Pharmacometabolomic Pathway Response of Effective Anticancer Agents on Different Diets in Rats with Induced Mammary Tumors

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
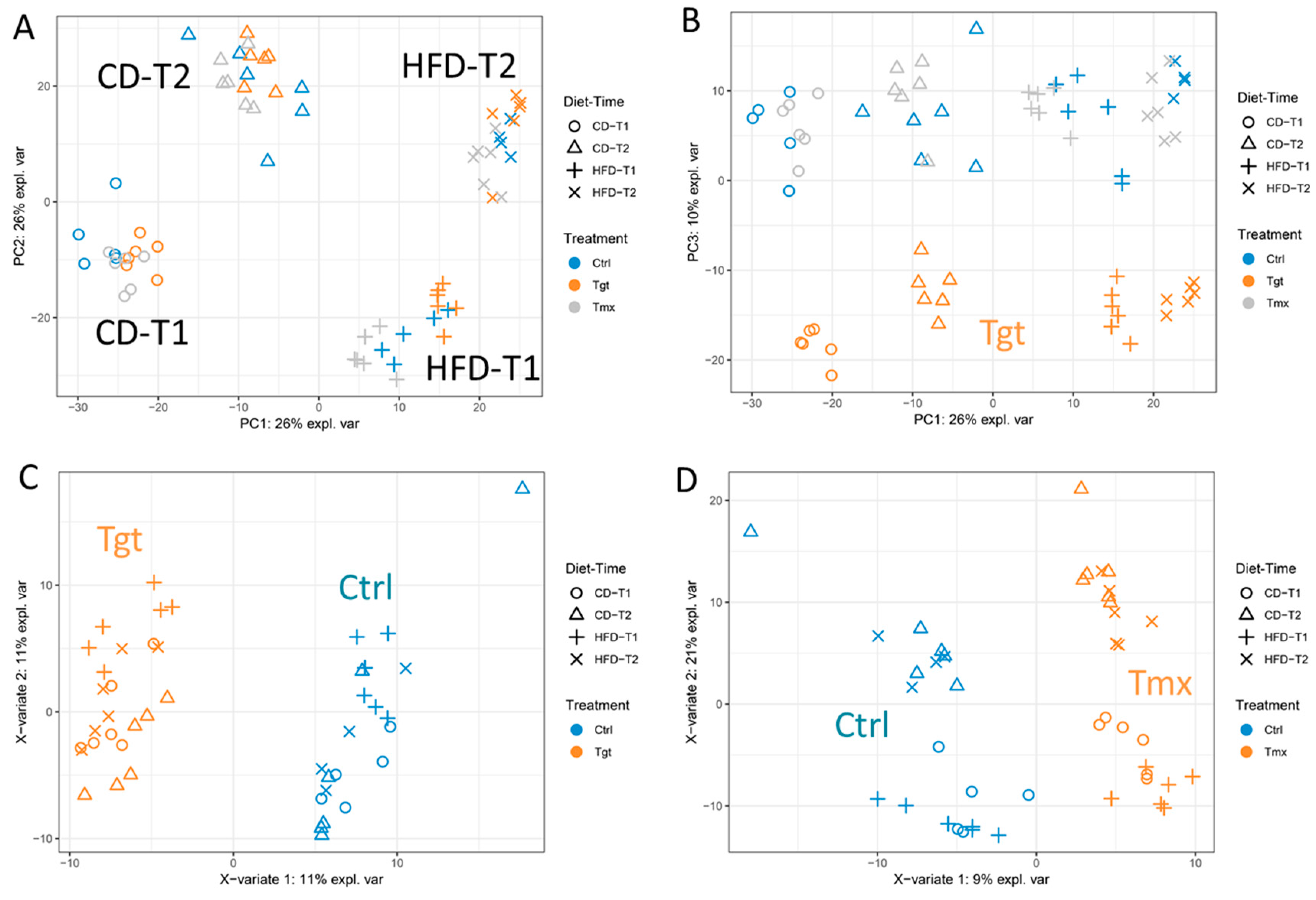
2.1. Principal Component Analysis (PCA) and Partial Least Square Discriminant Analysis (PLS-DA)
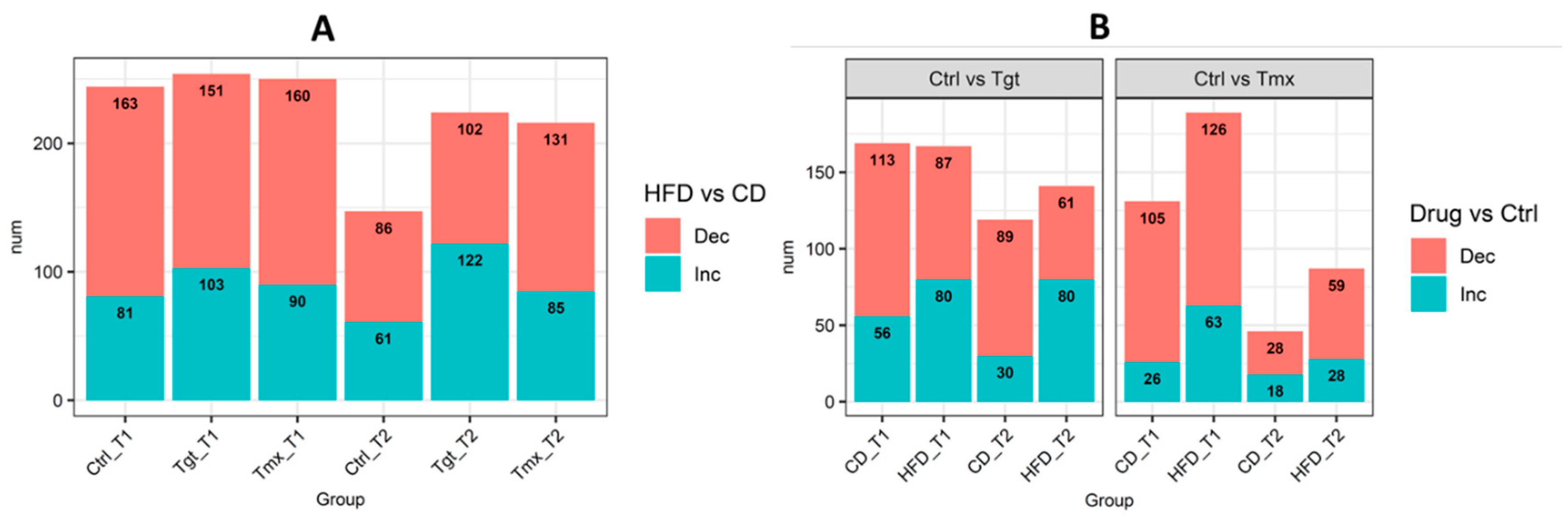
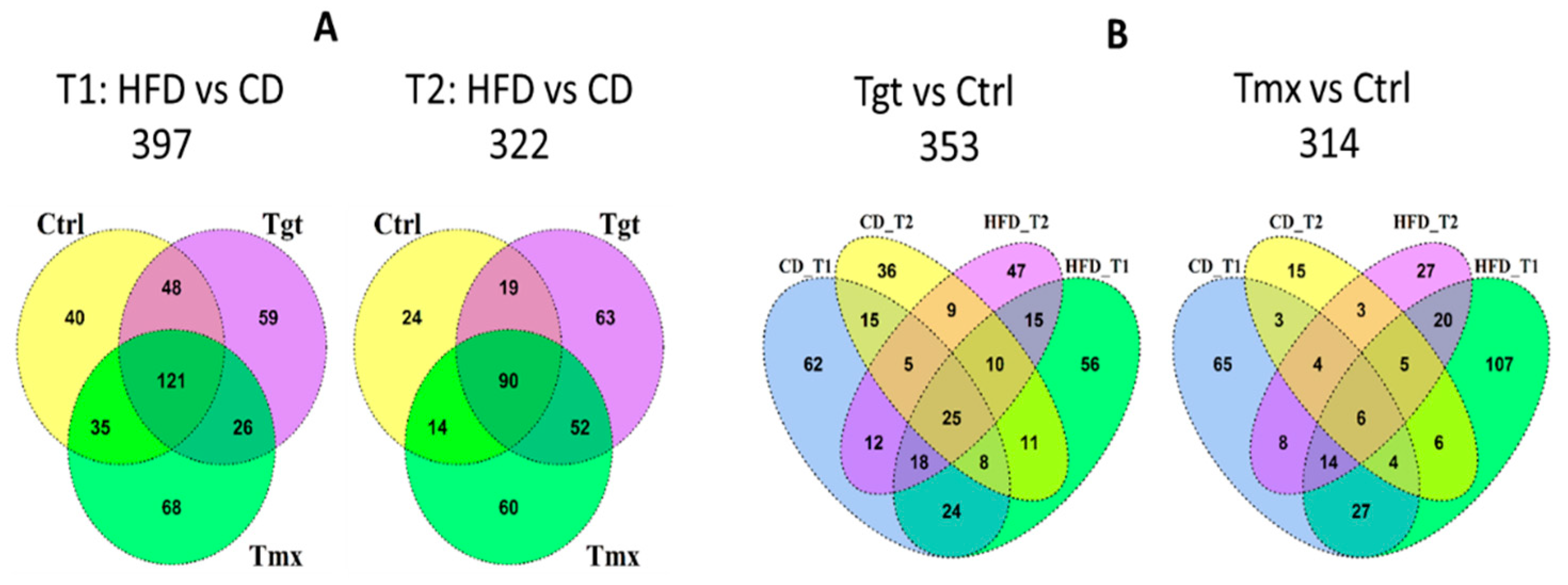
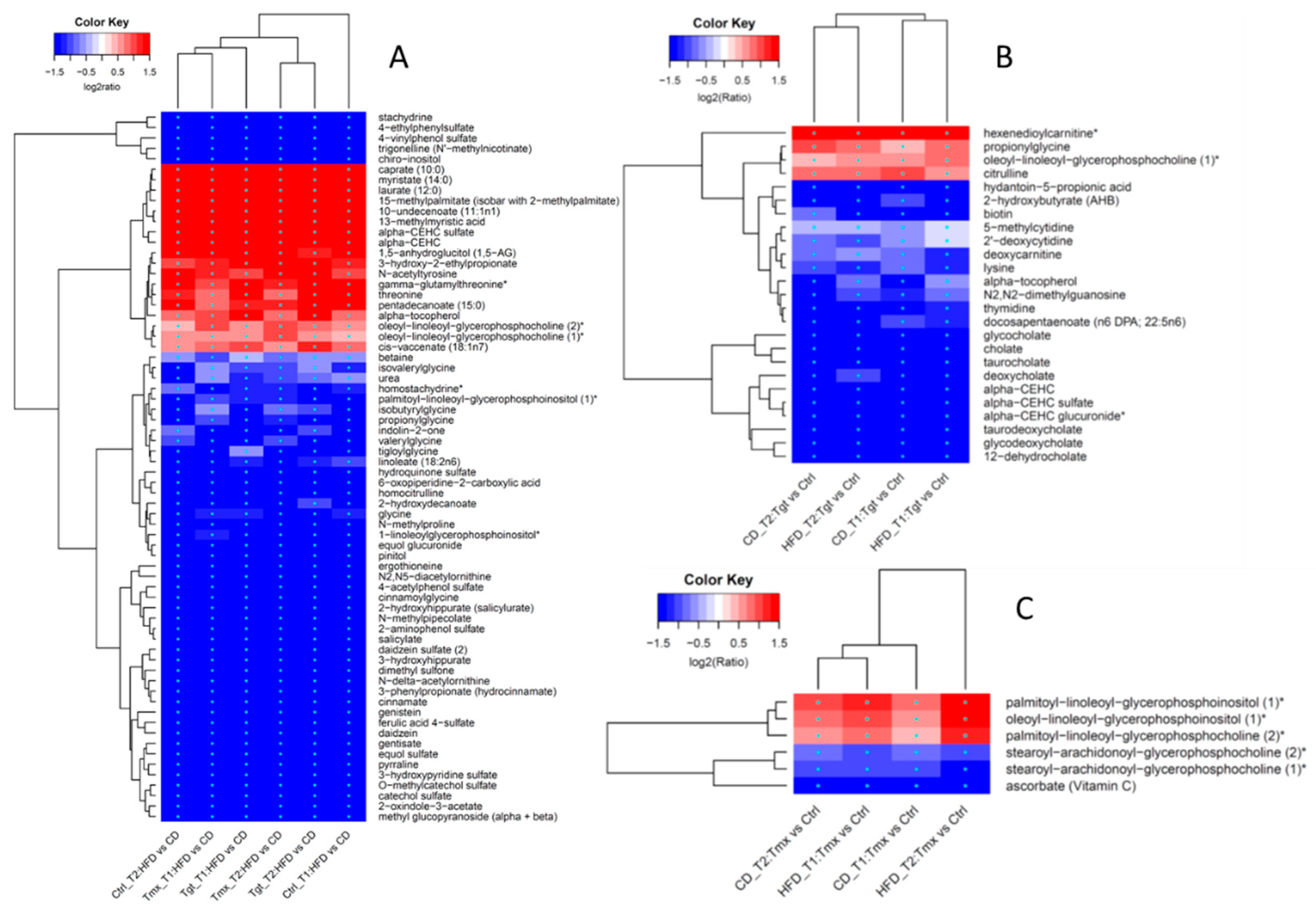
2.2. Differentially Regulated Metabolites
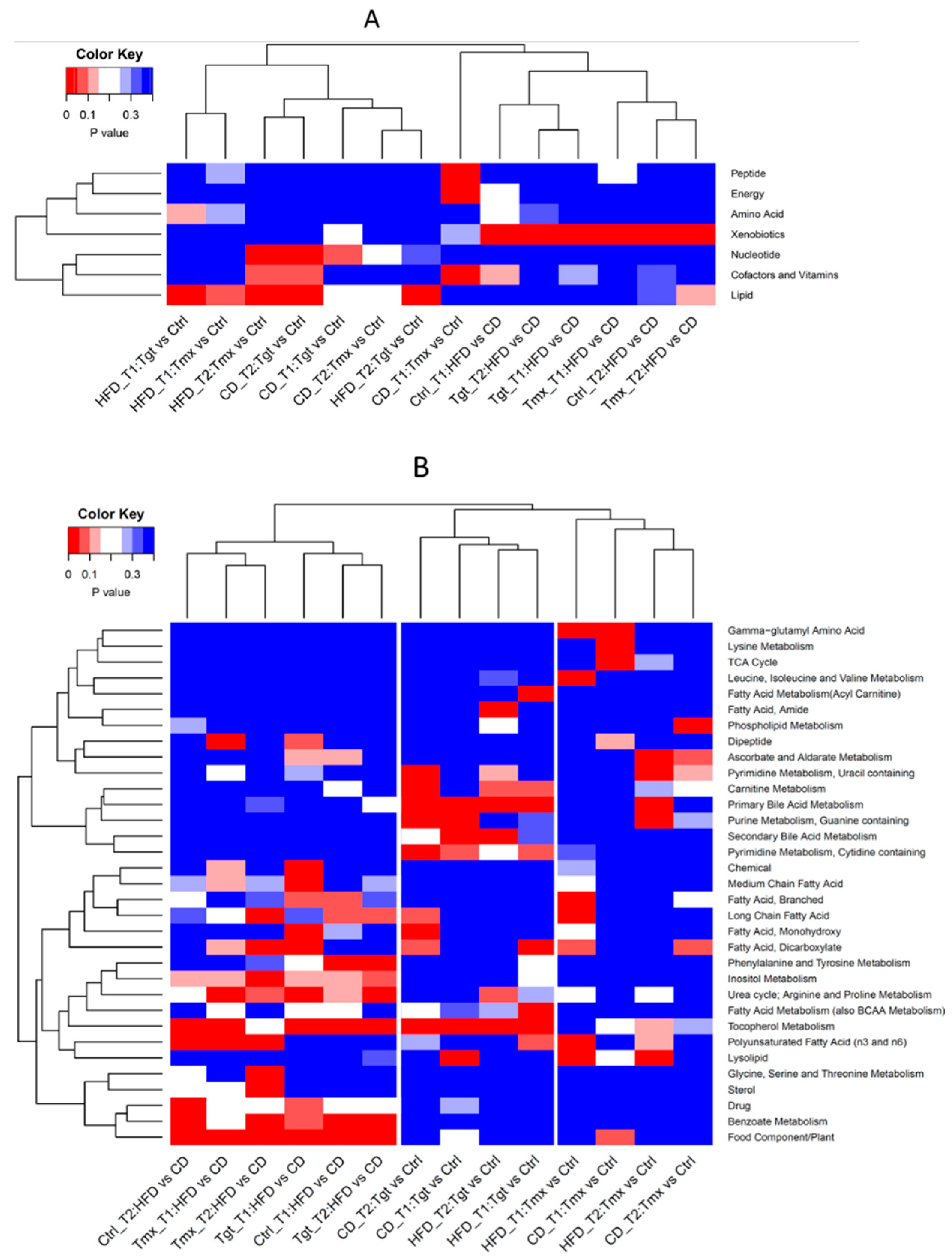
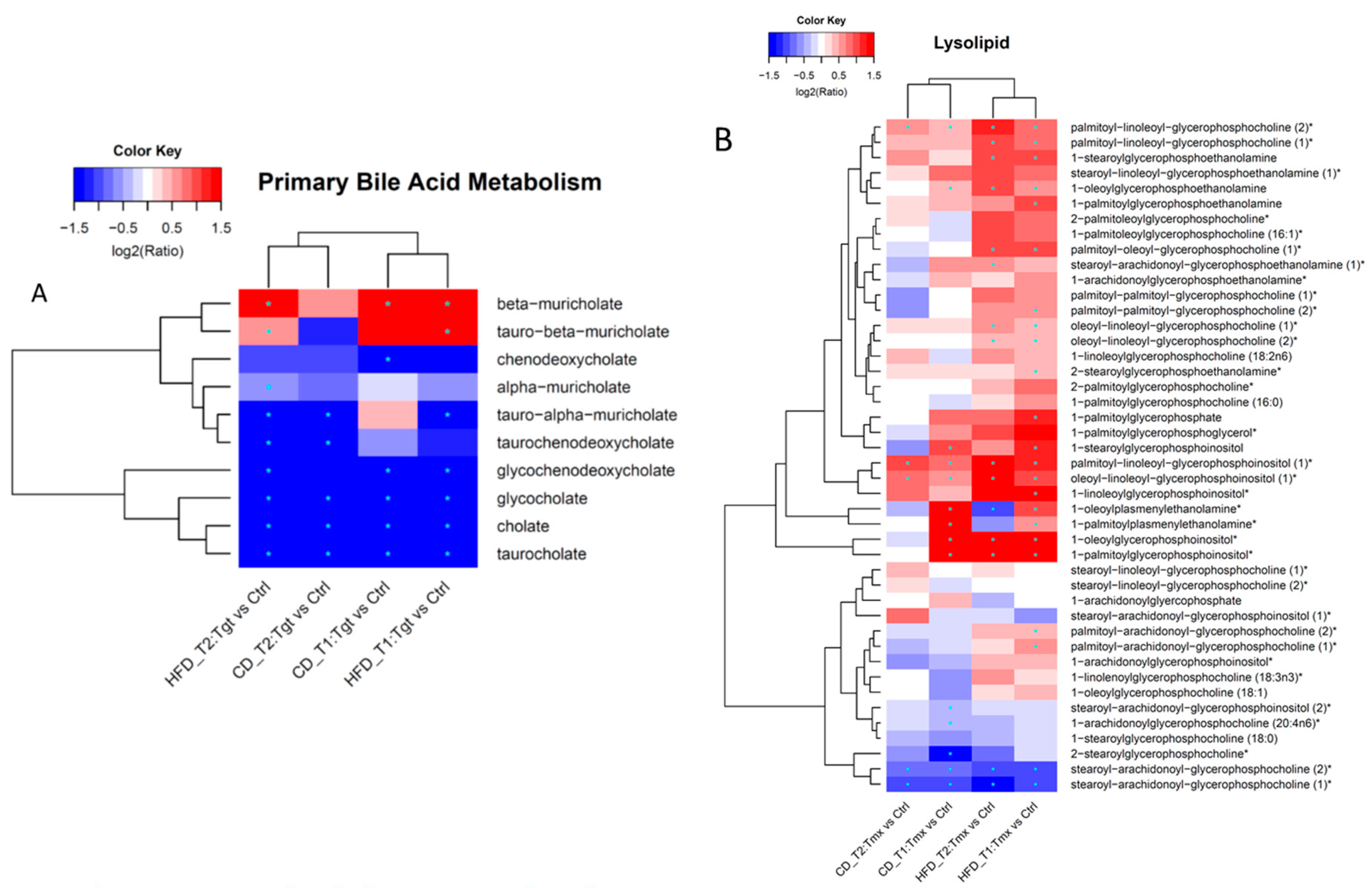
2.3. Pathway Enrichment Analysis
3. Discussion
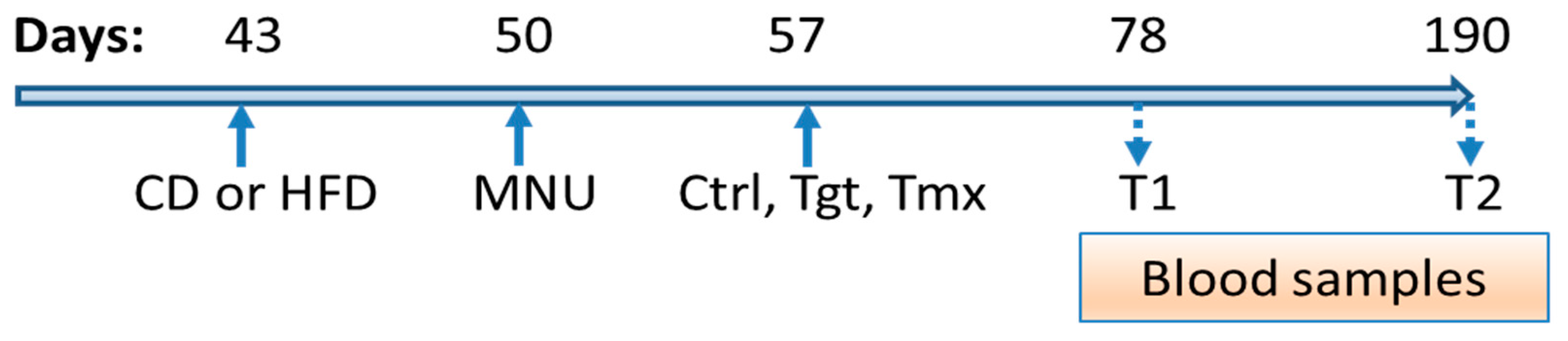
4. Materials and Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments and Disclaimer
Conflicts of Interest
References
- Gottardis, M.M.; Jordan, V.C. Antitumor actions of keoxifene and tamoxifen in the N-nitrosomethylurea-induced rat mammary carcinoma model. Cancer Res. 1987, 47, 4020–4024. [Google Scholar] [PubMed]
