Majumdar, S.; Elmore, J.; Pfister, N.; Compton, M.; Olson, S.; Resch, A.M.; Glover, C.V.C.; Graveley, B.R.; Terns, R.M.; et al. Essential features and rational design of CRISPR RNAs that function with the Cas RAMP module complex to cleave RNAs. Mol. Cell 2012, 45, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Terns, R.M.; Terns, M.P. The RNA-and DNA-targeting CRISPR-Cas immune systems of Pyrococcus furiosus. Biochem. Soc. Trans. 2013, 41, 1416–1421. [Google Scholar] [CrossRef] [Green Version]
- Ramia, N.F.; Spilman, M.; Li, H.; Stagg, S.M.; Ramia, N.F.; Spilman, M.; Tang, L.; Shao, Y.; Elmore, J.; Hale, C.; et al. Essential Structural and Functional Roles of the Cmr4 Subunit in RNA Cleavage by the Cmr CRISPR- Cas Complex Report Essential Structural and Functional Roles of the Cmr4 Subunit in RNA Cleavage by the Cmr CRISPR-Cas Complex. Cell Rep. 2014, 9, 1610–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.K.; Kim, Y.G.; Oh, B.H. Crystal structure and nucleic acid-binding activity of the CRISPR-Associated protein Csx1 of Pyrococcus furiosus. Proteins Struct. Funct. Bioinf. 2013, 81, 261–270. [Google Scholar] [CrossRef]
- Elmore, J.; Deighan, T.; Westpheling, J.; Terns, R.M.; Terns, M.P. DNA targeting by the type I-G and type I-A CRISPR-Cas systems of Pyrococcus furiosus. Nucleic Acids Res. 2015, 43, 10353–10363. [Google Scholar] [CrossRef] [Green Version]
- Grainy, J.; Garrett, S.; Graveley, B.R.; Terns, M.P. CRISPR repeat sequences and relative spacing specify DNA integration by Pyrococcus furiosus Cas1 and Cas2. Nucleic Acids Res. 2019, 47, 7518–7531. [Google Scholar] [CrossRef]
- Garrett, S.; Shiimori, M.; Watts, E.A.; Clark, L.; Graveley, B.R.; Terns, M.P. Primed CRISPR DNA uptake in Pyrococcus furiosus. Nucleic Acids Res. 2020, 48, 6120–6135. [Google Scholar] [CrossRef]
- Beloglazova, N.; Petit, P.; Flick, R.; Brown, G.; Savchenko, A.; Yakunin, A.F. Structure and activity of the Cas3 HD nuclease MJ0384, an effector enzyme of the CRISPR interference. EMBO J. 2011, 30, 4616–4627. [Google Scholar] [CrossRef]
- Hrle, A.; Su, A.A.H.; Ebert, J.; Benda, C.; Randau, L.; Conti, E. Structure and RNA-binding properties of the type III-A CRISPR-associated protein Csm3. RNA Biol. 2013, 10, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numata, T.; Inanaga, H.; Sato, C.; Osawa, T. Crystal structure of the Csm3-Csm4 subcomplex in the type III-A CRISPR-Cas interference complex. J. Mol. Biol. 2015, 427, 259–273. [Google Scholar] [CrossRef]
- Guy, C.P.; Majerník, A.I.; Chong, J.P.J.; Bolt, E.L. A novel nuclease-ATPase (Nar71) from archaea is part of a proposed thermophilic DNA repair system. Nucleic Acids Res. 2004, 32, 6176–6186. [Google Scholar] [CrossRef] [Green Version]
- Richter, H.; Zoephel, J.; Schermuly, J.; Maticzka, D.; Backofen, R.; Randau, L. Characterization of CRISPR RNA processing in Clostridium thermocellum and Methanococcus maripaludis. Nucleic Acids Res. 2012, 40, 9887–9896. [Google Scholar] [CrossRef] [Green Version]
