
The Effects of Rumen-Protected Choline and Rumen-Protected Nicotinamide on Liver Transcriptomics in Periparturient Dairy Cows

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Sample Acquisition
2.2. Culture of Human Hepatocyte Line LO2
2.3. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction
2.4. RNA Isolation and cDNA Library Construction
2.5. Analysis of Differential Expression of Genes
2.6. Analysis of DEG Enrichment
2.7. Statistical Analysis
3. Results
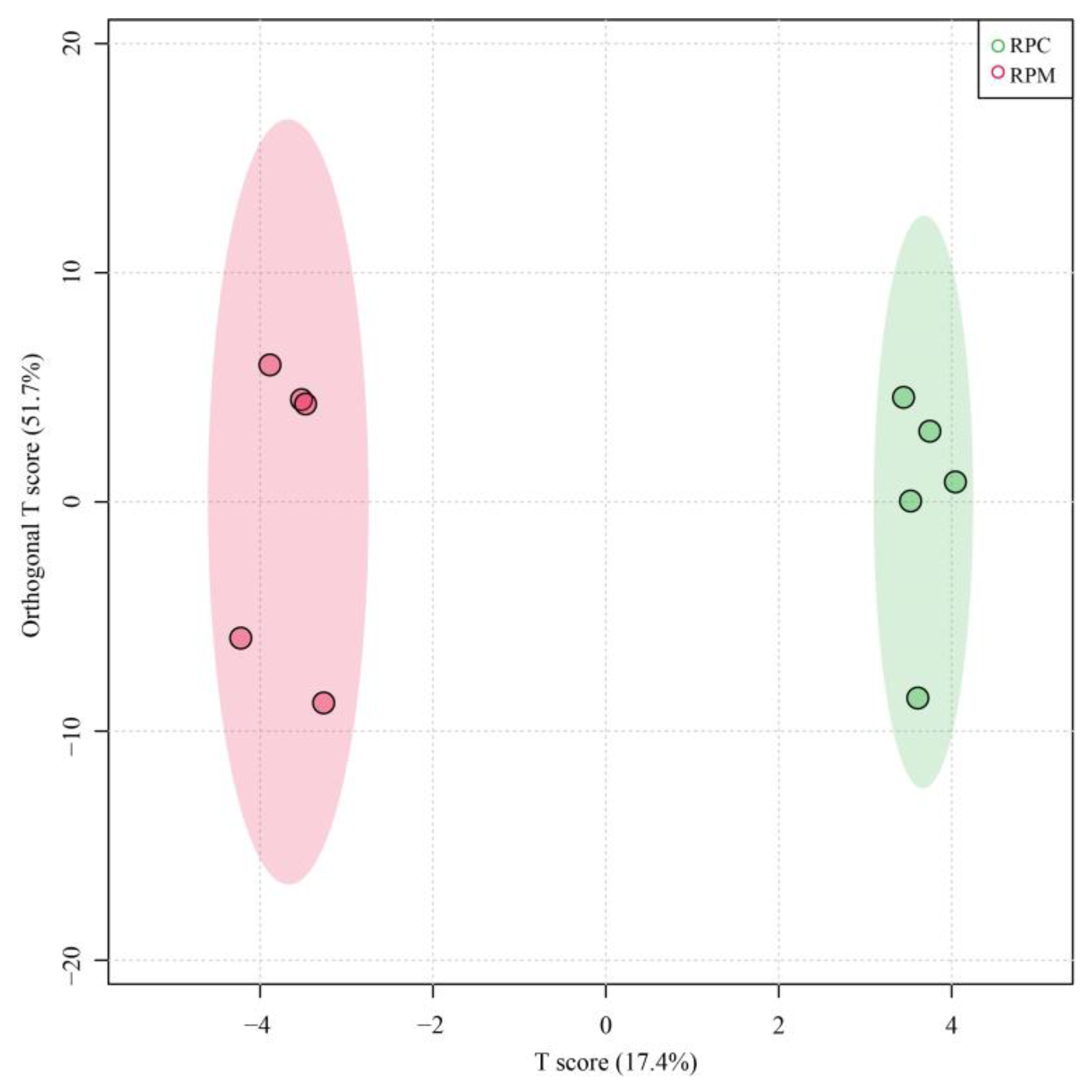
3.1. Sequencing Data Quality Control
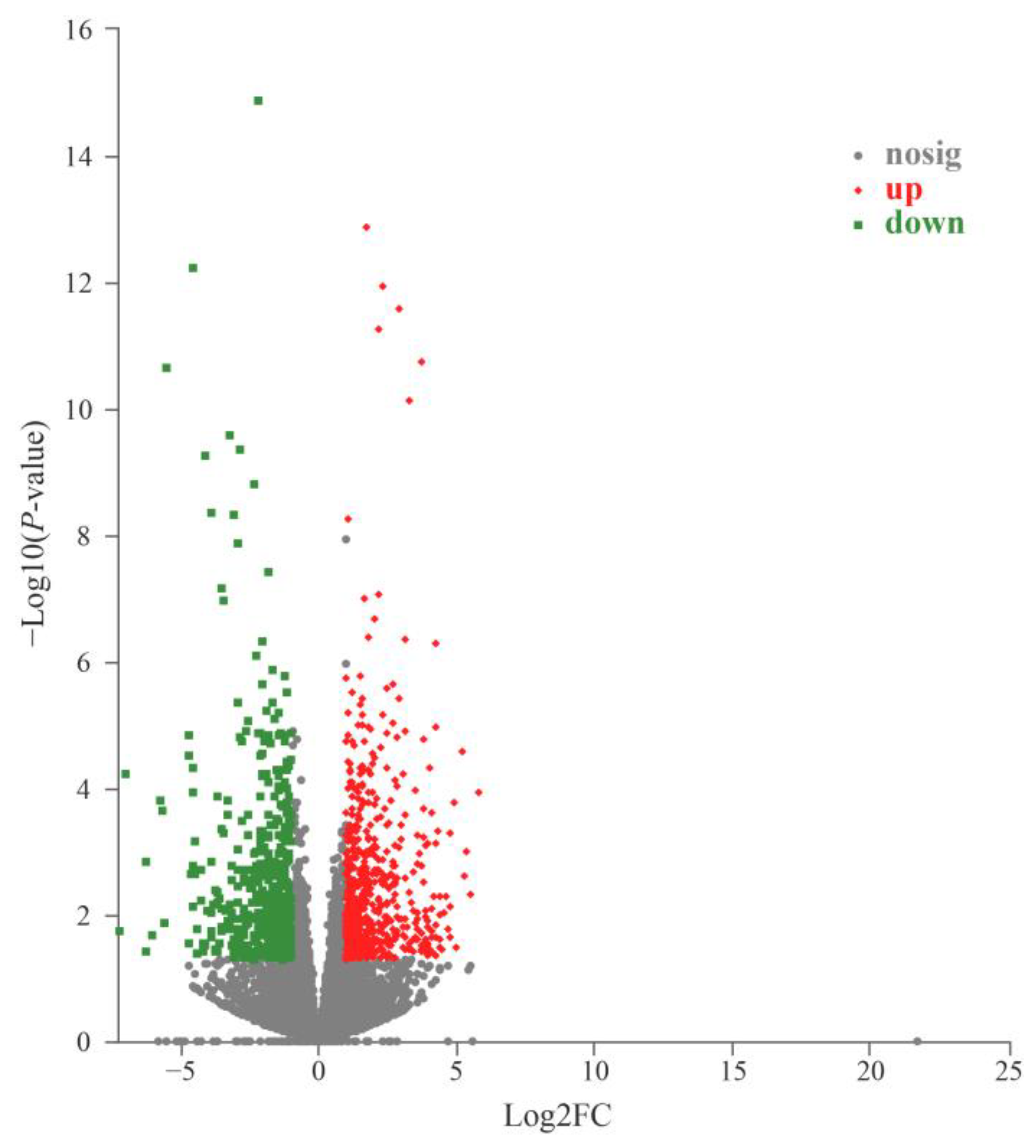
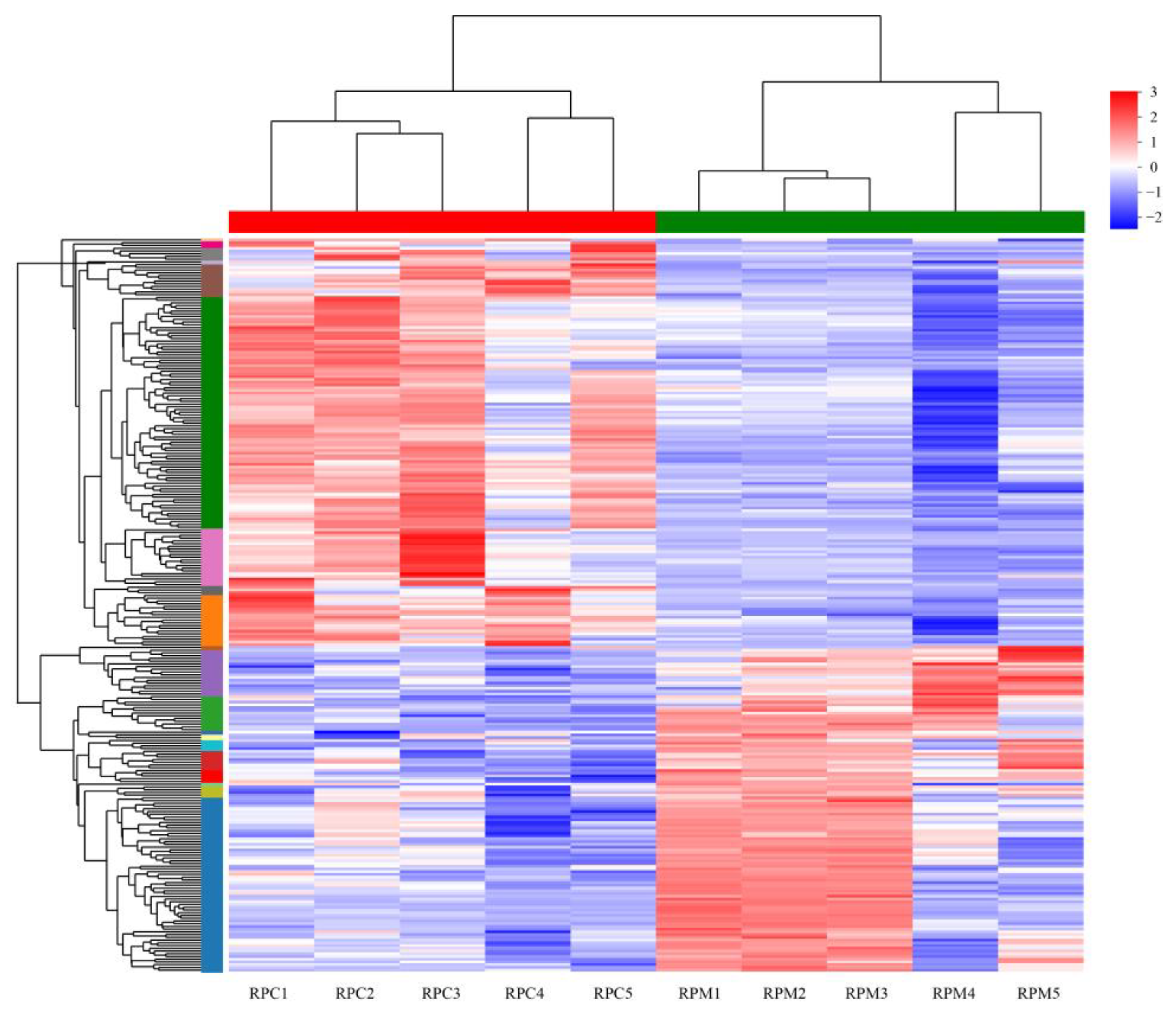
3.2. Analysis of DEGs
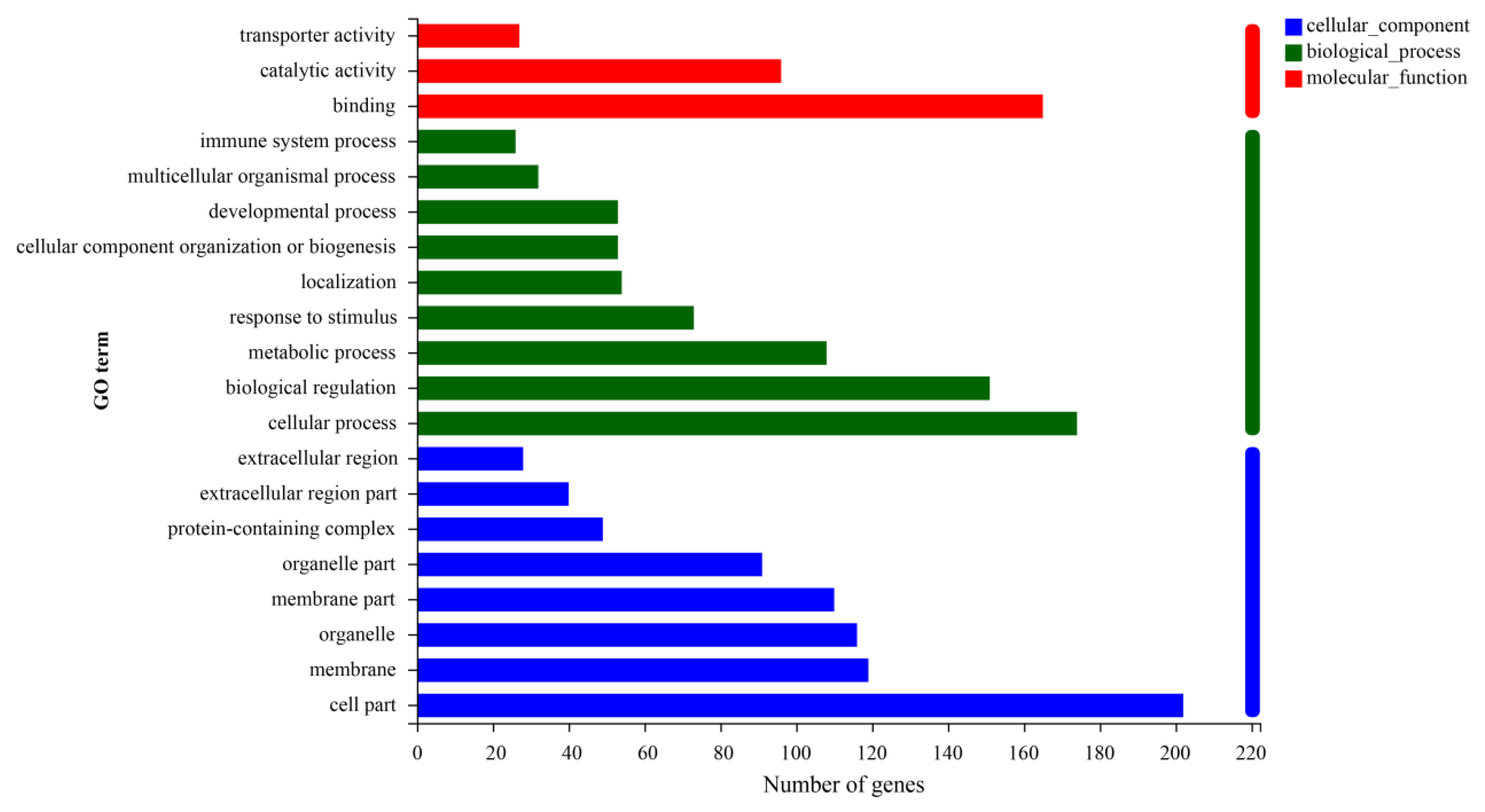
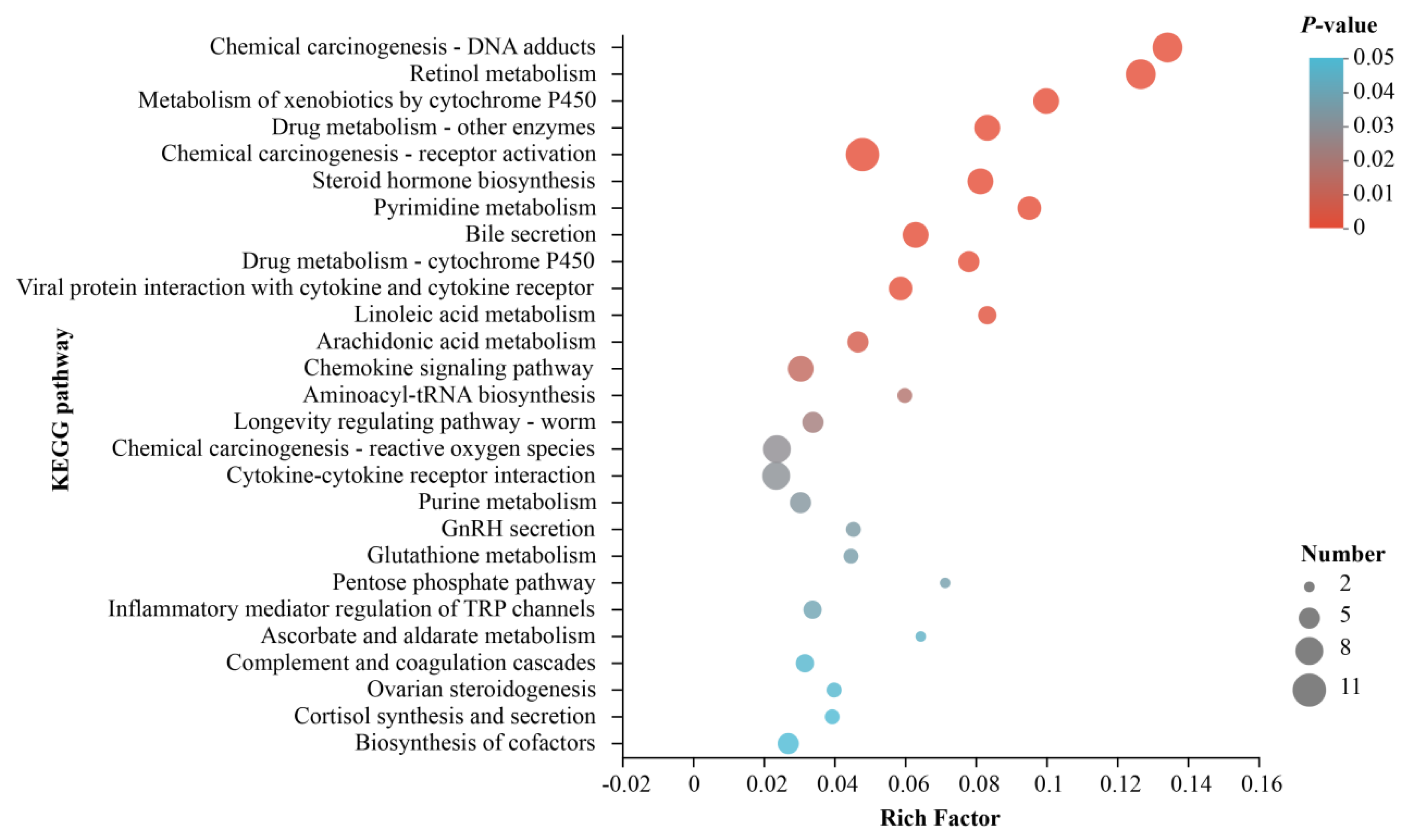
3.3. Analysis of Differential Gene Enrichment
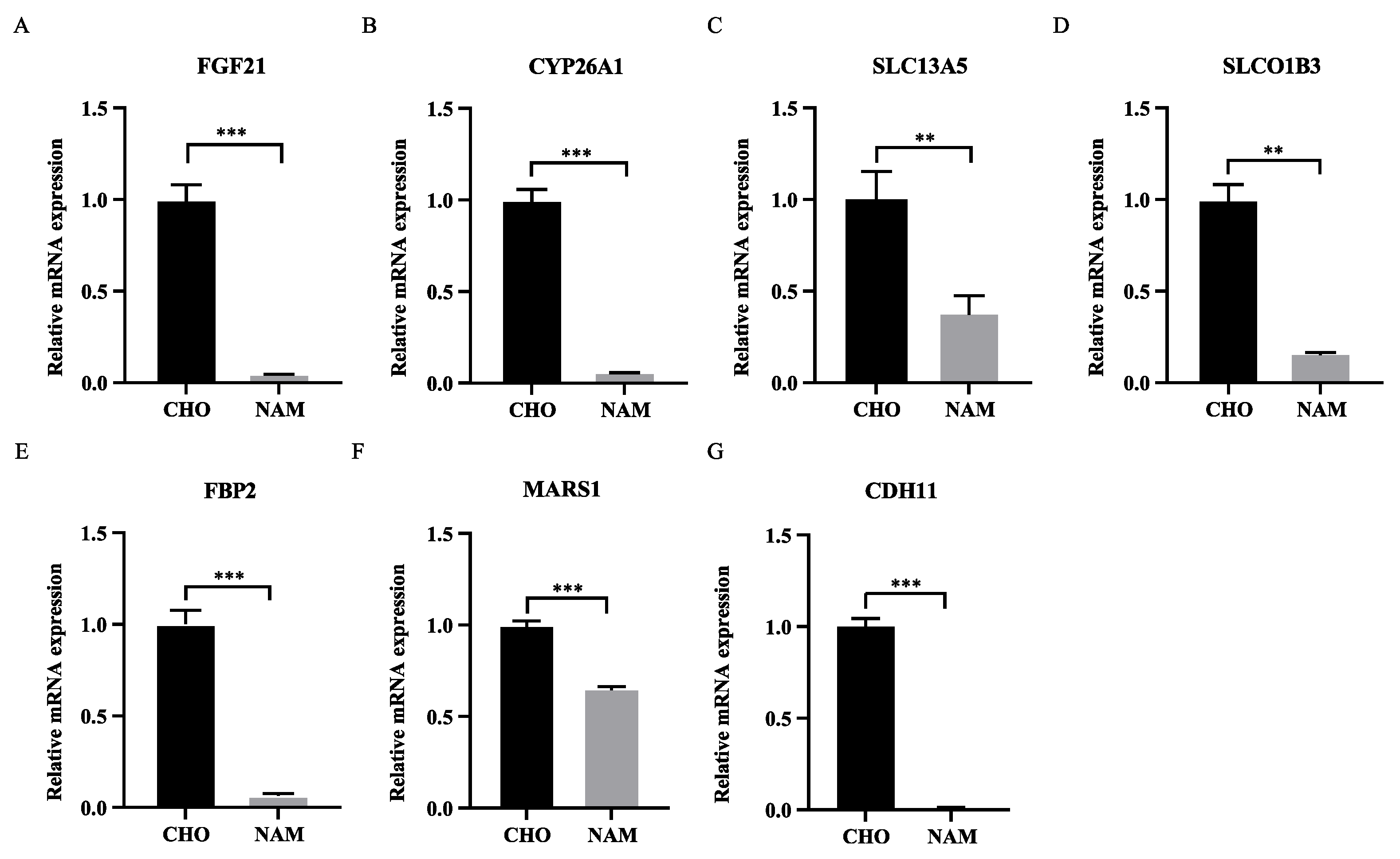
3.4. Effects of Choline and Nicotinamide on Gene Expression Related to Liver Metabolism
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bazzano, M.; Giannetto, C.; Fazio, F.; Arfuso, F.; Giudice, E.; Piccione, G. Metabolic profile of broodmares during late pregnancy and early post-partum. Reprod. Domest. Anim. 2014, 49, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Minuti, A.; Liotta, L.; Giannetto, C.; Trevisi, E.; Piccione, G.; Lopreiato, V. Stress and inflammatory response of cows and their calves during peripartum and early neonatal period. Theriogenology 2023, 196, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Tessari, R.; Berlanda, M.; Morgante, M.; Badon, T.; Gianesella, M.; Mazzotta, E.; Contiero, B.; Fiore, E. Changes of plasma fatty acids in four lipid classes to understand energy metabolism at different levels of non-esterified fatty acid (NEFA) in dairy cows. Animals 2020, 10, 1410. [Google Scholar] [CrossRef] [PubMed]
- Yepes, F.A.L.; Nydam, D.V.; Mann, S.; Caixeta, L.; Mcart, J.A.A.; Overton, T.R.; Wakshlag, J.; Huson, H.J. Longitudinal Phenotypes Improve Genotype Association for Hyperketonemia in Dairy Cattle. Animals 2019, 9, 1059. [Google Scholar] [CrossRef] [PubMed]
- Pascottini, O.B.; Leroy, J.L.M.R.; Opsomer, G. Metabolic stress in the transition period of dairy cows: Focusing on the prepartum period. Animals 2020, 10, 1419. [Google Scholar] [CrossRef]
- Van Saun, R.J.; Sniffen, C.J. Transition cow nutrition and feeding management for disease prevention. Vet. Clin. N. Am. Food. Anim. Pract. 2014, 30, 689–719. [Google Scholar] [CrossRef]
- Koeck, A.; Miglior, F.; Jamrozik, J.; Kelton, D.F.; Schenkel, F.S. Genetic associations of ketosis and displaced abomasum with milk production traits in early first lactation of Canadian Holsteins. J. Dairy Sci. 2013, 96, 4688–4696. [Google Scholar] [CrossRef]
- Jorritsma, R.; Wensing, T.; Kruip, T.A.; Vos, P.L.; Noordhuizen, J.P. Metabolic changes in early lactation and impaired reproductive performance in dairy cows. Vet. Res. Commun. 2003, 34, 11–26. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Moyes, K.M. Factors contributing to immunosuppression in the dairy cow during the periparturient period. JPN J. Vet. Res. 2015, 63 (Suppl. S1), S15–S24. [Google Scholar]
- Korsmo, H.W.; Jiang, X.; Caudill, M.A. Choline: Exploring the growing science on its benefits for moms and babies. Nutrients 2019, 11, 1823. [Google Scholar] [CrossRef]
