Non-Targeted Metabolic Profiling of Cerebellum in Spina Bifida Fetal Rats

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Experimental Design
Retinoic Acid-Induced Spina Bifida Rat Model
3. Procedure
3.1. Tissue Collection
3.2. Sample Preparation and NMR Data Collection
3.3. Metabolomics Data Analysis
4. Results
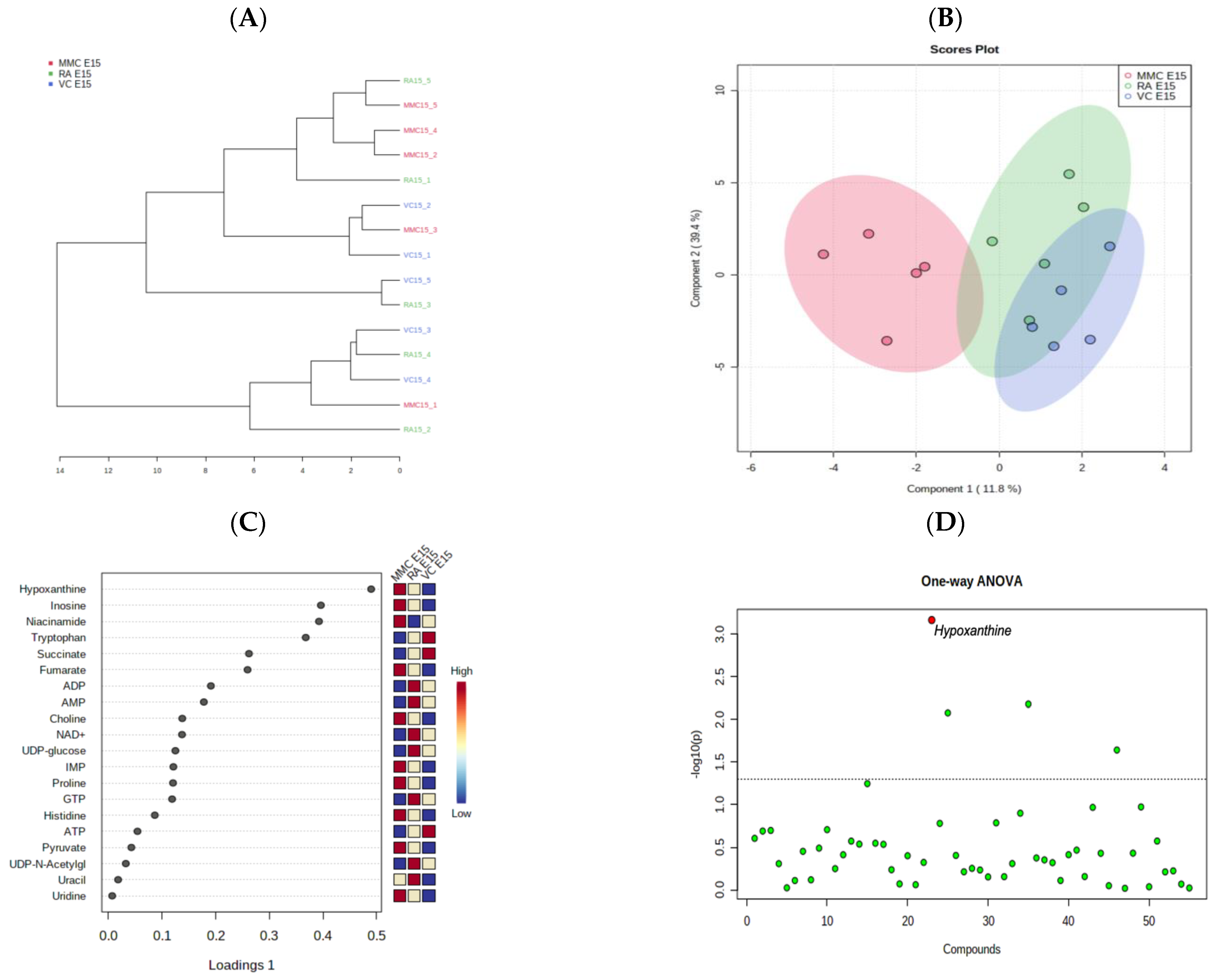
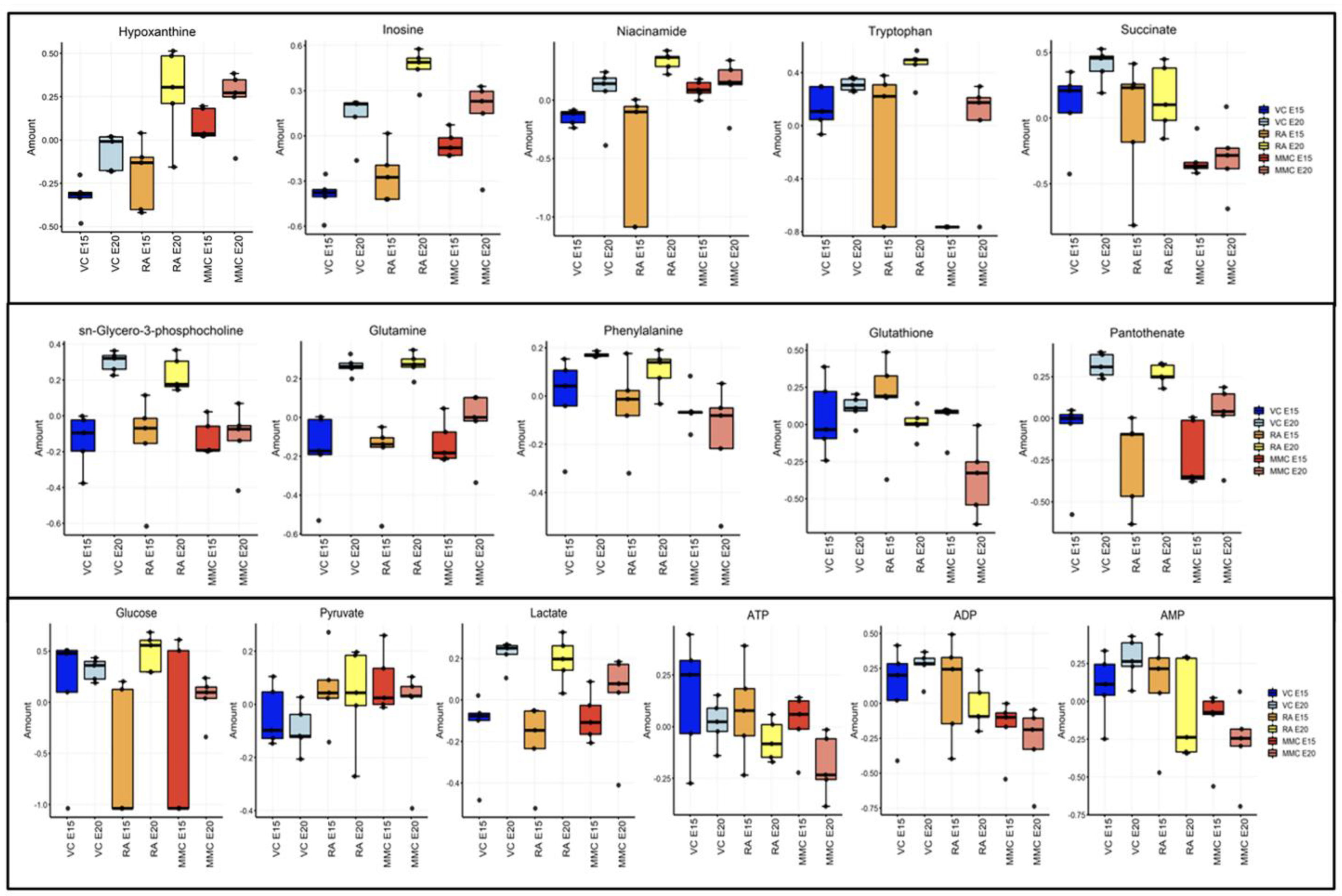
4.1. Metabolic Profile at Mid–Late Gestation (E15)
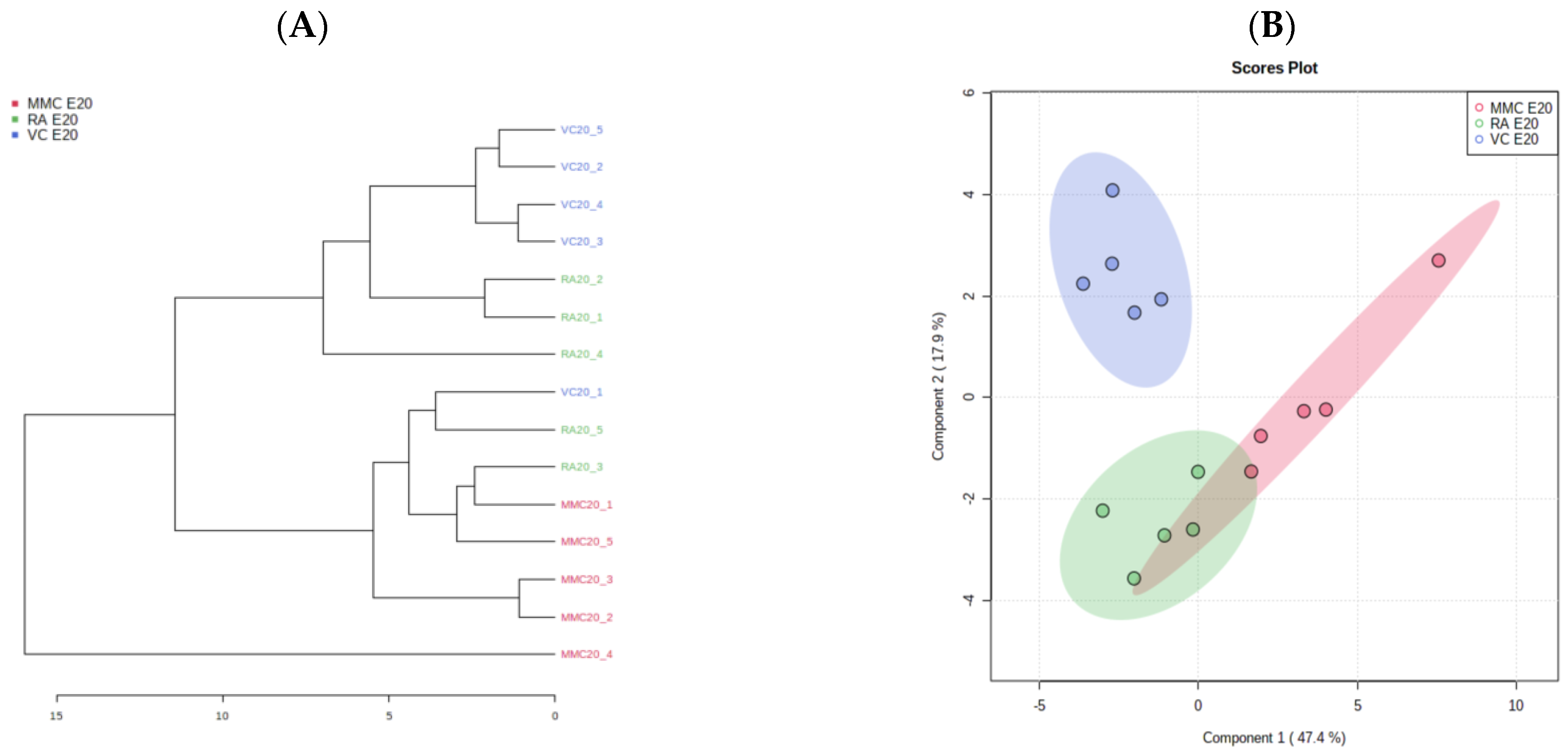
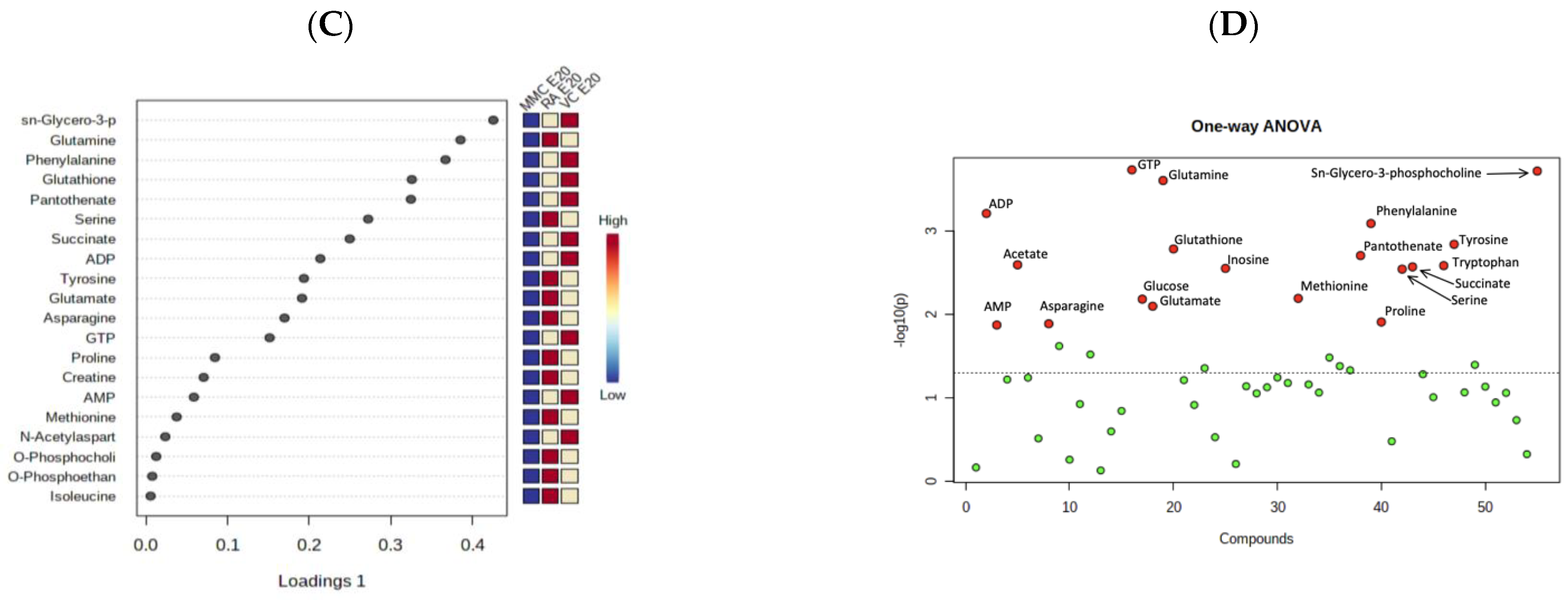
4.2. Metabolic Profile at Term Gestation (E20)
4.3. Metabolic Effect of Retinoic Acid (RA) Exposure
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Greene, N.D.; Copp, A.J. Neural tube defects. Annu. Rev. Neurosci. 2014, 37, 221–242. [Google Scholar] [CrossRef]
- Dyste, G.N.; Menezes, A.H. Symptomatic Chiari malformations: An analysis of presentation, management, and long-term outcome. J. Neurosurg. 1989, 71, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, H.J.; Hendrick, E.B. Manifestations and management of Arnold-Chiari malformation in patients with myelomeningocele. Childs Brain 1975, 1, 255–259. [Google Scholar] [CrossRef]
- Hung, C.F. The relationship between hydrocephalus and Chiari type II malformation in the experimental rat fetuses with Arnold-Chiari malformation. Proc. Natl. Sci. Counc. Repub. China B 1986, 10, 118–126. [Google Scholar]
- Danzer, E.; Schwarz, U. Retinoic acid induced myelomeningocele in fetal rats: Characterization by histopathological analysis and magnetic resonance imaging. Exp. Neurol. 2005, 192, 267–475. [Google Scholar] [CrossRef] [PubMed]
