
A Data-Driven Approach to Sugarcane Breeding Programs with Agronomic Characteristics and Amino Acid Constituent Profiling
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Cultivation Methods
2.2. Agronomic Characteristics of Sugarcane
2.3. Preparation of Sugarcane Juice and Bagasse
2.4. Evaluation of Sugarcane Juice Quality
2.5. Quantitative Analysis of Free Amino Acids in Sugarcane Juice
2.6. Statistical Analysis
3. Results
3.1. Data-Driven Approach to Evaluate the Agronomic Characteristics of Sugarcane
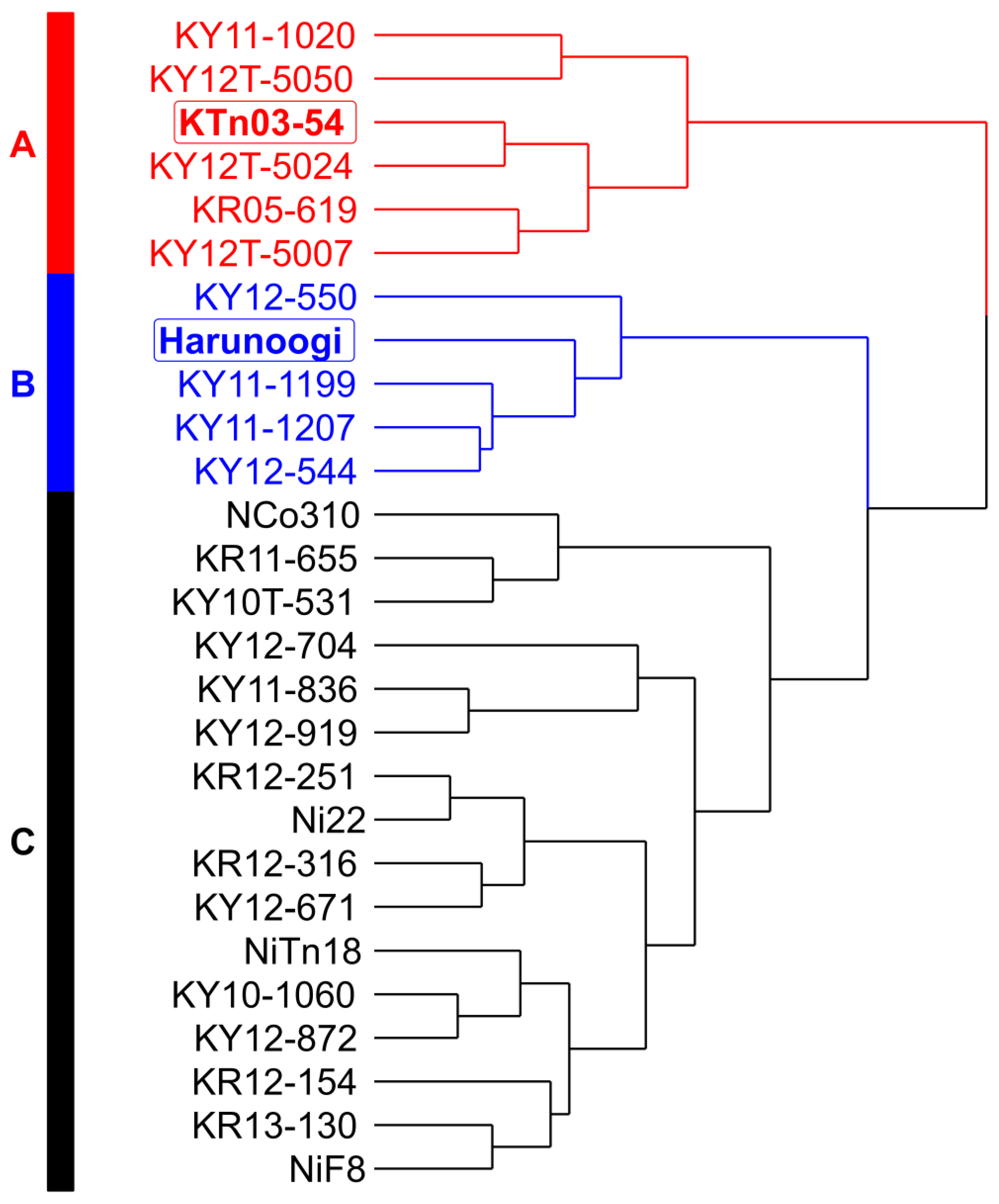
3.1.1. Visualization of Agronomic Characteristics Using a Data-Driven Approach
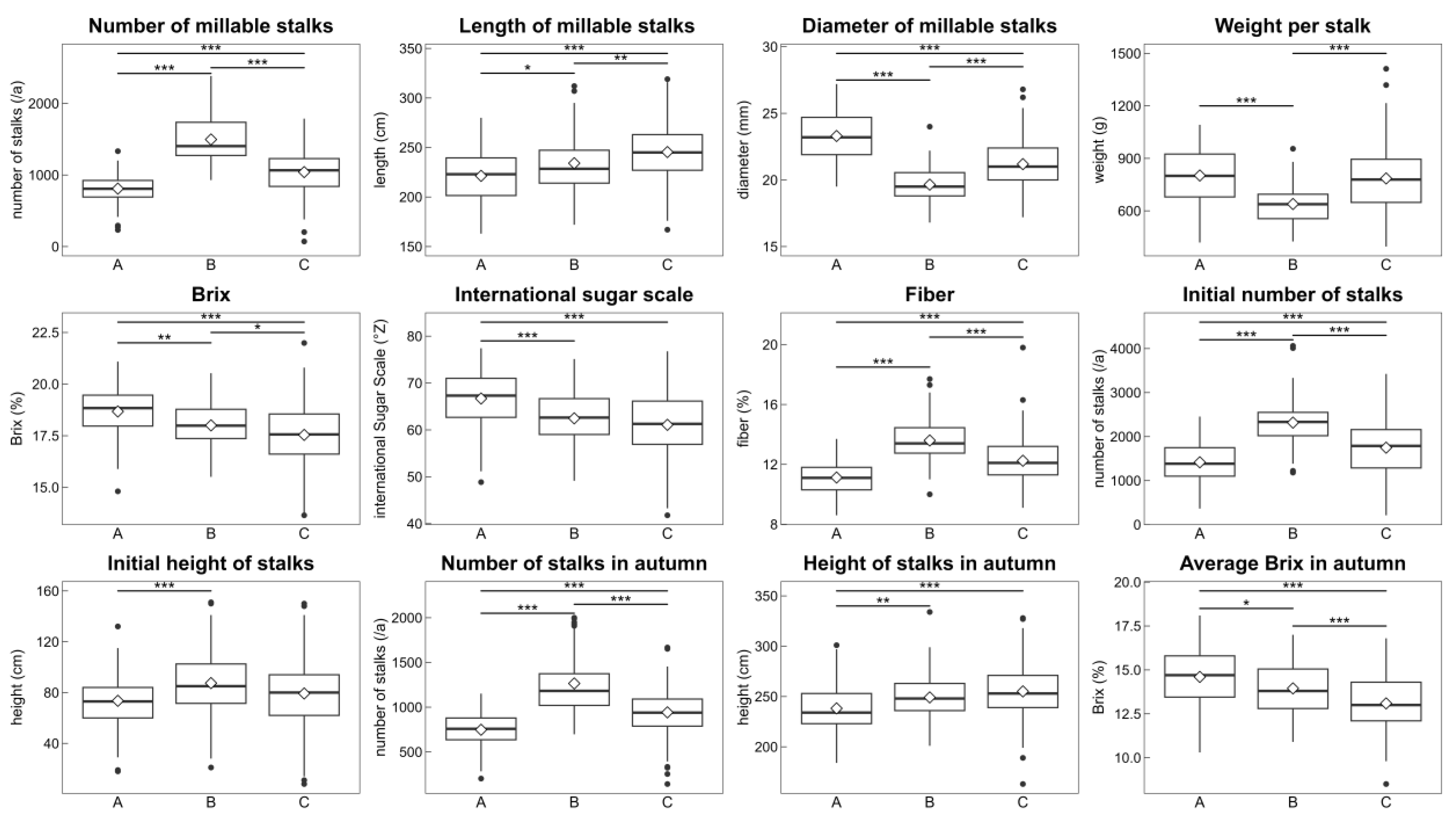
3.1.2. Analysis of the Agronomic Characteristics of the Three Groups of Sugarcane
3.2. Amino Acid Profiling of Sugarcane Juice
Features of Free Amino Acids among the Three Groups Have Different Agronomic Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Terajima, Y.; Hattori, T.; Shimatani, M.; Sato, M.; Takaragawa, H.; Sakaigaichi, T.; Umeda, M.; Naito, T.; Irei, S. Sugarcane breeding and supporting genetics research in Japan. Sugar Tech 2022, 24, 134–150. [Google Scholar] [CrossRef]
- Agricultural Industry and Horticulture Division, Department of Agricultural Administration, Kagoshima Prefectural Government. Reiwa 4 nensan Satokibi Oyobi Kanshato Seisan Jisseki (Actual Production of Sugarcane and Cane Sugar in Kagoshima (2022/2023) [In Japanese, Translated by the Author of This Article]). 2023. Available online: http://www.pref.kagoshima.jp/ag06/sangyo-rodo/nogyo/nosanbutu/satokibi/documents/41531_20231006165335-1.pdf (accessed on 1 March 2024). (In Japanese)
