
Childhood Obesity and the Cryptic Language of the Microbiota: Metabolomics’ Upgrading

Abstract
:1. Introduction
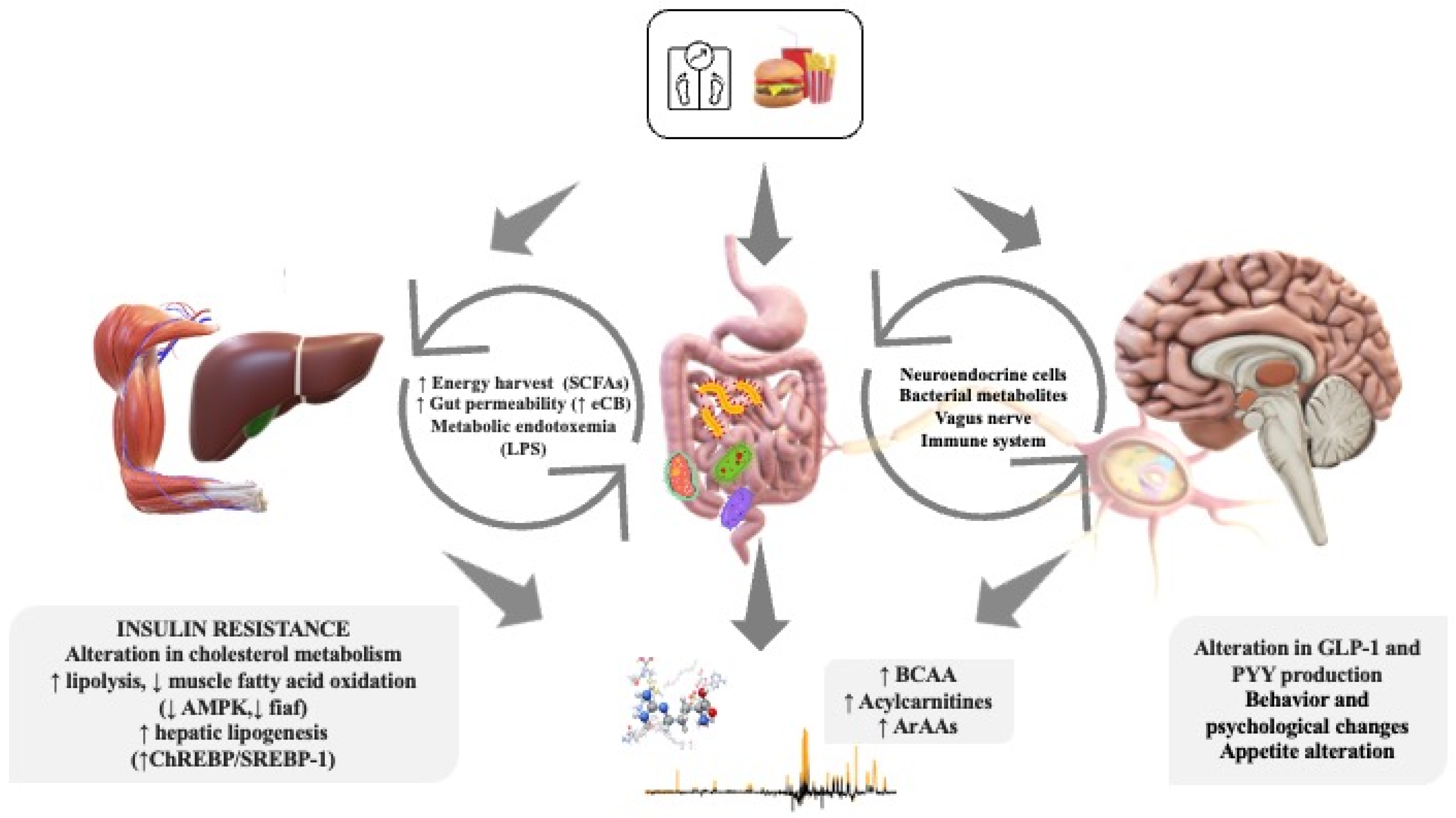
2. Childhood Obesity
3. Obesity and Microbiota in Children
4. Microbiota–Gut–Brain Axis in Children
5. Metabolomics in Childhood Obesity
6. Compared Metabolomics and Microbiomics Analysis of Childhood Obesity

{kind=link}
| Authors/Year | Patients | Samples | Technique | Main Metabolomics Findings | Microbiota Influence | Clinical Significance |
|---|---|---|---|---|---|---|
| Mastrangelo et al. [17] 2016 | 60 prepubertal OB children (30 girls/30 boys, 50% IR and 50% non-IR in each group, with similar BMIs) | Serum | LC–MS, GC–MS, CE–MS | IR group: ↑ LPs (), BCAAs, ArAAs, alanine, proline, pyruvate, ketoisocaproic acid, C3 and C4 acylcarnitines, ↓ free carnitine, bilirubin, nitro-octadecenoate, docosahexaenoate, docosapentanoate, and 3-hydroxybutyrate | IR group: ↑ glycodeoxycholate, Taurodeoxycholate, and piperidine | Metabolic pathways inherent to inflammation, central carbon metabolism along with some metabolites from the gut microbiota were more altered in obese children with IR, with alterations more pronounced for the female sex |
| Troisi et al. [115] 2017 | 36 children/adolescents (aged 5–16 years), 22 OB (including 10 without NAFLD and 12 with NAFLD) | Urine | GC–MS | In obese group: ↑ levels of glucose/1-methylhistidine ↓ levels of xylitol, phenyl acetic acid, and hydroquinone leucine/oxovalerate correlated with excess of visceral fat centimeters valine metabolites correlated with more deranged IP and SIBO | ↑ urinary PCS (an intestinal microbial metabolite) in obese children without NAFLD urinary PCS correlated negatively with the presence of SIBO | A complex network of urinary molecules appears to be correlated with clinical phenotype and distinguishes obese children between those with and without NAFLD. Individual or grouped metabolites interact with anthropometrics and variously aggregated GLA parameters |
| López-Contreras et al. [68] 2018 | 138 unrelated children, 67 HWC and 71 OB (80 boys and 58 girls, aged 6–12 years) | Serum + Stool | FIA–MS + 16sRNA | ↑ serum levels of BCAA (valine and leucine/isoleucine) and ArAAs (phenylalanine and tyrosine) in obese phenylalanine serum levels show a negative and significant correlation with both B. plebeius and unclassified Christensenellaceae abundance | No significant differences in phyla abundances or Firmicutes/Bacteroidetes ratios ↑ Bacteroides eggerthii abundance in obese that correlated positively with body fat percentage and negatively with insoluble fiber intake ↑ Bacteroides plebeius and unclassified Christensenellaceae abundances in normal weight | Identification of bacterial species associated with obesity and related metabolic alterations in order to design dietary intervention studies, which could eventually lead to translational dietary recommendations |
| Quiroga et al. [117] 2020 | 43 children (aged between 7 and 12 years), 29 OB and 14 HWC. OB group was randomly split into two categories (20 training participants followed a 12-week combined strength and endurance training program; the control obese group, 9, maintained their normal daily routines) | Stool | H1 NMR + BaseSpace Application 16 S Metagenomics v1.0 (Illumina Inc.) | exercise intervention modified the metabolic profile in obese patients, representing a dispersing factor: ↓ BCAAs (isoleucine and leucine) and