
Urinary Metabolomics for the Prediction of Radiation-Induced Cardiac Dysfunction
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
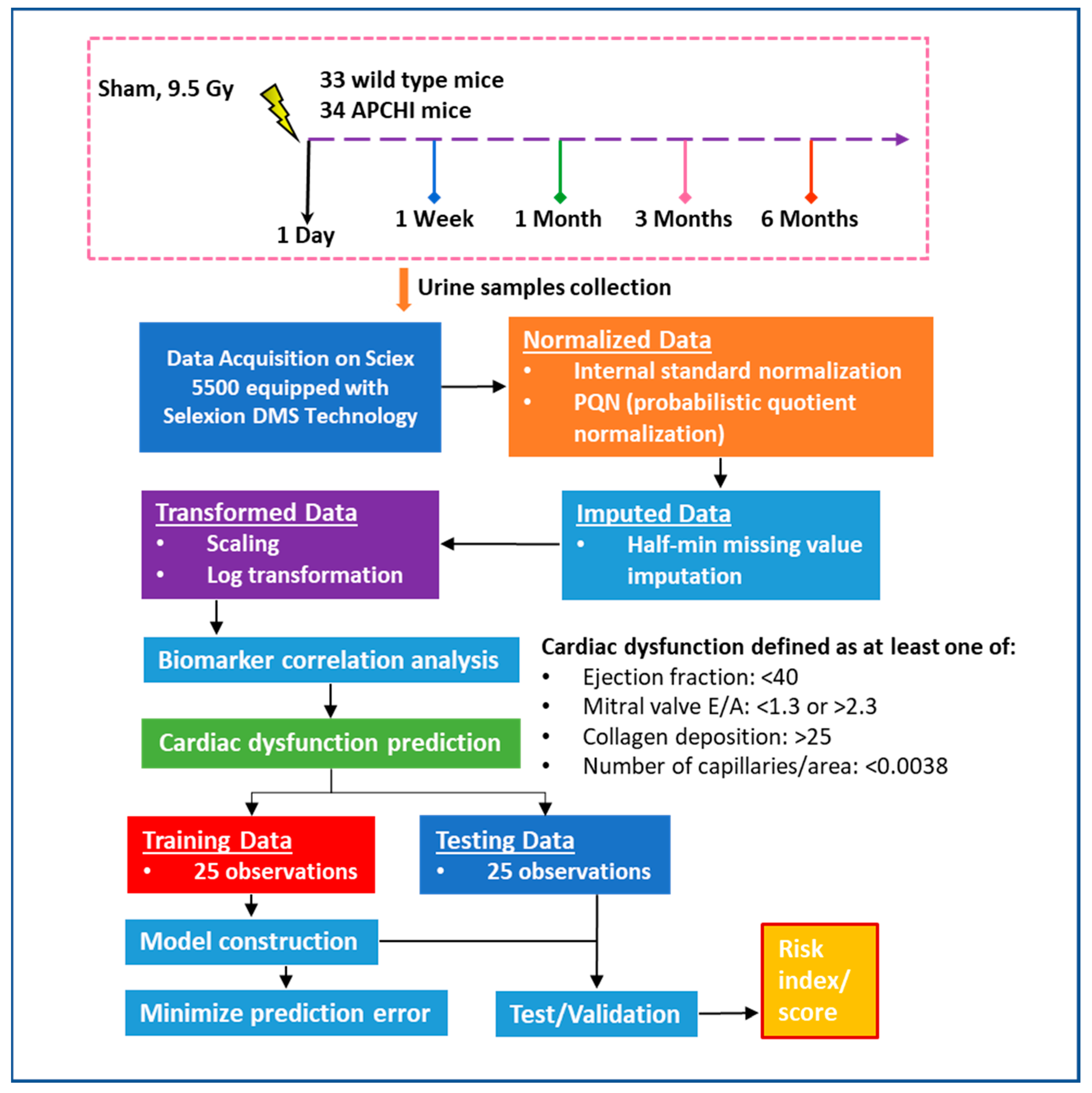
2.1. Animal Procedures
2.2. Chemicals
2.3. Targeted Metabolomics Analysis
2.4. Targeted Lipidomics Analysis
2.5. Data Processing and Statistical Analysis
3. Results
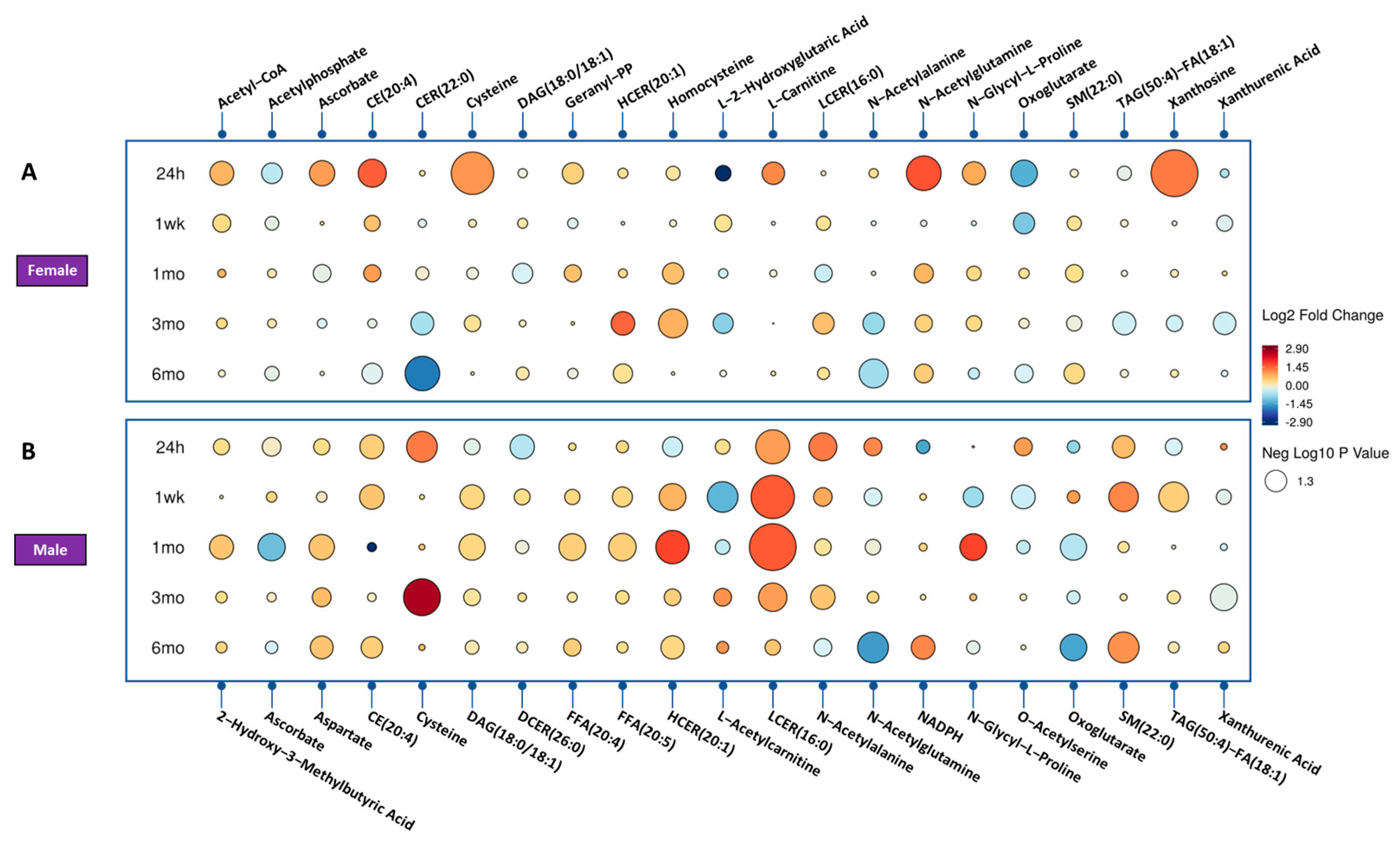
3.1. Radiation Induces a Robust Metabolic Response in the Urine Samples of Mice in a Time-Dependent Manner
3.2. Genotype Impacts the Response to Radiation Injury
3.3. Sex Impacts the Response to Radiation Injury
3.4. Prediction Model for Radiation-Induced Cardiac Dysfunction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Micewicz, E.D.; Iwamoto, K.S.; Ratikan, J.A.; Nguyen, C.; Xie, M.W.; Cheng, G.; Boxx, G.M.; Deriu, E.; Damoiseaux, R.D.; Whitelegge, J.P.; et al. The Aftermath of Surviving Acute Radiation Hematopoietic Syndrome and its Mitigation. Radiat. Res. 2019, 191, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Orschell, C.M.; Wu, T.; Patterson, A.M. Impact of Age, Sex, and Genetic Diversity in Murine Models of the Hematopoietic Acute Radiation Syndrome (H-ARS) and the Delayed Effects of Acute Radiation Exposure (DEARE). Curr. Stem Cell Rep. 2022, 8, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Orschell, C.M. The Delayed Effects of Acute Radiation Exposure (DEARE): Characteristics, mechanisms, animal models, and promising medical countermeasures. Int. J. Radiat. Biol. 2023, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cerveny, T.J.; MacVittie, T.M.; Young, R.W. Medical Consequences of Nuclear Warfare. In Textbook of Military Medicine; Cerveny, T.J., Walker, R.I., Eds.; TMM Publications, Office of the Surgeon General: Washington, DC, USA, 1989; Volume 2, pp. 15–36. [Google Scholar]
- Walden, T.L. Medical Consequences of Nuclear Warfare; Cerveny, T.J., Walker, R.I., Eds.; Office of Surgeon General, Department of the Army, TMM Publications: Washington, DC, USA, 1989; Chapter 9; pp. 171–226. [Google Scholar]
- Military Radiobiology; Conklin, J.J.; Walker, R.J. (Eds.) Academic Press Inc.: Cambridge, MA, USA, 1987; Chapter 20; pp. 379–391. [Google Scholar]
- Seed, T.M.; Kaspar, L.V.; Fritz, T.E.; Tolle, D.V. Cellular Responses in Chronic Radiation Leukemogenesis. In Carcinogenesis; Raven Press: New York, NY, USA, 1985; Volume 10, pp. 363–379. [Google Scholar]