- Sugar Industry and Agricultural Products Division, Department of Agriculture, Forestry and Fisheries, Okinawa Prefectural Government. Reiwa 4/5 nenki I Satokibi Oyobi Kanshato Seisan Jisseki (Actual Production of Sugarcane and Cane Sugar in Okinawa (2022/2023) [In Japanese, Translated by the Author of This Article]). 2023. Available online: https://www.pref.okinawa.lg.jp/_res/projects/default_project/_page_/001/010/408/01_seisanjisseki2.pdf (accessed on 1 March 2024). (In Japanese)
- Chauhan, O.P.; Singh, D.; Tyagi, S.M.; Balyan, D.K. Studies on preservation of sugarcane juice. Int. J. Food Prop. 2002, 5, 217–229. [Google Scholar] [CrossRef]
- Arif, S.; Batool, A.; Nazir, W.; Khan, R.S.; Khalid, N. 8—Physiochemical characteristics nutritional properties and health benefits of sugarcane juice. In Non-Alcoholic Beverages; Grumezescu, A.M., Holban, A.M., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 227–257. ISBN 978-0-12-815270-6. [Google Scholar] [CrossRef]
- Zidan, D.; Azlan, A. Non-Centrifugal Sugar (NCS) and Health: A review on functional components and health benefits. Appl. Sci. 2022, 12, 460. [Google Scholar] [CrossRef]
- Hirose, N.; Maeda, G.; Takara, K.; Wada, K. Study of the interrelation between variations in sugarcane juice contents and qualities of non-centrifugal brown Sugar “Kokuto” during production. Food Preserv. Sci. 2019, 45, 141–147, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Okutsu, K.; Yamamoto, Y.; Matsuo, F.; Yoshizaki, Y.; Futagami, T.; Tamaki, H.; Maeda, G.; Tsuchida, E.; Takamine, K. Characterization of aroma profiles of kokuto-shochu prepared from three different cultivars of sugarcane. J. Biosci. Bioeng. 