- Mignone Philpott, C.; Shannon, P. Diffusion-weighted imaging of the cerebellum in the fetus with Chiari II malformation. Am. J. Neuroradiol. 2013, 34, 1656–1660. [Google Scholar] [CrossRef]
- Oria, M.; Figueira, R.L. CD200-CD200R imbalance correlates with microglia and pro-inflammatory activation in rat spinal cords exposed to amniotic fluid in retinoid acid-induced spina bifida. Sci. Rep. 2018, 13, 10638. [Google Scholar] [CrossRef] [PubMed]
- Oria, M.; Pathak, B. Premature Neural Progenitor Cell Differentiation Into Astrocytes in Retinoic Acid-Induced Spina Bifida Rat Model. Front. Mol. Neurosci. 2022, 17, 888351. [Google Scholar] [CrossRef]
- Ceccarelli, G.; Pozzo, E. Molecular signature of amniotic fluid derived stem cells in the fetal sheep model of myelomeningocele. J. Pediatr. Surg. 2015, 50, 1521–1527. [Google Scholar] [CrossRef]
- Hansler, A.; Chen, Q. Untargeted metabolite profiling of murine embryos to reveal metabolic perturbations associated with neural tube closure defects. Birth Defects Res. A Clin. Mole Teratol. 2014, 100, 623–632. [Google Scholar] [CrossRef]
- Watanabe, M.; Meyer, K.A. Application of NMR-based metabolomics for environmental assessment in the Great Lakes using zebra mussel (Dreissena polymorpha). Metabolomics 2015, 11, 1302–1315. [Google Scholar] [CrossRef]
- Wu, H.; Southam, A.D. High-throughput tissue extraction protocol for NMR- and MS-based metabolomics. Anal. Biochem. 2008, 372, 204–212. [Google Scholar] [CrossRef]
