
Interactive Effect of Dietary Gamma-Aminobutyric Acid (GABA) and Water Temperature on Growth Performance, Blood Plasma Indices, Heat Shock Proteins and GABAergic Gene Expression in Juvenile Olive Flounder Paralichthys olivaceus
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Experimental Design
2.3. Experimental Fish and Condition
2.4. Sample Collection
2.5. Sample Analyses
2.6. Calculation and Statistical Analyses
3. Results
3.1. Effects of GABA and Water Temperature on Growth and Feed Utilization in Olive Flounder
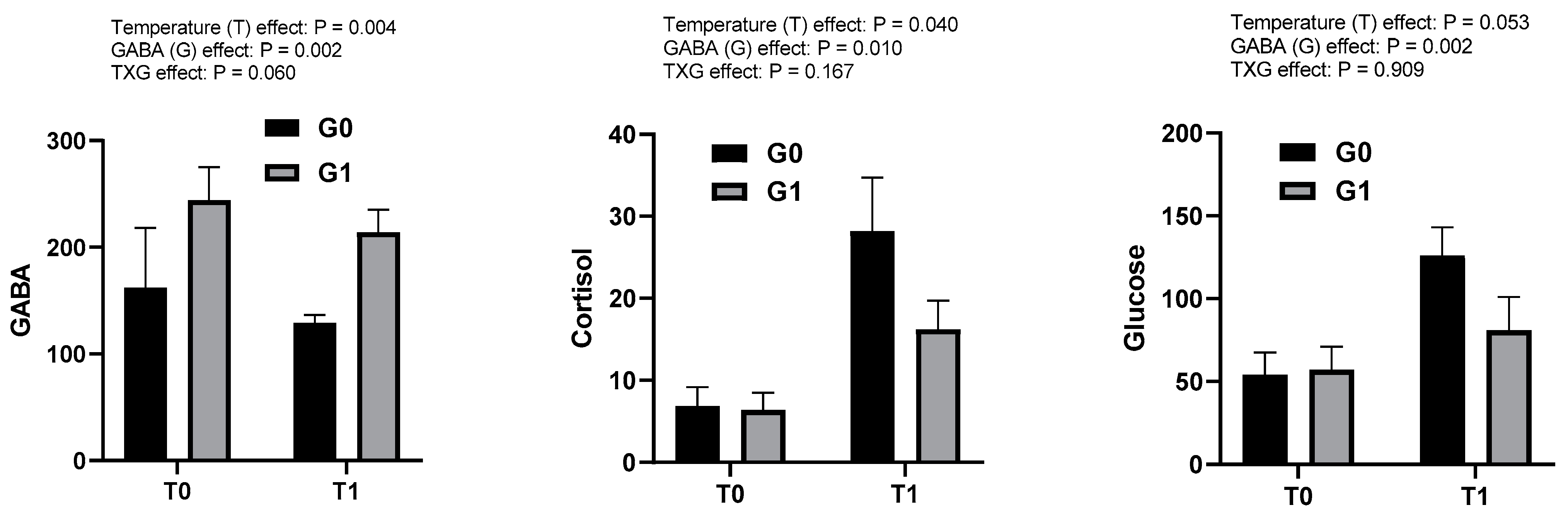
3.2. Effects of GABA and Water Temperature on Blood Plasma Indices in Olive Flounder
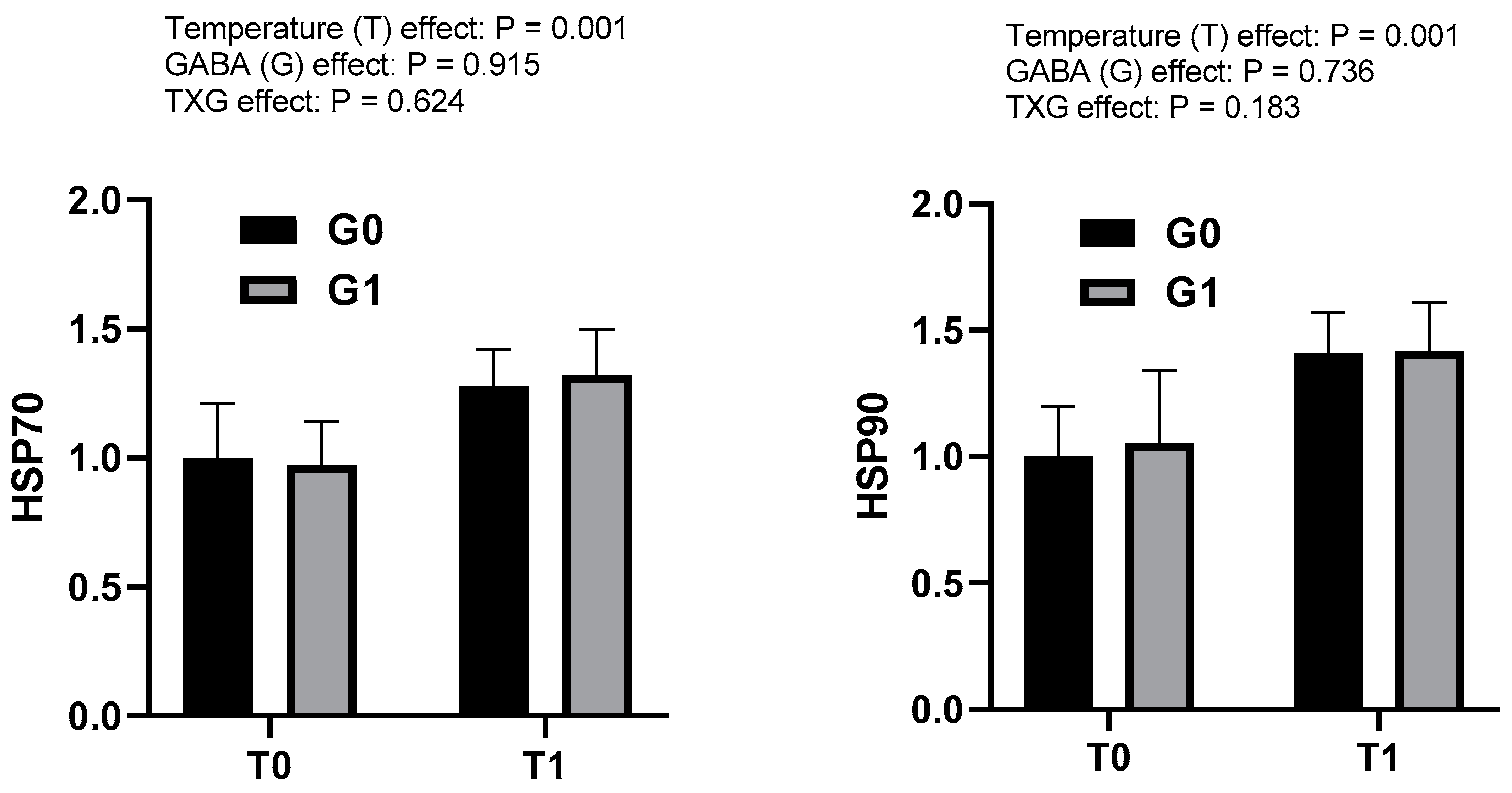
3.3. Effects of GABA and Water Temperature on Heat Shock Protein and GABA-Related Gene Expression in Olive Flounder
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, E.; Frankel, S. Aminobutyric acid in brain: Its formation from glutamic acid. J. Biol. Chem. 1950, 187, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Heli, Z.; Hongyu, C.; Dapeng, B.; Shin, T.Y.; Yejun, Z.; Xi, Z.; Yingying, W. Recent advances of γ-aminobutyric acid: Physiological and immunity function, enrichment, and metabolic pathway. Front. Nutr. 2022, 9, 1076223. [Google Scholar] [CrossRef] [PubMed]
- Tabor, R.; Yaksi, E.; Friedrich, R.W. Multiple functions of GABAA and GABAB receptors during pattern processing in the zebrafish olfactory bulb. Eur. J. Neurosci. 2008, 28, 117–127. [Google Scholar] [CrossRef]
- Mihic, S.J.; Harris, R.A. GABA and the GABAA Receptor. Alcohol Health Res. World 1997, 21, 127–131. [Google Scholar] [PubMed]
- Facciolo, R.M.; Crudo, M.; Zizza, M.; Giusi, G.; Canonaco, M. Feeding behaviors and ORXR–β-GABAAR subunit interactions in Carassius auratus. Neurotoxicol. Teratol. 2011, 33, 641–650. [Google Scholar] [CrossRef]
- Snigirov, S.; Sylantyev, S. GABAA receptors activate fish feeding behaviour via two distinct functional pathways. J. Exp. Biol. 2018, 221, jeb170514. [Google Scholar] [CrossRef] [PubMed]
- Sadamitsu, K.; Shigemitsu, L.; Suzuki, M.; Ito, D.; Kashima, M.; Hirata, H. Characterization of zebrafish GABAA receptor subunits. Sci. Rep. 2021, 11, 6242. [Google Scholar] [CrossRef] [PubMed]