- Moon, R.C.; Steele, V.E.; Kelloff, G.J.; Thomas, C.F.; Detrisac, C.J.; Mehta, R.G.; Lubet, R.A. Chemoprevention of MNU-induced mammary tumorigenesis by hormone response modifiers: Toremifene, RU 16117, tamoxifen, aminoglutethimide and progesterone. Anticancer Res. 1994, 14, 889–893. [Google Scholar] [PubMed]
- Lubet, R.A.; Steele, V.E.; DeCoster, R.; Bowden, C.; You, M.; Juliana, M.M.; Eto, I.; Kelloff, G.J.; Grubbs, C.J. Chemopreventive effects of the aromatase inhibitor vorozole (R 83842) in the methylnitrosourea-induced mammary cancer model. Carcinogenesis 1998, 19, 1345–1351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lubet, R.A.; Szabo, E.; Christov, K.; Bode, A.M.; Ericson, M.E.; Steele, V.E.; Juliana, M.M.; Grubbs, C.J. Effects of gefitinib (Iressa) on mammary cancers: Preventive studies with varied dosages, combinations with vorozole or targretin, and biomarker changes. Mol. Cancer Ther. 2008, 7, 972–979. [Google Scholar] [CrossRef] [PubMed]
- Lubet, R.A.; Christov, K.; Nunez, N.P.; Hursting, S.D.; Steele, V.E.; Juliana, M.M.; Eto, I.; Grubbs, C.J. Efficacy of Targretin on methylnitrosourea-induced mammary cancers: Prevention and therapy dose-response curves and effects on proliferation and apoptosis. Carcinogenesis 2005, 26, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.M.; Donnelly, L.A.; Emslie-Smith, A.M.; Alessi, D.R.; Morris, A.D. Metformin and reduced risk of cancer in diabetic patients. BMJ 2005, 330, 1304–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, M.D.; Grubbs, C.J.; Bode, A.M.; Reid, J.M.; McGovern, R.; Bernard, P.S.; Stijleman, I.J.; Green, J.E.; Bennett, C.; Juliana, M.M.; et al. Lack of effect of metformin on mammary carcinogenesis in nondiabetic rat and mouse models. Cancer Prev. Res. 2015, 8, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, E.A.; James, K.A.; Frei, J.V.; Carroll, K.K. Effects of