- McGuire, J.L.; DePasquale, E.A.K. Chronic Dysregulation of Cortical and Subcortical Metabolism After Experimental Traumatic Brain Injury. Mol. Neurobiol. 2019, 56, 2908–2921. [Google Scholar] [CrossRef]
- Kouskoumvekaki, I.; Panagiotou, G. Navigating the human metabolome for biomarker identification and design of pharmaceutical molecules. J. Biomed. Biotechnol. 2011, 2011, 525497. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed]
- Lammer, E.J.; Chen, D.T. Retinoic acid embryopathy. N. Engl. J. Med. 1985, 313, 837–841. [Google Scholar] [CrossRef]
- Ross, S.A.; McCaffery, P.J. Retinoids in embryonal development. Physiol. Rev. 2000, 80, 1021–1054. [Google Scholar] [CrossRef]
- Gessner, P.; Lum, J. The mammalian purine salvage pathway as an exploitable route for cerebral bioenergetic support after brain injury. Neuropharm 2023, 224, 109370. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, S.; Bakshi, R. Protection by inosine in a cellular model of Parkinson’s disease. Neuroscience 2014, 272, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Haun, S.E.; Segeleon, J.E. Inosine mediates the protective effect of adenosine in rat astrocyte cultures subjected to combined glucose–oxygen deprivation. J. Neurochem. 1996, 67, 2051–2059. [Google Scholar] [CrossRef]
- Litsky, M.L.; Hohl, C.M. Inosine and guanosine preserve neuronal and goal cell viability in mouse spinal cord cultures during chemical hypoxia. Brain Res. 1999, 821, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Ruhal, P.; Dhingra, D. Inosine improves