High-Intensity Interval Training Decreases Resting Urinary Hypoxanthine Concentration in Young Active Men—A Metabolomic Approach

 , ,
, ,
Abstract
:1. Introduction
2. Results
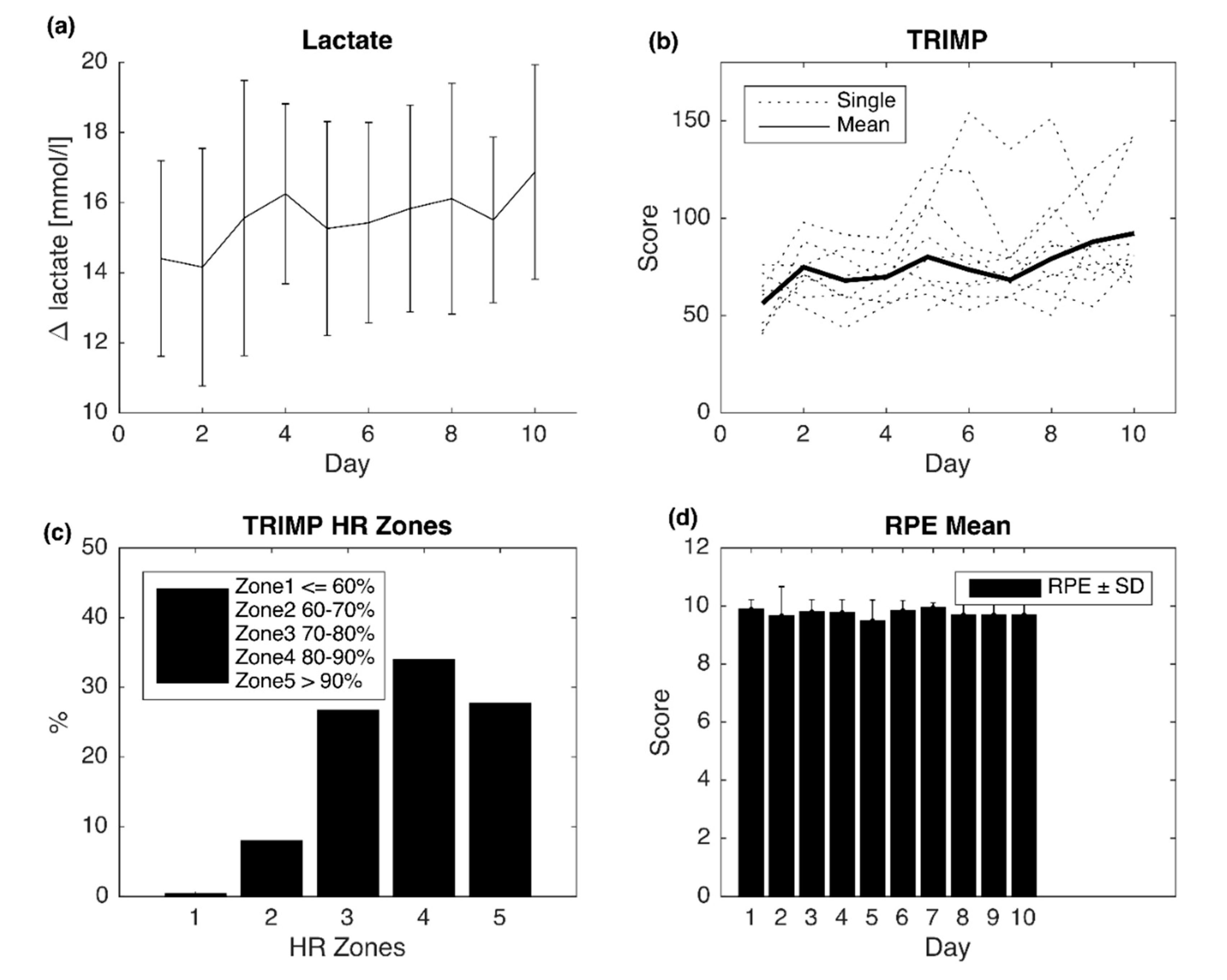
2.1. Training Parameters
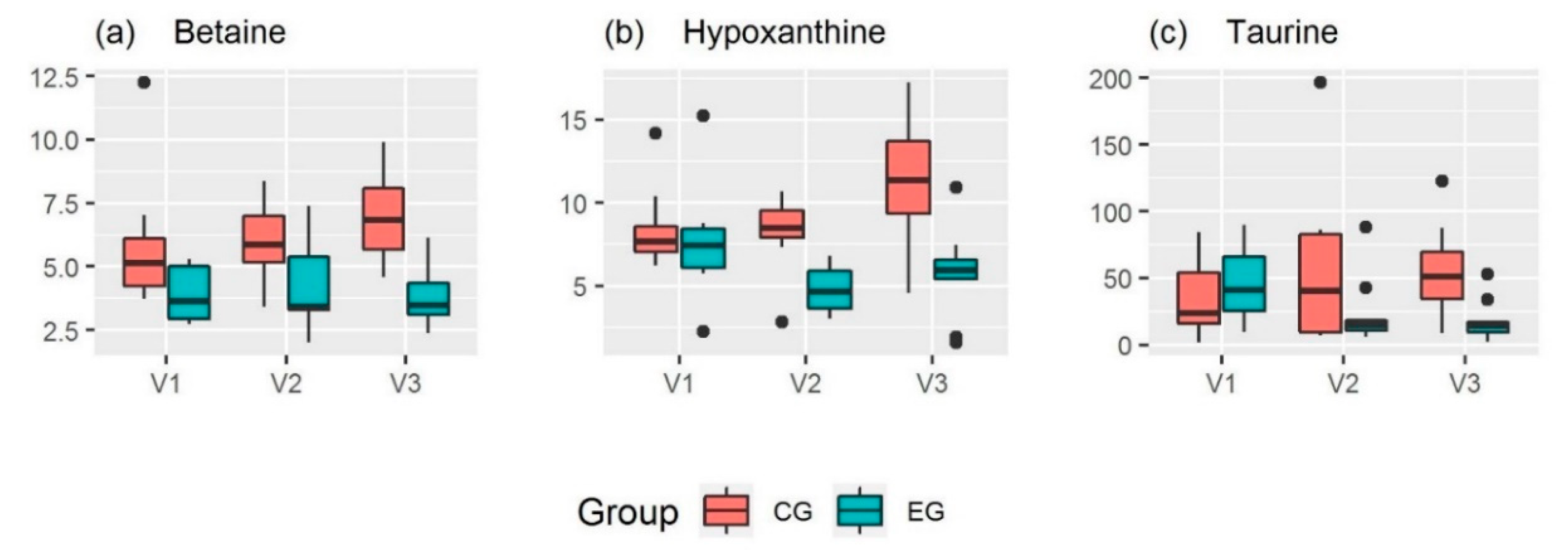
2.2. Urinary Metabolites
3. Discussion
4. Materials and Methods
4.1. Subjects and Study Design
4.2. Preliminary Testing
4.3. Experimental Protocol
4.4. Urine Sample Collection
4.5. Metabolomic Analyses
4.5.1. NMR
4.5.2. LC-MS
4.6. Data Handling and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADMA | asymmetric dimethylarginine |
| ATP | adenosine triphosphate |
| BMI | body mass index |
| CG | control group |
| EG | experimental group |
| GC-MS | gas chromatography-mass spectrometry |
| HGPRT | hypoxanthine-guanine phosphoribosyltransferase |
| HIIT | high-intensity interval training |
| HRmax | maximum heart rate |
| LC-MS | liquid chromatography-mass spectrometry |
| LLOQ | lower limit of quantification |
| MDBF | Mehrdimensionaler Befindlichkeitsfragebogen (German) |
| MSI-CE-MS | multi-segment injection-capillary electrophoresis-mass spectrometry |
| NMR | nuclear magnetic resonance |
| p | p-value of context-dependent test |
| PGC-1α | peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| Pmax | maximal power |
| QC | quality control |
| RPE | rating of perceived exertion |
| rpm | revolutions per minute |
| SDMA | symmetric dimethylarginine |
| TMAO | trimethylamine-N-oxide |
| Tmax | time to exhaustion at Pmax |
| TRIMP | training impulse |
| UPLC-MS/MS | ultra-performance liquid chromatography-tandem mass spectrometry |
| V1 | visit 1 (day before the training started) |
| V2 | visit 2 (day after the the last training session) |
| V3 | visit 3 (after four days of recovery); |
| VO2max | maximal oxygen consumption |
Appendix A


{kind=link}
{kind=link}
| EG (n = 10) | CG (n = 8) | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V1 | V2 | V3 | V1 | V2 | V3 | |||||||||||||
| β-Aminoisobutyrate 2, c) | 13 | ± | 16 | 10 | ± | 11 | 10 | ± | 9 | 3.2 | ± | 1.1 | 2.9 | ± | 1.7 | 4.6 | ± | 3.7 |
| γ-Aminobutyrate 2, c) | 0.10 | ± | 0.04 | 0.11 | ± | 0.05 | 0.10 | ± | 0.07 | 0.10 | ± | 0.07 | 0.13 | ± | 0.05 | 0.09 | ± | 0.03 |
| γ-Butyrobetaine 2, c) | 0.25 | ± | 0.22 | 0.14 | ± | 0.10 | 0.22 | ± | 0.14 | 0.15 | ± | 0.12 | 0.18 | ± | 0.11 | 0.22 | ± | 0.12 |
| π-Methylhistidine 2, c) | 42 | ± | 40 | 45 | ± | 40 | 39 | ± | 32 | 19 | ± | 27 | 19 | ± | 26 | 10 | ± | 8 |
| τ-Methylhistidine 2, c) | 18 | ± | 3 | 19 | ± | 2 | 18 | ± | 2 | 21 | ± | 7 | 18 | ± | 4 | 18 | ± | 3 |
| 1-Methylnicotinamide 1 | 3.1 | ± | 1.1 | 3.8 | ± | 1.6 | 3.9 | ± | 1.3 | 5.9 | ± | 3.9 | 4.3 | ± | 2.6 | 5.7 | ± | 2.7 |
| 2-Hydroxyisobutyrate 1 | 4.2 | ± | 0.7 | 4.2 | ± | 0.9 | 4.2 | ± | 0.8 | 4.0 | ± | 1.0 | 3.8 | ± | 0.9 | 4.3 | ± | 1.2 |
| 3-Hydroxyisovalerate 1 | 3.4 | ± | 1.0 | 3.4 | ± | 1.1 | 2.9 | ± | 0.6 | 2.9 | ± | 0.6 | 3.0 | ± | 0.8 | 3.5 | ± | 1.3 |
| 3-Indoxylsulfate 1 | 18 | ± | 8 | 21 | ± | 16 | 20 | ± | 11 | 19 | ± | 4 | 20 | ± | 8 | 17 | ± | 6 |
| 3-Methylxanthine 1 | 5.2 | ± | 3.5 | 6.4 | ± | 3.1 | 6.3 | ± | 5.4 | 4.3 | ± | 2.6 | 3.6 | ± | 1.6 | 5.5 | ± | 4.3 |
| 4-Hydroxyphenylacetate 1 | 8.7 | ± | 8.5 | 8.5 | ± | 4.0 | 7.9 | ± | 5.5 | 6.2 | ± | 1.6 | 6.9 | ± | 2.6 | 6.3 | ± | 2.5 |
| Acetate 1 | 2.8 | ± | 1.4 | 3.2 | ± | 2.4 | 2.9 | ± | 1.9 | 2.9 | ± | 1.7 | 3.5 | ± | 2.0 | 3.3 | ± | 1.0 |
| Acetone 1, e) | 1.1 | ± | 0.5 | 1.0 | ± | 0.6 | 1.0 | ± | 0.9 | 1.2 | ± | 1.0 | 1.8 | ± | 2.4 | 1.5 | ± | 0.8 |