dietary fat on long-term growth and mammary tumorigenesis in female Sprague-Dawley rats given a low dose of DMBA. Nutr. Cancer 1988, 11, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Clinton, S.K.; Imrey, P.B.; Alster, J.M.; Simon, J.; Truex, C.R.; Visek, W.J. The combined effects of dietary protein and fat on 7,12-dimethylbenz(a)anthracene-induced breast cancer in rats. J. Nutr. 1984, 114, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Lubet, R.A.; Beger, R.D.; Miller, M.S.; Luster, J.; Seifried, H.E.; Grubbs, C.J. Comparison of Effects of Diet on Mammary Cancer: Efficacy of Various Preventive Agents and Metabolomic Changes of Different Diets and Agents. Cancer Prev. Res. 2018, 11, 831–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vedell, P.T.; Townsend, R.R.; You, M.; Malone, J.P.; Grubbs, C.J.; Bland, K.I.; Muccio, D.D.; Atigadda, V.R.; Chen, Y.; Vignola, K.; et al. Global molecular changes in rat livers treated with RXR agonists: A comparison using transcriptomics and proteomics. Pharmacol. Res. Perspect. 2014, 2, e00074. [Google Scholar] [CrossRef]
- Saito, K.; Goda, K.; Kobayashi, A.; Yamada, N.; Maekawa, K.; Saito, Y.; Sugai, S. Arachidonic acid-containing phosphatidylcholine characterized by consolidated plasma and liver lipidomics as an early onset marker for tamoxifen-induced hepatic phospholipidosis. J. Appl. Toxicol. 2017, 37, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Lubet, R.A.; Boring, D.; Steele, V.E.; Ruppert, J.M.; Juliana, M.M.; Grubbs, C.J. Lack of efficacy of the statins atorvastatin and lovastatin in rodent mammary carcinogenesis. Cancer Prev. Res. 2009, 2, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Lubet, R.A.; Townsend, R.; Clapper, M.L.; Juliana, M.M.; Steele, V.E.; McCormick, D.L.; Grubbs, C.J. 5MeCDDO Blocks Metabolic Activation but Not Progression of Breast, Intestine, and Tongue Cancers. Is Antioxidant