
Oral Sources of Salivary Metabolites

Abstract
:1. Introduction
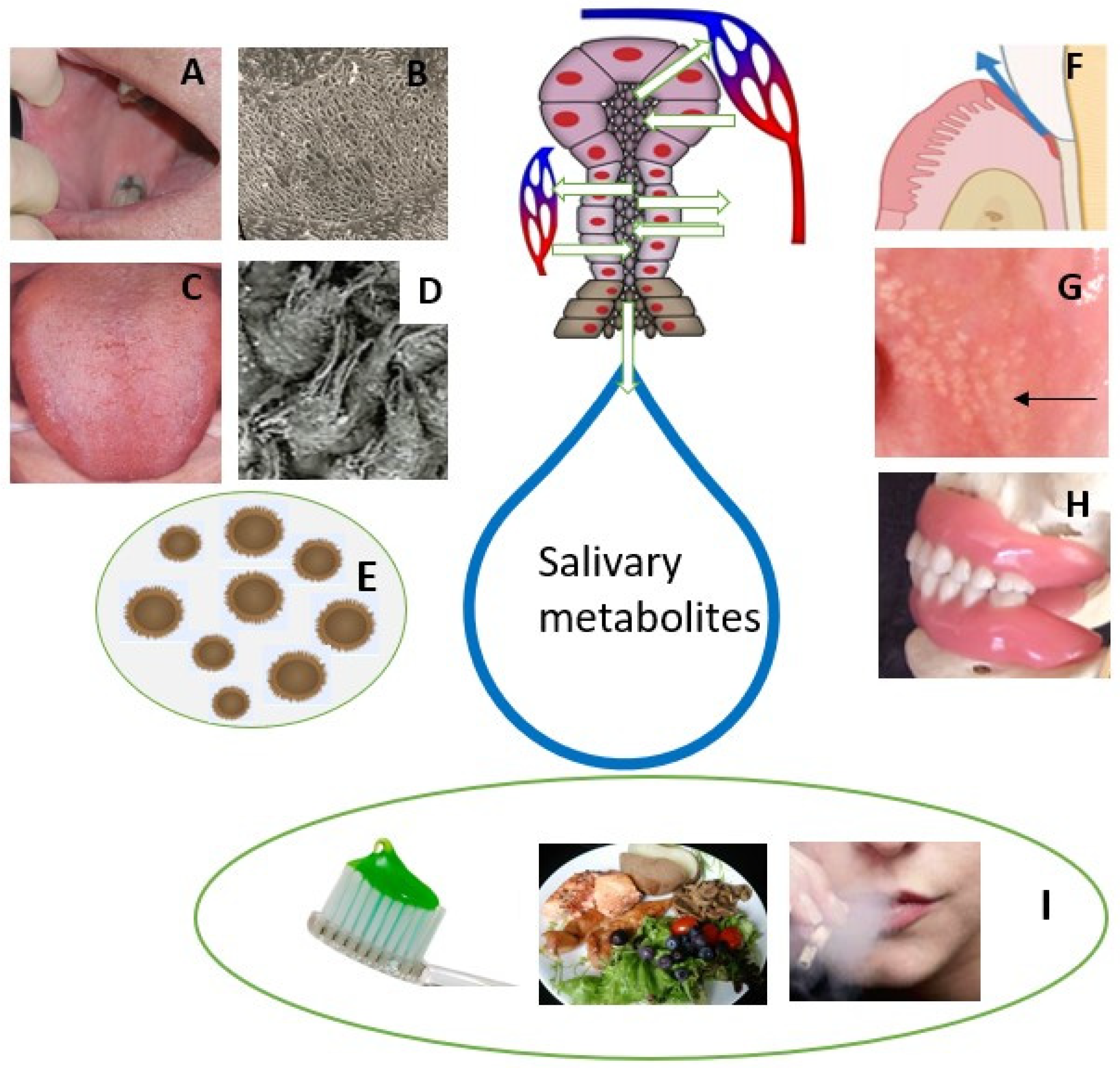
2. Sources of Salivary Metabolites in Healthy Subjects
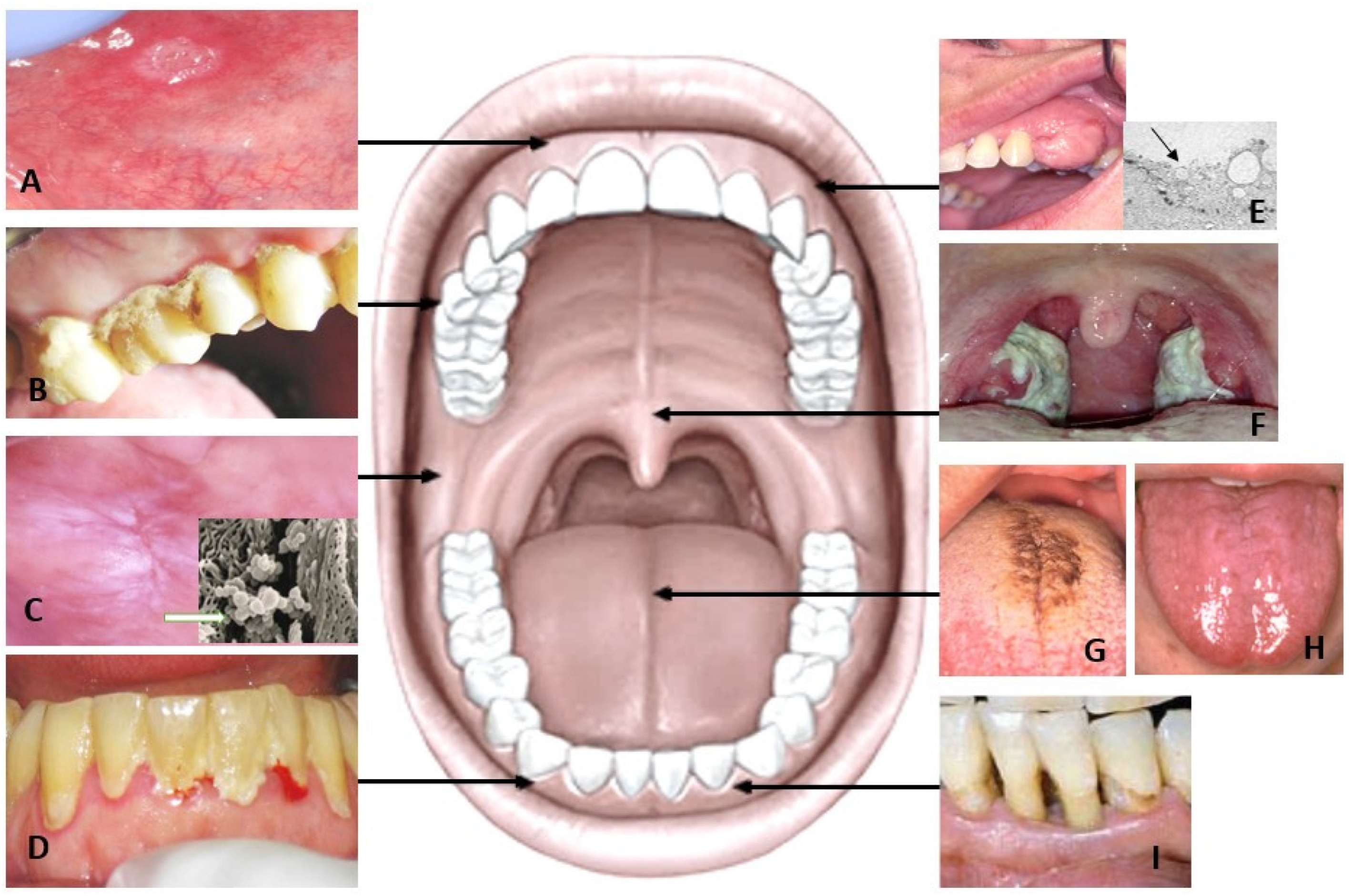
3. Salivary Metabolites in Patients with Oral Inflammation
4. Salivary Metabolomics in Oral Mucosal Diseases and Oral Cancer
4.1. Tongue Disorders
4.2. Leukoplakia and Lichen Planus
4.3. Oral Cancer
5. Discussion
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mikkonen, J.J.; Singh, S.P.; Herrala, M.; Lappalainen, R.; Myllymaa, S.; Kullaa, A.M. Salivary metabolomics in the diagnosis of oral cancer and periodontal diseases. J. Periodontal Res. 2016, 51, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Mikkonen, J.J.; Raittila, J.; Rieppo, L.; Lappalainen, R.; Kullaa, A.M.; Myllymaa, S. Fourier Transform Infrared Spectroscopy and Photoacoustic Spectroscopy for Saliva Analysis. Appl. Spectrosc. 2016, 70, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Hyvärinen, E.; Savolainen, M.; Mikkonen, J.J.W.; Kullaa, A.M. Salivary Metabolomics for Diagnosis and Monitoring Diseases: Challenges and Possibilities. Metabolites 2021, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J. Systems Biology of Metabolism. Annu. Rev. Biochem. 2017, 86, 245–275. [Google Scholar] [CrossRef]
- Gardner, A.; Parkes, H.G.; Carpenter, G.H.; So, P.W. Developing and standardizing a protocol for quantitative proton nuclear magnetic resonance (1H NMR) spectroscopy of saliva. J. Proteome Res. 2018, 17, 1521–1531. [Google Scholar] [CrossRef]
- Gardner, A.; Parkes, H.G.; So, P.W.; Carpenter, G.H. Determining bacterial and host contributions to the human salivary metabolome. J. Oral Microbiol. 2019, 11, 1617014. [Google Scholar] [CrossRef]
- Sampaio-Maia, B.; Caldas, I.M.; Pereira, M.L.; Pérez-Mongiovi, D.; Araujo, R. The Oral Microbiome in Health and Its Implication in Oral and Systemic Diseases. Adv. Appl. Microbiol. 2016, 97, 171–210. [Google Scholar]
- Mosaddad, S.A.; Tahmasebi, E.; Yazdanian, A.; Rezvani, M.B.; Seifalian, A.; Yazdanian, M.; Tebyanian, H. Oral microbial biofilms: An update. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 2005–2019. [Google Scholar] [CrossRef]
- Fábián, T.K.; Hermann, P.; Beck, A.; Fejérdy, P.; Fábián, G. Salivary defense proteins: Their network and role in innate and acquired oral immunity. Int. J. Mol. Sci. 2012, 13, 4295–4320. [Google Scholar] [CrossRef]
- Barros, S.P.; Williams, R.; Offenbacher, S.; Morelli, T. Gingival crevicular fluid as a source of biomarkers for periodontitis. Periodontol 2000 2016, 70, 53–64. [Google Scholar] [CrossRef]
- O’Donnell, L.E.; Smith, K.; Williams, C.; Nile, C.J.; Lappin, D.F.; Bradshaw, D.; Lambert, M.; Robertson, D.P.; Bagg, J.; Hannah, V.; et al. Dentures are a Reservoir for Respiratory Pathogens. J. Prosthodont. 2016, 25, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Tuganbaev, T.; Yoshida, K.; Honda, K. The effects of oral microbiota on health. Science 2022, 376, 934–936. [Google Scholar] [CrossRef] [PubMed]