cognitive function and decreases aging-induced oxidative stress and neuroinflammation in aged female rats. Inflammopharacology 2018, 26, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Kamat, J.P.; Devasagayam, T.P. Nicotinamide (vitamin B3) as an effective antioxidant against oxidative damage in rat brain mitochondria. Redox Rep. 1999, 4, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Li, X. High doses of nicotinamide prevent oxidative mitochondrial dysfunction in a cellular model and improve motor deficit in a Drosophilia model of Parkinson’s disease. J. Neurosci. Res. 2008, 86, 2083–2090. [Google Scholar] [CrossRef]
- Klimova, N.; Kristian, T. Multi-targeted Effect of Nicotinamide Mononucleotide on Brain Bioenergetic Metabolism. Neurochem. Res. 2019, 44, 2280–2287. [Google Scholar] [CrossRef]
- Gasperi, V.; Sibilano, M. Niacin in the Central Nervous System: An Update of Biological Aspects and Clinical Applications. Int. J. Mol. Sci. 2019, 20, 974. [Google Scholar] [CrossRef]
- Biasibetti, H.; Pierozan, P. Hypoxanthine Intrastriatal Administration Alters Neuroinflammatory Profile and Redox Status in Striatum of Infant and Young Adult Rats. Mol. Neurobiol. 2017, 54, 2790–2800. [Google Scholar] [CrossRef]
- Biasibett-Brendler, H.; Schmitz, F. Hypoxanthine Induces Neuroenergetic Impairment and Cell Death in Striatum of Young Adult Wistar Rats. Mol. Neurobiol. 2018, 55, 4098–4106. [Google Scholar] [CrossRef]
- Kim, Y.J.; Ryu, H. Hypoxanthine causes endothelial dysfunction through oxidative stress-induced apoptosis. Biochem. Biophys. Res. Commun. 2017, 482, 821–827. [Google Scholar] [CrossRef]