- Young, R.W. Acute Radiation Syndrome. In Military Radiobiology; Conklin, J.J., Walker, R.I., Eds.; Academic Press: Cambridge, MA, USA, 1987; pp. 165–190. [Google Scholar]
- Satyamitra, M.M.; DiCarlo, A.L.; Hollingsworth, B.A.; Winters, T.A.; Taliaferro, L.P. Development of Biomarkers for Radiation Biodosimetry and Medical Countermeasures Research: Current Status, Utility, and Regulatory Pathways. Radiat. Res. 2022, 197, 514–532. [Google Scholar] [CrossRef] [PubMed]
- Satyamitra, M.M.; Cassatt, D.R.; Hollingsworth, B.A.; Price, P.W.; Rios, C.I.; Taliaferro, L.P.; Winters, T.A.; DiCarlo, A.L. Metabolomics in Radiation Biodosimetry: Current Approaches and Advances. Metabolites 2020, 10, 328. [Google Scholar] [CrossRef]
- Griffin, J.H.; Zlokovic, B.V.; Mosnier, L.O. Activated Protein C, Protease Activated Receptor 1, and Neuroprotection. Blood 2018, 132, 159–169. [Google Scholar] [CrossRef]
- Geiger, H.; Pawar, S.A.; Kerschen, E.J.; Nattamai, K.J.; Hernandez, I.; Liang, H.P.; Fernandez, J.A.; Cancelas, J.A.; Ryan, M.A.; Kustikova, O.; et al. Pharmacological targeting of the thrombomodulin-activated protein C pathway mitigates radiation toxicity. Nat. Med. 2012, 18, 1123–1129. [Google Scholar] [CrossRef]
- Isermann, B.; Vinnikov, I.A.; Madhusudhan, T.; Herzog, S.; Kashif, M.; Blautzik, J.; Corat, M.A.; Zeier, M.; Blessing, E.; Oh, J.; et al. Activated protein C protects against diabetic nephropathy by inhibiting endothelial and podocyte apoptosis. Nat. Med. 2007, 13, 1349–1358. [Google Scholar] [CrossRef]
- Van Mens, T.E.; Liang, H.H.; Basu, S.; Hernandez, I.; Zogg, M.; May, J.; Zhan, M.; Yang, Q.; Foeckler, J.; Kalloway, S.; et al. Variable phenotypic penetrance of thrombosis in adult mice after tissue-selective and temporally controlled Thbd gene inactivation. Blood Adv. 2017, 1, 1148–1158. [Google Scholar] [CrossRef]
- Sridharan, V.; Johnson, K.A.; Landes, R.D.; Cao, M.; Singh, P.; Wagoner, G.; Hayar, A.; Sprick, E.D.; Eveld, K.A.; Bhattacharyya, A.; et al. Sex-dependent effects of genetic upregulation of activated protein C on delayed effects of acute radiation exposure in the mouse heart, small intestine, and skin. PLoS ONE 2021, 16, e0252142. [Google Scholar] [CrossRef]
- Cheema, A.K.; Hinzman, C.P.; Mehta, K.Y.; Hanlon, B.K.; Garcia, M.; Fatanmi, O.O.; Singh, V.K. Plasma Derived Exosomal Biomarkers of Exposure to Ionizing Radiation in Nonhuman Primates. Int. J. Mol. Sci. 2018, 19, 3427. [Google Scholar] [CrossRef]