- Gassmann, M.; Bettler, B. Regulation of neuronal GABAB receptor functions by subunit composition. Nat. Rev. Neurosci. 2012, 13, 380–394. [Google Scholar] [CrossRef]
- Wu, F.; Liu, M.; Chen, C.; Chen, J.; Tan, Q. Effects of dietary gamma aminobutyric acid on growth performance, antioxidant status, and feeding-related gene expression of juvenile grass carp, Ctenopharyngodon idellus. J. World Aquac. Soc. 2016, 47, 820–829. [Google Scholar] [CrossRef]
- Xie, S.-W.; Li, Y.-T.; Zhou, W.-W.; Tian, L.-X.; Zeng, S.-L.; Liu, Y.-J. Effect of γ-aminobutyric acid supplementation on growth performance, endocrine hormone and stress tolerance of juvenile Pacific white shrimp, Litopenaeus vannamei, fed low fishmeal diet. Aquac. Nutr. 2017, 23, 54–62. [Google Scholar] [CrossRef]
- Temu, V.; Kim, H.; Hamidoghli, A.; Park, M.; Won, S.; Oh, M.; Han, J.; Bai, S.C. Effects of dietary gamma-aminobutyric acid in juvenile Nile tilapia, Orechromis niloticus. Aquaculture 2019, 507, 475–480. [Google Scholar] [CrossRef]
- Bae, J.; Hamidoghli, A.; Farris, N.W.; Olowe, O.S.; Choi, W.; Lee, S.; Won, S.; Ohh, M.; Lee, S.H.; Bai, S.C. Dietary γ-aminobutyric acid (GABA) promotes growth and resistance to Vibrio alginolyticus in whiteleg shrimp Litopenaeus vannamei. Aquac. Nutr. 2022, 2022, 9105068. [Google Scholar] [CrossRef]
- Farris, N.W.; Hamidoghli, A.; Bae, J.; Choi, W.; Biro, J.; Lee, S.H.; Bai, S.C. Dietary supplementation with γ-aminobutyric acid improves growth, digestive enzyme activity, non-specific immunity and disease resistance against Streptococcus iniae in juvenile olive flounder, Paralichthys olivaceus. Animals 2022, 12, 248. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.E.; Hooker, N.; Duncan, H.; Jensen, L. γ-Aminobutyric acid, a neurotransmitter, induces planktonic abalone larvae to settle and begin metamorphosis. Science 1979, 204, 407–410. [Google Scholar] [CrossRef]
- Searcy-Bernal, R.; Anguiano-Beltrán, C. Optimizing the concentration of gamma-aminobutyric acid (GABA) for inducing larval metamorphosis in the red abalone Haliotis rufescens (Mollusca: Gastropoda). J. World Aquac. Soc. 1998, 29, 463–470. [Google Scholar] [CrossRef]
- Li, J.; Jiang, M.; Han, Q.; Peng, R.; Jiang, X. Effects of γ-aminobutyric acid supplementation on the growth performance, serum biochemical indices and antioxidant status of pharaoh cuttlefish, Sepia pharaonic. Aquac. Nutr. 2020, 26, 1026–1034. [Google Scholar] [CrossRef]