- Zeisel, S.H. Metabolic crosstalk between choline/1-carbon metabolism and energy homeostasis. Clin. Chem. Lab. Med. 2013, 51, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Agellon, L.B.; Allen, T.M.; Umeda, M.; Jewell, L.; Mason, A.; Vance, D.E. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis. Cell Metab. 2006, 3, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Duszka, K.; Gregor, A.; Guillou, H.; König, J.; Wahli, W. Peroxisome proliferator-activated receptors and caloric restriction-common pathways affecting metabolism, health, and longevity. Cells 2020, 9, 1708. [Google Scholar] [CrossRef] [PubMed]
- Hedtke, V.; Bakovic, M. Choline transport for phospholipid synthesis: An emerging role of choline transporter-like protein 1. Exp. Biol. Med. 2019, 244, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Snider, S.A.; Margison, K.D.; Ghorbani, P.; Leblond, N.D.; O’dwyer, C.; Nunes, J.R.C.; Nguyen, T.; Xu, H.; Bennett, S.A.L.; Fullerton, M. Choline transport links macrophage phospholipid metabolism and inflammation. J. Biol. Chem. 2018, 293, 11600–11611. [Google Scholar] [CrossRef]
- Ahmadzadeh-Gavahan, L.; Hosseinkhani, A. Feed restriction and supplementing with propylene glycol, monensin sodium and rumen-protected choline chloride in periparturient Ghezel ewes: Implications on production and performance of ewes and their offspring. Livest. Prod. Sci. 2022, 255, 104784. [Google Scholar] [CrossRef]
- Guo, C.Z.; Xue, Y.F.; Yin, Y.; Sun, D.M.; Xuan, H.P.; Liu, J.; Mao, S.Y. The effect of glycerol or rumen-protected choline chloride on rumen fermentation and blood metabolome in pregnant ewes suffering from negative energy balance. Anim. Feed Sci. Technol. 2020, 268, 114594. [Google Scholar] [CrossRef]
- Elek, P.; Newbold, J.R.; Gaal, T.; Wagner, L.; Husveth, F. Effects of rumen-protected choline supplementation on milk production and choline supply of periparturient dairy cows. Animal 2008, 2, 1595–1601. [Google Scholar] [CrossRef]
- Goselink, R.M.; van Baal, J.; Widjaja, H.C.; Dekker, R.A.; Zom, R.L.; de Veth, M.J.; van Vuuren, A.M. Effect of rumen-protected choline supplementation on liver and adipose gene expression during the transition period in dairy cattle. J. Dairy Sci. 2013, 96, 1102–1116. [Google Scholar] [CrossRef]
- Zom, R.L.; van Baal, J.; Goselink, R.M.; Bakker, J.A.; de Veth, M.J.; van Vuuren, A.M. Effect of rumen-protected choline on performance, blood metabolites, and hepatic triacylglycerols of periparturient dairy cattle. Clin. Chem. Lab. Med. 2011, 94, 4016–4027. [Google Scholar] [CrossRef]
- Guretzky, N.A.; Carlson, D.B.; Garrett, J.; Drackley, J.K. Lipid metabolite profiles and milk production for Holstein and Jersey cows fed rumen-protected choline during the periparturient period. J. Dairy Sci. 2006, 89, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.H.; Brown, N.E.; Martinez, C.M.; Cassidy, T.W.; Varga, G.A. Effects of rumen-protected choline and dry propylene glycol on feed intake and blood parameters for Holstein dairy cows in early lactation. J. Dairy Sci. 2009, 92, 2729–2736. [Google Scholar] [CrossRef] [PubMed]
- Arshad, U.; Zenobi, M.G.; Staples, C.R.; Santos, J.E.P. Meta-analysis of the effects of supplemental rumen-protected choline during the transition period on performance and health of parous dairy cows. J. Dairy Sci. 2020, 103, 282–300. [Google Scholar] [CrossRef] [PubMed]