2023, 135, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.E.; El Gedaily, R.A.; Mocan, A.; Farag, M.A.; El-Seedi, H.R. Profiling metabolites and biological activities of sugarcane (Saccharum officinarum Linn.) juice and its product molasses via a multiplex metabolomics approach. Molecules 2019, 24, 934. [Google Scholar] [CrossRef] [PubMed]
- Ujihara, K.; Yoshimoto, M.; Wada, K.; Nagai, R.; Hirose, N.; Teruya, R. Effect of roller material used in mill and liming treatments on color tone and quality of non-centrifugal sugar. J. Jpn. Soc. Food Sci. Technol. 2009, 56, 343–349, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Asikin, Y.; Takahara, W.; Takahashi, M.; Hirose, N.; Ito, S.; Wada, K. Compositional and electronic discrimination analyses of taste and aroma profiles of non-centrifugal cane brown sugars. Food Anal. Methods 2017, 10, 1844–1856. [Google Scholar] [CrossRef]
- Phaeon, N.; Chapanya, P.; Mueangmontri, R.; Pattamasuwan, A.; Lipan, L.; Carbonell-Barrachina, Á.A.; Sriroth, K.; Nitayapat, N. Acrylamide in non-centrifugal sugars and syrups. J. Sci. Food Agric. 2021, 101, 4561–4569. [Google Scholar] [CrossRef]
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef]
- Stadler, R.H.; Blank, I.; Varga, N.; Robert, F.; Hau, J.; Guy, P.A.; Robert, M.C.; Riediker, S. Acrylamide from Maillard reaction products. Nature 2002, 419, 449–450. [Google Scholar] [CrossRef] [PubMed]