- Chen, X.; Gao, C.; Du, X.; Xu, H.; Wang, G.; Zhang, D. Effects of dietary γ-aminobutyric acid levels on the growth, serum biochemical indexes, immune-related signaling molecules of Jian carp. Aquac. Res. 2021, 52, 1096–1105. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Wang, C.; Song, Y.; Pan, J.; Shi, Q.; Qin, J.; Chen, L. Gamma-aminobutyric acid regulates glucose homeostasis and enhances the hepatopancreas health of juvenile Chinese mitten crab (Eriocheir sinensis) under fasting stress. Gen. Comp. Endocrinol. 2021, 303, 113704. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Su, R.; He, J.; Liu, S.; Huang, Q.; Qin, C.; Zhang, M.; Qin, J.; Chen, L. Dietary gamma-aminobutyric acid (GABA) supplementation increases food intake, influences the expression of feeding-related genes and improves digestion and growth of Chinese mitten crab (Eriocheir sinensis). Aquaculture 2022, 546, 737332. [Google Scholar] [CrossRef]
- Lv, H.; Liu, Y.; Li, H.; Yin, X.; Wang, P.; Qu, X.; Gao, Y. Modulation of Antioxidant Enzymes, Heat Shock Protein, and Intestinal Microbiota of Large Yellow Croaker (Larimichthys crocea) Under Acute Cold Stress. Front. Mar. Sci. 2021, 8, 725899. [Google Scholar] [CrossRef]
- Li, C.; Tian, Y.; Ma, Q.; Zhang, B. Dietary gammaaminobutyric acid ameliorates growth impairment and intestinal dysfunction in turbot (Scophthalmus maximus L.) fed a high soybean meal diet. Food Funct. 2022, 13, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, G.E.; Lefevre, S. Physiological Challenges to Fishes in a Warmer and Acidified Future. Physiology 2016, 31, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Dickson, K.A.; Graham, J.B. Evolution and consequences of endothermy in fishes. Physiol. Biochem. Zool. 2004, 77, 998–1018. [Google Scholar] [CrossRef]
- Bai, S.C.; Kim, K.W. Effects of dietary animal protein sources on growth and body composition in Korean rockfish, Sebastes schlegelii. J. Aquac. 1997, 10, 77–85. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists (AOAC): Arlington, VA, USA, 1995. [Google Scholar]
- Goel, A.; Ncho, C.M.; Jeong, C.M.; Choi, Y.H. Embryonic thermal manipulation and in ovo gamma-aminobutyric acid supplementation regulating the chick weight and stress-related genes at hatch. Front. Vet. Sci. 2022, 8, 807450. [Google Scholar] [CrossRef] [PubMed]