xylose, glucose, and galactose moderate ↓ formate and alanine | In obese: no significant differences in phyla abundances or Firmicutes/ Bacteroidetes ratios ↑ phylum Proteobacteria ↓ genera Clostridium, Bifidobacterium, Coprococcus, Akkermansia, and Streptococcus ↑ Bacteroides, Prevotella, Phascolarctobacterium, and Paraprevotella exercise intervention: ↓ Proteobacteria phylum and Gammaproteobacteria class ↑ genera Blautia, Dialister, and Roseburia | Identification of an obesity-related deleterious microbiota profile that is positively modified by physical activity intervention |
| Jaimes et al. [91] 2021 | 52 children (aged 7 to 16 years), 16 HWC, 17 HW, 19 OB | Stool | H1 NMR + 16S rRNA | ↑fecal butyrate in the OB compared with the N group ↑ arabinose and galactose in OW and OB (strong positive correlation with each other, and both showed a significant positive correlation with the BMI z-score) ↑ TMA in the OW and OB | ↓Escherichia in relative abundance from the N to the OB group (genus includes both commensal and pathogenic species) ↑ Tyzzerella subgroup 3 in a relative abundance from the N to the OB group | Increased energy harvest in OB by the human gut microbiota |
| McCann et al. [118] 2021 | 54 adolescents (aged 10–18 years), 27 with BMI ≥ 95th percentile and 27 HWC. OB group are patients in the Healthy Lifestyles program, which includes visits to a multidisciplinary clinic and membership in a community-based fitness program (6 months of intervention) | Serum + stool | ISQ single quadruple GC–MS + UPLC/MS–MS + 16S rRNA | after FDR adjustment for multiple comparisons, no metabolites were significantly different between the OB and HWC groups nominally significantly different in OB: ↑ BCAA valine ↓ KIC and KMV | Significant differences in measurements of alpha and beta diversity between OB and HWC group 2 Lachnospiraceae families and a Lachnospira species characterized OB samples while members of the Christensenellaceae, Ruminococcae UCG_14 families and Alistipes species defined HWC | Suggestion of a metabolic signature of obesity unique to adolescents and confirmation of a metabolic and microbiome markers of obesity |
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Strauss, R.S.; Pollack, H.A. Epidemic increase in childhood overweight, 1986-1998. JAMA 2001, 286, 2845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afshin, A.; Forouzanfar, M.H.; Reitsma, M.B.; Sur, P.; Estep, K.; Lee, A.; Marczak, L.; Mokdad, A.H.; Moradi-Lakeh, M.; Naghavi, M.; et al. Health Effects of Overweight and Obesity in 195 Countries over 25 Years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [PubMed]
- Han, J.C.; Lawlor, D.A.; Kimm, S.Y. Childhood obesity. Lancet 2010, 375, 1737–1748. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Gang, X.; Liu, Y.; Sun, C.; Han, Q.; Wang, G. Using Metabolomic Profiles as Biomarkers for Insulin Resistance in Childhood Obesity: A Systematic Review. J. Diabetes Res. 2016, 2016, 8160545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concepcion, J.; Chen, K.; Saito, R.; Gangoiti, J.; Mendez, E.; Nikita, M.E.; Barshop, B.A.; Natarajan, L.; Sharma, K.; Kim, J.J. Identification of pathognomonic purine synthesis biomarkers by metabolomic profiling of adolescents with obesity and type 2 diabetes. PLoS ONE 2020, 15, e0234970. [Google Scholar] [CrossRef]
- World Health Organisation. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 23 November 2022).