- Henao Toro, S.J.; Gómez-Narváez, F.; Contreras-Calderón, J.; Arisseto, A.P. Acrylamide in sugar products. Curr. Opin. Food Sci. 2022, 45, 100841. [Google Scholar] [CrossRef]
- Keating, B.A.; Biggs, I.M.; Webster, A.J. Monitoring Cane at the Mill to Improve Nitrogen Management on the Farm; Sugar Research and Development Corporation Project CTA029 Final Report; CSIRO Sustainable Ecosystems: Brisbane, Australia, 2002; Available online: https://elibrary.sugarresearch.com.au/bitstream/handle/11079/13993/REDACTED%20CTA029%20Final%20Rep.pdf?sequence=1&isAllowed=y (accessed on 5 March 2024).
- Parish, D.H. The amino-acids of sugar cane. I.—The amino-acids of cane-juice and the effect of nitrogenous fertilisation on the levels of these substances. J. Sci. Food Agric. 1965, 16, 240–242. [Google Scholar] [CrossRef]
- Chapman, L.S.; Ham, G.; Hurney, A.P.; Sanders, D.H.; Leonard, G.J. Preliminary evaluation of field factors affecting amino acids in sugarcane juice. In Proceedings of the 1996 Conference of the Australian Society of Sugar Cane Technologists, Mackay, Australia, 30 April–3 May 1996; pp. 213–221. [Google Scholar]
- Keating, B.A.; Webster, A.J.; Biggs, I.M. Improving CCS in the Wet Tropics via Block-Specific Monitoring of Nitrogen in Cane Delivered to the Mill; Sugar Research and Development Corporation Project CTA045 Final Report; CSIRO Sustainable Ecosystems: Brisbane, Australia, 2003; Available online: https://elibrary.sugarresearch.com.au/bitstream/handle/11079/13978/REDACTED%20CTA045%20Fin%20rep.pdf?sequence=1&isAllowed=y (accessed on 4 March 2024).