- Ncho, C.M.; Goel, A.; Gupta, V.; Jeong, C.M.; Choi, Y.H. Embryonic manipulations modulate differential expressions of heat shock protein, fatty acid metabolism, and antioxidant-related genes in the liver of heat-stressed broilers. PLoS ONE 2022, 17, e0269748. [Google Scholar] [CrossRef]
- El-Naggar, K.; El-Kassas, S.; Abdo, S.E.; Kirrella, A.A.K.; Al-Wakeel, R.A. Role of gamma-aminobutyric acid in regulating feed intake in commercial broilers reared under normal and heat stress conditions. J. Therm. Biol. 2019, 84, 164–175. [Google Scholar] [CrossRef]
- Volkoff, H. The neuroendocrine regulation of food intake in fish: A review of current knowledge. Front. Neurosci. 2016, 10, 540. [Google Scholar] [CrossRef]
- Delgado, M.J.; Cerda-Reverter, J.M.; Soengas, J.L. Hypothalamic integration of metabolic, endocrine, and circadian signals in fish: Involvement in the control of food intake. Front. Neurosci. 2017, 11, 354. [Google Scholar] [CrossRef]
- Conde-Sieira, M.; Chivite, M.; Miguez, J.M.; Soengas, J.L. Stress effects on the mechanisms regulating appetite in teleost fish. Front. Endocrinol. 2018, 9, 631. [Google Scholar] [CrossRef]
- Dong, H.; Fang, L.X.; Chen, Y.X.; Jing, C.W.; Xuan, L.A.; Shan, H.E.; Min, X. Effects of γ-aminobutyric acid on food intake and appetite in mandarin fish Siniperca chuatsi. Acta Hydrobiol. Sin. 2017, 41, 1311–1317. [Google Scholar]
- Jentoft, S.; Aastveit, A.H.; Torjesen, P.A.; Andersen, O. Effects of stress on growth, cortisol and glucose levels in non-domesticated Eurasian perch (Perca fluviatilis) and domesticated rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Phycol. Part A 2002, 141, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Servili, A.; Leveque, E.; Mouchel, O.; Devergne, J.; Lebigre, C.; Roussel, S.; Mazurais, D.; Zambonino-Infante, J.L. Ocean acidification alters the acute stress response of a marine fish. Sci. Tot. Environ. 2023, 858, 159804. [Google Scholar] [CrossRef] [PubMed]
- Balasch, J.C.; Tort, L. Netting the stress responses in fish. Front. Endocrinol. 2019, 10, 62. [Google Scholar] [CrossRef] [PubMed]