- Richter, H.; Lange, S.J.; Backofen, R.; Randau, L. SF CRISPR: Comparative analysis of Cas6b processing and CRISPR RNA stability. RNA Biol. 2013, 10, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Nickel, L.; Weidenbach, K.; Jäger, D.; Backofen, R.; Lange, S.J.; Heidrich, N.; Schmitz, R.A. Two CRISPR-Cas systems in Methanosarcina mazei strain Gö1 display common processing features despite belonging to different types I and III. RNA Biol. 2013, 10, 779–791. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Richter, H.; Sun, S.; Sharma, K.; Urlaub, H.; Randau, L.; Li, H. A Non-Stem-Loop CRISPR RNA Is Processed by Dual Binding Cas6. Structure 2016, 24, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Richter, H.; Rompf, J.; Wiegel, J.; Rau, K.; Randau, L. Fragmentation of the CRISPR-Cas Type I-B signature protein Cas8b. Biochim. Biophys. Gen. Subj. 2017, 1861, 2993–3000. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Shmakov, S.; Makarova, K.S.; Forterre, P.; Koonin, E.V. Recent mobility of casposons, self-synthesizing transposons at the origin of the CRISPR-cas immunity. Genome Biol. Evol. 2016, 8, 375–386. [Google Scholar] [CrossRef]
- Krupovic, M.; Makarova, K.S.; Forterre, P.; Prangishvili, D.; Koonin, E.V. Casposons: A new superfamily of self-synthesizing DNA transposons at the origin of prokaryotic CRISPR-Cas immunity. BMC Biol. 2014, 12, 36. [Google Scholar] [CrossRef]
- Hickman, A.B.; Dyda, F. The casposon-encoded Cas1 protein from Aciduliprofundum boonei is a DNA integrase that generates target site duplications. Nucleic Acids Res. 2015, 43, 10576–10587. [Google Scholar] [CrossRef] [Green Version]
- Hickman, A.B.; Kailasan, S.; Genzor, P.; Haase, A.D.; Dyda, F. Casposase structure and the mechanistic link between DNA transposition and spacer acquisition by CRISPR-Cas. Elife 2020, 9, 1–26. [Google Scholar] [CrossRef]
- Ulbricht, A.; Nickel, L.; Weidenbach, K.; Vargas Gebauer, H.; Kießling, C.; Förstner, K.U.; Schmitz, R.A. The CARF Protein MM_0565 Affects Transcription of the Casposon-Encoded cas1-solo Gene in Methanosarcina mazei Gö1. Biomolecules 2020, 10, 1161. [Google Scholar] [CrossRef]
- Bolduc, B.; Shaughnessy, D.P.; Wolf, Y.I.; Koonin, E.V.; Roberto, F.F.; Young, M. Identification of novel positive-strand RNA viruses by metagenomic analysis of archaea-dominated Yellowstone hot springs. J. Virol. 2012, 86, 5562–5573. [Google Scholar] [CrossRef] [Green Version]
- Weidenbach, K.; Nickel, L.; Neve, H.; Alkhnbashi, O.S.; Künzel, S.; Kupczok, A.; Bauersachs, T.; Cassidy, L.; Tholey, A.; Backofen, R.; et al. Methanosarcina Spherical Virus, a Novel Archaeal Lytic Virus Targeting Methanosarcina Strains. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Garrett, R.; Shah, S.; Erdmann, S.; Liu, G.; Mousaei, M.; León-Sobrino, C.; Peng, W.; Gudbergsdottir, S.; Deng, L.; Vestergaard, G.; et al. CRISPR-Cas Adaptive Immune Systems of the Sulfolobales: Unravelling Their Complexity and Diversity. Life 2015, 5, 783–817. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; White, M.F. Hot and crispy: CRISPR-Cas systems in the hyperthermophile Sulfolobus solfataricus. Biochem. Soc. Trans. 2013, 41, 1422–1426. [Google Scholar] [CrossRef] [Green Version]
- Manica, A.; Schleper, C. CRISPR-mediated defense mechanisms in the hyperthermophilic archaeal genus Sulfolobus. RNA Biol. 2013, 10, 671–678. [Google Scholar] [CrossRef] [Green Version]