| ADMA 2, c) | 3.7 | ± | 0.4 †b | 3.6 | ± | 0.5 | 3.4 | ± | 0.4 †b | 3.8 | ± | 0.7 | 3.8 | ± | 0.6 | 3.7 | ± | 0.6 |
| Alanine 1 | 14 | ± | 6 | 15 | ± | 6 | 14 | ± | 6 | 14 | ± | 6 | 16 | ± | 5 | 13 | ± | 5 |
| Anserine 2, c) | 0.18 | ± | 0.18 | 0.23 | ± | 0.23 | 0.13 | ± | 0.10 | 0.29 | ± | 0.59 | 0.12 | ± | 0.12 | 0.06 | ± | 0.02 |
| Arginine 2, c) | 2.1 | ± | 0.9 | 1.9 | ± | 0.5 | 1.7 | ± | 0.6 | 1.8 | ± | 0.5 | 2.7 | ± | 2.7 | 2.1 | ± | 0.6 |
| Betaine 2, a) c) | 3.9 | ± | 1.1 | 4.3 | ± | 1.8 | 3.9 | ± | 1.3 * | 6.0 | ± | 3.0 | 6.0 | ± | 1.7 | 7.0 | ± | 1.9 * |
| Betonicine 2, c) | 0.64 | ± | 1.38 | 0.28 | ± | 0.34 | 0.19 | ± | 0.21 | 0.09 | ± | 0.10 | 0.10 | ± | 0.15 | 0.04 | ± | 0.03 |
| Carnitine 2, c) | 12 | ± | 10 | 10 | ± | 9 | 15 | ± | 11 | 10 | ± | 10 | 10 | ± | 6 | 15 | ± | 5 |
| Carnosine 2, c) | 1.0 | ± | 0.6 | 1.3 | ± | 0.7 | 1.2 | ± | 0.5 | 1.3 | ± | 0.7 | 1.4 | ± | 0.6 | 1.3 | ± | 0.7 |
| Choline 2, c) | 2.0 | ± | 1.1 | 1.9 | ± | 0.9 | 1.6 | ± | 0.5 | 1.7 | ± | 0.7 | 2.3 | ± | 1.7 | 1.8 | ± | 0.3 |
| cis-Aconitate 1 | 10 | ± | 2 | 11 | ± | 2 | 11 | ± | 3 | 11 | ± | 4 | 12 | ± | 4 | 13 | ± | 8 |
| Citrate 1 | 200 | ± | 130 | 200 | ± | 130 | 190 | ± | 100 | 200 | ± | 100 | 200 | ± | 70 | 230 | ± | 70 |
| Citrulline 2, c) | 0.7 | ± | 0.4 | 0.6 | ± | 0.3 | 0.6 | ± | 0.3 | 0.5 | ± | 0.3 †b | 1.3 | ± | 1.8 | 1.1 | ± | 0.7 †b |
| Creatine 2, a) c) | 2.6 | ± | 0.8 | 2.4 | ± | 0.6 | 3.2 | ± | 2.7 | 4.1 | ± | 3.4 | 4.2 | ± | 3.2 | 3.1 | ± | 1.4 |
| Dimethylamine 2, c) | 23 | ± | 7.8 | 21 | ± | 2 | 20 | ± | 3 | 21 | ± | 2 | 19 | ± | 2 | 22 | ± | 3 |
| N,N-Dimethylglycine 2, a) c) | 2.3 | ± | 0.8 | 2.7 | ± | 1.3 | 2.2 | ± | 0.9 | 2.5 | ± | 1.1 | 2.7 | ± | 0.7 | 2.8 | ± | 0.8 |
| Dimethylsulfone 1 | 4.2 | ± | 3.6 | 3.9 | ± | 2.8 | 3.9 | ± | 3.1 | 3.3 | ± | 1.4 | 4.5 | ± | 4.4 | 4.3 | ± | 2.2 |
| Formate 1 | 11 | ± | 5 | 13 | ± | 9 | 13 | ± | 7 | 13 | ± | 5 | 15 | ± | 9 | 16 | ± | 5 |
| Gluconate 1 | 24 | ± | 8 | 26 | ± | 6 | 24 | ± | 5 | 24 | ± | 5 | 23 | ± | 7 | 2 | ± | 7 |
| Glycine 1 | 80 | ± | 50 | 72 | ± | 40 | 78 | ± | 41 | 60 | ± | 22 | 61 | ± | 16 | 62 | ± | 24 |
| Glycolate 1 | 29 | ± | 8 | 31 | ± | 8 | 30 | ± | 11 | 37 | ± | 13 | 34 | ± | 9 | 34 | ± | 8 |
| Guanidoacetate 1 | 13 | ± | 4 | 11 | ± | 4 | 11 | ± | 5 | 13 | ± | 5 | 14 | ± | 6 | 13 | ± | 4 |
| Hippurate 1 | 140 | ± | 80 | 170 | ± | 110 | 140 | ± | 70 | 250 | ± | 130 | 190 | ± | 80 | 170 | ± | 110 |
| Histidine 2, c) | 58 | ± | 22 | 59 | ± | 21 | 55 | ± | 25 | 60 | ± | 28 | 60 | ± | 15 | 57 | ± | 12 |
| Hypoxanthine 1 | 7.6 | ± | 3.3 †a | 4.8 | ± | 1.4 * †a | 5.8 | ± | 2.6 * | 8.5 | ± | 2.7 †b | 8.2 | ± | 2.4 * †c | 11.4 | ± | 3.9 * †b †c |
| Isoleucine 1 | 0.72 | ± | 0.32 | 0.81 | ± | 0.22 | 0.61 | ± | 0.19 * | 0.70 | ± | 0.20 | 0.78 | ± | 0.20 | 0.95 | ± | 0.51 * |
| Lactate 1 | 3.6 | ± | 0.8 | 3.8 | ± | 1.2 | 3.8 | ± | 1.4 | 3.6 | ± | 1.1 | 4.3 | ± | 1.9 | 3.7 | ± | 0.8 |
| Leucine 1 | 2.0 | ± | 0.5 | 2.0 | ± | 0.4 | 1.9 | ± | 0.5 | 1.9 | ± | 0.2 | 2.0 | ± | 0.6 | 2.0 | ± | 0.3 |
| Mannitol 1, c) | 8.1 | ± | 4.8 | 10.4 | ± | 4.4 | 10.5 | ± | 5.7 | 5.3 | ± | 5.1 | 6.3 | ± | 2.5 | 10.2 | ± | 8.8 |
| Methanol 1 | 2.9 | ± | 2.5 | 4.4 | ± | 4.2 | 3.7 | ± | 4.5 | 2.6 | ± | 2.4 | 4.5 | ± | 6.1 | 3.6 | ± | 2.5 |
| Methylamine 1, b) | 2.9 | ± | 1.6 | 2.4 | ± | 1.2 | 2.3 | ± | 1.0 | 2.3 | ± | 0.8 | 2.4 | ± | 0.7 | 2.2 | ± | 0.7 |
| N-Methylarginine 2, c) | 0.03 | ± | 0.03 | 0.02 | ± | 0.01 | 0.02 | ± | 0.02 | 0.01 | ± | 0.01 †b | 0.04 | ± | 0.06 | 0.04 | ± | 0.03 †b |