- Swartz, T.H.; Bradford, B.J.; Malysheva, O.; Caudill, M.A.; Mamedova, L.K.; Estes, K.A. Effects of dietary rumen-protected choline supplementation on colostrum yields, quality, and choline metabolites from dairy cattle. JDS. Commun. 2022, 3, 296–300. [Google Scholar] [CrossRef]
- Ahmadzadeh-Gavahan, L.; Hosseinkhani, A.; Taghizadeh, A.; Ghasemi-Panahi, B.; Hamidian, G.; Cheraghi-Saray, S.; Vakili, A. Impact of supplementing feed restricted ewes’ diet with propylene glycol, monensin sodium and rumen-protected choline chloride during late pregnancy on blood biochemical indices, body condition score and body weight. Anim. Feed. Sci. Technol. 2021, 273, 114801. [Google Scholar] [CrossRef]
- Morrison, E.I.; Reinhardt, H.; Leclerc, H.; DeVries, T.J.; LeBlanc, S.J. Effect of rumen-protected B vitamins and choline supplementation on health, production, and reproduction in transition dairy cows. J. Dairy Sci. 2018, 101, 9016–9027. [Google Scholar] [CrossRef]
- Vailati-Riboni, M.; Zhou, Z.; Jacometo, C.B.; Minuti, A.; Trevisi, E.; Luchini, D.N.; Loor, J.J. Supplementation with rumen-protected methionine or choline during the transition period influences whole-blood immune response in periparturient dairy cows. J. Dairy Sci. 2017, 100, 3958–3968. [Google Scholar] [CrossRef]
- Jayaprakash, G.; Sathiyabarathi, M.; Robert, M.A.; Tamilmani, T. Rumen-protected choline: A significance effect on dairy cattle nutrition. Vet. World 2016, 9, 837–841. [Google Scholar] [CrossRef]
- Swartz, T.H.; Bradford, B.J.; Lemke, M.; Mamedova, L.K.; Agnew, R.; Fehn, J.; Owczarzak, E.; McGill, J.L.; Estes, K.A. Effects of prenatal dietary rumen-protected choline supplementation during late gestation on calf growth, metabolism, and vaccine response. J. Dairy Sci. 2022, 105, 9639–9651. [Google Scholar] [CrossRef]
- Chen, J.; Yang, Z.; Dong, G. Niacin nutrition and rumen-protected niacin supplementation in dairy cows: An updated review. Br. J. Nutr. 2019, 122, 1103–1112. [Google Scholar] [CrossRef]
- Ijichi, H.; Ichiyama, A.; Hayaishi, O. Studies on the biosynthesis of nicotinamide adenine dinucleotide. 3. Comparative in vivo studies on nicotinic acid, nicotinamide, and quinolinic acid as precursors of nicotinamide adenine dinucleotide. J. Biol. Chem. 1966, 241, 3701–3707. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. New Insights for nicotinamide: Metabolic disease, autophagy, and mTOR. Front. Biosci. 2020, 25, 1925–1973. [Google Scholar] [CrossRef]
- Bogan, K.L.; Brenner, C. Nicotinic acid, nicotinamide, and nicotinamide riboside: A molecular evaluation of NAD+ precursor vitamins in human nutrition. Annu. Rev. Nutr. 2008, 28, 115–130. [Google Scholar] [CrossRef]
- Graff, E.C.; Fang, H.; Wanders, D.; Judd, R.L. The absence of adiponectin alters niacin’s effects on adipose tissue inflammation in mice. Nutrients 2020, 12, 2427. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, M.L.; Ganji, S.; Nakra, N.K.; Kamanna, V.S. Niacin for treatment of nonalcoholic fatty liver disease (NAFLD): Novel use for an old drug? J. Clin. Lipidol. 2019, 13, 873–879. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.C. From the Editor Pellagra, Osler, Roberts, Goldberger, the Atherosclerotic Diet, Niacin, the Beginning of the Atherosclerotic Epidemic, and the First Lipid-Altering Drug. Am. J. Cardiol. 2019, 123, 697–700. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, M.; Shimotsuma, M.; Hirose, T. Separation of nicotinamide metabolites using a PBr column packed with pentabromobenzyl group modified silica gel. Anal. Biochem. 2022, 655, 114837. [Google Scholar] [CrossRef]