- Sonntag, D.; Ali, S.; De Bock, F. Lifetime indirect cost of childhood overweight and obesity: A decision analytic model. Obesity 2016, 24, 200–206. [Google Scholar] [CrossRef] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Musso, G.; Gambino, R.; Cassader, M. Obesity, diabetes, and gut microbiota: The hygiene hypothesis expanded? Diabetes Care 2010, 33, 2277–2284. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef]
- Puder, J.J.; Munsch, S. Psychological correlates of childhood obesity. Int. J. Obes. 2010, 34 (Suppl. S2), S37–S43. [Google Scholar] [CrossRef] [Green Version]
- Geier, A.B.; Foster, G.D.; Womble, L.G.; McLaughlin, J.; Borradaile, K.E.; Nachmani, J.; Sherman, S.; Kumanyika, S.; Shults, J. The relationship between relative weight and school attendance among elementary schoolchildren. Obesity 2007, 15, 2157–2161. [Google Scholar] [CrossRef] [PubMed]
- Caird, J.; Kavanagh, J.; Mara-eves, O.; Oliver, K.; Oliver, S.; Stansfield, C.; Thomas, J. Does Being Overweight Impede Academic Attainment? A Systematic Review. Health Educ. J. 2014, 73, 497–521. [Google Scholar] [CrossRef]
- Bomberg, E.M.; Addo, O.Y.; Sarafoglou, K.; Miller, B.S. Adjusting for Pubertal Status Reduces Overweight and Obesity Prevalence in the United States. J. Pediatr. 2021, 231, 200. [Google Scholar] [CrossRef] [PubMed]
- Hübel, C.; Herle, M.; Santos Ferreira, D.L.; Abdulkadir, M.; Bryant-Waugh, R.; Loos, R.J.F.; Bulik, C.M.; Lawlor, D.A.; Micali, N. Childhood overeating is associated with adverse cardiometabolic and inflammatory profiles in adolescence. Sci. Rep. 2021, 11, 12478. [Google Scholar] [CrossRef] [PubMed]
- Weiss, R.; Dziura, J.; Burgert, T.S.; Tamborlane, W.V.; Taksali, S.E.; Yeckel, C.W.; Allen, K.; Lopes, M.; Savoye, M.; Morrison, J.; et al. Obesity and the metabolic syndrome in children and adolescents. N. Engl. J. Med. 2004, 350, 2362–2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastrangelo, A.; Martos-Moreno, G.Á.; García, A.; Barrios, V.; Rupérez, F.J.; Chowen, J.A.; Barbas, C.; Argente, J. Insulin resistance in prepubertal obese children correlates with sex-dependent early onset metabolomic alterations. Int. J. Obes. 2016, 40, 1494–1502. [Google Scholar] [CrossRef] [Green Version]
- Ten, S.; Maclaren, N. Insulin resistance syndrome in children. J. Clin. Endocrinol. Metab. 2004, 89, 2526–2539. [Google Scholar] [CrossRef]
- Butte, N.F.; Liu, Y.; Zakeri, I.F.; Mohney, R.P.; Mehta, N.; Voruganti, V.S.; Göring, H.; Cole, S.A.; Comuzzie, A.G. Global metabolomic profiling targeting childhood obesity in the Hispanic population. Am. J. Clin. Nutr. 2015, 102, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Barlow, S.E.; Dietz, W.H. Obesity evaluation and treatment: Expert Committee recommendations. The Maternal and Child Health Bureau, Health Resources and Services Administration and the Department of Health and Human Services. Pediatrics 1998, 102, E29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardel, M.I.; Atkinson, M.A.; Taveras, E.M.; Holm, J.C.; Kelly, A.S. Obesity Treatment Among Adolescents: A Review of Current Evidence and Future Directions. JAMA Pediatr. 2020, 174, 609–617. [Google Scholar] [CrossRef] [PubMed]
- Berger, N.A. Young adult cancer: Influence of the obesity pandemic. Obesity 2018, 26, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Yu, Z.; Kleber, M.; Singmann, P.; Holzapfel, C.; He, Y.; Mittelstrass, K.; Polonikov, A.; Prehn, C.; Römisch-Margl, W.; et al. Childhood obesity is associated with changes in the serum metabolite profile. Obes. Facts 2012, 5, 660–670. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Pastor-Villaescusa, B.; Gil, A. Are we close to defining a metabolomic signature of human obesity? A systematic review of metabolomics studies. Metabolomics 2019, 15, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Mäkinen, V.P.; Luk, H.; Yang, X. Systems Biology Approaches and Applications in Obesity, Diabetes, and Cardiovascular Diseases. Curr. Cardiovasc. Risk Rep. 2013, 7, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Spiegeleer, M.; De Paepe, E.; Van Meulebroek, L.; Gies, I.; De Schepper, J.; Vanhaecke, L. Paediatric obesity: A systematic review and pathway mapping of metabolic alterations underlying early disease processes. Mol. Med. 2021, 27, 145. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, B.; Jain, V. Obesity in Children: Definition, Etiology and Approach. Indian J. Pediatr. 2018, 85, 463–471. [Google Scholar] [CrossRef]
- Blüher, S.; Schwarz, P. Metabolically healthy obesity from childhood to adulthood—Does weight status alone matter? Metabolism 2014, 63, 1084–1092. [Google Scholar] [CrossRef]