- Aimetti, M.; Cacciatore, S.; Graziano, A.; Tenori, L. Metabonomic analysis of saliva reveals generalized chronic periodontitis signature. Metabolomics 2012, 8, 465–474. [Google Scholar] [CrossRef]
- Rzeznik, M.; Triba, M.N.; Levy, P.; Jungo, S.; Botosoa, E.; Duchemann, B.; Le Moyec, L.; Bernaudin, J.F.; Savarin, P.; Guez, D. Identification of a discriminative metabolomic fingerprint of potential clinical relevance in saliva of patients with periodontitis using 1H nuclear magnetic resonance (NMR) spectroscopy. PLoS ONE 2017, 12, e0182767. [Google Scholar] [CrossRef] [PubMed]
- Romano, F.; Meoni, G.; Manavella, V.; Baima, G.; Tenori, L.; Cacciatore, S.; Aimetti, M. Analysis of salivary phenotypes of generalized aggressive and chronic periodontitis through nuclear magnetic resonance-based metabolomics. J. Periodontol. 2018, 89, 1452–1460. [Google Scholar] [CrossRef] [PubMed]
- García-Villaescusa, A.; Morales-Tatay, J.M.; Monleón-Salvadó, D.; González-Darder, J.M.; Bellot-Arcis, C.; Montiel-Company, J.M.; Almerich-Silla, J.M. Using NMR in saliva to identify possible biomarkers of glioblastoma and chronic periodontitis. PLoS ONE 2018, 13, e0188710. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.P.; Saxena, M.; Saimbi, C.S.; Siddiqui, M.H.; Roy, R. Post-periodontal surgery propounds early repair salivary biomarkers by 1 H NMR based metabolomics. Metabolomics 2019, 15, 141. [Google Scholar] [CrossRef] [PubMed]
- Gawron, K.; Wojtowicz, W.; Łazarz-Bartyzel, K.; Łamasz, A.; Qasem, B.; Mydel, P.; Chomyszyn-Gajewska, M.; Potempa, J.; Mlynarz, P. Metabolomic status of the oral cavity in chronic periodontitis. In Vivo 2019, 33, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Citterio, F.; Romano, F.; Meoni, G.; Iaderosa, G.; Grossi, S.; Sobrero, A.; Dego, F.; Corana, M.; Berta, C.N.; Tenori, L.; et al. Changes in the salivary metabolic profile of generalized periodontitis patients after non-surgical periodontal therapy: A metabolomic analysis using nuclear magnetic resonance spectroscopy. J. Clin. Med. 2020, 9, 3977. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H.J.; Song, Y.; Lee, H.A.; Kim, S.; Chung, J. Metabolic phenotyping of saliva to identify possible biomarkers of periodontitis using proton nuclear magnetic resonance. J. Clin. Periodontol. 2021, 48, 1240–1249. [Google Scholar] [CrossRef] [PubMed]
- Fidalgo, T.K.S.; Freitas-Fernandes, L.B.; Angeli, R.; Muniz, A.M.S.; Gonsalves, E.; Santos, R.; Nadal, J.; Almeida, F.C.L.; Valente, A.P.; Souza, I.P.R. Salivary metabolite signatures of children with and without dental caries lesions. Metabolomics 2013, 9, 657–666. [Google Scholar] [CrossRef]