- Hazim, R.A.; Paniagua, A.E.; Tang, L.; Yang, K.K.O.; Kim, K.; Stiles, L.; Divakaruni, A.S.; Williams, D. Vitamin B3, nicotinamide, enhances mitochondrial metabolism to promote differentiation of the retinal pigment epithelium. J. Biol. Chem. 2022, 298, 102286. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Chen, Y.; Xu, K.; Li, Y.; Riaz, F.; Lu, K.; Chen, Q.; Du, X.; Wu, L.; Cao, D.; et al. Cholesterol-induced leucine aminopeptidase 3 (LAP3) upregulation inhibits cell autophagy in pathogenesis of NAFLD. Aging 2022, 14, 3259–3275. [Google Scholar] [CrossRef]
- Caprarulo, V.; Erb, S.; Chandler, T.L.; Zenobi, M.G.; Barton, B.A.; Staples, C.R.; White, H.M. The effects of prepartum energy intake and peripartum rumen-protected choline supplementation on hepatic genes involved in glucose and lipid metabolism. J. Dairy Sci. 2020, 103, 11439–11448. [Google Scholar] [CrossRef]
- Zenobi, M.G.; Bollatti, J.M.; Lopez, A.M.; Barton, B.A.; Hixson, C.L.; Maunsell, F.P.; Thatcher, W.W.; Miller-Cushon, K.; Santos, J.E.P.; Staples, C.R.; et al. Effects of maternal choline supplementation on performance and immunity of progeny from birth to weaning. Clin. Chem. Lab. Med. 2022, 105, 9896–9916. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K. Nicotinamide as a Foundation for Treating Neurodegenerative Disease and Metabolic Disorders. Curr. Neurovasc. Res. 2021, 18, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Tselykh, T.; Mäkelä, J.; Ho, T.; Olkkonen, V.; Bornhauser, B.; Korhonen, L.; Zelcer, N.; Lindholm, D. Fibroblast growth factor-21 (FGF21) regulates low-density lipoprotein receptor (LDLR) levels in cells via the E3-ubiquitin ligase Mylip/Idol and the Canopy2 (Cnpy2)/Mylip-interacting saposin-like protein (Msap). J. Biol. Chem. 2012, 287, 12602–12611. [Google Scholar] [CrossRef] [PubMed]
- Bondurant, L.D.; Potthoff, M.J. Fibroblast growth factor 21: A versatile regulator of metabolic homeostasis. Annu. Rev. Nutr. 2018, 38, 173–196. [Google Scholar] [CrossRef]
- Badman, M.K.; Pissios, P.; Kennedy, A.R.; Koukos, G.; Flier, J.S.; Maratos-Flier, E. Hepatic fibroblast growth factor 21 is regulated by PPARalpha and is a key mediator of hepatic lipid metabolism in ketotic states. Cell Metab. 2007, 5, 426–437. [Google Scholar] [CrossRef]
- Badman, M.K.; Koester, A.; Flier, J.S.; Kharitonenkov, A.; Maratos-Flier, E. Fibroblast growth factor 21-deficient mice demonstrate impaired adaptation to ketosis. Endocrinology 2009, 150, 4931–4940. [Google Scholar] [CrossRef]
- Tillman, E.J.; Rolph, T. FGF21: An emerging therapeutic target for non-alcoholic steatohepatitis and related metabolic diseases. Front. Endocrinol. 2020, 11, 601290. [Google Scholar] [CrossRef]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Lipidol. 2005, 115, 1627–1635. [Google Scholar] [CrossRef]