| N-Methylproline 2, a) c) | 0.08 | ± | 0.07 | 0.20 | ± | 0.31 | 0.09 | ± | 0.04 | 0.25 | ± | 0.34 | 0.15 | ± | 0.15 | 0.19 | ± | 0.16 |
| Methylsuccinate 1 | 5.4 | ± | 1.4 | 6.1 | ± | 2.3 | 5.7 | ± | 1.4 | 6.1 | ± | 2.4 †b | 6.2 | ± | 1.1 | 7.4 | ± | 3.4 †b |
| Proline 2, a) c) | 0.61 | ± | 0.28 | 0.56 | ± | 0.15 | 0.53 | ± | 0.07 | 0.67 | ± | 0.46 | 1.10 | ± | 1.17 | 0.91 | ± | 0.43 |
| Pseudouridine 1 | 11 | ± | 1 | 11 | ± | 1 | 11 | ± | 1 | 11 | ± | 1 | 11 | ± | 1 | 11 | ± | 1 |
| Pyruvate 1 | 2.2 | ± | 0.8 | 2.2 | ± | 0.7 | 2.0 | ± | 0.5 | 2.0 | ± | 0.6 | 2.6 | ± | 1.3 | 1.8 | ± | 0.5 |
| Sarcosine 2, a) c) | 0.05 | ± | 0.02 | 0.05 | ± | 0.02 | 0.06 | ± | 0.05 | 0.09 | ± | 0.06 | 0.11 | ± | 0.11 | 0.07 | ± | 0.06 |
| SDMA 2, c) | 33 | ± | 4 | 35 | ± | 4 | 33 | ± | 3 | 34 | ± | 2 | 31 | ± | 1 | 33 | ± | 6 |
| Stachydrine 2, c) | 15 | ± | 19 | 28 | ± | 30 | 17 | ± | 12 | 17 | ± | 16 | 11 | ± | 11 | 12 | ± | 11 |
| Succinate 1 | 1.4 | ± | 0.7 | 2.1 | ± | 1.4 | 2.0 | ± | 1.1 | 2.5 | ± | 1.3 | 2.3 | ± | 1.8 | 2.3 | ± | 0.8 |
| Tartrate 1, b) d) | 1.8 | ± | 1.4 | 2.9 | ± | 3.0 | 2.1 | ± | 1.8 | 1.6 | ± | 1.1 | 1.9 | ± | 1.4 | 1.3 | ± | 0.5 |
| Taurine 1, c) | 44 | ± | 28 †b | 23 | ± | 25 | 18 | ± | 15 †b | 36 | ± | 32 | 61 | ± | 68 | 56 | ± | 38 |
| Threonine 1 | 6.8 | ± | 2.2 | 6.8 | ± | 1.8 | 7.2 | ± | 2.9 | 7.6 | ± | 2.4 | 7.6 | ± | 2.7 | 7.2 | ± | 3.4 |
| trans-Aconitate 1 | 3.2 | ± | 0.7 | 3.9 | ± | 1.4 | 3.5 | ± | 0.7 | 3.1 | ± | 0.6 | 2.9 | ± | 1.7 | 3.0 | ± | 0.8 |
| Trigonelline 2, c) | 15 | ± | 15 | 12 | ± | 10 | 14 | ± | 11 | 12 | ± | 8 | 11 | ± | 7 | 11 | ± | 10 |
| Trimethylamine 2, c) | 0.28 | ± | 0.17 | 0.25 | ± | 0.10 | 0.20 | ± | 0.07 | 0.25 | ± | 0.09 | 0.31 | ± | 0.19 | 0.20 | ± | 0.06 |
| TMAO 2, c) | 56 | ± | 55 | 45 | ± | 28 | 37 | ± | 23 | 36 | ± | 17 | 30 | ± | 12 | 58 | ± | 42 |
| Tyrosine 1 | 7.4 | ± | 2.6 | 7.5 | ± | 2.4 | 7.1 | ± | 2.5 | 8.0 | ± | 2.7 | 8.6 | ± | 3.7 | 8.1 | ± | 2.6 |
| Uracil 1 | 7.2 | ± | 2.8 | 6.9 | ± | 3.7 | 6.8 | ± | 3.1 | 6.0 | ± | 1.4 | 5.9 | ± | 2.5 | 6.0 | ± | 1.1 |
| Urea 1 | 2400 | ± | 1100 | 2700 | ± | 1100 | 2300 | ± | 800 | 2100 | ± | 500 †b | 2600 | ± | 1400 | 2900 | ± | 700 †b |
| Valine 1 | 2.3 | ± | 0.7 | 2.5 | ± | 0.6 | 2.4 | ± | 0.6 | 2.6 | ± | 0.4 | 2.6 | ± | 0.4 | 2.6 | ± | 0.5 |
References
- Laursen, P.B.; Jenkins, D.G. The Scientific Basis for High-Intensity Interval Training. Sports Med. 2002, 32, 53–73. [Google Scholar] [CrossRef] [PubMed]
- Gibala, M.J.; Little, J.P.; Macdonald, M.J.; Hawley, J.A. Physiological adaptations to low-volume, high-intensity interval training in health and disease. J. Physiol. 2012, 590, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, S.; Thoma, C.; Houghton, D.; Trenell, M.I. High-intensity interval training: A review of its impact on glucose control and cardiometabolic health. Diabetologia 2017, 60, 7–23. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; Carson, B.P.; Garcia-Roves, P.M.; Chibalin, A.V.; Sarsfield, F.M.; Barron, N.; McCaffrey, N.; Moyna, N.M.; Zierath, J.R.; O’Gorman, D.J. Exercise intensity-dependent regulation of peroxisome proliferator-activated receptor coactivator-1 mRNA abundance is associated with differential activation of upstream signalling kinases in human skeletal muscle. J. Physiol. 2010, 588, 1779–1790. [Google Scholar] [CrossRef] [PubMed]
- Little, J.P.; Safdar, A.; Bishop, D.; Tarnopolsky, M.A.; Gibala, M.J. An acute bout of high-intensity interval training increases the nuclear abundance of PGC-1alpha and activates mitochondrial biogenesis in human skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1303–R1310. [Google Scholar] [CrossRef] [PubMed]