- Garrett, R.A.; Shah, S.A.; Vestergaard, G.; Deng, L.; Gudbergsdottir, S.; Kenchappa, C.S.; Erdmann, S.; She, Q. CRISPR-based immune systems of the Sulfolobales: Complexity and diversity. Biochem. Soc. Trans. 2011, 39, 51–57. [Google Scholar] [CrossRef]
- Cannone, G.; Webber-Birungi, M.; Spagnolo, L. Electron microscopy studies of Type III CRISPR machines in Sulfolobus solfataricus. Biochem. Soc. Trans. 2013, 41, 1427–1430. [Google Scholar] [CrossRef] [Green Version]
- Prangishvili, D.; Bamford, D.H.; Forterre, P.; Iranzo, J.; Koonin, E.V.; Krupovic, M. The enigmatic archaeal virosphere. Nat. Rev. Microbiol. 2017, 15, 724–739. [Google Scholar] [CrossRef]
- Gudbergsdottir, S.; Deng, L.; Chen, Z.; Jensen, J.V.K.; Jensen, L.R.; She, Q.; Garrett, R.A. Dynamic properties of the Sulfolobus CRISPR/Cas and CRISPR/Cmr systems when challenged with vector-borne viral and plasmid genes and protospacers. Mol. Microbiol. 2011, 79, 35–49. [Google Scholar] [CrossRef] [Green Version]
- Manica, A.; Zebec, Z.; Teichmann, D.; Schleper, C. In vivo activity of CRISPR-mediated virus defence in a hyperthermophilic archaeon. Mol. Microbiol. 2011, 80, 481–491. [Google Scholar] [CrossRef]
- Jonuscheit, M.; Martusewitsch, E.; Stedman, K.M.; Schleper, C. A reporter gene system for the hyperthermophilic archaeon Sulfolobus solfataricus based on a selectable and integrative shuttle vector. Mol. Microbiol. 2003, 48, 1241–1252. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Li, H.; Hallstrøm, S.; Peng, N.; Liang, Y.X.; She, Q. Genetic determinants of PAM-dependent DNA targeting and pre-crRNA processing in Sulfolobus islandicus. RNA Biol. 2013, 10, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Mousaei, M.; Deng, L.; She, Q.; Garrett, R.A. Major and minor crRNA annealing sites facilitate low stringency DNA protospacer binding prior to Type I-A CRISPR-Cas interference in Sulfolobus. RNA Biol. 2016, 13, 1166–1173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Kenchappa, C.S.; Peng, X.; She, Q.; Garrett, R. a Modulation of CRISPR locus transcription by the repeat-binding protein Cbp1 in Sulfolobus. Nucleic Acids Res. 2012, 40, 2470–2480. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, Y.; Lin, J.; Pan, S.; Han, W.; Peng, N.; Liang, Y.X.; She, Q. Cmr1 enables efficient RNA and DNA interference of a III-B CRISPR-Cas system by binding to target RNA and crRNA. Nucleic Acids Res. 2017, 45, 11305–11314. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, K.; Wada, N.; Murakami, E.; Osakabe, Y. Genome editing in mammals using CRISPR type I-D nuclease. bioRxiv 2020, 991976. [Google Scholar] [CrossRef] [Green Version]
- Fusco, S.; Liguori, R.; Limauro, D.; Bartolucci, S.; She, Q.; Contursi, P. Transcriptome analysis of Sulfolobus solfataricus infected with two related fuselloviruses reveals novel insights into the regulation of CRISPR-Cas system. Biochimie 2015, 118, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Quax, T.E.F.; Voet, M.; Sismeiro, O.; Dillies, M.-A.; Jagla, B.; Coppee, J.-Y.; Sezonov, G.; Forterre, P.; van der Oost, J.; Lavigne, R.; et al. Massive Activation of Archaeal Defense Genes during Viral Infection. J. Virol. 2013, 87, 8419–8428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papathanasiou, P.; Erdmann, S.; Leon-Sobrino, C.; Sharma, K.; Urlaub, H.; Garrett, R.A.; Peng, X. Stable maintenance of the rudivirus SIRV3 in a carrier state in Sulfolobus islandicus despite activation of the CRISPR-Cas immune response by a second virus SMV1. RNA Biol. 2019, 16, 557–565. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.; Han, W.; She, Q. Tolerance of Sulfolobus SMV1 virus to the immunity of I-A and III-B CRISPR-Cas systems in Sulfolobus islandicus. RNA Biol. 2019, 16, 549–556. [Google Scholar] [CrossRef]