- Maida, A.; Zota, A.; Sjøberg, K.A.; Schumacher, J.; Sijmonsma, T.P.; Pfenninger, A.; Christensen, M.M.; Gantert, T.; Fuhrmeister, J.; Rothermel, U.; et al. A liver stress-endocrine nexus promotes metabolic integrity during dietary protein dilution. J. Clin. Investig. 2016, 126, 3263–3678. [Google Scholar] [CrossRef]
- Hall, J.A.; Grainger, J.R.; Spencer, S.P.; Belkaid, Y.J.I. The role of retinoic acid in tolerance and immunity. Immunity 2011, 35, 13–22. [Google Scholar] [CrossRef]
- Ross, A.C.; Zolfaghari, R. Cytochrome P450s in the regulation of cellular retinoic acid metabolism. Annu. Rev. Nutr. 2011, 31, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Zhong, G.; Hogarth, C.; Snyder, J.M.; Palau, L.; Topping, T.; Huang, W.; Czuba, L.C.; Lafrance, J.; Ghiaur, G.; Isoherranen, N. The retinoic acid hydroxylase Cyp26a1 has minor effects on postnatal vitamin A homeostasis, but is required for exogenous atRA clearance. J. Biol. Chem. 2019, 294, 11166–11179. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Zhuang, L.; Maddox, D.M.; Smith, S.B.; Ganapathy, V. Structure, function, and expression pattern of a novel sodium-coupled citrate transporter (NaCT) cloned from mammalian brain. J. Biol. Chem. 2002, 277, 39469–39476. [Google Scholar] [CrossRef]
- Rogers, R.P.; Rogina, B. The role of INDY in metabolism, health and longevity. Front. Genet. 2015, 6, 204. [Google Scholar] [CrossRef] [PubMed]
- Brachs, S.; Winkel, A.F.; Tang, H.; Birkenfeld, A.; Brunner, B.; Jahn-Hofmann, K.; Margerie, D.; Ruetten, H.; Schmoll, D.; Spranger, J. Inhibition of citrate cotransporter Slc13a5/mINDY by RNAi improves hepatic insulin sensitivity and prevents diet-induced non-alcoholic fatty liver disease in mice. Mol. Metab. 2016, 5, 1072–1082. [Google Scholar] [CrossRef]
- Neuschäfer-Rube, F.; Lieske, S.; Kuna, M.; Henkel, J.; Perry, R.J.; Erion, D.; Pesta, D.M.; Willmes, D.; Brachs, S.; von Loeffelholz, C.; et al. The mammalian INDY homolog is induced by CREB in a rat model of type 2 diabetes. Diabetes 2014, 63, 1048–1057. [Google Scholar] [CrossRef]
- Li, Z.; Li, D.; Choi, E.; Lapidus, R.; Zhang, L.; Huang, S.; Shapiro, P.; Wang, H. Silencing of solute carrier family 13 member 5 disrupts energy homeostasis and inhibits proliferation of human hepatocarcinoma cells. J. Biol. Chem. 2017, 292, 13890–13901. [Google Scholar] [CrossRef]
- König, J.; Cui, Y.; Nies, A.T.; Keppler, D. A novel human organic anion transporting polypeptide localized to the basolateral hepatocyte membrane. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 278, G156–G164. [Google Scholar] [CrossRef]
- Svoboda, M.; Riha, J.; Wlcek, K.; Jaeger, W.; Thalhammer, T. Organic anion transporting polypeptides (OATPs): Regulation of expression and function. Curr. Drug Metab. 2011, 12, 139–153. [Google Scholar] [CrossRef]
- Huang, Y.; Du, Y.; Zheng, Y.; Wen, C.; Zou, H.; Huang, J.; Zhou, H.; Zhao, H.; Wu, L. Ct-OATP1B3 promotes high-grade serous ovarian cancer metastasis by regulation of fatty acid beta-oxidation and oxidative phosphorylation. Cell Death Dis. 2022, 13, 556. [Google Scholar] [CrossRef]