- Gu, F.; Chauhan, V. Glutathione redox imbalance in brain disorders. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 89–95. [Google Scholar] [CrossRef]
- Mytilineou, C.; Kramer, B.C. Glutathione depletion and oxidative stress. Parkinsonism Relat. Disord. 2002, 8, 385–387. [Google Scholar] [CrossRef] [PubMed]
- Puppel, K.; Kapusta, A. The etiology of oxidative stress in the various species of animals, a review. J. Sci. Food Agric. 2015, 95, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.L.; Pazdro, R. Impact of Supplementary Amino Acids, Micronutrients, and Overall Diet on Glutathione Homeostasis. Nutrients 2019, 11, 1056. [Google Scholar] [CrossRef] [PubMed]
- Tretter, L.; Patocs, A. Succinate, an intermediate in metabolism, signal transduction, ROS, hypoxia, and tumorigenesis. Biochim. Biophys. Acta 2016, 1857, 1086–1101. [Google Scholar] [CrossRef]
- Aliu, E.; Kanungo, S. Amino acid disorders. Ann. Transl. Med. 2018, 6, 471. [Google Scholar] [CrossRef]
- Schultz, V.; Lowenstein, J.M. The purine nucleotide cycle. Studies of ammonia production and interconversions of adenine and hypoxanthine nucleotides and nucleosides by rat brain in situ. J. Biol. Chem. 1978, 253, 1938–1943. [Google Scholar] [CrossRef]
- Van den Berghe, G.; Bontemps, F. The purine nucleotide cycle and its molecular defects. Prog. Neurobiol. 1992, 39, 547–561. [Google Scholar] [CrossRef]