- Spiegelman, B.M. Transcriptional control of mitochondrial energy metabolism through the PGC1 coactivators. Novartis Found. Symp. 2007, 287, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Benton, C.R.; Nickerson, J.G.; Lally, J.; Han, X.X.; Holloway, G.P.; Glatz, J.F.; Luiken, J.J.; Graham, T.E.; Heikkila, J.J.; Bonen, A. Modest PGC-1alpha overexpression in muscle in vivo is sufficient to increase insulin sensitivity and palmitate oxidation in subsarcolemmal, not intermyofibrillar, mitochondria. J. Biol. Chem. 2008, 283, 4228–4240. [Google Scholar] [CrossRef] [PubMed]
- MacInnis, M.J.; Gibala, M.J. Physiological adaptations to interval training and the role of exercise intensity. J. Physiol. 2017, 595, 2915–2930. [Google Scholar] [CrossRef]
- Burgomaster, K.A.; Howarth, K.R.; Phillips, S.M.; Rakobowchuk, M.; Macdonald, M.J.; McGee, S.L.; Gibala, M.J. Similar metabolic adaptations during exercise after low volume sprint interval and traditional endurance training in humans. J. Physiol. 2008, 586, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Astorino, T.A.; Schubert, M.M. Changes in fat oxidation in response to various regimes of high intensity interval training (HIIT). Eur. J. Appl. Physiol. 2017, 118, 51–63. [Google Scholar] [CrossRef]
- Granata, C.; Oliveira, R.S.; Little, J.P.; Renner, K.; Bishop, D.J. Mitochondrial adaptations to high-volume exercise training are rapidly reversed after a reduction in training volume in human skeletal muscle. FASEB J. 2016, 30, 3413–3423. [Google Scholar] [CrossRef] [PubMed]
- Bujak, R.; Struck-Lewicka, W.; Markuszewski, M.J.; Kaliszan, R. Metabolomics for laboratory diagnostics. J. Pharm. Biomed. Anal. 2015, 113, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Rist, M.J.; Roth, A.; Frommherz, L.; Weinert, C.H.; Krüger, R.; Merz, B.; Bunzel, D.; Mack, C.; Egert, B.; Bub, A.; et al. Metabolite patterns predicting sex and age in participants of the Karlsruhe Metabolomics and Nutrition (KarMeN) study. PLoS ONE 2017, 12, e0183228. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; von Ruesten, A.; Drogan, D.; Schulze, M.B.; Prehn, C.; Adamski, J.; Pischon, T.; Boeing, H. Variation of serum metabolites related to habitual diet: A targeted metabolomic approach in EPIC-Potsdam. Eur. J. Clin. Nutr. 2013, 67, 1100–1108. [Google Scholar] [CrossRef] [PubMed]
- Chorell, E.; Svensson, M.B.; Moritz, T.; Antti, H. Physical fitness level is reflected by alterations in the human plasma metabolome. Mol. Biosyst. 2012, 8, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Kujala, U.M.; Makinen, V.P.; Heinonen, I.; Soininen, P.; Kangas, A.J.; Leskinen, T.H.; Rahkila, P.; Wurtz, P.; Kovanen, V.; Cheng, S.; et al. Long-term leisure-time physical activity and serum metabolome. Circulation 2013, 127, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.; Caimari, A.; del Bas, J.M.; Arola, L. Metabolomics: An emerging tool to evaluate the impact of nutritional and physiological challenges. TrAC Trends Anal. Chem. 2017, 96, 79–88. [Google Scholar] [CrossRef]
- Pechlivanis, A.; Kostidis, S.; Saraslanidis, P.; Petridou, A.; Tsalis, G.; Mougios, V.; Gika, H.G.; Mikros, E.; Theodoridis, G.A. 1H NMR-Based Metabonomic Investigation of the Effect of Two Different Exercise Sessions on the Metabolic Fingerprint of Human Urine. J. Proteome Res. 2010, 9, 6405–6416. [Google Scholar] [CrossRef] [PubMed]