- Allen-Baume, V.; Ségui, B.; Cockcroft, S. Current thoughts on the phosphatidylinositol transfer protein family. FEBS Lett. 2002, 531, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, C.; Cockcroft, S. The role of phosphatidylinositol transfer proteins (PITPs) in intracellular signalling. Trends Endocrinol. Metab. 1998, 9, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Pietras, Ł.; Stefanik, E.; Rakus, D.; Gizak, A. FBP2—A New Player in Regulation of Motility of Mitochondria and Stability of Microtubules in Cardiomyocytes. Cells 2022, 11, 1710. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, H.C.; Dalby, P.A. The Two-Species Model of transketolase explains donor substrate-binding, inhibition and heat-activation. Sci. Rep. 2020, 10, 4148. [Google Scholar] [CrossRef] [PubMed]
- Mirando, A.C.; Francklyn, C.S.; Lounsbury, K.M. Regulation of angiogenesis by aminoacyl-tRNA synthetases. Int. J. Mol. Sci. 2014, 15, 23725–23748. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Fox, P. Aminoacyl-tRNA synthetases in medicine and disease. EMBO. Mol. Med. 2013, 5, 332–343. [Google Scholar] [CrossRef]
- Hyeon, D.Y.; Kim, J.H.; Ahn, T.J.; Cho, Y.; Hwang, D.; Kim, S. Evolution of the multi-tRNA synthetase complex and its role in cancer. J. Biol. Chem. 2019, 294, 5340–5351. [Google Scholar] [CrossRef]
- Lee, J.M.; Kim, T.; Kim, E.Y.; Kim, A.; Lee, D.K.; Kwon, N.H.; Kim, S.; Chang, Y.S. Methionyl-tRNA synthetase is a useful diagnostic marker for lymph node metastasis in non-small cell lung cancer. Yonsei Med. J. 2019, 60, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Lee, H.J.; Kong, J.; Cho, H.Y.; Kim, S.; Kang, B.S. Structural basis for the dynamics of human methionyl-tRNA synthetase in multi-tRNA synthetase complexes. Nucleic. Acids. Res. 2021, 49, 6549–6568. [Google Scholar] [CrossRef]
- Buckner, F.S.; Ranade, R.M.; Gillespie, J.R.; Shibata, S.; Hulverson, M.A.; Zhang, Z.; Huang, W.; Choi, R.; Verlinde, C.L.M.J.; Hol, W.; et al. Cryptosporidium optimization of methionyl tRNA-synthetase inhibitors for treatment of infection. Antimicrob. Agents Chemother. 2019, 63, e02061-18. [Google Scholar] [CrossRef]
- Sun, K.; Tordjman, J.; Clément, K.; Scherer, P. Fibrosis and adipose tissue dysfunction. Cell Met. 2013, 18, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T. Integrating mechanisms of pulmonary fibrosis. J. Exp. Med. 2011, 208, 1339–1350. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Pedroza, M.; Lafyatis, R.; George, A.T.; Mayes, M.D.; Assassi, S.; Tan, F.K.; Brenner, M.B.; Agarwal, S.K. Identification of cadherin 11 as a mediator of dermal fibrosis and possible role in systemic sclerosis. Arthritis Rheumatol. 2014, 66, 1010–1021. [Google Scholar] [CrossRef] [PubMed]
- Schneider, D.J.; Wu, M.; Le, T.; Cho, S.H.; Brenner, M.B.; Blackburn, M.R.; Agarwal, S.K. Cadherin-11 contributes to pulmonary fibrosis: Potential role in TGF-β production and epithelial to mesenchymal transition. FASEB J. 2012, 26, 503–512. [Google Scholar] [CrossRef]
- Garcia, M.; Mamedova, L.K.; Barton, B.; Bradford, B.J. In vitro choline regulates the function of bovine immune cells and alters the mRNA abundance of enzymes and receptors involved in its metabolism. Front. Immunol. 2018, 9, 2448. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Forward (5′-3′) | Reverse (5′-3′) |
|---|---|---|
| FBP2 | GTCACGTTAACGCTTCCTGC | CTGCACTGCTGGCGTTTTAG |
| SLCO1B3 | CACACTTGGGTGAATGCCCA | ATGTGGTACCTCCTGTTGCAG |
| MARS1 | GGGCTTCCAGCTGATGCTAT | TGGACTCTCTGTAGCCACCA |