- Valerio, G.; Balsamo, A.; Baroni, M.G.; Brufani, C.; Forziato, C.; Grugni, G.; Licenziati, M.R.; Maffeis, C.; Miraglia Del Giudice, E.; Morandi, A.; et al. on the behalf of the Childhood Obesity Group of the Italian Society of Pediatric Endocrinology and Diabetology. Childhood obesity classification systems and cardiometabolic risk factors: A comparison of the Italian, World Health Organization and International Obesity Task Force references. Ital. J. Pediatr. 2017, 43, 19. [Google Scholar] [PubMed] [Green Version]
- Wan Mohd Zin, R.M.; Jalaludin, M.Y.; Yahya, A.; Nur Zati Iwani, A.K.; Md Zain, F.; Hong, J.Y.H.; Mokhtar, A.H.; Wan Mohamud, W.N. Prevalence and clinical characteristics of metabolically healthy obese versus metabolically unhealthy obese school children. Front. Endocrinol. 2022, 13, 971202. [Google Scholar] [CrossRef]
- Bervoets, L.; Massa, G. Classification and clinical characterization of metabolically “healthy” obese children and adolescents. J. Pediatr. Endocrinol. Metab. 2016, 29, 553–560. [Google Scholar] [CrossRef]
- Prince, R.L.; Kuk, J.L.; Ambler, K.A.; Dhaliwal, J.; Ball, G.D. Predictors of metabolically healthy obesity in children. Diabetes Care 2014, 37, 1462–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bervoets, L.; Massa, G.; Guedens, W.; Reekmans, G.; Noben, J.P.; Adriaensens, P. Identification of metabolic phenotypes in childhood obesity by 1H NMR metabolomics of blood plasma. Future Sci. OA 2018, 4, FSO310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dessì, A.; Cesare Marincola, F.; Masili, A.; Gazzolo, D.; Fanos, V. Clinical metabolomics and nutrition: The new frontier in neonatology and pediatrics. Biomed. Res. Int. 2014, 2014, 981219. [Google Scholar] [CrossRef]
- Dessì, A.; Fanos, V. Pediatric obesity: Could metabolomics be a useful tool? J. Pediatr. Neonat. Individual. Med. 2013, 2, e020205. [Google Scholar]
- Cho, K.; Moon, J.S.; Kang, J.H.; Jang, H.B.; Lee, H.J.; Park, S.I.; Yu, K.S.; Cho, J.Y. Combined untargeted and targeted metabolomic profiling reveals urinary biomarkers for discriminating obese from normal-weight adolescents. Pediatr. Obes. 2017, 12, 93–101. [Google Scholar] [CrossRef]
- Rupérez, F.J.; Martos-Moreno, G.Á.; Chamoso-Sánchez, D.; Barbas, C.; Argente, J. Insulin Resistance in Obese Children: What Can Metabolomics and Adipokine Modelling Contribute? Nutrients 2020, 12, 3310. [Google Scholar] [CrossRef]
- Oliveira, M.H.; Pereira, D.; Melo, D.S.; Silva, J.C.; Conde, W.L. Accuracy of international growth charts to assess nutritional status in children and adolescents: A systematic review. Rev. Paul. Pediatr. 2022, 40, e2021016. [Google Scholar] [CrossRef]
- Harrington, D.M.; Staiano, A.E.; Broyles, S.T.; Gupta, A.K.; Katzmarzyk, P.T. BMI percentiles for the identification of abdominal obesity and metabolic risk in children and adolescents: Evidence in support of the CDC 95th percentile. Eur. J. Clin. Nutr. 2013, 67, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.J.; Braun, W.; Enderle, J.; Bosy-Westphal, A. Beyond BMI: Conceptual Issues Related to Overweight and Obese Patients. Obes. Facts 2016, 9, 193–205. [Google Scholar] [CrossRef]
- Gätjens, I.; Schmidt, S.; Plachta-Danielzik, S.; Bosy-Westphal, A.; Müller, M.J. Body Composition Characteristics of a Load-Capacity Model: Age-Dependent and Sex-Specific Percentiles in 5- to 17-Year-Old Children. Obes. Facts 2021, 14, 593–603. [Google Scholar] [CrossRef] [PubMed]
- Cole, T.J.; Green, P.J. Smoothing reference centile curves: The LMS method and penalized likelihood. Stat. Med. 1992, 11, 1305–1319. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, K.; Kassis, A.; Major, G.; Chou, C.J. Is the gut microbiota a new factor contributing to obesity and its metabolic disorders? J. Obes. 2012, 2012, 14. [Google Scholar]
- Blaser, M.J.; Devkota, S.; McCoy, K.D.; Relman, D.A.; Yassour, M.; Young, V.B. Lessons learned from the pre- natal microbiome controversy. Microbiome 2021, 9, 1–7. [Google Scholar] [CrossRef]
- Walter, J.; Hornef, M.W. A philosophical perspective on the prenatal in utero microbiome debate. Microbiome 2021, 9, 1–9. [Google Scholar] [CrossRef]
- Yao, Y.; Cai, X.; Ye, Y.; Wang, F.; Chen, F.; Zheng, C. The Role of Microbiota in Infant Health: From Early Life to Adulthood. Front. Immunol. 2021, 12, 708472. [Google Scholar] [CrossRef] [PubMed]
- Vangay, P.; Ward, T.; Gerber, J.S.; Knights, D. Antibiotics, pediatric dysbiosis, and disease. Cell Host Microbe. 2015, 17, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.M.; Blaser, M.J. Pathways in microbe-induced obesity. Cell Metab. 2013, 17, 883–894. [Google Scholar] [CrossRef] [Green Version]