- Struzynska, L.; Sulkowski, G. Relationships between glutamine, glutamate, and GABA in nerve endings under Pb-toxicity conditions. J. Inorg. Biochem. 2004, 98, 951–958. [Google Scholar] [CrossRef]
- McKenna, M.C. The glutamate-glutamine cycle is not stoichiometric: Fates of glutamate in brain. J. Neurosci. Res. 2007, 85, 3347–3358. [Google Scholar] [CrossRef]
- Fernstrom, J.D.; Fernstrom, M.H. Tyrosine, phenylalanine, and catecholamine synthesis and function in the brain. J. Nutr. 2007, 137 (Suppl. 1), 1539S–1548S. [Google Scholar] [CrossRef]
- Benavides, J.; Gimenez, C. Effect of phenylalanine metabolites on the activities of enzymes of ketone-body utilization in brain of suckling rats. Biochem. J. 1976, 160, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.; Kureishy, N. Vitamin B5 (d-pantothenic acid) localizes in myelinated structures of the rat brain: Potential role for cerebral vitamin B5 stores in local myelin homeostasis. Biochem. Biophys. Res. Commun. 2020, 522, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Patassini, S. Cerebral deficiency of vitamin B5 (d-pantothenic acid; pantothenate) as a potentially-reversible cause of neurodegeneration and dementia in sporadic Alzheimer's disease. Biochem. Biophys. Res. Commun. 2020, 527, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Della-Flora Nunes, G.; Mueller, L. Acetyl-CoA production from pyruvate is not necessary for preservation of myelin. Glia 2017, 65, 1626–1639. [Google Scholar] [CrossRef]
- Saher, G.; Brügger, B. High cholesterol level is essential for myelin membrane growth. Nat. Neurosci. 2005, 8, 468–475. [Google Scholar] [CrossRef]