- Pereira, J.L.; Duarte, D.; Carneiro, T.J.; Ferreira, S.; Cunha, B.; Soares, D.; Costa, A.L.; Gil, A.M. Saliva NMR metabolomics: Analytical issues in pediatric oral health research. Oral Dis. 2019, 25, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Jakubovics, N.S. Saliva as the Sole Nutritional Source in the Development of Multispecies Communities in Dental Plaque. Microbiol. Spectr. 2015, 3, 263–277. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.D. Microbial Ecology of Dental Plaque and its Significance in Health and Disease. Adv. Dent. Res. 1994, 8, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Schulz, A.; Lang, R.; Behr, J.; Hertel, S.; Reich, M.; Kümmerer, K.; Hannig, M.; Hannig, C.; Hofmann, T. Targeted metabolomics of pellicle and saliva in children with different caries activity. Sci. Rep. 2020, 10, 697. [Google Scholar] [CrossRef]
- Tomita, Y.; Miyake, N.; Yamanaka, S. Lipids in human parotid saliva with regard to caries experience. J. Oleo. Sci. 2008, 57, 115–121. [Google Scholar] [CrossRef]
- Van Houte, J. Role of micro-organisms in caries etiology. J. Dent. Res. 1994, 73, 672–681. [Google Scholar] [CrossRef]
- Estreicher, A.; Broggiato, A.; Duroux, P.; Andersen, E.; Cimasoni, G. Low molecular-weight proteins in human gingival crevicular fluid. Arch. Oral Biol. 1996, 41, 733–738. [Google Scholar] [CrossRef]
- Niederman, R.; Zhang, J.; Kashket, S. Short-chain carboxylic-acid-stimulated, PMN-mediated gingival inflammation. Crit. Rev. Oral Biol. Med. 1997, 8, 269–290. [Google Scholar] [CrossRef]
- Linden, G.J.; Lyons, A.; Scannapieco, F.A. Periodontal systemic associations: Review of the evidence. J. Periodontol. 2013, 84, S8–S19. [Google Scholar] [CrossRef]
- Han, Y.W.; Wang, X. Mobile microbiome: Oral bacteria in extra-oral infections and inflammation. J. Dent. Res. 2013, 92, 485–491. [Google Scholar] [CrossRef]
- Cui, R.Z.; Bruce, A.J.; Rogers, R.S., 3rd. Recurrent aphthous stomatitis. Clin. Dermatol. 2016, 34, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Akintoye, S.O.; Greenberg, M.S. Recurrent aphthous stomatitis. Dent. Clin. N. Am. 2014, 58, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Pärssinen, M.; Jäsberg, H.; Mikkonen, J.J.W.; Kullaa, A.M. Oral mucosal pellicle as an immune protection against micro-organisms in patients with recurrent aphthous stomatitis: A hypothesis. Med. Hypotheses 2021, 146, 110449. [Google Scholar] [CrossRef] [PubMed]
- Slebioda, Z.; Szponar, E.; Kowalska, A. Etiopathogenesis of recurrent aphthous stomatitis and the role of immunologic aspects: Literature review. Arch. Immunol. Ther. Exp. 2014, 62, 205–215. [Google Scholar] [CrossRef]
- Babaee, N.; Hosseinkazemi, H.; Pouramir, M.; Baboli, O.K.; Salehi, M.; Khadir, F.; Bijani, A.; Mehryari, M. Salivary oxidant/ antioxidant status and hematological parameters in patients with recurrent aphthous stomatitis. Caspian. J. Intern. Med. 2016, 7, 13–18. [Google Scholar]
- Li, Y.; Wang, D.; Zeng, C.; Liu, Y.; Huang, G.; Mei, Z. Salivary metabolomics profile of patients with recurrent aphthous ulcer as revealed by liquid chromatography-tandem mass spectrometry. J. Int. Med. Res. 2018, 46, 1052–1062. [Google Scholar] [CrossRef]
- Kullaa-Mikkonen, A.; Sorvari, T.E. A scanning electron microscopic study of fissured tongue. J. Oral Pathol. 1986, 15, 93–97. [Google Scholar] [CrossRef]