- Cox, L.M.; Yamanishi, S.; Sohn, J.; Alekseyenko, A.V.; Leung, J.M.; Cho, I.; Kim, S.G.; Li, H.; Gao, Z.; Mahana, D.; et al. Altering the intestinal microbiota during a critical developmental window has lasting metabolic conse- quences. Cell 2014, 158, 705–721. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [Green Version]
- Beller, L.; Deboutte, W.; Falony, G.; Vieira-Silva, S.; Tito, R.Y.; Valles-Colomer, M.; Rymenans, L.; Jansen, D.; Van Espen, L.; Papadaki, M.I.; et al. Successional Stages in Infant Gut Microbiota Maturation. mBio 2021, 12, e0185721. [Google Scholar] [CrossRef]
- Gupta, A.; Osadchiy, V.; Mayer, E.A. Brain-gut-microbiome interactions in obesity and food addiction. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 655–672. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Liu, L.; Zhu, Y.; Huang, G.; Wang, P.P. The association between breastfeeding and childhood obesity: A meta-analysis. BMC Public Health 2014, 14, 1267. [Google Scholar] [CrossRef] [Green Version]
- Walker, W.A. The importance of appropriate initial bacterial colonization of the intestine in newborn, child, and adult health. Pediatr. Res. 2017, 82, 387e95. [Google Scholar] [CrossRef] [Green Version]
- Human Microbiome Project Consortium. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Knoll, R.L.; Forslund, K.; Kultima, J.R.; Meyer, C.U.; Kullmer, U.; Sunagawa, S.; Bork, P.; Gehring, S. Gut microbiota differs between children with inflammatory bowel disease and healthy siblings in taxonomic and functional composition: A metagenomic analysis. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G327e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.; Cope, J.L.; Nagy-Szakal, D.; Dowd, S.; Versalovic, J.; Hollister, E.B.; Kellermayer, R. Composition and function of the pediatric colonic mucosal microbiome in untreated patients with ulcer- ative colitis. Gut. Microb. 2016, 7, 384e96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun, H.M.; Bridgman, S.L.; Chari, R.; Field, C.J.; Guttman, D.S.; Becker, A.B.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P.; Sears, M.R.; et al. Roles of birth mode and infant gut microbiota in intergenerational transmission of overweight and obesity from mother to offspring. JAMA Pediatr. 2018, 172, 368e77. [Google Scholar] [CrossRef]
- Orbe-Orihuela, Y.C.; Lagunas-Martínez, A.; Bahena-Román, M.; Madrid-Marina, V.; Torres-Poveda, K.; Flores-Alfaro, E.; Méndez-Padrón, A.; Díaz-Benítez, C.E.; Peralta-Zaragoza, O.; Antúnez-Ortiz, D.; et al. High relative abundance of firmicutes and increased TNF-α levels correlate with obesity in children. Salud. Publica. Mex. 2018, 60, 5–11. [Google Scholar] [CrossRef] [Green Version]
- West, C.E.; Rydén, P.; Lundin, D.; Engstrand, L.; Tulic, M.K.; Prescott, S.L. Gut microbiome and innate immune response patterns in IgE- associated eczema. Clin. Exp. Allergy 2015, 45, 1419e29. [Google Scholar] [CrossRef] [Green Version]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Björkstén, B.; Engstrand, L.; Jenmalm, M.C. Low diversity of the gut microbiota in infants with atopic eczema. J. Allergy Clin. Immunol. 2012, 129, 434–440.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Losada, M.; Castro-Nallar, E.; Bendall, M.L.; Freishtat, R.J.; Crandall, K.A. Dual Transcriptomic Profiling of Host and Microbiota during Health and Disease in Pediatric Asthma. PLoS ONE 2015, 10, e0131819. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.J.; Gerasimidis, K.; Edwards, C.A.; Shaikh, M.G. Role of Gut Microbiota in the Aetiology of Obesity: Proposed Mechanisms and Review of the Literature. J. Obes. 2016, 2016, 7353642. [Google Scholar] [CrossRef] [Green Version]
- Indiani, C.M.D.S.P.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.C.; Darrieux, M.; Parisotto, T.M. Childhood Obesity and Firmicutes/Bacteroidetes Ratio in the Gut Microbiota: A Systematic Review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef]
- López-Contreras, B.E.; Morán-Ramos, S.; Villarruel-Vázquez, R.; Macías-Kauffer, L.; Villamil-Ramírez, H.; León-Mimila, P.; Vega-Badillo, J.; Sánchez-Muñoz, F.; Llanos-Moreno, L.E.; Canizalez-Román, A.; et al. Composition of gut microbiota in obese and normal-weight Mexican school-age children and its association with metabolic traits. Pediatr. Obes. 2018, 13, 381–388. [Google Scholar] [CrossRef]