- Cheema, A.K.; Mehta, K.Y.; Fatanmi, O.O.; Wise, S.Y.; Hinzman, C.P.; Wolff, J.; Singh, V.K. A Metabolomic and Lipidomic Serum Signature from Nonhuman Primates Administered with a Promising Radiation Countermeasure, Gamma-Tocotrienol. Int. J. Mol. Sci. 2017, 19, 79. [Google Scholar] [CrossRef]
- Upadhyay, M.; Rajagopal, M.; Gill, K.; Li, Y.; Bansal, S.; Sridharan, V.; Tyburski, J.B.; Boerma, M.; Cheema, A.K. Identification of Plasma Lipidome Changes Associated with Low Dose Space-Type Radiation Exposure in a Murine Model. Metabolites 2020, 10, 252. [Google Scholar] [CrossRef]
- Cheema, A.K.; Li, Y.; Girgis, M.; Jayatilake, M.; Simas, M.; Wise, S.Y.; Olabisi, A.O.; Seed, T.M.; Singh, V.K. Metabolomic studies in tissues of mice treated with amifostine and exposed to gamma-radiation. Sci. Rep. 2019, 9, 15701. [Google Scholar] [CrossRef]
- Sündermann, A.; Eggers, L.F.; Schwudke, D. Liquid Extraction: Bligh and Dyer. In Encyclopedia of Lipidomics; Wenk, M.R., Ed.; Springer: Dordrecht, The Netherlands, 2016; pp. 1–4. [Google Scholar]
- Viswanathan, C.T.; Bansal, S.; Booth, B.; DeStefano, A.J.; Rose, M.J.; Sailstad, J.; Shah, V.P.; Skelly, J.P.; Swann, P.G.; Weiner, R. Quantitative bioanalytical methods validation and implementation: Best practices for chromatographic and ligand binding assays. Pharm. Res. 2007, 24, 1962–1973. [Google Scholar] [CrossRef]
- Johansson, B.; Lundin, F.; Tegerback, R.; Bojo, L. E/a ratio a simple detector of left ventricular dysfunction in patients with decreased ejection fraction. Scand. Cardiovasc. J. 2018, 52, 20–27. [Google Scholar] [CrossRef]
- Datta, K.; Suman, S.; Kallakury, B.V.; Fornace, A.J., Jr. Exposure to heavy ion radiation induces persistent oxidative stress in mouse intestine. PLoS ONE 2012, 7, e42224. [Google Scholar] [CrossRef]
- Suman, S.; Johnson, M.D.; Fornace, A.J., Jr.; Datta, K. Exposure to ionizing radiation causes long-term increase in serum estradiol and activation of PI3K-Akt signaling pathway in mouse mammary gland. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, 500–507. [Google Scholar] [CrossRef]
- Rola, R.; Sarkissian, V.; Obenaus, A.; Nelson, G.A.; Otsuka, S.; Limoli, C.L.; Fike, J.R. High-LET radiation induces inflammation and persistent changes in markers of hippocampal neurogenesis. Radiat. Res. 2005, 164, 556–560. [Google Scholar] [CrossRef]