- Bautista, M.A.; Zhang, C.; Whitaker, R.J. Virus-induced dormancy in the archaeon Sulfolobus islandicus. MBio 2015, 6. [Google Scholar] [CrossRef]
- Maaty, W.S.; Steffens, J.D.; Heinemann, J.; Ortmann, A.C.; Reeves, B.D.; Biswas, S.K.; Dratz, E.A.; Grieco, P.A.; Young, M.J.; Bothner, B. Global analysis of viral infection in an archaeal model system. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Bhoobalan-chitty, Y.; Van, L.B.; Kjeldsen, A.L.; Dedola, M.; Makarova, K.S.; Koonin, E.V.; Brodersen, D.E.; Peng, X. viruses inhibit subtype I-D immunity. Nat. Microbiol. 2018, 3. [Google Scholar]
- Wang, R.; Zheng, H.; Preamplume, G.; Shao, Y.; Li, H. The impact of CRISPR repeat sequence on structures of a Cas6 protein-RNA complex. Protein Sci. 2012, 21, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Elmore, J.R.; Yokooji, Y.; Sato, T.; Olson, S.; Glover, C.V.C.; Graveley, B.R.; Atomi, H.; Terns, R.M.; Terns, M.P. Programmable plasmid interference by the CRISPR-Cas system in Thermococcus kodakarensis. RNA Biol. 2013, 10, 828–840. [Google Scholar] [CrossRef] [Green Version]
- Park, K.; An, Y.; Jung, T.; Baek, I.; Noh, H.; Ahn, W.; Hebert, H.; Song, J.; Kim, J.; Oh, B.; et al. RNA activation-independent DNA targeting of the Type III CRISPR-Cas system by a Csm complex. EMBO Rep. 2017, 18, 826–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, D.D.; Metcalf, W.W. Cas9-mediated genome editing in the methanogenic archaeon Methanosarcina acetivorans. Proc. Natl. Acad. Sci. USA 2017, 114, 2976–2981. [Google Scholar] [CrossRef] [Green Version]
- Dhamad, A.E.; Lessner, D.J. A CRISPRi-dCas9 system for archaea and its use to examine gene function during nitrogen fixation by Methanosarcina acetivorans. Appl. Environ. Microbiol. 2020. [Google Scholar] [CrossRef]
- Su, A.A.H.; Tripp, V.; Randau, L. RNA-Seq analyses reveal the order of tRNA processing events and the maturation of C/D box and CRISPR RNAs in the hyperthermophile Methanopyrus kandleri. Nucleic Acids Res. 2013, 41, 6250–6258. [Google Scholar] [CrossRef] [Green Version]
- Stachler, A.-E.; Marchfelder, A. Gene Repression in Haloarchaea Using the CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats)-Cas I-B System. J. Biol. Chem. 2016, 291, 15226–15242. [Google Scholar] [CrossRef] [Green Version]
- Cheng, F.; Gong, L.; Zhao, D.; Yang, H.; Zhou, J.; Li, M.; Xiang, H. Harnessing the native type I-B CRISPR-Cas for genome editing in a polyploid archaeon. J. Genet. Genom. 2017, 44, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.Y.; Shin, M.; Huynh Thi Yen, L.; Kim, J.S. Crystal structure of Cas1 from Archaeoglobus fulgidus and characterization of its nucleolytic activity. Biochem. Biophys. Res. Commun. 2013, 441, 720–725. [Google Scholar] [CrossRef]