- De Abreu, M.S.; Demin, K.A.; Giacomini, A.C.V.V.; Amstislavskaya, T.G.; Strekalova, T.; Maslov, G.O.; Kositsin, Y.; Petersen, E.V.; Kalueff, A.V. Understanding how stress responses and stress-related behaviors have evolved in zebrafish and mammals. Neurobiol. Stress 2021, 15, 1000405. [Google Scholar] [CrossRef]
- Sohrabipour, S.; Sharifi, M.R.; Talebi, A.; Sharifi, M.; Soltani, N. GABA dramatically improves glucose tolerance in streptozotocin-induced diabetic rats fed with high-fat diet. Eur. J. Pharmacol. 2018, 826, 75–84. [Google Scholar] [CrossRef]
- Dai, S.F.; Gao, F.; Zhang, W.H.; Song, S.X.; Xu, X.L.; Zhou, G.H. Effects of dietary glutamine and gamma-aminobutyric acid on performance, carcass characteristics and serum parameters in broilers under circular heat stress. Anim. Feed Sci. Technol. 2011, 168, 51–60. [Google Scholar] [CrossRef]
- Facciolo, R.M.; Crudo, M.; Giusi, G.; Canonaco, M. GABAergic influences on ORX receptor-dependent abnormal motor behaviors and neurodegenerative events in fish. Toxicol. Appl. Pharmacol. 2010, 243, 77–86. [Google Scholar] [CrossRef]
- Martyniuk, C.J.; Crawford, A.B.; Hogan, N.S.; Trudeau, V.L. GABAergic modulation of the expression of genes involved in GABA synaptic transmission and stress in the hypothalamus and telencephalon of the female goldfish (Carassius auratus). J. Neuroendocrinol. 2005, 17, 269–275. [Google Scholar] [CrossRef]
- Grone, B.P.; Maruska, K.P. Three Distinct Glutamate Decarboxylase Genes in Vertebrates. Sci. Rep. 2016, 6, 30507. [Google Scholar] [CrossRef]
- Hamilton, T.J.; Holcombe, A.; Tresguerres, M. CO2-induced ocean acidification increases anxiety in Rockfish via alteration of GABAA receptor functioning. Proc. R. Soc. B 2013, 281, 20132509. [Google Scholar] [CrossRef] [PubMed]
- Schunter, C.; Ravasi, T.; Munday, P.L.; Nilsson, G.E. Neural effects of elevated CO2 in fish may be amplified by a vicious cycle. Conserv. Physiol. 2019, 7, coz100. [Google Scholar] [CrossRef] [PubMed]
- Hossain-Javaheri, N.; Buck, L.T. GABA receptor inhibition and severe hypoxia induce a paroxysmal depolarization shift in goldfish neurons. J. Neurophysiol. 2021, 125, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J.; Attwell, D. The effect of simulated ischaemia on spontaneous GABA release in area CA1 of the juvenile rat hippocampus. J. Physiol. 2004, 561, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.J.; Xie, X.Z.; Zhou, Y.W.; Liang, C.; Jiang, Y.Y.; Chen, Z. Effect of Heat Stress on the Expression of GABA Receptor mRNA in the HPG Axis of Wenchang Chickens. Braz. J. Poult. Sci. 2016, 18, 277–282. [Google Scholar] [CrossRef]