- Tseng, B.P.; Giedzinski, E.; Izadi, A.; Suarez, T.; Lan, M.L.; Tran, K.K.; Acharya, M.M.; Nelson, G.A.; Raber, J.; Parihar, V.K.; et al. Functional consequences of radiation-induced oxidative stress in cultured neural stem cells and the brain exposed to charged particle irradiation. Antioxid. Redox Signal. 2014, 20, 1410–1422. [Google Scholar] [CrossRef]
- Snyder, A.R.; Morgan, W.F. Gene expression profiling after irradiation: Clues to understanding acute and persistent responses? Cancer Metastasis Rev. 2004, 23, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Azzam, E.I.; Jay-Gerin, J.P.; Pain, D. Ionizing radiation-induced metabolic oxidative stress and prolonged cell injury. Cancer Lett. 2012, 327, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Coeytaux, K.; Bey, E.; Christensen, D.; Glassman, E.S.; Murdock, B.; Doucet, C. Reported radiation overexposure accidents worldwide, 1980-2013: A systematic review. PLoS ONE 2015, 10, e0118709. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, A.; Tanigawa, K.; Ohtsuru, A.; Yabe, H.; Maeda, M.; Shigemura, J.; Ohira, T.; Tominaga, T.; Akashi, M.; Hirohashi, N.; et al. Health effects of radiation and other health problems in the aftermath of nuclear accidents, with an emphasis on Fukushima. Lancet 2015, 386, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Reyes, E.H.; Baciu, F.; Benderitter, M.; Lataillade, J.J.; Bey, E.; Trompier, F.; Tamarat, R. Medical Response to Radiological Accidents in Latin America and International Assistance. Radiat. Res. 2016, 185, 359–365. [Google Scholar] [CrossRef]
- Holm, A.; Aabenhus, R. Urine sampling techniques in symptomatic primary-care patients: A diagnostic accuracy review. BMC Fam. Pract. 2016, 17, 72. [Google Scholar] [CrossRef]
- Katsimbri, P.; Korakas, E.; Kountouri, A.; Ikonomidis, I.; Tsougos, E.; Vlachos, D.; Papadavid, E.; Raptis, A.; Lambadiari, V. The Effect of Antioxidant and Anti-Inflammatory Capacity of Diet on Psoriasis and Psoriatic Arthritis Phenotype: Nutrition as Therapeutic Tool? Antioxidants 2021, 10, 157. [Google Scholar] [CrossRef]
- Ping, Z.; Peng, Y.; Lang, H.; Xinyong, C.; Zhiyi, Z.; Xiaocheng, W.; Hong, Z.; Liang, S. Oxidative Stress in Radiation-Induced Cardiotoxicity. Oxidative Med. Cell. Longev. 2020, 2020, 3579143. [Google Scholar] [CrossRef]