- Tang, T.H.; Bachellerie, J.P.; Rozhdestvensky, T.; Bortolin, M.L.; Huber, H.; Drungowski, M.; Elge, T.; Brosius, J.; Hüttenhofer, A. Identification of 86 candidates for small non-messenger RNAs from the archaeon Archaeoglobus fulgidus. Proc. Natl. Acad. Sci. USA 2002, 99, 7536–7541. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, S.; Garrett, R.A. Selective and hyperactive uptake of foreign DNA by adaptive immune systems of an archaeon via two distinct mechanisms. Mol. Microbiol. 2012, 85, 1044–1056. [Google Scholar] [CrossRef]
- Rollie, C.; Schneider, S.; Brinkmann, A.S.; Bolt, E.L.; White, M.F. Intrinsic sequence specificity of the Cas1 integrase directs new spacer acquisition. Elife 2015, 4, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.H.; Polacek, N.; Zywicki, M.; Huber, H.; Brugger, K.; Garrett, R.; Bachellerie, J.P.; Hüttenhofer, A. Identification of novel non-coding RNAs as potential antisense regulators in the archaeon Sulfolobus solfataricus. Mol. Microbiol. 2005, 55, 469–481. [Google Scholar] [CrossRef]
- Han, D.; Krauss, G. Characterization of the endonuclease SSO2001 from Sulfolobus solfataricus P2. FEBS Lett. 2009, 583, 771–776. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, S.; Le Moine Bauer, S.; Garrett, R.A. Inter-viral conflicts that exploit host CRISPR immune systems of Sulfolobus. Mol. Microbiol. 2014, 91, 900–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Pan, S.; Zhang, Y.; Ren, M.; Feng, M.; Peng, N.; Chen, L.; Liang, Y.X.; She, Q. Harnessing Type I and Type III CRISPR-Cas systems for genome editing. Nucleic Acids Res. 2015, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayo-Muñoz, D.; He, F.; Jørgensen, J.B.; Madsen, P.K.; Bhoobalan-Chitty, Y.; Peng, X. Anti-crispr-based and crispr-based genome editing of sulfolobus islandicus rod-shaped virus 2. Viruses 2018, 10, 695. [Google Scholar] [CrossRef] [Green Version]
- Lillestøl, R.K.; Redder, P.; Garrett, R.A.; Brügger, K. A putative viral defence mechanism in archaeal cells. Archaea 2006, 2, 59–72. [Google Scholar] [CrossRef] [Green Version]
- Zink, I.A.; Fouqueau, T.; Risa, G.T.; Werner, F.; Baum, B.; Bläsi, U.; Schleper, C. Comparative CRISPR type III-based knockdown of essential genes in hyperthermophilic Sulfolobales and the evasion of lethal gene silencing hyperthermophilic Sulfolobales and the evasion of lethal gene silencing. RNA Biol. 2020, 1–14. [Google Scholar] [CrossRef]
- Plagens, A.; Tjaden, B.; Hagemann, A.; Randau, L.; Hensel, R. Characterization of the CRISPR/Cas subtype I-A system of the hyperthermophilic crenarchaeon Thermoproteus tenax. J. Bacteriol. 2012, 194, 2491–2500. [Google Scholar] [CrossRef] [Green Version]
- Plagens, A.; Tripp, V.; Daume, M.; Sharma, K.; Klingl, A.; Hrle, A.; Conti, E.; Urlaub, H.; Randau, L. In vitro assembly and activity of an archaeal CRISPR-Cas type I-A Cascade interference complex. Nucleic Acids Res. 2014, 42, 5125–5138. [Google Scholar] [CrossRef]