- Wetzel, S.L.; Wollenberg, J. Oral Potentially Malignant Disorders. Dent. Clin. N. Am. 2020, 64, 25–37. [Google Scholar] [CrossRef]
- Wei, J.; Xie, G.; Zhou, Z.; Shi, P.; Qiu, Y.; Zheng, X.; Chen, T.; Su, M.; Zhao, A.; Jia, W. Salivary metabolite signatures of oral cancer and leukoplakia. Int. J. Cancer 2011, 129, 2207–2217. [Google Scholar] [CrossRef]
- Sridharan, G.; Ramani, P.; Patankar, S.; Vijayaraghavan, R. Evaluation of salivary metabolomics in oral leukoplakia and oral squamous cell carcinoma. J. Oral Pathol. Med. 2019, 48, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Sugimoto, M.; Edamatsu, K.; Sugano, A.; Kitabatake, K.; Iino, M. Discrimination of oral squamous cell carcinoma from oral lichen planus by salivary metabolomics. Oral Dis. 2020, 26, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Tantray, S.; Sharma, S.; Prabhat, K.; Nasrullah, N.; Gupta, M. Salivary metabolite signatures of oral cancer and leukoplakia through gas chromatography-mass spectrometry. J. Oral Maxillofac. Pathol. 2022, 26, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef]
- Wang, Q.; Gao, P.; Wang, X.; Duan, Y. The early diagnosis and monitoring of squamous cell carcinoma via saliva metabolomics. Sci. Rep. 2014, 4, 6802. [Google Scholar] [CrossRef]
- Ishikawa, S.; Sugimoto, M.; Kitabatake, K.; Sugano, A.; Nakamura, M.; Kaneko, M.; Ota, S.; Hiwatari, K.; Enomoto, A.; Soga, T.; et al. Identification of salivary metabolomic biomarkers for oral cancer screening. Sci. Rep. 2016, 19, 31520. [Google Scholar] [CrossRef]
- Lohavanichbutr, P.; Zhang, Y.; Wang, P.; Gu, H.; Nagana Gowda, G.A.; Djukovic, D.; Buas, M.F.; Raftery, D.; Chen, C. Salivary metabolite profiling distinguishes patients with oral cavity squamous cell carcinoma from normal controls. PLoS ONE 2018, 13, e0204249. [Google Scholar] [CrossRef]
- Ishikawa, S.; Wong, D.T.W.; Sugimoto, M.; Gleber-Netto, F.O.; Li, F.; Tu, M.; Zhang, Y.; Akin, D.; Iino, M. Identification of salivary metabolites for oral squamous cell carcinoma and oral epithelial dysplasia screening from persistent suspicious oral mucosal lesions. Clin. Oral Investig. 2019, 23, 3557–3563. [Google Scholar] [CrossRef]
- Song, X.; Yang, X.; Narayanan, R.; Shankar, V.; Ethiraj, S.; Wang, X.; Duan, N.; Ni, Y.H.; Hu, Q.; Zare, R.N. Oral squamous cell carcinoma diagnosed from saliva metabolic profiling. Proc. Natl. Acad. Sci. USA 2020, 117, 16167–16173. [Google Scholar] [CrossRef]
- Wang, X.; Liu, L.; Du, Q.; Sun, Z.; Yue, E.; Xue, P.; Zhao, H. Human Saliva Metabolome for Oral Lichen Planus Biomarker Identification. Recent Pat. Anticancer Drug Discov. 2021, 16, 417–425. [Google Scholar] [CrossRef]
- Yatsuoka, W.; Ueno, T.; Miyano, K.; Enomoto, A.; Ota, S.; Sugimoto, M.; Uezono, Y. Time-Course of Salivary Metabolomic Profiles during Radiation Therapy for Head and Neck Cancer. J. Clin. Med. 2021, 10, 2631. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer tatistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Chocolatewala, N.; Chaturvedi, P.; Desale, R. The role of bacteria in oral cancer. Indian J. Med. Paediatr. Oncol. 2010, 31, 126–131. [Google Scholar] [CrossRef] [PubMed]