- Pechlivanis, A.; Kostidis, S.; Saraslanidis, P.; Petridou, A.; Tsalis, G.; Veselkov, K.; Mikros, E.; Mougios, V.; Theodoridis, G.A. 1H NMR study on the short- and long-term impact of two training programs of sprint running on the metabolic fingerprint of human serum. J. Proteome Res. 2013, 12, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Pechlivanis, A.; Papaioannou, K.G.; Tsalis, G.; Saraslanidis, P.; Mougios, V.; Theodoridis, G.A. Monitoring the Response of the Human Urinary Metabolome to Brief Maximal Exercise by a Combination of RP-UPLC-MS and (1)H NMR Spectroscopy. J. Proteome Res. 2015, 14, 4610–4622. [Google Scholar] [CrossRef] [PubMed]
- Zafeiridis, A.; Chatziioannou, A.C.; Sarivasiliou, H.; Kyparos, A.; Nikolaidis, M.G.; Vrabas, I.S.; Pechlivanis, A.; Zoumpoulakis, P.; Baskakis, C.; Dipla, K.; et al. Global Metabolic Stress of Isoeffort Continuous and High Intensity Interval Aerobic Exercise: A Comparative (1)H NMR Metabonomic Study. J. Proteome Res. 2016, 15, 4452–4463. [Google Scholar] [CrossRef] [PubMed]
- Peake, J.M.; Tan, S.J.; Markworth, J.F.; Broadbent, J.A.; Skinner, T.L.; Cameron-Smith, D. Metabolic and hormonal responses to isoenergetic high-intensity interval exercise and continuous moderate-intensity exercise. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E539–E552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danaher, J.; Gerber, T.; Wellard, R.M.; Stathis, C.G.; Cooke, M.B. The use of metabolomics to monitor simultaneous changes in metabolic variables following supramaximal low volume high intensity exercise. Metabolomics 2015, 12. [Google Scholar] [CrossRef]
- Siopi, A.; Deda, O.; Manou, V.; Kellis, S.; Kosmidis, I.; Komninou, D.; Raikos, N.; Christoulas, K.; Theodoridis, G.A.; Mougios, V. Effects of Different Exercise Modes on the Urinary Metabolic Fingerprint of Men with and without Metabolic Syndrome. Metabolites 2017, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Saoi, M.; Percival, M.; Nemr, C.; Li, A.; Gibala, M.; Britz-McKibbin, P. Characterization of the Human Skeletal Muscle Metabolome for Elucidating the Mechanisms of Bicarbonate Ingestion on Strenuous Interval Exercise. Anal. Chem. 2019, 91, 4709–4718. [Google Scholar] [CrossRef] [PubMed]
- Kuehnbaum, N.L.; Gillen, J.B.; Gibala, M.J.; Britz-McKibbin, P. Personalized metabolomics for predicting glucose tolerance changes in sedentary women after high-intensity interval training. Sci. Rep. 2014, 4, 6166. [Google Scholar] [CrossRef] [PubMed]
- Kuehnbaum, N.L.; Gillen, J.B.; Kormendi, A.; Lam, K.P.; DiBattista, A.; Gibala, M.J.; Britz-McKibbin, P. Multiplexed separations for biomarker discovery in metabolomics: Elucidating adaptive responses to exercise training. Electrophoresis 2015, 36, 2226–2236. [Google Scholar] [CrossRef]
- Hawley, J.A.; Myburgh, K.H.; Noakes, T.D.; Dennis, S.C. Training techniques to improve fatigue resistance and enhance endurance performance. J. Sports Sci. 1997, 15, 325–333. [Google Scholar] [CrossRef]
- Bouatra, S.; Aziat, F.; Mandal, R.; Guo, A.C.; Wilson, M.R.; Knox, C.; Bjorndahl, T.C.; Krishnamurthy, R.; Saleem, F.; Liu, P.; et al. The Human Urine Metabolome. PLoS ONE 2013, 8, e73076. [Google Scholar] [CrossRef]
- Wu, J.; Gao, Y. Physiological conditions can be reflected in human urine proteome and metabolome. Expert Rev. Proteom. 2015, 12, 623–636. [Google Scholar] [CrossRef]
- Gerber, T.; Borg, M.L.; Hayes, A.; Stathis, C.G. High-intensity intermittent cycling increases purine loss compared with workload-matched continuous moderate intensity cycling. Eur. J. Appl. Physiol. 2014, 114, 1513–1520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastin, P.B. Energy system interaction and relative contribution during maximal exercise. Sports Med. 2001, 31, 725–741. [Google Scholar] [CrossRef] [PubMed]