- Matsuoka, M. Sugarcane cultivation and sugar industry in Japan. Sugar Tech 2006, 8, 3–9. [Google Scholar] [CrossRef]
- Han, S.Y.; Bishop, T.F.A.; Filippi, P. Data-driven, early-season forecasts of block sugarcane yield for precision agriculture. Field Crops Res. 2022, 276, 108360. [Google Scholar] [CrossRef]
- Misawa, T.; Date, Y.; Kikuchi, J. Human metabolic, mineral, and microbiota fluctuations across daily nutritional intake visualized by a data-driven approach. J. Proteome Res. 2015, 14, 1526–1534. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Kitagawa, H. Free amino acids in stalks, roots and leaves of sugarcanes. Tech. Bull. Fac. Agric. Kagawa Univ. 1989, 41, 69–74. Available online: https://kagawa-u.repo.nii.ac.jp/records/3334 (accessed on 5 March 2024). (In Japanese with English Abstract).
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen supply influences photosynthesis establishment along the sugarcane leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Dong, F.; Pang, Z.; Fallah, N.; Zhou, Y.; Li, Z.; Hu, C. Integrated metabolomics and transcriptome analyses unveil pathways involved in sugar content and rind color of two sugarcane varieties. Front. Plant Sci. 2022, 13, 921536. [Google Scholar] [CrossRef]
- Biggs, I.M. An Investigation of Sugarcane Nitrogen Physiology: Sources, Uptake, and Metabolism. Ph.D. Thesis, University of Queensland, Brisbane, Queensland, Australia, 2003. [Google Scholar] [CrossRef]
- Ferreira, D.A.; Martins, M.C.M.; Cheavegatti-Gianotto, A.; Carneiro, M.S.; Amadeu, R.R.; Aricetti, J.A.; Wolf, L.D.; Hoffmann, H.P.; de Abreu, L.G.F.; Caldana, C. Metabolite profiles of sugarcane culm reveal the relationship among metabolism and axillary bud outgrowth in genetically related sugarcane commercial cultivars. Front. Plant Sci. 2018, 9, 857. [Google Scholar] [CrossRef]
- Rao, M.J.; Duan, M.; Wang, J.; Han, S.; Ma, L.; Mo, X.; Li, M.; Hu, L.; Wang, L. Transcriptomic and widely targeted metabolomic approach identified diverse group of bioactive compounds, antiradical activities, and their associated genes in six sugarcane varieties. Antioxidants 2022, 11, 1319. [Google Scholar] [CrossRef] [PubMed]
- Perlo, V.; Botha, F.C.; Furtado, A.; Hodgson-Kratky, K.; Henry, R.J. Metabolic changes in the developing sugarcane culm associated with high yield and early high sugar content. Plant Direct 2020, 4, e00276. [Google Scholar] [CrossRef] [PubMed]
- Date, Y.; Ishikawa, C.; Umeda, M.; Tarumoto, Y.; Okubo, M.; Tamura, Y.; Ono, H. Sugarcane metabolome compositional stability in pretreatment processes for NMR measurements. Metabolites 2022, 12, 862. [Google Scholar] [CrossRef] [PubMed]
- Hattori, T.; Terajima, Y.; Sakaigaichi, T.; Terauchi, T.; Tarumoto, Y.; Adachi, K.; Hayano, M.; Tanaka, M.; Ishikawa, S.; Umeda, M.; et al. High ratoon yield sugarcane cultivar “Harunoogi” developed for Kumage region by using an interspecific hybrid between a commercial cultivar and Saccharum spontaneum L. J. NARO Res. Dev. 2019, 2, 21–44, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Sakaigaichi, T.; Terauchi, T.; Matsuoka, M.; Terajima, Y.; Hattori, T.; Irei, S.; Ujihara, K.; Sugimoto, A.; Ishikawa, S.; Tanaka, M.; et al. Stalk weight type sugarcane variety “KTn03-54” with early maturing in the Kumage region of Kagoshima Prefecture. Bull. NARO Agric. Res. Kyushu Okinawa Reg. 2017, 66, 1–20, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Irei, S.; Terajima, Y.; Sakaigaichi, T.; Ujihara, K.; Matsuoka, M.; Sugimoto, A.; Takemure, M.; Fukui, K.; Shirasawa, S. New early maturing sugarcane variety “Ni22” with high yield under ratooning conditions. Bull. NARO Agric. Res. Kyushu Okinawa Reg. 2010, 54, 1–22, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Terajima, Y.; Sugimoto, A.; Matsuoka, M.; Ujihara, K.; Sakaigaichi, T.; Fukuhara, S.; Maeda, H.; Katsuta, Y.; Oka, M.; Shimoda, S.; et al. New sugarcane cultivar “NiTn18” with excellent ratooning ability in mulch-free cultivation. Bull. NARO Agric. Res. Kyushu Okinawa Reg. 2010, 54, 23–41, (In Japanese with English Abstract). [Google Scholar] [CrossRef]