- Gallardo-Becerra, L.; Cornejo-Granados, F.; García-López, R.; Valdez-Lara, A.; Bikel, S.; Canizales-Quinteros, S.; López-Contreras, B.E.; Mendoza-Vargas, A.; Nielsen, H.; Ochoa-Leyva, A. Metatranscriptomic analysis to define the Secrebiome, and 16S rRNA profiling of the gut microbiome in obesity and metabolic syndrome of Mexican children. Microb. Cell Fact. 2020, 19, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trasande, L.; Blustein, J.; Liu, M.; Corwin, E.; Cox, L.M.; Blaser, M.J. Infant antibiotic exposures and early-life body mass. Int. J. Obes. 2013, 37, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Ajslev, T.A.; Andersen, C.S.; Gamborg, M.; Sørensen, T.I.; Jess, T. Child-hood overweight after establishment of the gut microbiota: The role of delivery mode, pre-pregnancy weight and early adminis- tration of antibiotics. Int. J. Obes. 2011, 35, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Yallapragada, S.G.; Nash, C.B.; Robinson, D.T. Early-life exposure to antibiotics, alterations in the intestinal microbiome, and risk of metabolic disease in children and adults. Pediatr. Ann. 2015, 44, e265–e269. [Google Scholar] [CrossRef]
- Chambers, E.S.; Morrison, D.J.; Frost, G. Control of appetite and energy intake by SCFA: What are the potential underlying mechanisms? Proc. Nutr. Soc. 2015, 74, 328–336. [Google Scholar] [CrossRef] [Green Version]
- Houtman, T.A.; Eckermann, H.A.; Smidt, H.; de Weerth, C. Gut microbiota and BMI throughout childhood: The role of firmicutes, bacteroidetes, and short-chain fatty acid producers. Sci. Rep. 2022, 12, 3140. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, K.V.; Sherwin, E.; Schellekens, H.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Feeding the microbiota-gut-brain axis: Diet, microbiome, and neuropsychiatry. Transl. Res. 2017, 179, 223–244. [Google Scholar] [CrossRef]
- Margolis, K.G.; Cryan, J.F.; Mayer, E.A. The Microbiota-Gut-Brain Axis: From Motility to Mood. Gastroenterology 2021, 160, 1486–1501. [Google Scholar] [CrossRef]
- Generoso, J.S.; Giridharan, V.V.; Lee, J.; Macedo, D.; Barichello, T. The role of the microbiota-gut-brain axis in neuropsychiatric disorders. Braz. J. Psychiatry 2021, 43, 293–305. [Google Scholar] [CrossRef]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Gao, K.; Mu, C.L.; Farzi, A.; Zhu, W.Y. Tryptophan Metabolism: A Link Between the Gut Microbiota and Brain. Adv. Nutr. 2020, 11, 709–723. [Google Scholar] [CrossRef]
- Marin, I.A.; Goertz, J.E.; Ren, T.; Rich, S.S.; Onengut-Gumuscu, S.; Farber, E.; Wu, M.; Overall, C.C.; Kipnis, J.; Gaultier, A. Microbiota alteration is associated with the development of stress-induced despair behavior. Sci. Rep. 2017, 7, 43859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Sadanala, K.C.; Kim, E.K. A Metabolomic Approach to Understanding the Metabolic Link between Obesity and Diabetes. Mol. Cells 2015, 38, 587–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hivert, M.F.; Perng, W.; Watkins, S.M.; Newgard, C.S.; Kenny, L.C.; Kristal, B.S.; Patti, M.E.; Isganaitis, E.; DeMeo, D.L.; Oken, E.; et al. Metabolomics in the developmental origins of obesity and its cardiometabolic consequences. J. Dev. Orig. Health Dis. 2015, 6, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Handakas, E.; Lau, C.H.; Alfano, R.; Chatzi, V.L.; Plusquin, M.; Vineis, P.; Robinson, O. A systematic review of metabolomic studies of childhood obesity: State of the evidence for metabolic determinants and consequences. Obes. Rev. 2022, 23 (Suppl. S1), e13384. [Google Scholar] [CrossRef]
- Fattuoni, C.; Palmas, F.; Noto, A.; Fanos, V.; Barberini, L. Perinatal asphyxia: A review from a metabolomics perspective. Molecules 2015, 20, 7000–7016. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Fanos, V. How could metabolomics change pediatric health? Ital. J. Pediatr. 2020, 46, 37. [Google Scholar] [CrossRef] [Green Version]
- Ellul, S.; Wake, M.; Clifford, S.A.; Lange, K.; Würtz, P.; Juonala, M.; Dwyer, T.; Carlin, J.B.; Burgner, D.P.; Saffery, R. Metabolomics: Population epidemiology and concordance in Australian children aged 11-12 years and their parents. BMJ Open 2019, 9 (Suppl. S3), 106–117. [Google Scholar] [CrossRef]
- Newgard, C.B.; An, J.; Bain, J.R.; Muehlbauer, M.J.; Stevens, R.D.; Lien, L.F.; Haqq, A.M.; Shah, S.H.; Arlotto, M.; Slentz, C.A.; et al. A branched-chain amino acid-related metabolic signature that differentiates obese and lean humans and contributes to insulin resistance. Cell Metab. 2009, 9, 311–326, Erratum in: Cell Metab. 2009, 9, 565–566. [Google Scholar] [CrossRef] [Green Version]
- Klein, M.S.; Shearer, J. Metabolomics and Type 2 Diabetes: Translating Basic Research into Clinical Application. J. Diabetes Res. 2016, 2016, 3898502. [Google Scholar] [CrossRef] [Green Version]
- Gawlik, A.; Salonen, A.; Jian, C.; Yanover, C.; Antosz, A.; Shmoish, M.; Wasniewska, M.; Bereket, A.; Wudy, S.A.; Hartmann, M.F.; et al. Personalized approach to childhood obesity: Lessons from gut microbiota and omics studies. Narrative review and insights from the 29th European childhood obesity congress. Pediatr. Obes. 2021, 16, e12835. [Google Scholar] [CrossRef] [PubMed]