| FGF21 | ATCGCTCCACTTTGACCCTG | GGGCTTCGGACTGGTAAACA |
| CYP26A1 | CCCTATGCTGTGGCTGCAAT | CCAAGGGCTGACAAACTGGA |
| CDH11 | GGGCTTCCAGCTGATGCTAT | TGGACTCTCTGTAGCCACCA |
| SLC13A5 | TGATGACGTAGGCACACCTG | TTGACAATCCATGCCAGCCG |
| β-actin | GCACTCTTCCAGCCTTCCTT | AATGCCAGGGTACATGGTGG |
| Reagent Name | Volume (μL) |
|---|---|
| 2× SYBR® Green Pro Taq HS Premix | 5.0 |
| Forward primer (10 μM) | 0.5 |
| Reverse primer (10 μM) | 0.5 |
| cDNA | 4.0 |
| Reaction conditions: 95 °C 30 s; 2 95 °C 5 s, 60 °C 30 s (40 cycles) (two-step algorithm) Dissociation stage: 95 °C 10 s, 65 °C 60 s, 97 °C 1 s | |
| Gene Name | Gene Description | Log2FC (RPC/RPM) * | p-Value |
|---|---|---|---|
| FARS2 | phenylalanyl-tRNA synthetase 2, mitochondrial | 1.757 | 1.31 × 1013 |
| FGF21 | fibroblast growth factor 21 | −4.535 | 5.77 × 1013 |
| NOTUM | notum, palmitoleoyl-protein carboxylesterase | 2.310 | 1.13 × 1012 |
| CYP2C19 | cytochrome P450, family 2, subfamily C, polypeptide 19 | 2.894 | 2.58 × 1012 |
| KCNN2 | potassium calcium-activated channel subfamily N member 2 | 2.163 | 5.49 × 1012 |
| FUT1 | fucosyltransferase 1 | −5.524 | 2.32 × 1012 |
| CYP26A1 | cytochrome P450, family 26, subfamily A, polypeptide 1 | 3.315 | 7.46 × 1011 |
| CDH11 | cadherin 11 | −3.201 | 2.64 × 1010 |
| DTX1 | deltex E3 ubiquitin ligase 1 | −2.872 | 4.29 × 1010 |
| ATP6V1C2 | ATPase H+ transporting V1 subunit C2 | −4.079 | 5.57 × 1010 |
| SLC13A5 | solute carrier family 13 member 5 | −2.369 | 1.53 × 109 |
| LRRC73 | leucine-rich repeat-containing 73 | −3.907 | 4.41 × 109 |
| HOPX | HOP homeobox | −3.070 | 4.68 × 109 |
| SLCO1B3 | solute carrier organic anion transporter family member 1B3 | 1.089 | 5.55 × 109 |
| MICAL2 | microtubule-associated monooxygenase, calponin and LIM domain-containing 2 | −2.959 | 1.36 × 108 |
| GPC3 | glypican 3 | -1.830 | 3.69 × 108 |
| ADCY2 | adenylate cyclase 2 | −3.537 | 7.04 × 107 |
| HRG | histidine-rich glycoprotein | 1.666 | 1.02 × 107 |
| GLI1 | GLI family zinc finger 1 | −3.446 | 1.10 × 107 |
| CES1 | carboxylesterase 1 (monocyte/macrophage serine esterase 1) | 2.000 | 2.08 × 107 |
| CD1D | CD1D antigen, d polypeptide | 1.810 | 4.24 × 107 |
| FBP2 | fructose-bisphosphatase 2 | 3.167 | 4.36 × 107 |
| MARS1 | methionyl-tRNA synthetase 1 | −2.070 | 4.71 × 107 |
| SEC14L3 | SEC14-like lipid binding 3 | 4.21 | 5.31 × 107 |
| INHBE | inhibin subunit beta E | −2.277 | 8.08 × 107 |
| CARS1 | cysteinyl-tRNA synthetase 1 | −1.218 | 1.71 × 106 |
| PLEK | pleckstrin | 1.545 | 1.72 × 106 |
| TKT | transketolase | 1.009 | 1.75 × 106 |
| CITED4 | Cbp/p300-interacting transactivator with Glu/Asp-rich carboxy-terminal domain 4 | −2.026 | 2.22 × 106 |
| BIRC5 | baculoviral IAP-repeat-containing 5 | 2.703 | 2.34 × 106 |
| GO ID | Category | Description | p-Value | Counts |
|---|---|---|---|---|
| GO:0006629 | BP | lipid metabolic process | 1.18 × 106 | 29 |
| GO:0044281 | BP | small molecule metabolic process | 1.56 × 106 | 40 |
| GO:0034754 | BP | cellular hormone metabolic process | 3.39 × 106 | 8 |
| GO:0006805 | BP | xenobiotic metabolic process | 4.33 × 106 | 7 |
| GO:0005615 | CC | extracellular space | 2.60 × 106 | 33 |
| GO:0004497 | MF | monooxygenase activity | 3.61 × 106 | 10 |
| GO:0005737 | CC | cytoplasm | 5.73 × 106 | 88 |