- Wang, H.; Wei, J.; Zheng, Q.; Meng, L.; Xin, Y.; Yin, X.; Jiang, X. Radiation-induced heart disease: A review of classification, mechanism and prevention. Int. J. Mol. Sci. 2019, 15, 2128–2138. [Google Scholar] [CrossRef]
- Dynlacht, J.R.; Valluri, S.; Lopez, J.; Greer, F.; Desrosiers, C.; Caperell-Grant, A.; Mendonca, M.S.; Bigsby, R.M. Estrogen protects against radiation-induced cataractogenesis. Radiat. Res. 2008, 170, 758–764. [Google Scholar] [CrossRef]
- Prithivirajsingh, S.; Story, M.D.; Bergh, S.A.; Geara, F.B.; Ang, K.K.; Ismail, S.M.; Stevens, C.W.; Buchholz, T.A.; Brock, W.A. Accumulation of the common mitochondrial DNA deletion induced by ionizing radiation. FEBS Lett. 2004, 571, 227–232. [Google Scholar] [CrossRef]
- Eaton, S.; Bartlett, K.; Pourfarzam, M. Mammalian mitochondrial beta-oxidation. Biochem. J. 1996, 320 Pt 2, 345–357. [Google Scholar] [CrossRef]
- Rutkowsky, J.M.; Knotts, T.A.; Ono-Moore, K.D.; McCoin, C.S.; Huang, S.; Schneider, D.; Singh, S.; Adams, S.H.; Hwang, D.H. Acylcarnitines activate proinflammatory signaling pathways. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E1378–E1387. [Google Scholar] [CrossRef]
- Gomez, L.A.; Heath, S.H.; Hagen, T.M. Acetyl-L-carnitine supplementation reverses the age-related decline in carnitine palmitoyltransferase 1 (CPT1) activity in interfibrillar mitochondria without changing the L-carnitine content in the rat heart. Mech. Ageing Dev. 2012, 133, 99–106. [Google Scholar] [CrossRef]
- Bernard, A.; Rigault, C.; Mazue, F.; Le Borgne, F.; Demarquoy, J. L-carnitine supplementation and physical exercise restore age-associated decline in some mitochondrial functions in the rat. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2008, 63, 1027–1033. [Google Scholar] [CrossRef]
- Yuan, D.; Chu, J.; Lin, H.; Zhu, G.; Qian, J.; Yu, Y.; Yao, T.; Ping, F.; Chen, F.; Liu, X. Mechanism of homocysteine-mediated endothelial injury and its consequences for atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 1109445. [Google Scholar] [CrossRef]
- Herrmann, W.; Herrmann, M. The Controversial Role of HCY and Vitamin B Deficiency in Cardiovascular Diseases. Nutrients 2022, 14, 1412. [Google Scholar] [CrossRef]
- Kaplan, P.; Tatarkova, Z.; Sivonova, M.K.; Racay, P.; Lehotsky, J. Homocysteine and Mitochondria in Cardiovascular and Cerebrovascular Systems. Int. J. Mol. Sci. 2020, 21, 7698. [Google Scholar] [CrossRef]