- Le, S.Q.; Nestrasil, I. Myelin and Lipid Composition of the Corpus Callosum in Mucopolysaccharidosis Type I Mice. Lipids 2020, 55, 627–637. [Google Scholar] [CrossRef]
- Skripuletz, T.; Manzel, A. Pivotal role of choline metabolites in remyelination. Brain 2015, 138, 398–413. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Significantly Altered Metabolites (E20)—MMC vs. VC | ||
|---|---|---|
| Metabolite | FC | p Value (FDR) |
| ADP | 0.31567 | 0.016892 |
| AMP | 0.31439 | 0.029109 |
| Glutamine | 0.53706 | 0.021256 |
| Glutathione | 0.38811 | 0.021256 |
| GTP | 0.43598 | 0.016892 |
| Pantothenate | 0.52425 | 0.029109 |
| Phenylalanine | 0.50395 | 0.038424 |
| sn-Glycero-3-phosphocholine | 0.39806 | 0.0062079 |
| Succinate | 0.22668 | 0.016892 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thielen, E.; Oria, M.; Watanabe-Chailland, M.; Lampe, K.; Romick-Rosendale, L.; Peiro, J.L. Non-Targeted Metabolic Profiling of Cerebellum in Spina Bifida Fetal Rats. Metabolites 2023, 13, 670. https://doi.org/10.3390/metabo13050670
Thielen E, Oria M, Watanabe-Chailland M, Lampe K, Romick-Rosendale L, Peiro JL. Non-Targeted Metabolic Profiling of Cerebellum in Spina Bifida Fetal Rats. Metabolites. 2023; 13(5):670. https://doi.org/10.3390/metabo13050670
Chicago/Turabian StyleThielen, Evan, Marc Oria, Miki Watanabe-Chailland, Kristin Lampe, Lindsey Romick-Rosendale, and Jose L. Peiro. 2023. "Non-Targeted Metabolic Profiling of Cerebellum in Spina Bifida Fetal Rats" Metabolites 13, no. 5: 670. https://doi.org/10.3390/metabo13050670