- Zieliński, J.; Kusy, K. Pathways of Purine Metabolism: Effects of Exercise and Training in Competitive Athletes. Trends Sport Sci. 2015, 3, 103–112. [Google Scholar]
- Sahlin, K.; Tonkonogi, M.; Söderlund, K. Plasma hypoxanthine and ammonia in humans during prolonged exercise. Eur. J. Appl. Physiol. Occupat. Physiol. 1999, 80, 417–422. [Google Scholar] [CrossRef] [PubMed]
- Manfredi, J.; Holmes, E.W. Purine salvage pathways in myocardium. Ann. Rev. Physiol. 1985, 47, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Snow, R.J.; Stathis, C.G.; Febbraio, M.A.; Carey, M.F. Muscle adenine nucleotide metabolism during and in recovery from maximal exercise in humans. J. Appl. Physiol. 2000, 88, 1513–1519. [Google Scholar] [CrossRef] [PubMed]
- Sutton, J.R.; Toews, C.J.; Ward, G.R.; Fox, I.H. Purine metabolism during strenuous muscular exercise in man. Metabolism 1980, 29, 254–260. [Google Scholar] [CrossRef] [Green Version]
- Stathis, C.G.; Zhao, S.; Carey, M.F.; Snow, R.J. Purine loss after repeated sprint bouts in humans. J. Appl. Physiol. 1999, 87, 2037–2042. [Google Scholar] [CrossRef] [PubMed]
- Stathis, C.G.; Carey, M.F.; Snow, R.J. The influence of allopurinol on urinary purine loss after repeated sprint exercise in man. Metabolism 2005, 54, 1269–1275. [Google Scholar] [CrossRef]
- Zieliński, J.; Kusy, K. Training-induced adaptation in purine metabolism in high-level sprinters vs. triathletes. J. Appl. Physiol. 2012, 112, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Zielinski, J.; Kusy, K. Hypoxanthine: A Universal Metabolic Indicator of Training Status in Competitive Sports. Exerc. Sport Sci. Rev. 2015, 43, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Stathis, C.G.; Carey, M.F.; Hayes, A.; Garnham, A.P.; Snow, R.J. Sprint training reduces urinary purine loss following intense exercise in humans. Appl. Physiol. Nutr. Metab. 2006, 31, 702–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stathis, C.G.; Febbraio, M.A.; Carey, M.F.; Snow, R.J. Influence of sprint training on human skeletal muscle purine nucleotide metabolism. J. Appl. Physiol. 1994, 76, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Spencer, M.; Bishop, D.; Lawrence, S. Longitudinal assessment of the effects of field-hockey training on repeated sprint ability. J. Sci. Med. Sport 2004, 7, 323–334. [Google Scholar] [CrossRef]
- Hellsten-Westing, Y.; Balsom, P.D.; Norman, B.; Sjodin, B. The effect of high-intensity training on purine metabolism in man. Acta Physiol. Scand. 1993, 149, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Hellsten-Westing, Y.; Norman, B.; Balsom, P.D.; Sjodin, B. Decreased resting levels of adenine nucleotides in human skeletal muscle after high-intensity training. J. Appl. Physiol. 1993, 74, 2523–2528. [Google Scholar] [CrossRef]
- Giacomello, A.; Salerno, C. Hypoxanthine uptake by human erythrocytes. FEBS Lett. 1979, 107, 203–204. [Google Scholar] [CrossRef] [Green Version]
- Dudzinska, W.; Suska, M.; Lubkowska, A.; Jakubowska, K.; Olszewska, M.; Safranow, K.; Chlubek, D. Comparison of human erythrocyte purine nucleotide metabolism and blood purine and pyrimidine degradation product concentrations before and after acute exercise in trained and sedentary subjects. J. Physiol. Sci. 2018, 68, 293–305. [Google Scholar] [CrossRef]