Response Element a Prevention Target? Cancer Prev. Res. 2016, 9, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Ohta, T.; Masutomi, N.; Tsutsui, N.; Sakairi, T.; Mitchell, M.; Milburn, M.V.; Ryals, J.A.; Beebe, K.D.; Guo, L. Untargeted metabolomic profiling as an evaluative tool of fenofibrate-induced toxicology in Fischer 344 male rats. Toxicol. Pathol. 2009, 37, 521–535. [Google Scholar] [CrossRef] [PubMed]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, nontargeted ultrahigh performance liquid chromatography/electrospray ionization tandem mass spectrometry platform for the identification and relative quantification of the small-molecule complement of biological systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef] [PubMed]
- Dehaven, C.D.; Evans, A.M.; Dai, H.; Lawton, K.A. Organization of GC/MS and LC/MS metabolomics data into chemical libraries. J. Cheminform. 2010, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, Y.; Benjamini, Y. More powerful procedures for multiple significance testing. Stat. Med. 1990, 9, 811–818. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Z.; Miller, M.S.; Lubet, R.A.; Grubbs, C.J.; Beger, R.D. Pharmacometabolomic Pathway Response of Effective Anticancer Agents on Different Diets in Rats with Induced Mammary Tumors. Metabolites 2019, 9, 149. https://doi.org/10.3390/metabo9070149
Cao Z, Miller MS, Lubet RA, Grubbs CJ, Beger RD. Pharmacometabolomic Pathway Response of Effective Anticancer Agents on Different Diets in Rats with Induced Mammary Tumors. Metabolites. 2019; 9(7):149. https://doi.org/10.3390/metabo9070149
Chicago/Turabian StyleCao, Zhijun, Mark Steven Miller, Ronald A. Lubet, Clinton J. Grubbs, and Richard D. Beger. 2019. "Pharmacometabolomic Pathway Response of Effective Anticancer Agents on Different Diets in Rats with Induced Mammary Tumors" Metabolites 9, no. 7: 149. https://doi.org/10.3390/metabo9070149