- Umeda, M.; Sakaigaichi, T.; Tanaka, M.; Tarumoto, Y.; Adachi, K.; Hattori, T.; Hayano, M.; Takahashi, H.; Tamura, Y.; Kimura, T.; et al. Detection of a major QTL related to smut disease resistance inherited from a Japanese wild sugarcane using GRAS-Di technology. Breed. Sci. 2021, 71, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Kyushu National Agricultural Experiment Station; Kagoshima Agricultural Experiment Station; Okinawa Prefectural Agricultural Experiment Station; Kanmi Shigen Shinkokai (Sugar Crop Development Fund). Satoukibi ni Kansuru Chousa Kijun (Research Standards of Sugarcane); [In Japanese, translated by the author of this article]; Kyushu National Agricultural Experiment Station: Koshi, Japan, 1982; pp. 1–19, 22–25. (In Japanese)
- Plant Variety Protection Office, Intellectual Property Division, Export and International Bureau, Ministry of Agriculture, Forestry and Fisheries. Test Guideline (Saccharum L.). Available online: http://www.hinshu2.maff.go.jp/info/sinsakijun/kijun/1554.pdf (accessed on 4 March 2024). (In Japanese)
- International Union for the Protection of New Varieties of Plants. Test Guidelines Sugarcane (Saccharum L.) (Revision). 2023. Available online: https://www.upov.int/edocs/mdocs/upov/en/twa_52/tg_186_2_proj_4.pdf (accessed on 11 March 2024).
- Terajima, Y.; Sugimoto, A.; Tippayawat, A.; Irei, S.; Hayashi, H. Root distribution and fibre composition of intergeneric F1 hybrid between sugarcane and E. arundinaceus. Field Crops Res. 2023, 297, 108920. [Google Scholar] [CrossRef]
- Legendre, B.L.; Clarke, M.A. Comparison of clarification reagents for polarization analysis of sugarcane juice. J. Am. Soc. Sugar Cane Technol. 1991, 11, 75–81. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 20 June 2023).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC: Boston, MA, USA, 2021; Available online: http://www.rstudio.com/ (accessed on 20 June 2023).
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H.; Bryan, J. readxl: Read Excel Files. R package version 1.4.2. 2023. Available online: https://CRAN.R-project.org/package=readxl (accessed on 16 March 2024).
- Schneider, G.; Chicken, E.; Becvarik, R. NSM3: Functions and Datasets to Accompany Hollander, Wolfe, and Chicken—Nonparametric Statistical Methods, Third Edition. R package version 1.17. 2022. Available online: https://CRAN.R-project.org/package=NSM3 (accessed on 11 March 2024).
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research; R Package Version 2.1.9; Northwestern University: Evanston, IL, USA, 2021; Available online: https://CRAN.R-project.org/package=psych (accessed on 17 March 2024)R Package Version 2.1.9.
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 11 March 2024).
- Hall, M. ggarchery: Flexible Segment Geoms with Arrows for ‘ggplot2’. R Package Version 0.4.2. 2023. Available online: https://CRAN.R-project.org/package=ggarchery (accessed on 11 March 2024).
- Slowikowski, K. ggrepel: Automatically Position Non-Overlapping Text Labels with ‘ggplot2’. R Package Version 0.9.1. 2021. Available online: https://CRAN.R-project.org/package=ggrepel (accessed on 11 March 2024).