- Zielinski, J.; Kusy, K.; Rychlewski, T. Effect of training load structure on purine metabolism in middle-distance runners. Med. Sci. Sports Exerc. 2011, 43, 1798–1807. [Google Scholar] [CrossRef]
- Zielinski, J.; Rychlewski, T.; Kusy, K.; Domaszewska, K.; Laurentowska, M. The effect of endurance training on changes in purine metabolism: A longitudinal study of competitive long-distance runners. Eur. J. Appl. Physiol. 2009, 106, 867–876. [Google Scholar] [CrossRef]
- Steyer, R.; Schwenkmezger, P.; Notz, P.; Eid, M. Der Mehrdimensionale Befindlichkeitsfragebogen (MDBF). Handanweisung [A Multidimensional Questionnaire for Subjective Well-being (MDBF). Manual]; Hogrefe: Gottingen, Germany, 1997. [Google Scholar]
- Lambert, I.H.; Kristensen, D.; Holm, J.B.; Mortensen, O.H. Physiological role of taurine–from organism to organelle. Acta Physiol. 2015, 213, 191–212. [Google Scholar] [CrossRef] [PubMed]
- Peacock, A.; Martin, F.H.; Carr, A. Energy drink ingredients. Contribution of caffeine and taurine to performance outcomes. Appetite 2013, 64, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, U.; Huebbe, P.; Rimbach, G. Taurine: A Regulator of Cellular Redox Homeostasis and Skeletal Muscle Function. Mol. Nutr. Food Res. 2018, 1800569. [Google Scholar] [CrossRef] [PubMed]
- Cuisinier, C.; De Welle, J.M.; Verbeeck, R.K.; Poortmans, J.R.; Ward, R.; Sturbois, X.; Francaux, M. Role of taurine in osmoregulation during endurance exercise. Eur. J. Appl. Physiol. 2002, 87, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Cuisinier, C.; Ward, R.J.; Francaux, M.; Sturbois, X.; de Witte, P. Changes in plasma and urinary taurine and amino acids in runners immediately and 24 h after a marathon. Amino Acids 2001, 20, 13–23. [Google Scholar] [CrossRef]
- Han, X.; Patters, A.; Jones, D.; Zelikovic, I.; Chesney, R. The taurine transporter: Mechanisms of regulation. Acta Physiol. 2006, 187, 61–73. [Google Scholar] [CrossRef]
- Teerlink, T. ADMA metabolism and clearance. Vascul. Med. 2005, 10, S73–S81. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, S.; Sonntag, S.R.; Lieb, W.; Maas, R. Asymmetric and Symmetric Dimethylarginine as Risk Markers for Total Mortality and Cardiovascular Outcomes: A Systematic Review and Meta-Analysis of Prospective Studies. PLoS ONE 2016, 11, e0165811. [Google Scholar] [CrossRef]
- Nijveldt, R.J.; van Leeuwen, P.A.M.; van Guldener, C.; Stehouwer, C.D.A.; Rauwerda, J.A.; Teerlink, T. Net renal extraction of asymmetrical (ADMA) and symmetrical (SDMA) dimethylarginine in fasting humans. Nephrol. Dial. Transplant. 2002, 17, 1999–2002. [Google Scholar] [CrossRef]
- Riccioni, G.; Scotti, L.; Guagnano, M.T.; Bosco, G.; Bucciarelli, V.; Di Ilio, E.; Speranza, L.; Martini, F.; Bucciarelli, T. Physical exercise reduces synthesis of ADMA, SDMA, and L-Arg. Front. Biosci. Elite Ed. 2015, 7, 417–422. [Google Scholar] [CrossRef]
- Wang, H.; Ran, J.; Jiang, T. Urea. In Urea Transporters; Yang, B., Sands, J.M., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 7–29. [Google Scholar]
- Lever, M.; Slow, S. The clinical significance of betaine, an osmolyte with a key role in methyl group metabolism. Clin. Biochem. 