- Tuominen, H.; Rautava, J. Oral Microbiota and Cancer Development. Pathobiology 2021, 88, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef]
- Ahn, J.; Chen, C.Y.; Hayes, R.B. Oral microbiome and oral and gastrointestinal cancer risk. Cancer Causes Control 2012, 23, 399–404. [Google Scholar] [CrossRef]
- Meurman, J.H. Oral microbiota and cancer. J. Oral Microbiol. 2010, 10, 2. [Google Scholar] [CrossRef]
- Assad, D.X.; Mascarenhas, E.C.P.; de Lima, C.L.; de Toledo, I.P.; Chardin, H.; Combes, A.; Acevedo, A.C.; Guerra, E.N.S. Salivary metabolites to detect patients with cancer: A systematic review. Int. J. Clin. Oncol. 2020, 25, 1016–1036. [Google Scholar] [CrossRef]
- Hannig, C.; Hannig, M.; Kensche, A.; Carpenter, G. The mucosal pellicle—An underestimated factor in oral physiology. Arch. Oral Biol. 2017, 80, 144–152. [Google Scholar] [CrossRef]
- Li, X.; Kolltveit, K.M.; Tronstad, L.; Oslen, I. Systemic diseases caused by oral infection. Clin. Microbiol. Rev. 2000, 13, 547–558. [Google Scholar] [CrossRef]
- Hajishengallis, G. Periodontitis: From microbial immune subversion to systemic inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Asher, S.; Stephen, R.; Mäntylä, P.; Suominen, A.L.; Solomon, A. Periodontal health, cognitive decline, and dementia: A systematic review and meta-analysis of longitudinal studies. J. Am. Geriatr. Soc. 2022, 70, 2695–2709. [Google Scholar] [CrossRef] [PubMed]
- Qiqiang, L.; Huanxin, M.; Xuejun, G. Longitudinal study of volatile fatty acids in the gingival crevicular fluid of patients with periodontitis before and after nonsurgical therapy. J. Periodontal. Res. 2012, 47, 740–749. [Google Scholar] [CrossRef] [PubMed]
- Cueno, M.E.; Ochiai, K. Gingival Periodontal Disease (PD) Level-Butyric Acid Affects the Systemic Blood and Brain Organ: Insights Into the Systemic Inflammation of Periodontal Disease. Front. Immunol. 2018, 4, 1158. [Google Scholar] [CrossRef]
- De La Peña, V.A.; Diz Dios, P.; Tojo Sierra, R. Relationship between lactate dehydrogenase activity in saliva and oral health status. Arch. Oral Biol. 2007, 52, 911–915. [Google Scholar] [CrossRef]
- Piao, Y.; Lu, L.; de Groot, J. AMPA receptors promote perivascular glioma invasion via beta1 integrin-dependent adhesion to the extracellular matrix. Neuro. Oncol. 2009, 11, 260–273. [Google Scholar] [CrossRef]
- Lyssiotis, C.A.; Kimmelman, A.C. Metabolic Interactions in the Tumor Microenvironment. Trends Cell Biol. 2017, 27, 863–875. [Google Scholar] [CrossRef]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef]