- Heuer, R.M.; Grosell, M. Physiological impacts of elevated carbon dioxide and ocean acidification on fish. Am. J. Physiol. Reg. Int. Comp. Physiol. 2014, 307, R1061–R1084. [Google Scholar] [CrossRef]
- Goodman, A.; Wong, R.Y. Differential effects of ethanol on behavior and GABAA receptor expression in adult zebrafish (Danio rerio) with alternative stress coping styles. Sci. Rep. 2020, 10, 13076. [Google Scholar] [CrossRef]
- Rahman, M.M.; Hyeonho, Y.; Moniruzzaman, M.; Ferreira, F.; Kim, K.W.; Bai, S.C. Effects of Feeding Rate and Water Temperature on Growth and Body Composition of Juvenile Korean Rockfish, Sebastes schlegeli (Hilgendorf). Asian Australas. J. Anim. Sci. 2014, 27, 690–699. [Google Scholar]
- Oksala, N.K.J.; Ekmekci, F.G.; Ozsoy, E.; Kirankaya, S.; Kokkola, T.; Emecen, G.; Lappalainen, J.; Kaarniranta, K.; Atalay, M. Natural thermal adaptation increases heat shock protein levels and decreases oxidative stress. Redox Biol. 2014, 3, 25–28. [Google Scholar] [CrossRef]
- Basu, N.; Todgham, A.E.; Ackerman, P.A.; Bibeau, M.R.; Nakano, K.; Schulte, P.M. Heat shock protein genes and their functional significance in fish. Gene 2002, 295, 173–183. [Google Scholar] [CrossRef]
- Lei, L.; Yu, J.; Bao, E. Expression of heat shock protein 90 (hsp90) and transcription of its corresponding mrna in broilers exposed to high temperature. Br. Poult. Sci. 2009, 50, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Morita, V.D.S.; de Almeida, V.R.; Matos, J.B.J.; Vicentini, T.I.; van den Brand, H.; Boleli, I.C. Incubation temperature during fetal development influences morphophysiological characteristics and preferred ambient temperature of chicken hatchlings. PLoS ONE 2016, 11, e0154928. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Ingredients | Diets | |
|---|---|---|
| CON | GABA | |
| Fish meal (sardine) 1 | 50.0 | 50.0 |
| Poultry by-product meal 2 | 6.0 | 6.0 |
| Soybean meal 2 | 8.0 | 8.0 |
| Soy protein concentrate 2 | 3.0 | 3.0 |
| Corn protein concentrate 2 | 3.0 | 3.0 |
| Wheat gluten meal 2 | 1.5 | 1.5 |