- Bernick, D.L.; Cox, C.L.; Dennis, P.P.; Lowe, T.M. Comparative genomic and transcriptional analyses of CRISPR systems across the genus Pyrobaculum. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, S.; Shah, S.A.; Garrett, R. SMV1 virus-induced CRISPR spacer acquisition from the conjugative plasmid pMGB1 in Sulfolobus solfataricus P2. Biochem. Soc. Trans. 2013, 41, 1449–1458. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Li, Y.; Wang, X.; Ye, Q.; Li, H.; Liang, Y.; She, Q.; Peng, N. Transcriptional regulator-mediated activation of adaptation genes triggers CRISPR de novo spacer acquisition. Nucleic Acids Res. 2015, 43, 1044–1055. [Google Scholar] [CrossRef]
- León-Sobrino, C.; Kot, W.P.; Garrett, R.A. Transcriptome changes in STSV2-infected Sulfolobus islandicusREY15A undergoing continuous CRISPR spacer acquisition. Mol. Microbiol. 2016, 99, 719–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; She, Q.; Garrett, R.A. Diverse CRISPR-Cas responses and dramatic cellular DNA changes and cell death in pKEF9-conjugated Sulfolobus species. Nucleic Acids Res. 2016, 44, 4233–4242. [Google Scholar] [CrossRef] [Green Version]
- Bikard, D.; Jiang, W.; Samai, P.; Hochschild, A.; Zhang, F.; Marraffini, L. a Programmable repression and activation of bacterial gene expression using an engineered CRISPR-Cas system. Nucleic Acids Res. 2013, 41, 7429–7437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komor, A.C.; Badran, A.H.; Liu, D.R. CRISPR-Based Technologies for the Manipulation of Eukaryotic Genomes. Cell 2017, 168, 20–36. [Google Scholar] [CrossRef] [Green Version]
- Stachler, A.-E.; Schwarz, T.S.; Schreiber, S.; Marchfelder, A. CRISPRi as an efficient tool for gene repression in archaea. Methods 2019. [Google Scholar] [CrossRef]
- Zebec, Z.; Zink, I.A.; Kerou, M.; Schleper, C. Efficient CRISPR-Mediated Post-transcriptional Gene Silencing in a Hyperthermophilic Archaeon Using Multiplexed crRNA Expression. G3 (Bethesda) 2016, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Feng, X.; She, Q. Reverse gyrase functions in genome integrity maintenance by protecting DNA breaks in vivo. Int. J. Mol. Sci. 2017, 18, 1340. [Google Scholar] [CrossRef] [Green Version]
- Zink, I.A.; Pfeifer, K.; Wimmer, E.; Sleytr, U.B.; Schuster, B.; Schleper, C. CRISPR-mediated gene silencing reveals involvement of the archaeal S-layer in cell division and virus infection. Nat. Commun. 2019, 10, 4797. [Google Scholar] [CrossRef] [Green Version]
- Bassani, F.; Zink, I.A.; Pribasnig, T.; Wolfinger, M.T.; Romagnoli, A.; Resch, A.; Schleper, C.; Bläsi, U.; La Teana, A. Indications for a moonlighting function of translation factor aIF5A in the crenarchaeum Sulfolobus solfataricus. RNA Biol. 2019, 16. [Google Scholar] [CrossRef] [Green Version]
- Stetter, K.O. Hyperthermophiles in the history of life. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 1837–1842. [Google Scholar] [CrossRef]
- Krupovic, M.; Cvirkaite-Krupovic, V.; Iranzo, J.; Prangishvili, D.; Koonin, E.V. Viruses of archaea: Structural, functional, environmental and evolutionary genomics. Virus Res. 2018, 244, 181–193. [Google Scholar] [CrossRef]
- Quemin, E.R.J.; Quax, T.E.F. Archaeal viruses at the cell envelope: Entry and egress. Front. Microbiol. 2015, 6, 552. [Google Scholar] [CrossRef] [Green Version]
- Dewerff, S.J.; Bautista, M.A.; Pauly, M.; Zhang, C.; Whitaker, R.J. Killer archaea: Virus-mediated antagonism to CRISPR-immune populations results in emergent virus-host mutualism. MBio 2020, 11. [Google Scholar] [CrossRef]