- Carley, A.N.; Lewandowski, E.D. Triacylglycerol turnover in the failing heart. Biochim. Biophys. Acta 2016, 1861, 1492–1499. [Google Scholar] [CrossRef]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef]
- Truscott, R.J.W.; Malegan, D.; McCairns, E.; Burke, D.; Hick, L.; Sims, P.; Halpern, B.; Tanaka, K.; Sweetman, L.; Nyhan, W.L.; et al. New metabolites in isovaleric acidemia. Clin. Chim. Acta 1981, 110, 187–203. [Google Scholar] [CrossRef] [PubMed]
- De Fabiani, E. The true story of palmitoleic acid: Between myth and reality. Eur. J. Lipid Sci. Technol. 2011, 113, 809–811. [Google Scholar] [CrossRef]
- Abraham, R.; Riemersma, R.A.; Wood, D.; Elton, R.; Oliver, M.F. Adipose fatty acid composition and the risk of serious ventricular arrhythmias in acute myocardial infarction. Am. J. Cardiol. 1989, 63, 269–272. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Week Regression Model | ||||
|---|---|---|---|---|
| Name | p-Value | FDR | Fold Change | Log2(FC) |
| FFA(16:1) | 0.00031 | 0.067 | 0.493 | −1.020 |
| Homocysteine | 0.00083 | 0.081 | 0.375 | −1.415 |
| TAG56:7-FA20:4 | 0.00114 | 0.081 | 1.883 | 0.913 |
| SM(24:0) | 0.00056 | 0.079 | 2.385 | 1.254 |
| SM(22:0) | 0.00003 | 0.011 | 2.314 | 1.211 |
| 1 Month Regression Model | ||||
| Name | p-Value | FDR | Fold Change | Log2(FC) |
| TAG54:4-FA16:0 | 0.00077 | 0.327 | 2.094 | 1.067 |
| Spermidine | 0.00709 | 0.511 | 0.670 | −0.578 |
| CE(22:6) | 0.00929 | 0.511 | 2.981 | 1.576 |
| Anthranilic acid | 0.03527 | 0.511 | 0.660 | −0.601 |
| Mesaconic acid | 0.22154 | 0.595 | 1.392 | 0.477 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Bansal, S.; Sridharan, V.; Bansal, S.; Jayatilake, M.M.; Fernández, J.A.; Griffin, J.H.; Boerma, M.; Cheema, A.K. Urinary Metabolomics for the Prediction of Radiation-Induced Cardiac Dysfunction. Metabolites 2023, 13, 525. https://doi.org/10.3390/metabo13040525
Li Y, Bansal S, Sridharan V, Bansal S, Jayatilake MM, Fernández JA, Griffin JH, Boerma M, Cheema AK. Urinary Metabolomics for the Prediction of Radiation-Induced Cardiac Dysfunction. Metabolites. 2023; 13(4):525. https://doi.org/10.3390/metabo13040525
Chicago/Turabian StyleLi, Yaoxiang, Shivani Bansal, Vijayalakshmi Sridharan, Sunil Bansal, Meth M. Jayatilake, Jose A. Fernández, John H. Griffin, Marjan Boerma, and Amrita K. Cheema. 2023. "Urinary Metabolomics for the Prediction of Radiation-Induced Cardiac Dysfunction" Metabolites 13, no. 4: 525. https://doi.org/10.3390/metabo13040525