- Harber, K.J.; de Goede, K.E.; Verberk, S.G.S.; Meinster, E.; de Vries, H.E.; van Weeghel, M.; de Winther, M.P.J.; Van den Bossche, J. Succinate Is an Inflammation-Induced Immunoregulatory Metabolite in Macrophages. Metabolites 2020, 10, 372. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, F.; Thomas, S.C.; Zhang, Y.; Paul, B.; Sakilam, S.; Chae, S.; Li, P.; Almeter, C.; Kamer, A.R.; et al. Targeting the succinate receptor effectively inhibits periodontitis. Cell Rep. 2022, 40, 111389. [Google Scholar] [CrossRef]
- Sonveaux, P.; Végran, F.; Schroeder, T.; Wergin, M.C.; Verrax, J.; Rabbani, Z.N.; De Saedeleer, C.J.; Kennedy, K.M.; Diepart, C.; Jordan, B.F.; et al. Targeting lactate-fueled respiration selectively kills hypoxic tumor cells in mice. J. Clin. Investig. 2008, 118, 3930–3942. [Google Scholar] [CrossRef] [PubMed]
- Schuller, H.M.; Al-Wadei, H.A.; Majidi, M. Gamma-aminobutyric acid, a potential tumor suppressor for small airway-derived lung adenocarcinoma. Carcinogenesis 2008, 29, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Niggemann, B.; Zaenker, K.S.; Entschladen, F. The neurotransmitter gamma-aminobutyric acid is an inhibitory regulator for the migration of SW 480 colon carcinoma cells. Cancer Res. 2002, 62, 6467–6469. [Google Scholar] [PubMed]
- Tamashiro, P.M.; Furuya, H.; Shimizu, Y.; Iino, K.; Kawamori, T. The impact of sphingosine kinase-1 in head and neck cancer. Biomolecules 2013, 3, 481–513. [Google Scholar] [CrossRef]
- Arima, Y.; Nishigori, C.; Takeuchi, T.; Oka, S.; Morimoto, K.; Utani, A.; Miyachi, Y. 4-Nitroquinoline 1-oxide forms 8-hydroxydeoxyguanosine in human fibroblasts through reactive oxygen species. Toxicol. Sci. 2006, 91, 382–392. [Google Scholar] [CrossRef]
- Roy, D.; Cai, Q.; Felty, Q.; Narayan, S. Estrogen-induced generation of reactive oxygen and nitrogen species, gene damage, and estrogen-dependent cancers. J. Toxicol. Environ. Health B Crit. Rev. 2007, 10, 235–257. [Google Scholar] [CrossRef]
- Parys, J.B.; Decuypere, J.P.; Bultynck, G. Role of the inositol 1,4,5-trisphosphate receptor/Ca2+-release channel in autophagy. Cell Commun. Signal 2012, 10, 17. [Google Scholar] [CrossRef]
- Salaspuro, M. Acetaldehyde and gastric cancer. J. Dig. Dis. 2011, 12, 51–59. [Google Scholar] [CrossRef]
- Miyoshi, E.; Moriwaki, K.; Nakagawa, T. Biological function of fucosylation in cancer biology. J. Biochem. 2008, 143, 725–729. [Google Scholar] [CrossRef]
- Manjula, S.; Monteiro, F.; Rao Aroor, A.; Rao, S.; Annaswamy, R.; Rao, A. Assessment of serum L-fucose in brain tumor cases. Ann. Indian Acad. Neurol. 2010, 13, 33–36. [Google Scholar] [CrossRef]
- Listinsky, J.J.; Siegal, G.P.; Listinsky, C.M. The emerging importance of α-L-fucose in human breast cancer: A review. Am. J. Transl. Res. 2011, 3, 292–322. [Google Scholar] [PubMed]
- Shah, M.; Telang, S.; Raval, G.; Shah, P.; Patel, P.S. Serum fucosylation changes in oral cancer and oral precancerous conditions: Alpha-L-fucosidase as a marker. Cancer 2008, 113, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Shetty, R.K.; Bhandary, S.K.; Kali, A. Significance of Serum L-fucose Glycoprotein as Cancer Biomarker in Head and Neck Malignancies without Distant Metastasis. J. Clin. Diagn. Res. 2013, 7, 2818–2820. [Google Scholar]