- Alonso, A.; Marsal, S.; Julià, A. Analytical methods in untargeted metabolomics: State of the art in 2015. Front. Bioeng. Biotechnol. 2015, 3, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaimes, J.D.; Slavíčková, A.; Hurych, J.; Cinek, O.; Nichols, B.; Vodolánová, L.; Černý, K.; Havlík, J. Stool metabolome-microbiota evaluation among children and adolescents with obesity, overweight, and normal-weight using 1H NMR and 16S rRNA gene profiling. PLoS ONE 2021, 16, e0247378. [Google Scholar] [CrossRef] [PubMed]
- Sohn, M.J.; Chae, W.; Ko, J.S.; Cho, J.Y.; Kim, J.E.; Choi, J.Y.; Jang, H.B.; Lee, H.J.; Park, S.I.; Park, K.H.; et al. Metabolomic Signatures for the Effects of Weight Loss Interventions on Severe Obesity in Children and Adolescents. Metabolites 2021, 12, 27. [Google Scholar] [CrossRef]
- Kim, J.A.; Wei, Y.; Sowers, J.R. Role of mitochondrial dysfunction in insulin resistance. Circ. Res. 2008, 102, 401–414. [Google Scholar] [CrossRef]
- Martos-Moreno, G.Á.; Mastrangelo, A.; Barrios, V.; García, A.; Chowen, J.A.; Rupérez, F.J.; Barbas, C.; Argente, J. Metabolomics allows the discrimination of the pathophysiological relevance of hyperinsulinism in obese prepubertal children. Int. J. Obes. 2017, 41, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Tricò, D.; Prinsen, H.; Giannini, C.; de Graaf, R.; Juchem, C.; Li, F.; Caprio, S.; Santoro, N.; Herzog, R.I. Elevated α-Hydroxybutyrate and Branched-Chain Amino Acid Levels Predict Deterioration of Glycemic Control in Adolescents. J. Clin. Endocrinol. Metab. 2017, 102, 2473–2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormack, S.E.; Shaham, O.; McCarthy, M.A.; Deik, A.A.; Wang, T.J.; Gerszten, R.E.; Clish, C.B.; Mootha, V.K.; Grinspoon, S.K.; Fleischman, A. Circulating branched-chain amino acid concentrations are associated with obesity and future insulin resistance in children and adolescents. Pediatr. Obes. 2013, 8, 52–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Cairano, E.S.; Davalli, A.M.; Perego, L.; Sala, S.; Sacchi, V.F.; La Rosa, S.; Finzi, G.; Placidi, C.; Capella, C.; Conti, P.; et al. The glial glutamate transporter 1 (GLT1) is expressed by pancreatic beta-cells and prevents glutamate-induced beta-cell death. J. Biol. Chem. 2011, 286, 14007–14018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozefsky, T.; Felig, P.; Tobin, J.D.; Soeldner, J.S.; Cahill, G.F., Jr. Amino acid balance across tissues of the forearm in postabsorptive man. Effects of insulin at two dose levels. J. Clin. Investig. 1969, 48, 2273–2282. [Google Scholar] [CrossRef] [Green Version]
- Newbern, D.; Gumus Balikcioglu, P.; Balikcioglu, M.; Bain, J.; Muehlbauer, M.; Stevens, R.; Ilkayeva, O.; Dolinsky, D.; Armstrong, S.; Irizarry, K.; et al. Sex differences in biomarkers associated with insulin resistance in obese adolescents: Metabolomic profiling and principal components analysis. J. Clin. Endocrinol. Metab. 2014, 99, 4730–4739, Erratum in: J. Clin. Endocrinol. Metab. 2015, 100, 1709. [Google Scholar] [CrossRef] [Green Version]
- Ritter, O.; Jelenik, T.; Roden, M. Lipid-mediated muscle insulin resistance: Different fat, different pathways? J. Mol. Med. 2015, 93, 831–843. [Google Scholar] [CrossRef]
- Hellmuth, C.; Kirchberg, F.F.; Lass, N.; Harder, U.; Peissner, W.; Koletzko, B.; Reinehr, T. Tyrosine Is Associated with Insulin Resistance in Longitudinal Metabolomic Profiling of Obese Children. J. Diabetes Res. 2016, 2016, 2108909. [Google Scholar] [CrossRef] [Green Version]
- Perng, W.; Gillman, M.W.; Fleisch, A.F.; Michalek, R.D.; Watkins, S.M.; Isganaitis, E.; Patti, M.E.; Oken, E. Metabolomic profiles and childhood obesity. Obesity 2014, 22, 2570–2578. [Google Scholar] [CrossRef] [Green Version]
- Isganaitis, E.; Rifas-Shiman, S.L.; Oken, E.; Dreyfuss, J.M.; Gall, W.; Gillman, M.W.; Patti, M.E. Associations of cord blood metabolites with early childhood obesity risk. Int. J. Obes. 2015, 39, 1041–1048. [Google Scholar] [CrossRef] [Green Version]
- Hellmuth, C.; Uhl, O.; Standl, M.; Demmelmair, H.; Heinrich, J.; Koletzko, B.; Thiering, E. Cord Blood Metabolome Is Highly Associated with Birth Weight, but Less Predictive for Later Weight Development. Obes. Facts 2017, 10, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Sorrow, P.; Maguire, R.; Murphy, S.K.; Belcher, S.M.; Hoyo, C. Elevated metabolites of acetaminophen in cord blood of children with obesity. Pediatr. Obes. 2019, 14, e12465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rzehak, P.; Hellmuth, C.; Uhl, O.; Kirchberg, F.F.; Peissner, W.; Harder, U.; Grote, V.; Weber, M.; Xhonneux, A.; Langhendries, J.P.; et al. European Childhood Obesity Trial Study Group. Rapid growth and childhood obesity are strongly associated with lysoPC(14:0). Ann. Nutr. Metab. 2014, 64, 294–303. [Google Scholar] [CrossRef] [Green Version]