| GO:0006082 | BP | organic acid metabolic process | 7.52 × 106 | 23 |
| GO:0043436 | BP | oxoacid metabolic process | 1.41 × 105 | 22 |
| GO:0042445 | BP | hormone metabolic process | 1.64 × 105 | 9 |
| GO:0019752 | BP | carboxylic acid metabolic process | 1.78 × 105 | 21 |
| GO:0110165 | CC | cellular anatomical entity | 2.00 × 105 | 236 |
| GO:0032787 | BP | monocarboxylic acid metabolic process | 3.31 × 105 | 15 |
| GO:0016712 | MF | oxidoreductase activity, acting on paired donors, with incorporation or reduction in molecular oxygen, reduced flavin or flavoprotein as one donor, and incorporation of one atom of oxygen | 2.96 × 105 | 5 |
| GO:0046906 | MF | tetrapyrrole binding | 3.25 × 105 | 10 |
| GO:0008009 | MF | chemokine activity | 4.86 × 105 | 5 |
| GO:0008202 | BP | steroid metabolic process | 8.63 × 105 | 9 |
| GO:0098754 | BP | detoxification | 9.17 × 105 | 7 |
| GO:0102033 | MF | fatty acid omega-hydroxylase activity | 9.08 × 105 | 2 |
| GO:0008395 | MF | steroid hydroxylase activity | 9.33 × 105 | 5 |
| Pathway ID | KEGG Pathways | p-Value | Counts |
|---|---|---|---|
| map00830 | Retinol metabolism | 5.95 × 108 | 9 |
| map05204 | Chemical carcinogenesis - DNA adducts | 3.54 × 108 | 9 |
| map00980 | Metabolism of xenobiotics by cytochrome P450 | 9.19 × 106 | 7 |
| map00140 | Steroid hormone biosynthesis | 3.57 × 105 | 7 |
| map05207 | Chemical carcinogenesis - receptor activation | 3.44 × 105 | 11 |
| map00983 | Drug metabolism - other enzymes | 3.07 × 105 | 7 |
| map00240 | Pyrimidine metabolism | 5.39 × 105 | 6 |
| map04976 | Bile secretion | 0.00018 | 7 |
| map00982 | Drug metabolism - cytochrome P450 | 0.00058 | 5 |
| map04061 | Viral protein interaction with cytokine and cytokine receptor | 0.00076 | 6 |
| map00591 | Linoleic acid metabolism | 0.00165 | 4 |
| map00590 | Arachidonic acid metabolism | 0.00563 | 5 |
| map04062 | Chemokine signaling pathway | 0.01122 | 7 |
| map00970 | Aminoacyl-tRNA biosynthesis | 0.01591 | 3 |
| map04212 | Longevity regulating pathway - worm | 0.02022 | 5 |
| map04060 | Cytokine-cytokine receptor interaction | 0.02835 | 8 |
| map04750 | Inflammatory mediator regulation of TRP channels | 0.03684 | 4 |
| map00030 | Pentose phosphate pathway | 0.03520 | 2 |
| map05208 | Chemical carcinogenesis - reactive oxygen species | 0.02709 | 8 |
| map00230 | Purine metabolism | 0.03055 | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.; Cui, Z.; Zhang, R.; Zhao, K.; Wang, L.; Yao, J.; Liu, S.; Cai, C.; Cao, Y. The Effects of Rumen-Protected Choline and Rumen-Protected Nicotinamide on Liver Transcriptomics in Periparturient Dairy Cows. Metabolites 2023, 13, 594. https://doi.org/10.3390/metabo13050594
Du X, Cui Z, Zhang R, Zhao K, Wang L, Yao J, Liu S, Cai C, Cao Y. The Effects of Rumen-Protected Choline and Rumen-Protected Nicotinamide on Liver Transcriptomics in Periparturient Dairy Cows. Metabolites. 2023; 13(5):594. https://doi.org/10.3390/metabo13050594
Chicago/Turabian StyleDu, Xue’er, Zhijie Cui, Rui Zhang, Keliang Zhao, Lamei Wang, Junhu Yao, Shimin Liu, Chuanjiang Cai, and Yangchun Cao. 2023. "The Effects of Rumen-Protected Choline and Rumen-Protected Nicotinamide on Liver Transcriptomics in Periparturient Dairy Cows" Metabolites 13, no. 5: 594. https://doi.org/10.3390/metabo13050594