- Pedersen, T.L. patchwork: The Composer of Plots. R Package Version 1.1.1. 2020. Available online: https://CRAN.R-project.org/package=patchwork (accessed on 11 March 2024).
- Galili, T. dendextend: An R package for visualizing, adjusting, and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [PubMed]
- Ahlmann-Eltze, C.; Patil, I. ggsignif: R package for displaying significance brackets for ‘ggplot2’. PsyArXiv 2021. [Google Scholar] [CrossRef]
- Neuwirth, E. RColorBrewer: ColorBrewer Palettes. R Package Version 1.1-2. 2014. Available online: https://CRAN.R-project.org/package=RColorBrewer (accessed on 17 March 2024).
- Nagatomi, S.; Kodama, S. Studies on the selection methods for sugarcane breeding III. On the correlations between yield components, stalk form, juice quality and leaf form characters. Jpn. J. Trop. Agric. 1971, 15, 6–10. [Google Scholar] [CrossRef]
- Dinh, T.H.; Takaragawa, H.; Kawamitsu, Y. Nitrogen use efficiency and drought tolerant ability of various sugarcane varieties under drought stress at early growth stage. Plant Prod. Sci. 2019, 22, 250–261. [Google Scholar] [CrossRef]
- Singh, J.; Chandra, P.; Singh, S.B. Studies on some biochemical variations during tillering of sugarcane. Indian Sugar 1994, 43, 775–778. [Google Scholar]
- Wiggins, L.F.; Williams, J.H. Nutritional value of crops, amino acid content of West Indies sugar cane. J. Agric. Food Chem. 1955, 3, 341–345. [Google Scholar] [CrossRef]
- Ujihara, K.; Masuda, R.; Sugiura, M.; Nagata, S.; Sugimoto, A.; Terajima, Y.; Fukuhara, S. The seasonal change of sugarcane juice constituents with growth and relationships between sugarcane juice constituents and juice products constituents. Rep. Kyushu Br. Crop Sci. Soc. Jpn. 2004, 70, 75–77. Available online: https://dl.ndl.go.jp/pid/11088358/1/1 (accessed on 7 March 2024). (In Japanese).
- Fujiwara-Arasaki, T.; Mino, N. Comparison of the free amino acids in sugar cane during the growth periods. J. Jpn. Soc. Food Nutr. 1982, 35, 363–366. [Google Scholar] [CrossRef]
- Lea, P.J.; Sodek, L.; Parry, M.A.J.; Shewry, P.R.; Halford, N.G. Asparagine in plants. Ann. Appl. Biol. 2007, 150, 1–26. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ishikawa, C.; Date, Y.; Umeda, M.; Tarumoto, Y.; Okubo, M.; Morimitsu, Y.; Tamura, Y.; Nishiba, Y.; Ono, H. A Data-Driven Approach to Sugarcane Breeding Programs with Agronomic Characteristics and Amino Acid Constituent Profiling. Metabolites 2024, 14, 243. https://doi.org/10.3390/metabo14040243
Ishikawa C, Date Y, Umeda M, Tarumoto Y, Okubo M, Morimitsu Y, Tamura Y, Nishiba Y, Ono H. A Data-Driven Approach to Sugarcane Breeding Programs with Agronomic Characteristics and Amino Acid Constituent Profiling. Metabolites. 2024; 14(4):243. https://doi.org/10.3390/metabo14040243
Chicago/Turabian StyleIshikawa, Chiaki, Yasuhiro Date, Makoto Umeda, Yusuke Tarumoto, Megumi Okubo, Yasujiro Morimitsu, Yasuaki Tamura, Yoichi Nishiba, and Hiroshi Ono. 2024. "A Data-Driven Approach to Sugarcane Breeding Programs with Agronomic Characteristics and Amino Acid Constituent Profiling" Metabolites 14, no. 4: 243. https://doi.org/10.3390/metabo14040243