- Mikkonen, J.J.W.; Singh, S.P.; Akhi, R.; Salo, T.; Lappalainen, R.; González-Arriagada, W.A.; Ajudarte Lopes, M.; Kullaa, A.M.; Myllymaa, S. Potential role of nuclear magnetic resonance spectroscopy to identify salivary metabolite alterations in patients with head and neck cancer. Oncol. Lett. 2018, 16, 6795–6800. [Google Scholar] [CrossRef] [PubMed]
- Martina, E.; Campanati, A.; Diotallevi, F.; Offidani, A. Saliva and Oral Diseases. J. Clin. Med. 2020, 9, 466. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Oral Diseases N (HC/D) * | Saliva | Elevated Salivary Metabolites | Lowered Salivary Metabolites | Reference |
|---|---|---|---|---|
| Periodontitis 54 (22/32) | WS | acetate, c-aminobutyrate, n-butyrate, succinate, trimethylamine, propionate, valine | pyruvate, N-acetyl groups | [13] |
| Periodontitis 52 (26/26) | SWS | butyrate | fucose, lactate, acetate, N-acetyl of glycoprotein, GABA, 3-hydroxybutyrate, pyruvate, methanol, threonine, ethanol | [14] |
| Aggressive chronic periodontitis 100 (39/61) | USWS | proline, phenylalanine, isoleucine, valine, tyrosine | pyruvate, N-acetyl groups, lactate | [15] |
| Chronic periodontitis 130 (120/10) | USWS | caproate, isocaproate, butyrate, isovalerate, isoleucine, isopropanol, methanol, 4-aminobutyrate, choline, sucrose, glycine, lysine, lactate, proline | [16] | |
| Chronic periodontitis, post-surgery 176 (52/124) | WS | lactate, ethanol, succinate, glutamate | [17] | |
| Chronic periodontitis 45 (15/30) | US | propylene glycol, ethanol, lactate, acetoin, succinate, methanol, glycerol, formate | valine, propionate, isopropanol, alanine, pyruvate, acetate, acetone, choline, taurine, glycine, glucose | [18] |
| Periodontitis after therapy 23 (11/12) | USWS | alanine, glycine, taurine, proline, leucine, valine, isovalerate, tyrosine, methylamine, phenylalanine, isoleucine, lactate, formate, glucose, sarcosine, hypoxanthine, uracil | acetate, propionate, butyrate, ethanol, succinate, acetoin, galactose, aspartate, creatine, choline, methanol, pyruvate, isopropanol | [19] |
| Periodontitis 221 (92/129) | SWS | taurine, glucose, butyrate, isovalerate, glycolate, formate | ethanol, acetate, acetone, acetoin, choline, pyruvate, proline, lysine, propionate | [20] |
| Dental caries 30 (10/20) | USWS | butyrate, ambiguous, lysine, saccharide region, phenylalanine, and propionate. | [21] | |
| Dental caries in children 38 (38/NM) | USWS SWS | alanine, aspartate, glutamine, glycine, isoleucine, leucine, proline, taurine, tyrosine, fucose, galactose, glucose, xylose, choline, dimethylsulfone, hypoxanthine, menthol, N-acetyls, uracil | butyrate, acetone | [22] |
| Oral Diseases N (HC/D) * | Saliva | Elevated Salivary Metabolites | Lowered Salivary Metabolites | Reference |
|---|---|---|---|---|
| OSCC 87/215 | WS | alanine, taurine, pipecolic acid, leucine, isoleucine, histidine, valine, tryptophan, glutamic acid, threonine, carnitine | [44] | |