| Wheat flour 2 | 19.2 | 19.2 |
| Fish oil (menhaden) 3 | 6.0 | 6.0 |
| Choline chloride (60%) 2 | 0.6 | 0.6 |
| Vitamin C (Stay C, 35%) | 0.2 | 0.2 |
| Vitamin premix 4 | 1.0 | 1.0 |
| Mineral premix 5 | 1.0 | 1.0 |
| Cellulose 6 | 0.5 | 0.0 |
| GABA 7 (40,000 ppm) | 0.0 | 0.5 |
| Proximate analysis (% dry matter basis) | ||
| Moisture | 7.3 | 7.5 |
| Crude ash | 10.9 | 11.1 |
| Crude lipid | 10.9 | 11.1 |
| Crude protein | 52.0 | 51.8 |
| Primers | Sense | Sequences (5′-3′) | Product Size (bp) | Accession Number | Annealing Temperature (°C) |
|---|---|---|---|---|---|
| gabarap a | Forward | AGTGATGAGAGTGTGTATGGG | 204 | XM_020095803 | 60 |
| Reverse | AGAAATGGATGGGAGAAGGG | ||||
| gabbr1 b | Forward | TCCTTTGCCTTTGCCTCTC | 161 | XM_020104954 | 60 |
| Reverse | CCTCGTCGTTGTTGTTGTC | ||||
| gad1 c | Forward | AGCAGGATCGTGGGTTCCCT | 105 | XM_020089610 | 60 |
| Reverse | GAGAAGTCCGTCTCCGTGCG | ||||
| hsp70 d | Forward | CAGTCCAGGCTGCTATCCTCAT | 102 | AB010871 | 60 |
| Reverse | TCATGACTCCACCAGCAGTCTC | ||||
| hsp90 e | Forward | GAGCGAGACAAGGAGGTGAG | 101 | KY856948 | 60 |
| Reverse | CTGGCTTGTCTTCGTCCTTC | ||||
| β-actin | Forward | CAGCATCATGAAGTGTGACGTG | 200 | HQ386788.1 | 60 |
| Reverse | CTTCTGCATACGGTCAGCAATG |
| Treatments | IW (g/fish) | FBW (g/fish) | WG (g/fish) | WG (%/fish) | SGR (%/day) | Feed Intake (g/fish) | FCR | Survival (%) | |
|---|---|---|---|---|---|---|---|---|---|
| Temperature | Dietary GABA | ||||||||
| Normal Temperature | 0 | 36.8 ± 0.2 a | 62.4 ± 0.8 a | 25.6 ± 0.7 a | 69.6 ± 1.5 a | 2.20 ± 0.1 a | 28.9 ± 0.2 a | 1.13 ± 0.1 c | 100 ± 0.0 a |
| 200 | 36.8 ± 0.7 a | 62.7 ± 0.2 a | 25.9 ± 0.5 a | 70.6 ± 2.9 a | 2.23 ± 0.1 a | 29.0 ± 0.3 a | 1.12 ±0.1 c | 100 ± 0.0 a | |
| High Temperature | 0 | 37.0 ± 0.1 a | 58.1 ± 0.5 c | 21.1 ± 0.5 c | 56.8 ± 1.1 c | 1.87 ± 0.1 c | 28.6 ± 0.3 a | 1.36 ± 0.1 a | 95.6 ± 3.1 a |
| 200 | 36.9 ± 0.3 a | 60.3 ± 0.2 b | 23.4 ± 0.2 b | 63.5 ± 0.9 b | 2.05 ± 0.1 b | 28.9 ± 0.5 a | 1.23 ± 0.1 b | 97.8 ± 3.1 a | |
| Means of main effect a | |||||||||
| Temperature | Normal | 36.8 a | 62.6 a | 25.7 a | 70.1 a | 2.21 a | 29.0 a | 1.12 b | 100 a |
| High | 36.9 a | 59.2 b | 22.2 b | 60.1 b | 1.96 b | 28.7 a | 1.30 a | 96.7 a | |
| GABA | GABA0 | 36.9 a | 60.2 b | 23.3 b | 63.2 b | 2.04 b | 28.8 a | 1.24 a | 97.8 a |