- Pauly, M.D.; Bautista, M.A.; Black, J.A.; Whitaker, R.J. Diversified local CRISPR-Cas immunity to viruses of Sulfolobus islandicus. Philos. Trans. R. Soc. B Biol. Sci. 2019, 374, 20180093. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Zhao, S.; Ren, C.; Zhu, Y.; Zhou, H.; Lai, Y.; Zhou, F.; Jia, Y.; Zheng, K.; Huang, Z. CRISPRminer is a knowledge base for exploring CRISPR-Cas systems in microbe and phage interactions. Commun. Biol. 2018, 1. [Google Scholar] [CrossRef] [Green Version]
- Athukoralage, J.S.; Graham, S.; Rouillon, C.; Grüschow, S.; Czekster, C.M.; White, M.F. The dynamic interplay of host and viral enzymes in type iii crispr-mediated cyclic nucleotide signalling. Elife 2020, 9, 1–16. [Google Scholar] [CrossRef]
- Samolygo, A.; Athukoralage, J.S.; Graham, S.; White, M.F. Fuse to defuse: A self-limiting ribonuclease-ring nuclease fusion for type III CRISPR defence. Nucleic Acids Res. 2020, 48, 6149–6156. [Google Scholar] [CrossRef]
- He, F.; Vestergaard, G.; Peng, W.; She, Q.; Peng, X. CRISPR-Cas type I-A Cascade complex couples viral infection surveillance to host transcriptional regulation in the dependence of Csa3b. Nucleic Acids Res. 2017, 45, 1902–1913. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Wootton, J.C.; Gertz, E.M.; Agarwala, R.; Morgulis, A.; Schäffer, A.A.; Yu, Y.K. Protein database searches using compositionally adjusted substitution matrices. FEBS J. 2005, 272, 5101–5109. [Google Scholar] [CrossRef]
- Medvedeva, S.; Liu, Y.; Koonin, E.V.; Severinov, K.; Prangishvili, D.; Krupovic, M. Virus-borne mini-CRISPR arrays are involved in interviral conflicts. Nat. Commun. 2019, 10, 5204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Garrett, R.A.; Saunders, S.J.; Backofen, R. CRISPRstrand: Predicting repeat orientations to determine the crRNA-encoding strand at CRISPR loci. Bioinformatics 2014, 30, i489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, S.J.; Alkhnbashi, O.S.; Rose, D.; Will, S.; Backofen, R. CRISPRmap: An automated classification of repeat conservation in prokaryotic adaptive immune systems. Nucleic Acids Res. 2013, 41, 8034–8044. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silas, S.; Lucas-Elio, P.; Jackson, S.A.; Aroca-Crevillén, A.; Hansen, L.L.; Fineran, P.C.; Fire, A.Z.; Sánchez-Amat, A. Type III CRISPR-Cas systems can provide redundancy to counteract viral escape from type I systems. Elife 2017, 6. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zink, I.A.; Wimmer, E.; Schleper, C. Heavily Armed Ancestors: CRISPR Immunity and Applications in Archaea with a Comparative Analysis of CRISPR Types in Sulfolobales. Biomolecules 2020, 10, 1523. https://doi.org/10.3390/biom10111523
Zink IA, Wimmer E, Schleper C. Heavily Armed Ancestors: CRISPR Immunity and Applications in Archaea with a Comparative Analysis of CRISPR Types in Sulfolobales. Biomolecules. 2020; 10(11):1523. https://doi.org/10.3390/biom10111523
Chicago/Turabian StyleZink, Isabelle Anna, Erika Wimmer, and Christa Schleper. 2020. "Heavily Armed Ancestors: CRISPR Immunity and Applications in Archaea with a Comparative Analysis of CRISPR Types in Sulfolobales" Biomolecules 10, no. 11: 1523. https://doi.org/10.3390/biom10111523