2010, 43, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.R.; Ratamess, N.A.; Kang, J.; Rashti, S.L.; Faigenbaum, A.D. Effect of betaine supplementation on power performance and fatigue. J. Int. Soc. Sports Nutr. 2009, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Craig, S.A. Betaine in human nutrition. Am. J. Clin. Nutr. 2004, 80, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, R.; Brennan, L.; Price, R.K.; Wallace, J.M.W.; Strain, J.J.; Gibney, M.J.; Shewry, P.R.; Ward, J.L.; Garg, L.; Welch, R.W. Using NMR-Based Metabolomics to Evaluate Postprandial Urinary Responses Following Consumption of Minimally Processed Wheat Bran or Wheat Aleurone by Men and Women. Nutrients 2016, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Laursen, P.B.; Shing, C.M.; Peake, J.M.; Coombes, J.S.; Jenkins, D.G. Interval training program optimization in highly trained endurance cyclists. Med. Sci. Sports Exerc. 2002, 34, 1801–1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banister, E.W. Physiological Testing of Elite Athletes; Human Kinetics: Champaign, IL, USA, 1991. [Google Scholar]
- Edwards, S. The Heart Rate Monitor Book; Fleet Feet Press: Sacramento, CA, USA, 1994; pp. 113–129. [Google Scholar]
- Foster, C.; Florhaug, J.A.; Franklin, J.; Gottschall, L.; Hrovatin, L.A.; Parker, S.; Doleshal, P.; Dodge, C. A new approach to monitoring exercise training. J. Strength Cond. Res. 2001, 15, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Krüger, R.; Merz, B.; Rist, M.J.; Ferrario, P.G.; Bub, A.; Kulling, S.E.; Watzl, B. Associations of current diet with plasma and urine TMAO in the KarMeN study: Direct and indirect contributions. Mol. Nutr. Food Res. 2017, 61, 1700363. [Google Scholar] [CrossRef] [PubMed]
- Emwas, A.-H.M. The Strengths and Weaknesses of NMR Spectroscopy and Mass Spectrometry with Particular Focus on Metabolomics Research. In Metabonomics: Methods and Protocols; Bjerrum, J.T., Ed.; Springer: New York, NY, USA, 2015; pp. 161–193. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org (accessed on 10 July 2019).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-131. 2017. Available online: https://CRAN.R-project.org/package=nlme (accessed on 27 June 2019).
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef]

| Total (n = 18) | EG (n = 10) | CG (n = 8) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Age (years) | 30.2 | ± | 7.6 | 30.0 | ± | 8.0 | 30.4 | ± | 8.0 |
| Height (cm) | 182 | ± | 7 | 181 | ± | 7 | 184 | ± | 7 |
| Weight (kg) | 79.7 | ± | 7.6 | 77.1 | ± | 8.3 | 82.9 | ± | 5.5 |
| BMI (kg/m2) | 24.0 | ± | 2.2 | 23.6 | ± | 1.6 | 24.6 | ± | 2.8 |
| VO2max (mL/(kg/min)) | 54.1 | ± | 8.2 | 53.0 | ± | 6.1 | 55.5 | ± | 10.6 |
| Pmax (W) | 351 | ± | 30 | 348 | ± | 31 | 355 | ± | 30 |
| HRmax (bpm) | 182 | ± | 11 | 181 | ± | 13 | 185 | ± | 8 |
| 65% HRmax (bpm) | - | − | - | 124 | ± | 5 | - | − | - |
| Tmax (s) | - | − | - | 162 | ± | 23 | - | − | - |
| 60% Tmax (s) | - | − | - | 97 | ± | 14 | - | − | - |
| Sets | Intensity | Duration | Rest |
|---|---|---|---|
| 8 | Pmax | 60% Tmax | 65% HRmax |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kistner, S.; Rist, M.J.; Krüger, R.; Döring, M.; Schlechtweg, S.; Bub, A. High-Intensity Interval Training Decreases Resting Urinary Hypoxanthine Concentration in Young Active Men—A Metabolomic Approach. Metabolites 2019, 9, 137. https://doi.org/10.3390/metabo9070137
Kistner S, Rist MJ, Krüger R, Döring M, Schlechtweg S, Bub A. High-Intensity Interval Training Decreases Resting Urinary Hypoxanthine Concentration in Young Active Men—A Metabolomic Approach. Metabolites. 2019; 9(7):137. https://doi.org/10.3390/metabo9070137
Chicago/Turabian StyleKistner, Sina, Manuela J. Rist, Ralf Krüger, Maik Döring, Sascha Schlechtweg, and Achim Bub. 2019. "High-Intensity Interval Training Decreases Resting Urinary Hypoxanthine Concentration in Young Active Men—A Metabolomic Approach" Metabolites 9, no. 7: 137. https://doi.org/10.3390/metabo9070137