| GABA200 | 36.8 a | 61.5 a | 24.7 a | 67.0 a | 2.14 a | 28.9 a | 1.18 b | 98.9 a | |
| Two-way ANOVA (p value) | |||||||||
| Temperature effect | 0.601 | <0.001 | <0.001 | <0.001 | <0.001 | 0.356 | <0.001 | 0.067 | |
| GABA effect | 0.675 | 0.010 | 0.005 | 0.018 | 0.018 | 0.420 | 0.005 | 0.500 | |
| Temperature × GABA | 0.916 | 0.029 | 0.022 | 0.062 | 0.034 | 0.752 | 0.012 | 0.500 | |
| Treatments | Gabarap | Gabbr1 | Gad1 | |
|---|---|---|---|---|
| Temperature | Dietary GABA | |||
| Normal Temperature | 0 | 1.00 ± 0.2 a | 1.00 ± 0.3 b | 1.00 ± 0.2 a |
| 200 | 0.97 ± 0.3 a | 1.22 ± 0.3 ab | 0.73 ± 0.1 a | |
| High Temperature | 0 | 0.96 ± 0.1 a | 1.08 ± 0.1 ab | 0.95 ± 0.2 a |
| 200 | 0.94 ± 0.1 a | 1.49 ± 0.3 a | 0.73 ± 0.3 a | |
| Means of main effect a | ||||
| Temperature | Normal | 0.99 a | 1.11 a | 0.87 a |
| High | 0.95 a | 1.28 a | 0.84 a | |
| GABA | GABA0 | 0.98 a | 1.04 b | 0.97 a |
| GABA200 | 0.96 a | 1.36 a | 0.73 b | |
| Two-way ANOVA (p value) | ||||
| Temperature | 0.714 | 0.177 | 0.783 | |
| GABA | 0.777 | 0.017 | 0.018 | |
| Temperature × GABA | 0.953 | 0.446 | 0.800 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Moniruzzaman, M.; Farris, N.; Min, T.; Bai, S.C. Interactive Effect of Dietary Gamma-Aminobutyric Acid (GABA) and Water Temperature on Growth Performance, Blood Plasma Indices, Heat Shock Proteins and GABAergic Gene Expression in Juvenile Olive Flounder Paralichthys olivaceus. Metabolites 2023, 13, 619. https://doi.org/10.3390/metabo13050619
Lee S, Moniruzzaman M, Farris N, Min T, Bai SC. Interactive Effect of Dietary Gamma-Aminobutyric Acid (GABA) and Water Temperature on Growth Performance, Blood Plasma Indices, Heat Shock Proteins and GABAergic Gene Expression in Juvenile Olive Flounder Paralichthys olivaceus. Metabolites. 2023; 13(5):619. https://doi.org/10.3390/metabo13050619
Chicago/Turabian StyleLee, Seunghan, Mohammad Moniruzzaman, Nathaniel Farris, Taesun Min, and Sungchul C. Bai. 2023. "Interactive Effect of Dietary Gamma-Aminobutyric Acid (GABA) and Water Temperature on Growth Performance, Blood Plasma Indices, Heat Shock Proteins and GABAergic Gene Expression in Juvenile Olive Flounder Paralichthys olivaceus" Metabolites 13, no. 5: 619. https://doi.org/10.3390/metabo13050619