- Fleddermann, M.; Demmelmair, H.; Hellmuth, C.; Grote, V.; Trisic, B.; Nikolic, T.; Koletzko, B. Association of infant formula composition and anthropometry at 4 years: Follow-up of a randomized controlled trial (BeMIM study). PLoS ONE 2018, 13, e0199859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal-Witt, M.J.; Ramon-Krauel, M.; Samino, S.; Llobet, M.; Cuadras, D.; Jimenez-Chillaron, J.C.; Yanes, O.; Lerin, C. Untargeted metabolomics identifies a plasma sphingolipid-related signature associated with lifestyle intervention in prepubertal children with obesity. Int. J. Obes. 2018, 42, 72–78. [Google Scholar] [CrossRef]
- Leal-Witt, M.J.; Llobet, M.; Samino, S.; Castellano, P.; Cuadras, D.; Jimenez-Chillaron, J.C.; Yanes, O.; Ramon-Krauel, M.; Lerin, C. Lifestyle Intervention Decreases Urine Trimethylamine N-Oxide Levels in Prepubertal Children with Obesity. Obesity 2018, 26, 1603–1610. [Google Scholar] [CrossRef] [Green Version]
- Pathmasiri, W.; Pratt, K.J.; Collier, D.N.; Lutes, L.D.; McRitchie, S.; Sumner, S.C.J. Integrating metabolomic signatures and psychosocial parameters in responsivity to an immersion treatment model for ado- lescent obesity. Metabolomics 2012, 8, 1037–1051. [Google Scholar] [CrossRef]
- Short, K.R.; Chadwick, J.Q.; Teague, A.M.; Tullier, M.A.; Wolbert, L.; Coleman, C.; Copeland, K.C. Effect of Obesity and Exercise Training on Plasma Amino Acids and Amino Metabolites in American Indian Adolescents. J. Clin. Endocrinol. Metab. 2019, 104, 3249–3261. [Google Scholar] [CrossRef]
- Wahl, S.; Holzapfel, C.; Yu, Z.; Breier, M.; Kondofersky, I.; Fuchs, C.; Singmann, P.; Prehn, C.; Adamsky, J.; Grallert, H.; et al. Metabolomics reveals determinants of weight loss during lifestyle intervention in obese children. Metabolomics 2013, 9, 1157–1167. [Google Scholar] [CrossRef] [Green Version]
- Reinehr, T.; Wolters, B.; Knop, C.; Lass, N.; Hellmuth, C.; Harder, U.; Peissner, W.; Wahl, S.; Grallert, H.; Adamski, J.; et al. Changes in the serum metabolite profile in obese children with weight loss. Eur. J. Nutr. 2015, 54, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, T.; Kulle, A.; Rothermel, J.; Knop-Schmenn, C.; Lass, N.; Bosse, C.; Holterhus, P.M. Longitudinal analyses of the steroid metabolome in obese PCOS girls with weight loss. Endocr. Connect. 2017, 6, 213–224. [Google Scholar] [CrossRef] [Green Version]
- Troisi, J.; Pierri, L.; Landolfi, A.; Marciano, F.; Bisogno, A.; Belmonte, F.; Palladino, C.; Nuzio, S.G.; Campiglia, P.; Vajro, P. Urinary Metabolomics in Pediatric Obesity and NAFLD Identifies Metabolic Pathways/Metabolites Related to Dietary Habits and Gut-Liver Axis Perturbations. Nutrients 2017, 9, 485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, K.P.; Luo, F.J.; Plummer, N.S.; Hostetter, T.H.; Meyer, T.W. The production of p-cresol sulfate and indoxyl sulfate in vegetarians versus omnivores. Clin. J. Am. Soc. Nephrol. 2012, 7, 982–988. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, R.; Nistal, E.; Estébanez, B.; Porras, D.; Juárez-Fernández, M.; Martínez-Flórez, S.; García-Mediavilla, M.V.; de Paz, J.A.; González-Gallego, J.; Sánchez-Campos, S.; et al. Exercise training modulates the gut microbiota profile and impairs inflammatory signaling pathways in obese children. Exp. Mol. Med. 2020, 52, 1048–1061. [Google Scholar] [CrossRef]
- McCann, J.R.; Bihlmeyer, N.A.; Roche, K.; Catherine, C.; Jawahar, J.; Kwee, L.C.; Younge, N.E.; Silverman, J.; Ilkayeva, O.; Sarria, C.; et al. The Pediatric Obesity Microbiome and Metabolism Study (POMMS): Methods, Baseline Data, and Early Insights. Obesity 2021, 29, 569–578. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bosco, A.; Loi, M.; Pinna, G.; Pintus, R.; Fanos, V.; Dessì, A. Childhood Obesity and the Cryptic Language of the Microbiota: Metabolomics’ Upgrading. Metabolites 2023, 13, 414. https://doi.org/10.3390/metabo13030414
Bosco A, Loi M, Pinna G, Pintus R, Fanos V, Dessì A. Childhood Obesity and the Cryptic Language of the Microbiota: Metabolomics’ Upgrading. Metabolites. 2023; 13(3):414. https://doi.org/10.3390/metabo13030414
Chicago/Turabian StyleBosco, Alice, Michele Loi, Giulia Pinna, Roberta Pintus, Vassilios Fanos, and Angelica Dessì. 2023. "Childhood Obesity and the Cryptic Language of the Microbiota: Metabolomics’ Upgrading" Metabolites 13, no. 3: 414. https://doi.org/10.3390/metabo13030414