| OSCC/OL 34/69 | NM | alanine, lactic acid, 3-indolepropionic acid, n-eicosanoic acid | valine, proline, isoleucine, leucine, n-tetradecanoic acid, proline, threonine, phenylalanine, γ-aminobutyric acid | [40] |
| OSCC 30/30 | USWS | lactic acid, hydroxyphenynactic acid, N-nonanoylglycine, 5-hydroxymethyluracil, succinic acid, ornithine, hexanoylcarnitine, propionylcholine | carnitine, 4-hydroxy-L-glutamic acid, acetylphenylalanine, spihingarine, phytosphingosine, S-carboxymethyl-L-cystein | [45] |
| Oral cancer 44/24 | USWS | 3PG, pipecolate, spermidine, met, SAM, 2 AB, trp, val, hypoxanthine, gly-gly, trimetrhylamine N-oxide, guanine, guanosine, tautine, choline, cadaverine, thr | [46] | |
| OSCC 35/101 | NM | glycine, proline, citrulline, ornithine | [47] | |
| OSCC (OED, PSOML) NA/48 | USWS | ornithine, carnitine, arginine, o-hydroxybenzoate, N-acetylglucosamine-1-phosphate, and ribose 5-phosphate (R5P) | [48] | |
| OL and OSCC 18/43 | USWS | d-glycerate-2-phosphate, estrone-3-glucuronide, 4-nitroquinoline-1-oxide, sphin-ganine-1 phosphate,1-methyl histidine, inositol 1,3,4-triphosphate, d-glycerate-2-phosphate, 2-oxoarginine, norcocaine nitroxide, pseudouridine | S-ureidoglycolic acid, p-chlorphenylalanine, d-urobilinogen, N-(3-Indolylacetyl)-l-isoleucine, tetradecanedioic acid, 1-hexadecyl hexadecanoate, l- homocysteic acid, ubiquinone, neuraminic acid, and estradiol valerate. | [41] |
| OSCC 124/249 | USWS | putrescine, cadaverine, thymidine, adenosine, 5-aminopentoate | hippuric acid, phosphocholine, glucose, serine, adrenic acid. | [49] |
| OLP/OSCC NA/60 | USWS | trimethylamine N-oxide, putrescine, creatinine, 5-aminovalerate, pipecolate, N-acetylputrescine, gamma-butyrobetaine, indole-3-acetate, N1-acetylspermine, 2′-deoxyinosine, ethanolamine phosphate, N-acetylglucosamine | N-acetylhistidine, o-acetylcarnitine | [42] |
| OLP 125/120 | USWS | 6 amino acid metabolites, 2 carnitines, 2 lipid metabolites and 9 other metabolites. | [50] | |
| HNC with RT NA/9 | USWS | histidine, tyrosine, urocanate, glycine, glutamic acid, aspartic acid, tryptophan, lysine, methionine, gamma-aminobutyric acid (GABA), butyrate, 2-isopropaylate, 2-aminobutyric acids | [51] | |
| Oral cancer and OL 30/60 | USWS | decanedioic acid, 2-methyloctacosane, Eicosane, Octane, 3,5-dimethyl, pentadecane, hentriacontane, 5,5-diethylpentadecane, nonadecane, oxalic acid, 6-phenylundecanea, L-proline, 2-furancarboxamide, 2-isopropyl-5-methyl-1-heptanol, pentanoic acid, docosane. | [43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyvärinen, E.; Kashyap, B.; Kullaa, A.M. Oral Sources of Salivary Metabolites. Metabolites 2023, 13, 498. https://doi.org/10.3390/metabo13040498
Hyvärinen E, Kashyap B, Kullaa AM. Oral Sources of Salivary Metabolites. Metabolites. 2023; 13(4):498. https://doi.org/10.3390/metabo13040498
Chicago/Turabian StyleHyvärinen, Eelis, Bina Kashyap, and Arja M. Kullaa. 2023. "Oral Sources of Salivary Metabolites" Metabolites 13, no. 4: 498. https://doi.org/10.3390/metabo13040498